Introduction
Cutaneous melanoma, developing in melanocytes that
produce melanin, is a most pernicious tumors in skin malignancy and
have a poor prognosis for survival (1). Emerging evidences indicate that
melanoma gives rise to the majority of skin tumor-associated death
(2–4), incontrollable growth and metastasis
early are the main characteristics in the progression of cutaneous
melanoma (5). The etiology of
cutaneous melanoma is multifactorial and complicated including
environmental and genetic factors (6,7).
Therefore, it is extremely crucial that further investigation of
gene expression and regulation in the development of melanoma.
miRNAs are small and noncoding RNA molecules which play key roles
in post-transcriptional regulation of gene expression by binding to
the 3′-UTR of targeted mRNA (8–10).
Previous studies manifested microRNAs were involved in a range of
biological activities including cell proliferation, transformation,
and tumorigenesis (11). Emerging
evidences identified that abnormal miRNAs expression was associated
with the initiation and procession of tumors (12–14).
PTEN, known as phosphatase and tensin homolog
deleted on chromosome 10, is a phosphoinositide phosphatase that
plays a tumor suppressor role in malignant melanoma (15). Increasing evidences revealed that
microRNAs were involved in the tumorigenesis of tumors by
regulating the expression of PTEN. For example, miR-500a promoted
the invasion and migration of hepatocarcinoma via suppressing the
expression of PTEN (16). miR-20b
enhanced prostate cancer growth through targeting PTEN (17). So far, the regulatory relationship
remains obscure and needs to be elucidated carefully between
microRNAs and PTEN in malignant cutaneous melanoma.
miR-367 had been reported to be involved in the
tumorigenesis and transformation of different cancers (18,19),
and it played tumor promotor or tumor suppressor roles in diverse
tumors. For instance, overexpression of miR-367 was associated with
a unfavorable prognosis and enhanced non-small cell lung cancer
growth by binding to FBXW7 (20).
In contrast, miR-367 was found to be related with TNM stage of
patients and suppress gastric cancer development via targeting
Rab23 (21). Up to date, the
effect of miR-367 on cutaneous melanoma remains unclear. The
present report aims to detect the expression of miR-367 in
cutaneous melanoma tissues and cell lines, and investigate the
regulatory mechanism of miR-367 on the development of cutaneous
melanoma.
Materials and methods
Patients and tissues
Fifty melanoma tissues and 25 benign nevi tissues
were collected from the Affiliated Hospital of Hubei University of
Chinese Medicine and these samples were immediately frozen in
liquid nitrogen until use. Written informed consents were obtained
from all of recipients enrolled. These patients did not receive any
radiation or chemotherapy before operation. All tissue sections
were reviewed at least by two pathologists. The use of these
specimens was approved by the Ethics Committee of the Affiliated
Hospital of Hubei University of Chinese Medicine.
Cell culture
Human melanoma cell lines A375, WM35, SK-MEL-5 and
SK-MEL-2 were incubated in Dulbecco's modified Eagle's medium
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 100
U/ml streptomycin at 37°C in 5% CO2 atmosphere. Human
epidermal melanocytes (HEMa-LP) were obtained from Cascade
Biologics, Inc. (Mansfield, UK) and were cultured in medium with
HMGS-2 in a humidified atmosphere of 5% CO2 at 37°C.
Cell transfection
A375 cells were transiently transfected with
Lipofectamine 2000 according to the manufacturer's instructions
(Invitrogen; Thermo Fisher Scientific, Inc.). Cells were
transfected with miR-367 mimics, miR-367 inhibitor, control vectors
or pcDNA3.1 vector encoding PTEN (GeneCopoeia, Inc., Rockville, MY,
USA). The transfected cells were incubated at 37°C and 5%
CO2. After 4 to 6 h, the transfection solution was
replaced with fresh culture medium for subsequent experiments.
RNA extraction and reverse
transcription-quantitative polymerease chain reaction
The total RNA of the tissue samples and cells was
extracted using the TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
cDNA was converted and amplified using the TaqMan miRNA reverse
transcription kit according to the manufacturer's assay procedure.
Quantitative real-time PCR was performed with TaqMan human MiRNA
assay kit. miR-367 expression was calculated relative to the
expression of U6. The expression changes were determined using
2−∆∆Ct method. miR-367 forward,
5′-TTCTCCGAACTTGTCACGTTT-3′ and reverse,
5′-ACGTGACACGTTCGGAGAATT-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′
and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
Cell proliferation assay
Melanoma cells proliferation was measured by using a
cell proliferation kit, Cell Counting Kit-8, according to the
manufacturer's instructions. Cells after transfection were seeded
into 96-well culture plates and incubated at 37°C. Fresh culture
media (90 µl) and 10 µl CCK-8 solutions were added to each sample
at 12, 24, 48 and 72 h. Subsequently, the transfected cells were
cultured at 37°C for 2 h. Cell growth was calculated by a
microplate reader at a wave length of 450 nanometer.
Cell migration assay
Wound healing assay was used to study cell
migration. Melanoma cells were seeded and incubated in 6-well
plates until monolayers were developed. A plastic scriber was used
to make an artificial wound, and the image of the wound was served
as baseline. Cells were then washed twice with PBS and incubated in
DMEM with 10% fetal bovine serum at 37°C for 48 h. Cell migration
capacity was assessed by subtracting the final wound width from the
initial wound width.
Transwell invasion assay
Transwell invasion assay was performed to evaluate
cell invasion ability. A Transwell chamber was precoated with
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA). Cells
(2×105) were seeded into the upper chamber and incubated
in serum free medium. The incubating media with 20% fetal bovine
serum were added into the lower chamber to attract cells. After
being cultured for 48 h, cells on the upper chamber were cleared
with a swab. Cells after migration were fixed and stained with 0.5%
crystal violet, photographed and counted under a microscope.
Luciferase reporter analysis
The 3′-UTR region of PTEN or the mutated sequence
was cloned into pMIR-REPORT vectors. miR-367 mimics, or
corresponding control was transfected together with reporter
plasmid and pRL-SV40 (Promega, Madison, WI, USA) into A375 cells.
The active luciferase level was assessed 48 h after transfection by
a Luciferase Reporter Assay (Promega).
Western blot analysis
The concentration of protein was determined by the
BCA kit (Pierce, Rockford, IL USA). SDS-PAGE assay was performed to
separate the extracted protein mixtures. The proteins were
transferred to a PVDF membrane and incubated with the corresponding
antibodies. Antibodies listed below were used to determine the
protein expression: PTEN (1:500; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA), AKT (1:500), and p-AKT (1:500) (both from Abcam,
Cambridge, MA, USA). Anti-GAPDH (1:2,000; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). Horseradish peroxidase-conjugated
secondary antibody (1:2,000; Abcam). Band intensity was evaluated
using an ECL chemiluminescent kit (Millipore, Billerica, MA,
USA).
Tumor growth in vivo
Nude mice were prepared to determine the melanoma
growth (25–30 g, 6-week-old), A375 cells after transfection were
harvested by centrifugation, washed with PBS and re-suspended, A375
cells were subcutaneously injected into nude mice (n=6), mice were
sacrificed on 5, 10, 15, 20 and 25 days, and tumor tissues were
excised for further study. The weight and volume of the tumor mass
were measured and recorded.
Statistical analysis
SPSS 16.0 (SPSS, Inc., Chicago, IL, USA) was
performed for statistical analyses, including t-test, one-way
analysis of variance with post hoc contrasts by
Student-Newman-Keuls test, Fisher exact probability test, Pearson
correlation analysis and Kaplan-Meier plot. All data were
represented as mean ± standard deviation. P<0.05 was considered
to indicate a statistically significant difference.
Results
Overexpression of miR-367 was
confirmed in cutaneous melanoma and was correlated with poor
prognosis in patients with cutaneous melanoma
qRT-PCR was used to determine the expression of
miR-367 in 50 of cutaneous melanoma tissues and 25 of benign nevi
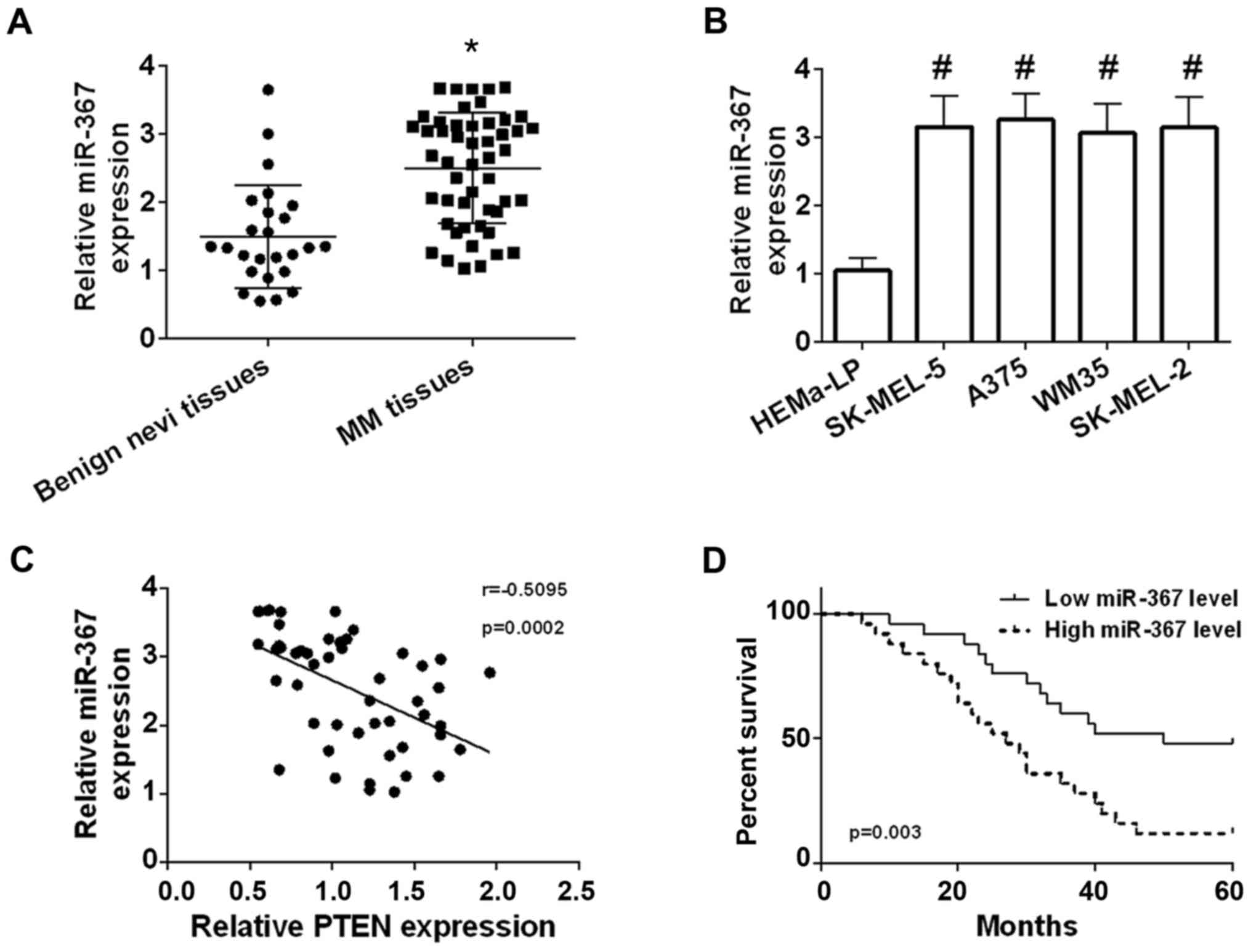
tissue. Our results identified that the miR-367 level was
significantly elevated in melanoma tissues in comparison with
benign nevi tissues (Fig. 1A).
High expression of miR-367 was confirmed in A375 cells, WM35 cells,
SK-MEL-2 cells and SK-MEL-5 cells compared with HEMa-LP cells, and
the highest level of miR-367 was found in A375 cells line (Fig. 1B). The miR-367 level was negatively
correlated with the PTEN level in melanoma tissues (Fig. 1C). Patients with melanoma were
separated into high miR-367 expression group (n=25) and low miR-367
expression (n=25) according to the relative median value of miR-367
(median, 2.665; Min-Max, 1.030–3.680). The correlation was analyzed
between the miR-367 expression and clinical characteristics of
patients with melanoma. The correlation was not statistically
significant between the miR-367 level and age, sex of patients with
melanoma, but high expression of miR-367 was significantly
correlated with tumor thickness, TNM stage, lymph node involvement
and distant metastasis of malignant melanoma as shown in Table I. Besides, high miR-367 level was
positively correlated with decreased overall survival (Fig. 1D). The results manifested that
miR-367 played a promoting role in the development of melanoma.
 | Table I.Association between clinical findings
and the miR-367 level in individuals with cutaneous melanoma. |
Table I.
Association between clinical findings
and the miR-367 level in individuals with cutaneous melanoma.
|
|
| miR-367 |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
findings | Patients (n=50) | High | Low | P-value |
|---|
| Age (years) |
|
|
|
|
| ≤60 | 30 | 17 | 13 |
|
|
>60 | 20 | 8 | 12 | 0.387 |
| Sex |
|
|
|
|
| Male | 28 | 15 | 13 |
|
|
Female | 22 | 10 | 12 | 0.776 |
| Tumor thickness
(mm) |
|
|
|
|
| ≤1 | 11 | 2 | 9 |
|
|
>1 | 39 | 23 | 16 | 0.037 |
| TNM
classification |
|
|
|
|
| I+II | 13 | 2 | 11 |
|
|
III+IV | 37 | 23 | 14 | 0.008 |
| Lyph node
involvement |
|
|
|
|
| No | 17 | 4 | 13 |
|
| Yes | 33 | 21 | 12 | 0.016 |
| Distant
metastasis |
|
|
|
|
| No | 39 | 16 | 23 |
|
| Yes | 11 | 9 | 2 | 0.037 |
High level of miR-367 enhanced the
growth and invasion of A375 cells
A375 cells were selected for further experiments due
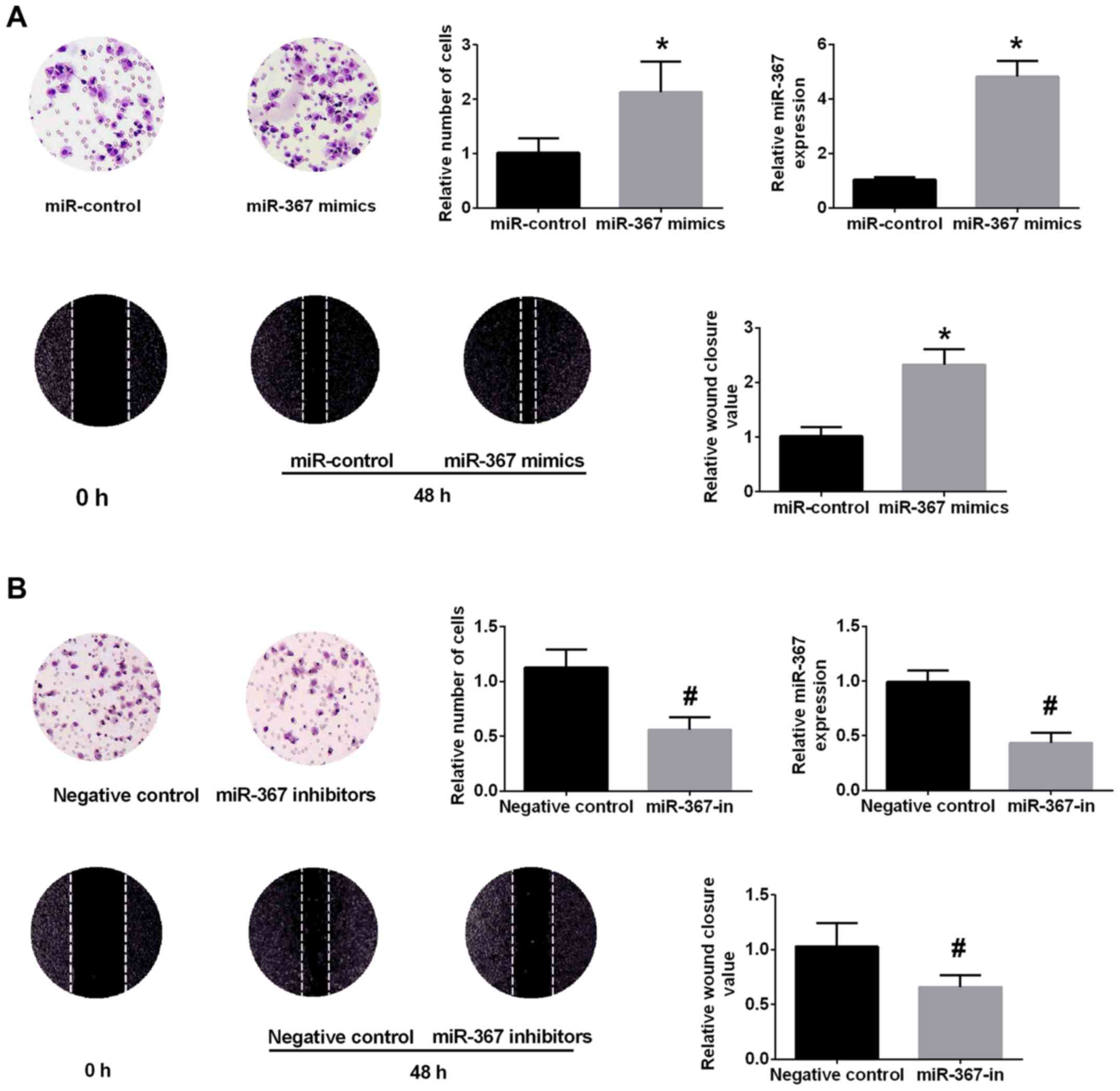
to the highest level of miR-367 in A375 cells. The miR-367 level
was significantly upregulated in miR-367 mimics group in comparison
with miR-control group (Fig. 2A).
CCK-8 assay, Transwell assays and wound healing assay confirmed
that upregulation of miR-367 distinctly promoted the proliferation,
invasion and migration of A375 cells (Fig. 2A). Conversely, the miR-367 level
was decreased in A375 cells after transfection with miR-367
inhibitor (Fig. 2B). The
transfection of miR-367 inhibitor attenuated the proliferation,
invasion and migration of A375 cells (Fig. 2B). These data identified that
miR-367 play a critical role in the growth, invasion and migration
of A375 cells.
PTEN mRNA was identified as the
functional downstream target of miR-367
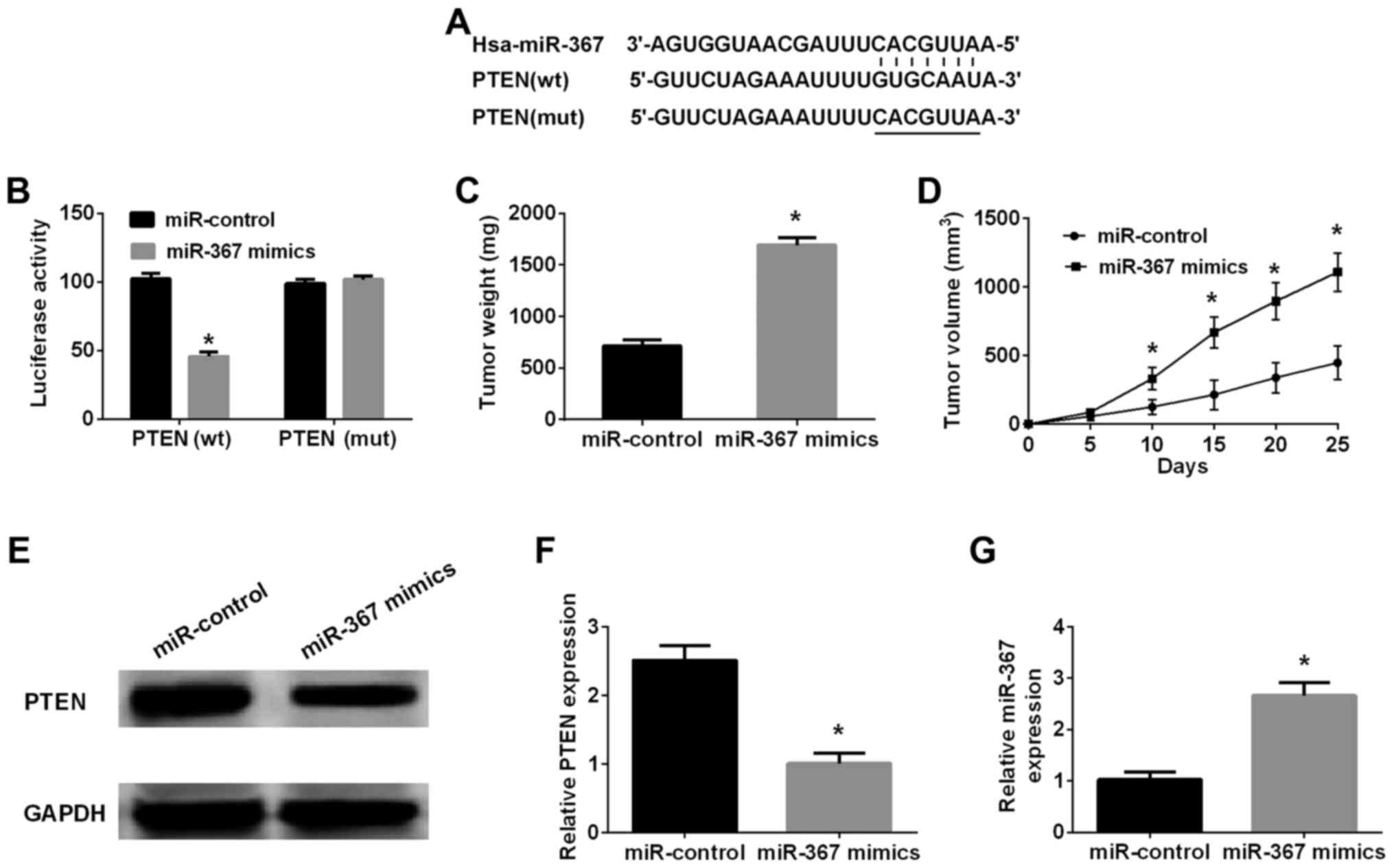
Bioinformatics analysis (TargetScan 7.0 and miRanda)
was performed to investigate the downstream targets of miR-367 in
melanoma cells. Retrieval results manifested that 3′-UTR of PTEN
contained a direct binding sequence of miR-367 (Fig. 3), miR-367 interaction with binding
sequence of 3′-UTR-PTEN was confirmed by luciferase reporter assay.
The results indicated that miR-367 distinctly inhibited luciferase
activity of PTEN in wild-type, however, had no effect on that of
PTEN in mutated type (Fig. 3B).
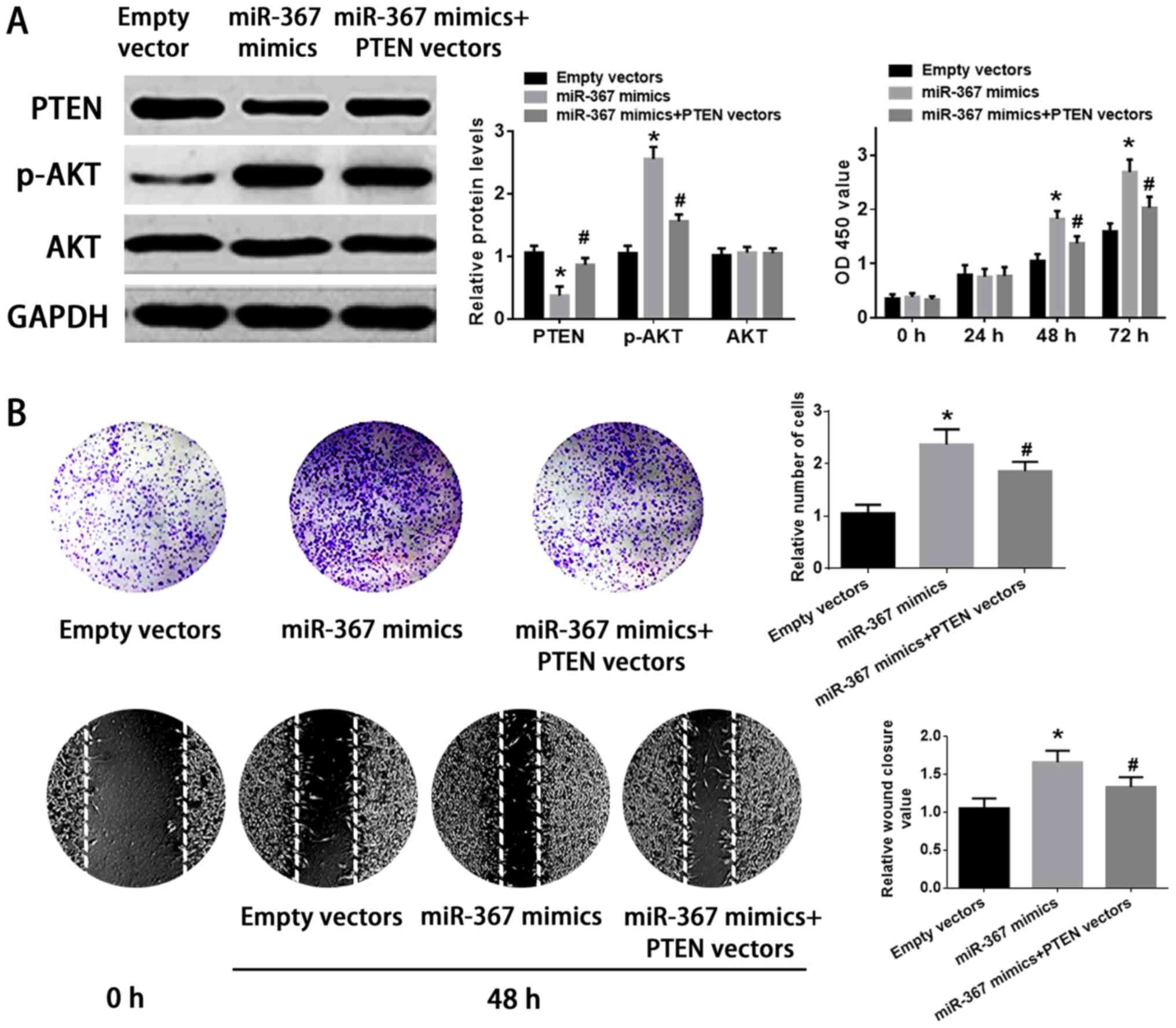
Western blot analysis was implemented to identify whether miR-367
could regulate PTEN expression in A375 cells or not. The results
further identified that high level of miR-367 remarkably suppressed
PTEN expression in A375 cells. Furthermore, the phosphorylation of
AKT was increased by the transfection of miR-367 mimics into A375
cells (Fig. 4A). On the other
hand, the PTEN expression was significantly elevated and the
phosphorylation of AKT was downregulated in A375 cells after
transfection with miR-367 inhibitors (Fig. 4B). Collectively, the results
identified that PTEN was an important downstream targeting mRNA of
miR-367 in A375 cells.
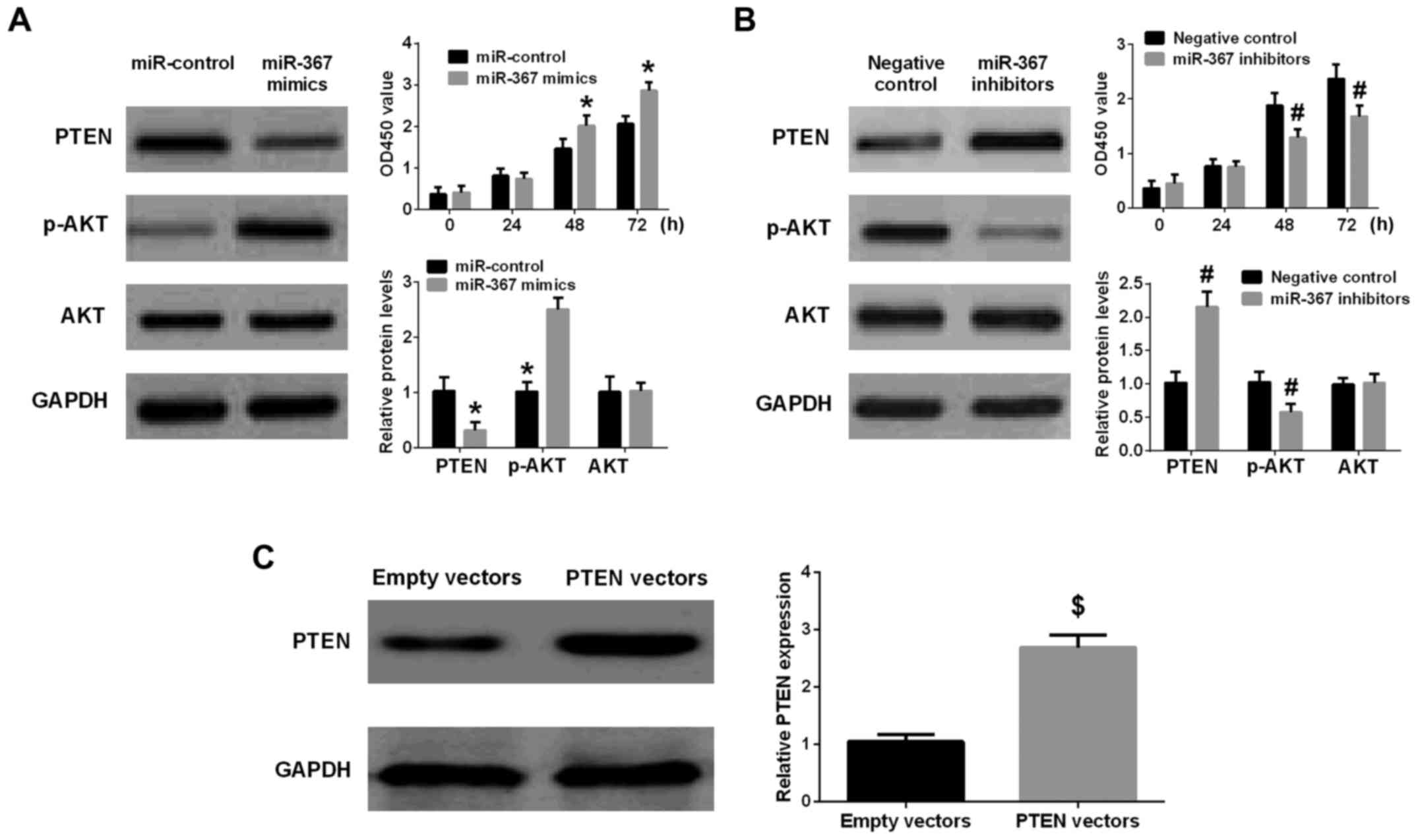 | Figure 4.miR-367 promotes PTEN expression and
inhibits the phosphorylation of AKT in A375 cells, and enhanced
cell proliferation. (A) Upregulation of miR-367 distinctly reduced
PTEN level, promoted the phosphorylation of AKT, and increased cell
proliferation. (B) Downregulation of miR-367 markedly raised the
PTEN level, attenuated the phosphorylation of AKT and inhibited
cell proliferation. (C) The PTEN level was significantly raised in
A375 cells transfected with PTEN vectors in comparison with empty
vectors group. *P<0.01, compared with miR-control group;
#P<0.01, compared with negative control group;
$P<0.01, compared with empty vectors group. miR,
microRNA; PTEN, phosphatase and tensin homolog; AKT, protein kinase
B. |
Upregulation of PTEN attenuated the
effect of miR-367 on the proliferation and invasion of A375
cells
The transfection of PTEN vectors significantly
increased the expression of PTEN in PTEN vectors group compared
with empty vectors group (Fig.
4C). The transfection of miR-367 mimics remarkably suppressed
the expression of PTEN and enhanced the phosphorylation of AKT in
A375 cells, but the cotransfection of miR-367 mimics with PTEN
vectors attenuated the effect of miR-367 mimics on the expression
of PTEN and the phosphorylation of AKT in A375 cells. In addition,
upregulation of miR-367 enhanced the growth, invasion and migration
of A375 cells, however, PTEN vectors ameliorated the ability of
growth, invasion, and migration of A375 cells transfected with
miR-367 mimics. Finally, PTEN vectors and miR-367 mimics had no
significant effect on AKT level in A375 cells (Fig. 5). Briefly, miR-367 was able to
promote the proliferation, invasion and migration of A375 cells via
targeting PTEN.
The effect of miR-367 on melanoma
growth in a xenograft tumor model
A xenograft tumor model was applied to confirm the
promoting function of miR-367 in the progression of melanoma. The
xenograft tumors were collected for determination of weight and
volume. The result showed that the volume of tumor in miR-367
mimics group was significantly increased in comparison with
miR-control group at 10, 15, 20 and 25 days after A375 cells
injection (Fig. 3D), and the
weight of tumor in miR-367 mimics group was markedly larger than
that in miR-control group at 25 days after A375 cells injection
(Fig. 3C). Therefore, it was
identified strongly that forced expression of miR-367 markedly
augmented the growth of xenograft tumors. In addition, the PTEN
expression in tissues was significantly decreased, but the miR-367
level in tissues was distinctly increased in miR-367 mimics group
compared with miR-control group (Fig.
3E-G). Accordingly, enhancing function of miR-367 in the
malignant melanoma growth was confirmed in vivo.
Discussion
miRNA has been identified as a critical
transcriptional mediator in many biological activities including
the development of tumorgenesis by binding to 3′-UTR of its
targeting mRNA. The aberrant expression of miR-367 was found
definitely in many types of tumors (22,23).
The association between miR-367 and cutaneous malignant melanoma
has not been clearly elucidated up to date. Accordingly, we
investigated the expression and underlying regulatory function of
miR-367 in malignant melanoma. As a result, we found that miR-367
was distinctly overexpressed in cutaneous melanoma tissues and cell
lines, and negatively correlated with the PTEN level in cutaneous
melanoma tissues. High expression of miR-367 was markedly related
with the malignant clinical characteristics of individuals with
melanoma. Next, overexpression of miR-367 enhanced A375 cells
proliferation and Transwell migration, and inhibited PTEN
expression, upregulation of PTEN alleviated partially the enhancing
effect of miR-367 on the growth, invasion and migration of A375
cells. Finally, nude mice models study identified in vivo
that overexpression of miR-367 promoted xenograft tumor
development. All of these results combined manifested that miR-367
promoted melanoma progression by, at least in part, regulating the
PTEN expression.
PTEN is a crucial tumor inhibitor gene that is
involved in the occurence and progression of many types of
malignant tumor (24) including
malignant melanoma (25). The
protein derived from the gene can reduce
phosphatidylinositol-3,4,5-trisphosphate level and exert a tumor
inhibitor role through PTEN/AKT pathway (26). In this study, PTEN was identified
to be an important functional target of miR-367 in melanoma cells.
Firstly, the binding target of miR-367 was discovered in 3′UTR-PTEN
mRNA. Secondly, miR-367 decreased distinctly the luciferase
activity in the wild PTEN, however, did not affect luciferase
activity in the mutated PTEN. Thirdly, forced expression of miR-367
inhibited expression of PTEN and promoted the phosphorylation of
AKT, whereas downregulation of miR-367 distinctly promoted the
expression of PTEN and alleviated the phosphorylation of AKT.
Upregulation of PTEN suppressed the pro-tumor function of miR-367
in A375 cells. Finally, The PTEN expression was distinctly reduced
but the miR-367 level was markedly raised in xenograft tumors in
miR-367 mimics group in comparison with miR-control group. All
results combined identified that miR-367 played a pro-tumor role in
A375 cells via regulating PTEN expression. PTEN was a potential
mediator of the PTEN/PI3K/AKT pathway (27). These results further confirmed
miR-367 could regulate the phosphorylation of AKT level by
targeting PTEN mRNA, and be involved in malignant behaviors of A375
cells via regulating PTEN/AKT pathway.
Together, this study confirms the miR-367 level is
distinctly upregulated in cutaneous melanoma. Overexpression of
miR-367 is correlated with malignant clinical characteristics of
individuals with cutaneous melanoma. miR-367 augments the
proliferation and invasion of melanoma. Furthermore, miR-367 plays
promoting roles on the development of melanoma via modulating PTEN
expression. Regulation of miR-367 expression may be a novel and
crucial strategy for cutaneous melanoma treatment.
References
|
1
|
Isola AL, Eddy K and Chen S: Biology,
therapy and implications of tumor exosomes in the progression of
melanoma. Cancers. 8:E1102016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Giblin AV and Thomas JM: Incidence,
mortality and survival in cutaneous melanoma. J Plast Reconstr
Aesthet Surg. 60:32–40. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peterson M, Albertini MR and Remington P:
Incidence, survival and mortality of malignant Cutaneous melanoma
in Wisconsin, 1995–2011. WMJ. 114:196–201. 2015.PubMed/NCBI
|
|
4
|
Downing A, Newton-Bishop JA and Forman D:
Recent trends in cutaneous malignant melanoma in the Yorkshire
region of England; incidence, mortality and survival in relation to
stage of disease, 1993–2003. Br J Cancer. 95:91–95. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leiter U, Meier F, Schittek B and Garbe C:
The natural course of cutaneous melanoma. J Surg Oncol. 86:172–178.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luo C, Weber CE, Osen W, Bosserhoff AK and
Eichmuller SB: The role of microRNAs in melanoma. Eur J Cell Biol.
93:11–22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kashani-Sabet M: Molecular markers in
melanoma. Br J Dermatol. 170:31–35. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mannucci C, Casciaro M, Minciullo PL,
Calapai G, Navarra M and Gangemi S: Involvement of microRNAs in
skin disorders: A literature review. Allergy Asthma Proc. 38:pp.
9–15. 2017; View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Greenberg E, Nemlich Y and Markel G:
MicroRNAs in cancer: Lessons from melanoma. Curr Pharm Des.
20:5246–5259. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carthew RW: Gene regulation by microRNAs.
Curr Opin Genet Dev. 16:203–208. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakahara K and Carthew RW: Expanding roles
for miRNAs and siRNAs in cell regulation. Curr Opin Cell Biol.
16:127–133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Long J, Menggen Q, Wuren Q, Shi Q and Pi
X: miR-219-5p inhibits the growth and metastasis of malignant
melanoma by targeting BCL-2. Biomed Res Int. 2017:90325022017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He J, Zhao J, Zhu W, Qi D, Wang L, Sun J,
Wang B, Ma X, Dai Q and Yu X: MicroRNA biogenesis pathway genes
polymorphisms and cancer risk: A systematic review and
meta-analysis. Peer J. 4:e27062016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fattore L, Costantini S, Malpicci D,
Ruggiero CF, Ascierto PA, Croce CM, Mancini R and Ciliberto G:
MicroRNAs in melanoma development and resistance to target therapy.
Oncotarget. 8:22262–22278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stahl JM, Cheung M, Sharma A, Trivedi NR,
Shanmugam S and Robertson GP: Loss of PTEN promotes tumor
development in malignant melanoma. Cancer Res. 63:2881–2890.
2003.PubMed/NCBI
|
|
16
|
Zhao Y and Wang Y and Wang Y: Up-regulated
microRNA-500a enhances hepatocarcinoma metastasis by repressing
PTEN expression. Biosci Rep. 37:BSR201708372017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo J, Xiao Z, Yu X and Cao R: miR-20b
promotes cellular proliferation and migration by directly
regulating phosphatase and tensin homolog in prostate cancer. Oncol
Lett. 14:6895–6900. 2017.PubMed/NCBI
|
|
18
|
Guan Y, Chen L, Bao Y, Qiu B, Pang C, Cui
R and Wang Y: High miR-196a and low miR-367 cooperatively correlate
with unfavorable prognosis of high-grade glioma. Int J Clin Exp
Pathol. 8:6576–6588. 2015.PubMed/NCBI
|
|
19
|
Sun J, Song K, Feng X and Gao S:
MicroRNA-367 is a potential diagnostic biomarker for patients with
esophageal squamous cell carcinoma. Biochem Biophys Res Commun.
473:363–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu J, Wu W, Wang J, Huang C, Wen W, Zhao
F, Xu X, Pan X, Wang W, Zhu Q and Chen L: miR-367 promotes the
proliferation and invasion of non-small cell lung cancer via
targeting FBXW7. Oncol Rep. 37:1052–1058. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bin Z, Dedong H, Xiangjie F, Hongwei X and
Qinghui Y: The microRNA-367 inhibits the invasion and metastasis of
gastric cancer by directly repressing Rab23. Genet Test Mol
Biomarkers. 19:69–74. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Campayo M, Navarro A, Vinolas N, Diaz T,
Tejero R, Gimferrer JM, Molins L and Cabanas ML: Low miR-145 and
high miR-367 are associated with unfavourable prognosis in resected
nonsmall cell lung cancer. Eur Respir J. 41:1172–1178. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kaid C, Silva PB, Cortez BA, Rodini CO,
Semedo-Kuriki P and Okamoto OK: miR-367 promotes proliferation and
stem-like traits in medulloblastoma cells. Cancer Sci.
106:1188–1195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Milella M, Falcone I, Conciatori F, Cesta
Incani U, Del Curatolo A, Inzerilli N, Nuzzo CM, Vaccaro V, Vari S,
Cognetti F and Ciuffreda L: PTEN: Multiple functions in human
malignant tumors. Front Oncol. 5:242015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang H, Kircher DA, Kim KH, Grossmann AH,
VanBrocklin MW, Holmen SL and Robinson JP: Activated MEK cooperates
with Cdkn2a and Pten loss to promote the development and
maintenance of melanoma. Oncogene. 36:3842–3851. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song MS, Salmena L and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor. Nat Rev Mol
Cell Biol. 13:283–296. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|