Introduction
Osteoporosis is a set of bone diseases caused by
variety of factors featuring normal calcification, normal
proportions of calcium salt and substrate, decreased bone quantity
per unit volume, and metabolic bone lesions (1). Reports have shown that osteoporosis
affects a higher proportion of individuals in the elderly
population (2). Statistics
indicate that osteoporosis is becoming a serious health problem in
the aging population due to weakened bone structure and fractures
in elderly individuals (3,4). The results of pathological
investigations indicate that osteoclast activity and subtypes are
crucial for the function of physiology and pathological
implications for future treatment of osteoporosis (5). Osteoporosis readily leads to hip
fractures, which are associated with functional impairment,
maintenance of bone damage, loss of independence and increased
mortality rates (6,7). Furthermore, clinical practice
guidelines recommend that all patients with osteoporosis receive
chronic oral glucocorticoid treatment. However, the therapeutic
efficacy is limited in patients with osteoporosis who receive
osteoporosis pharmacotherapy in terms of bone mineral density
testing and bone strength. Therefore, examining more efficient
molecular agents for the clinical treatment of osteoporosis is
urgently required.
Tanshinone IIA (Tan-IIA) is a traditional Chinese
medicine extracted from Danshen, which has been used clinically for
the treatment of various human diseases (8,9). A
previous study indicated that Tan-IIA decreased the protein
expression of epidermal growth factor receptor and insulin-like
growth factor 1 receptor, inhibiting the phosphoinositide
3-kinase/Akt/mammalian target of rapamycin pathway in gastric
carcinoma in vitro and in vivo (10). In addition, Tan-IIA was shown to
exert inhibitory effects on the hypoxia-induced enhancement of
store-operated calcium entry in pulmonary arterial smooth muscle
cells through the protein kinase G-proliferator-activated
receptor-γ signaling pathway (11). The pharmacological and therapeutic
properties of Tan-IIA in the cardiovascular system have attracted
interest (12). A previous study
showed that Tan-IIA treatment alleviated rat gingival connective
tissue overgrowth induced by cyclosporine A (13). Emerging experimental investigations
and clinical trials have demonstrated that Tan-IIA prevents cardiac
injury, hypertrophy and atherogenesis through estrogen
receptor-α-induced oxidative stress (14). Although the efficacy of Tan-IIA has
been observed in different diseases (15,16),
the efficacy and molecular mechanism underlying the effect of
Tan-IIA have not been reported previously. In the present study,
the effects and molecular mechanisms underlying the effects of
Tan-IIA on osteocytes were evaluated in vitro and in
vivo. The present study also examined the efficacy of Tan-IIA
on anti-apoptotic effects in the treatment of osteoporosis. Current
evidence suggests that Tan-IIA is an efficient anti-apoptotic agent
in osteoporosis.
Oxidative and reductive stress are essential for the
dynamic phases experienced by cells undergoing adaptation towards
an endogenous or exogenous noxious stimulus (17). Mitochondrial malfunction is the
common denominator arising from the aberrant functioning of the
rheostat, which maintains homeostasis between oxidative and
reductive stress in osteoblasts (18,19).
Previous evidence suggests that the mechanism underlying
osteoporosis is important for osteoclast activation in the
treatment of osteoporosis. Maladaptation during oxidative stress
may be pivotal in the pathophysiology of osteoporosis (20,21).
A previous study provided evidence that targeting antioxidant drug
therapy is an efficient clinical regimen for reducing the
deterioration of osteoporosis in the aforementioned oxidative
stress signaling pathway (22,23).
Evidence has also revealed that endoplasmic reticulum stress is a
key risk factor in the pathogenic progression of osteoporosis.
Therefore, the present study investigated Tan-IIA-mediated
oxidative stress in osteoblasts from mice with osteoporosis. The
data showed that Tan-IIA may be an efficient anti-osteoporotic
agent in vitro and in vivo.
The present study also investigated whether Tan-IIA
has an anti-apoptotic and protective role in the treatment of
osteoporosis in a mouse model. The data suggested that Tan-IIA
ameliorated osteoblast activity and functioned through the nuclear
factor (NF)-κB signaling pathway. Taken together, the data
suggested that Tan-IIA exerted protective effects against oxidative
stress in osteoblastic differentiation of mice with osteoporosis
via regulation of the NF-κB signaling pathway and endoplasmic
reticulum stress-dependent pathways, which may contribute to the
treatment of osteoporosis.
Materials and methods
Ethics statement
The present study was performed in strict accordance
with the recommendations in the Guide for the Care and Use of
Laboratory Animals of Taihe Hospital Affiliated to Hubei University
of Medicine (Hubei, China). All experimental protocols on animals
were performed in accordance with the National Institutes of Health
(24) and approved by the
Committee on the Ethics of Animal Experiments Defence Research of
Tianjin Medical University General Hospital (Tianjin, China). All
surgery and euthanasia were made to minimize suffering.
Animal experiments
A total of 64 female Wnt1sw/sw mice
(10-week-old, 32–40 g) with osteoporosis were purchased from the
Jackson Laboratory (Ben Harbor, ME, USA) on a mixed C57BL/6
background. All animals were fed under pathogen-free conditions.
All animals were housed under controlled temperatures in a 12 h
light/dark cycle with free access to food and water. The
Wnt1sw/sw mice with osteoporosis were randomly divided
into three groups (n=32 in each experimental group) and received
treatment with Tan-IIA (10 mg/kg), Alendronate (ADN) or PBS.
Treatments were performed via an intraperitoneal injection manner
(200 µl) once a day. The detailed procedures were according to
those of a previous report (25).
The therapeutic efficacies of Tan-IIA were analyzed according to a
previous study (26).
Reverse
transcription-quantitative-polymerase chain reaction (RT-qPCR)
analysis
Osteoblasts were isolated from experimental mice
following treatment with Tan-IIA, ADN or PBS as described
previously (27). Total RNA from
the osteoblasts was extracted using an RNAeasy Mini kit (Qiagen
Sciences, Inc., Gaithersburg, MD, USA) according to the
manufacturer's protocols. The expression levels of B-cell lymphoma
(Bcl)-2, alkaline phosphatase (ALP), p53, NF-κB, inducible nitric
oxide synthase (iNOS), cyclooxygenase (COX)2, tumor necrosis factor
(TNF) receptor-associated factor 1 (TRAF-1), TNF-α, Caspase-3,
Apaf-1, and inhibitor of apoptosis protein (IAP) in the osteoblasts
were determined using RT-qPCR analysis (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). All forward and reverse
primers were synthesized by Invitrogen; Thermo Fisher Scientific,
Inc. (Table I). RT-qPCR was
performed using a TaqPath™ 1-Step RT-qPCR Master Mix RT-qPCR kit
(cat. no. A15300; Thermo Fisher Scientific, Inc.). Thermocycling
conditions included 45 amplification cycles, denaturation at 95°C
for 120 sec, primer annealing at 62.5°C for 30 sec with touchdown
to 56.5°C for 45 sec and applicant extension at 72°C for 60 sec).
Changes in relative mRNA expression levels were calculated using
the 2−ΔΔCq method (28). The results are expressed as the
n-fold change, compared with the control (β-actin).
 | Table I.Sequences of primers in the present
study. |
Table I.
Sequences of primers in the present
study.
|
| Sequence |
|---|
|
|
|
|---|
| Gene name | Reverse | Forward |
|---|
| P53 |
5′-TTAAGCTTTTTGCGTTCGGGCTGG-3′ |
5-ATGGTGGCATGAACCTGTGG-3G |
| Bcl-2 |
5-CGTCATAACTAAAGACACCCC-3′ |
5-TTCATCTCCAGTATCCGACT-3′ |
| Caspase-3 |
5′-ATGGAGAACAACAAAACCTCAGT-3′ |
5-TTGCTCCCATGTATGGTCTTTAC-3′ |
| Apaf-1 |
5-AGCAAGTTGGTGTCATCCTCCGAT-3r |
5-ATAGCAACAAAGCTCTCCGGTGGA-3r |
| ALP |
5-CACCAATCACCTGCGGTACA-3G |
5-CAGATCACGTCATCGCAC-3- |
| TNF-α |
5′-CAATCCCTTTATTACCC-3′ |
3′-GTCTTCTCAAGTCCTGC-3′ |
| iNOS |
5′-CTGCAGGTCTTTGACGCTCGG-3′ |
5′-GTGGAACACAGGGGTGATG-3′ |
| COX2 |
5′-TGAACACGGACTTGCTCACTTTG-3′ |
5′-AGGCCTTTGCCACTGCTTGTA-3′ |
| TRAF-1 |
5′-AGAACCCGAGGAATGGCGA-3′ |
5′-TGAAGGAGCAGCCGACACC-3′ |
| IAP |
5′-GGCAGATTATGAAGCACGGATC-3′ |
5′-GGCTTCCAATCAGTTAGCCCTC-3′ |
| β-actin |
5-ACGGTCAGGTCATCACTATCG-3′ |
5′-GGCATAGAGGTCTTTACGGATG-3′ |
Western blot analysis
The osteoblasts from the experimental mice with
osteoporosis treated with Tan-IIA, ADN and PBS were homogenized in
lysate buffer containing protease-inhibitor and were centrifuged at
8,000 × g at 4°C for 10 min. The supernatant of the mixture was
used for analysis of proteins of interest. For detection of
proteins, the transmembrane proteins were extracted using a
transmembrane protein extraction kit (Qiagen Sciences, Inc.)
according to the manufacturer's protocols. Protein concentrations
were determined using a Bicinchoninic Acid protein assay (Pierce;
Thermo Fisher Scientific, Inc.) and protein samples (40 µg) was
separated by 12.5% polyacrylamide gel electrophoresis. Proteins
were transferred onto a nitrocellulose membrane (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The SDS assays were
performed as previously described (29). For the western blot analysis,
primary antibodies: p65 (1:1,000, ab16502; Abcam, Cambridge, UK),
IKK-β (1:1,000, ab178870; Abcam), IκBα (1:1,000, ab109300; Abcam)
and β-actin (1:1,000, ab8227; Abcam) were added following blocking
(5% skimmed milk) for 1 h at 37°C, following which they were
incubated with horseradish peroxidase-conjugated immunoglobulin G
(cat. no. STAR206P; Bio-Rad Laboratories, Inc.) was used at a
1:5,000 dilution for 24 h at 4°C. The results were visualized using
a chemiluminescence detection system.
Immunohistochemical staining
Bone tissues were obtained from experimental mice
after treatment as described previously (30). Immunohistochemical staining was
performed using an avidin-biotin-peroxidase technique.
Paraffin-embedded tissue sections (4 µm) were prepared and epitope
retrieval was performed for further analysis. The paraffinized
sections were subjected to H2O2 (3%) for
10–15 min, and were subsequently blocked in regular blocking
solution for 10–15 min at 37°C. Finally, the sections were
incubated in anti-cluster of differentiation 31 (1:1,200, ab28364;
Abcam), and anti-Ki67 (1:1,200, ab15580; Abcam), respectively, at
4°C for 12 h following blocking. All sections were washed three
times and incubated with horseradish peroxidase-conjugated
Immunoglobulin G (1:2,000, product code: ab97057; Abcam) for 1 h at
37°C and were observed in six randomly selected views under a
fluorescence microscope at 488 nm (Olympus BX41; Olympus
Corporation, Tokyo, Japan).
Oxidative stress assay
The examination of intracellular oxidative stress
was performed using flow cytometry and DCFH-DA. The osteoblasts
were harvested from the experimental mice on day 60, and
trypsinized and labeled with DCFH-DA for 45 min at 37°C.
Subsequently, the osteoblasts were treated with
H2O2 in the presence Tan-IIA, ADN or PBS for
30 min at room temperature. The data were further analyzed with
CellQuest Pro software (version 3.2; BD Biosciences, San Jose, CA,
USA).
Phenotypic characterization of
osteoblast differentiation
The osteoblasts were seeded at a density of
1×105 cells/cm2 for 12 h at 37°C. At 85%
confluence, the cells were cultured in osteogenic medium
(Invitrogen; Thermo Fisher Scientific, Inc.) containing 5% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) with Tan-IIA,
ADN or PBS (1.5 mg/ml) in the presence of the indicated reagents.
The procedures used for characterizing osteoblast differentiation
were as described in a previous study (31).
Statistical analysis
All data are presented as the mean ± standard error
of the mean. Unpaired data were analyzed using Student's t-test.
Comparisons of data between multiple groups were analyzed using
one-way analysis of variance followed by a Bonferroni post hoc
test. Statistical analyses were performed using SPSS 19.0 (IBM
SPSS, Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Tan-IIA ameliorates the decreased
viability of osteoblasts and inhibits osteoclast
differentiation
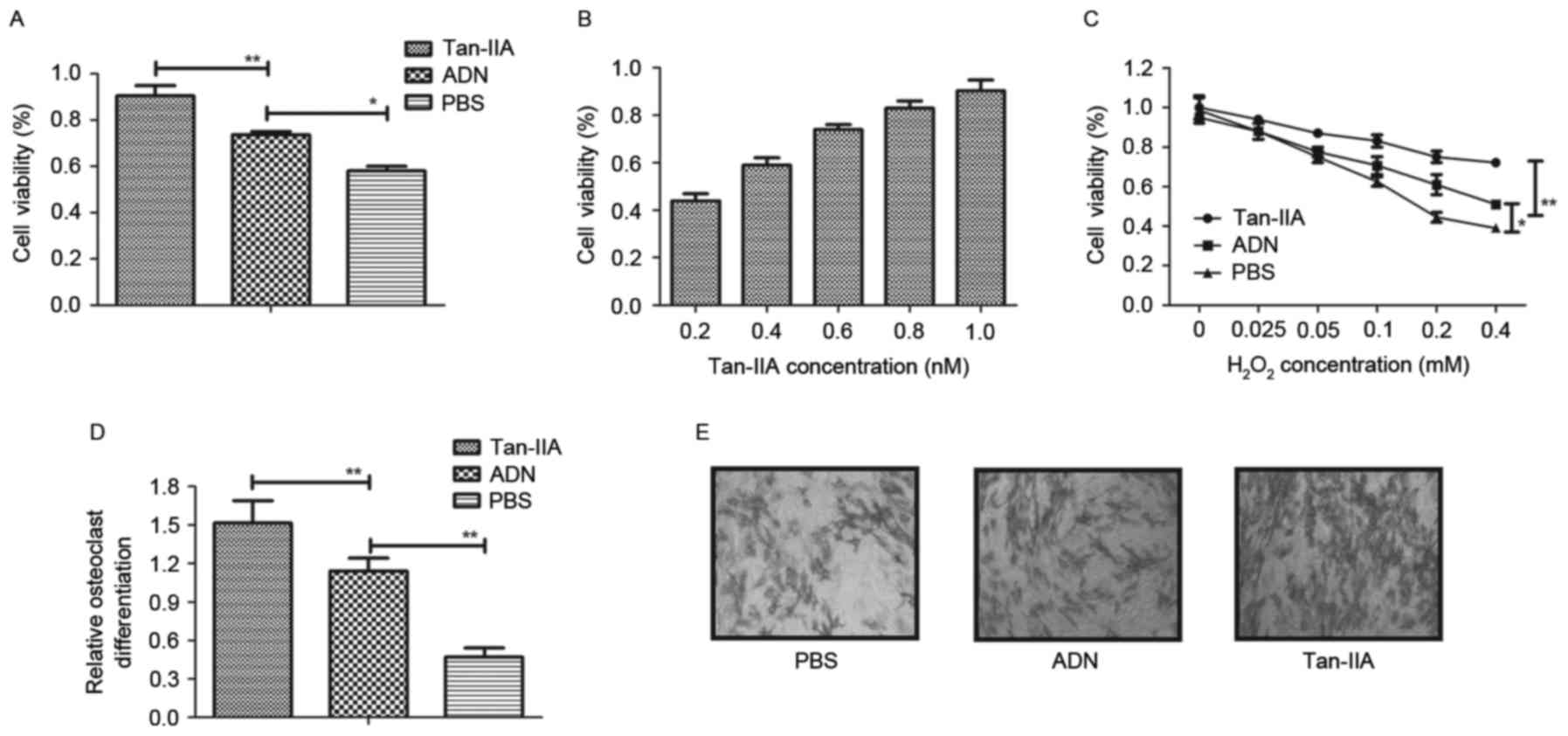
To investigate the effects of Tan-IIA on osteocyte
growth, osteoblasts and osteoclasts were treated with Tan-IIA for
24 h. As shown in Fig. 1A and B,
the results showed that Tan-IIA treatment increased the viability
of the osteoblasts in a dose-dependent manner, compared with ADN
and PBS. The viability of osteoblasts was also improved by Tan-IIA
following treatment with increasing concentrations of
H2O2 between 0.025 and 1.2 mM for 24 h
(Fig. 1C). The present study also
investigated the effects of Tan-IIA on osteoclastogenesis. The
results (Fig. 1D) showed that
Tan-IIA treatment inhibited osteoclast differentiation. The
morphology of osteoclasts also confirmed the efficacy of Tan-IIA on
osteoclast differentiation (Fig.
1E). Collectively, these results suggested that Tan-IIA not
only ameliorated the decreased viability of osteoblasts, but
effectively inhibited osteoclast differentiation.
Tan-IIA treatment inhibits the
bone-resorbing activity and apoptosis of osteoclasts in vitro
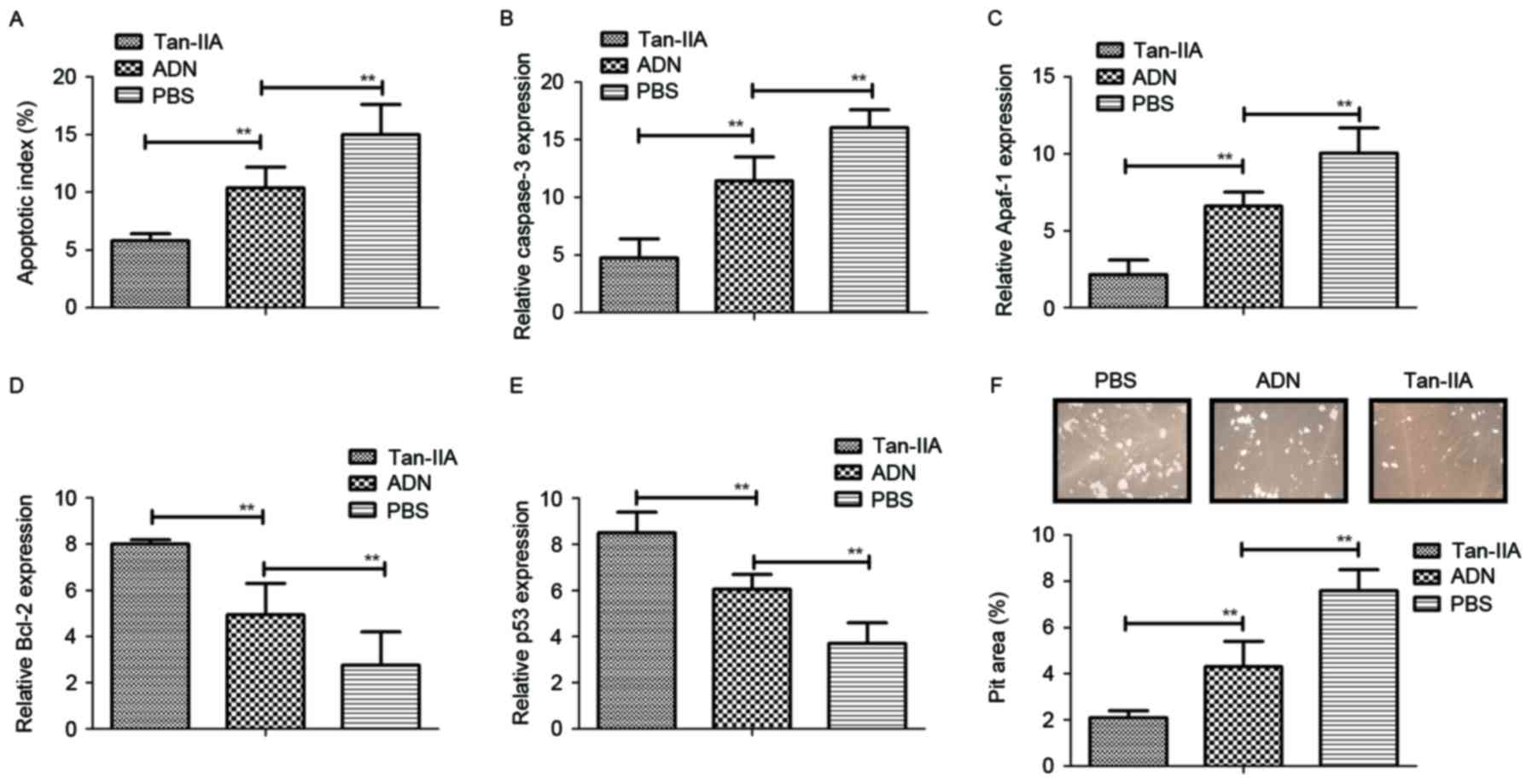
The present study also investigated whether Tan-IIA
affects the function of osteoblasts and osteoclasts. It was found
that Tan-IIA treatment inhibited the apoptosis of osteoblasts
induced by receptor activator of NF-κB ligand (RANKL), whereas the
apoptosis of osteoclasts was promoted by treatment with Tan-IIA
(Fig. 2A). In addition, the
expression levels of caspase-3 and apoptotic protease-activating
factor 1 were significantly upregulated in osteoclasts, but
downregulated in osteoblasts (Fig. 2B
and C). It was also observed that the expression levels of the
anti-apoptotic genes Bcl-2 and p53 were increased and decreased in
osteoblasts, respectively (Fig. 2D and
E). Furthermore, Tan-IIA treatment significantly increased the
bone-resorbing activity for osteoblasts, however, no effects were
observed in osteoclasts in vitro (Fig. 2F). Taken together, these data
suggested that Tan-IIA treatment was beneficial in bone-resorbing
activity through regulation of the apoptosis of osteoblasts and
osteoclasts.
Tan-IIA treatment shows beneficial
effects in mice with osteoporosis
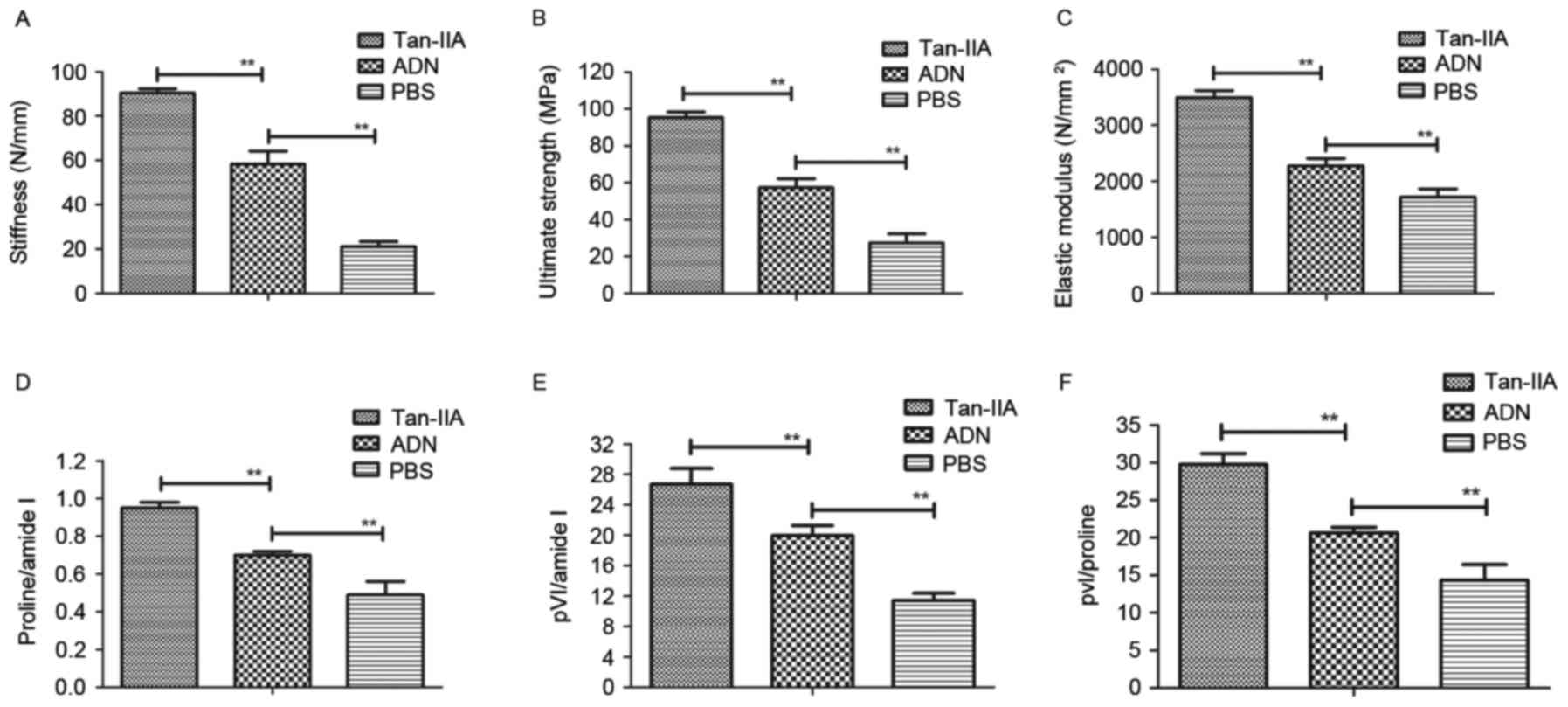
The biomechanical properties of Wnt1sw/sw
bones were examined to assess the in vivo effects of Tan-IIA
on osteoporosis. First, bone strength was assessed in experimental
mice treated with Tan-IIA, ADN and PBS. The data showed that ADN
increased bone strength, compared with that in the PBS group,
however, Tan-IIA increased the bone strength of mice with
osteoporosis, compared with that in the ADN and PBS groups
(Fig. 3A-C). In addition, the
present study analyzed the bone mineral and matrix composition of
the experimental mice treated with Tan-IIA, ADN or PBS. The results
(Fig. 3D) showed that Tan-IIA
treatment led to an increasing ratio of proline to amide I. The
relative mineral content was calculated by the ratio of phosphate
to amide I, which was also increased in the Tan-IIA-treated
Wnt1sw/sw mice (Fig.
3E). The evidence showed that Tan-IIA treatment increased the
ratio of phosphate to proline in the Wnt1sw/sw mice
(Fig. 3F). Overall, these data
suggested that Tan-IIA was beneficial for the treatment of mice
with osteoporosis by decreasing the mineral and collagen
composition of the bone matrix.
Tan-IIA improves osteoporosis by
regulating oxidative stress in osteoblasts from experimental
mice
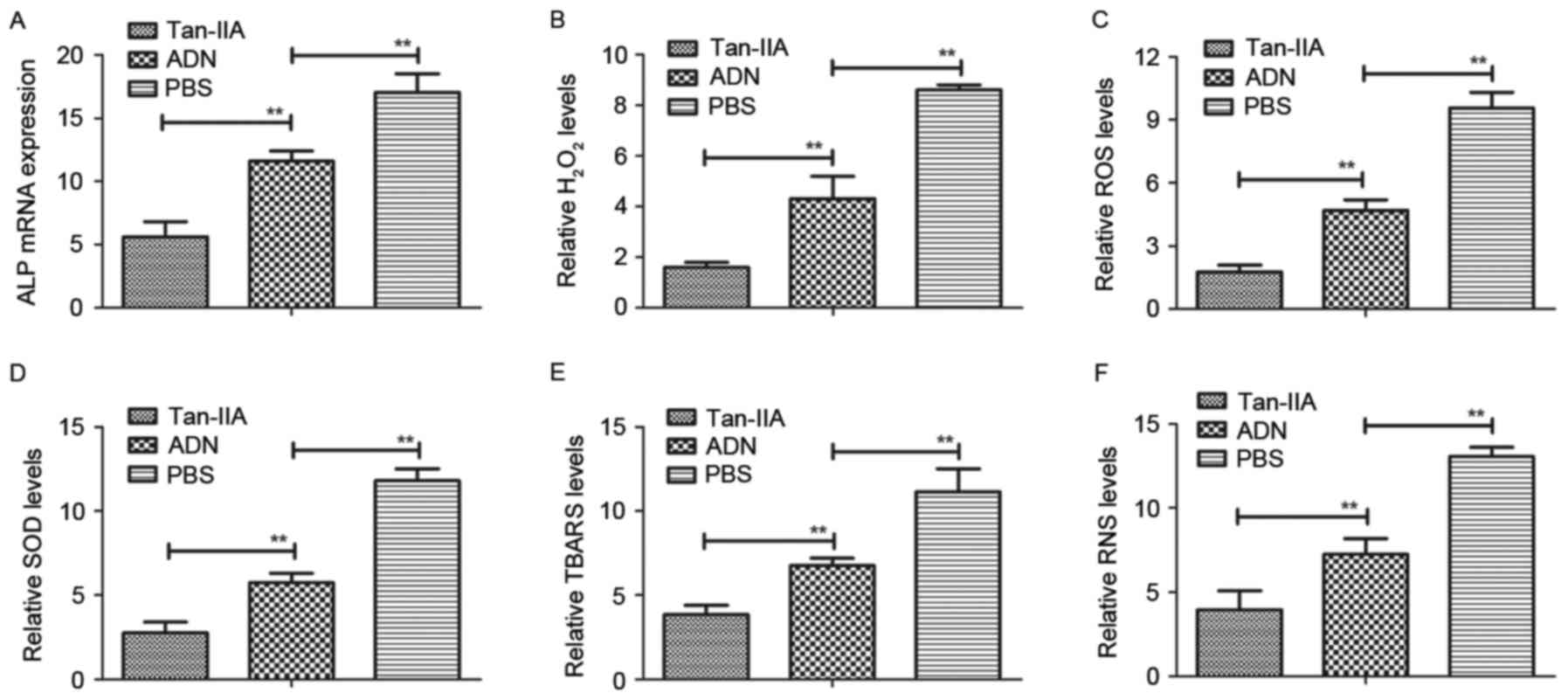
In order to analyze the efficacy of Tan-IIA in
osteoblasts and osteoclasts from experimental mice treated with
Tan-IIA, ADN and PBS, the present study analyzed oxidative stress
in the mice with osteoporosis. The mRNA expression of ALP, a
biomarker of osteoblastogenesis, was downregulated by Tan-IIA
treatment for 24 h (Fig. 4A). The
levels of H2O2 and accumulation of ROS were
also decreased in the osteoblasts from the experimental mice
(Fig. 4B and C). In addition, the
results revealed that the expression levels of superoxide dismutase
(SOD) and thiobarbituric acid reactive substances (TBARS) were
decreased in the osteoblasts from the experimental mice treated
with Tan-IIA, compared with those in mice treated with ADN or PBS
(Fig. 4D and E). The results also
demonstrated that the levels of reactive nitrogen species (RNS)
were decreased in Tan-IIA-mediated oxidative stress in osteoblasts
from the experimental mice (Fig.
4F). These data suggested that Tan-IIA inhibited the
deleterious effects on the osteoblasts of experimental mice
triggered by oxidative stress.
Tan-IIA improves osteoporosis via
regulation of the NF-κB signaling pathway
To understand the mechanism underlying the
Tan-IIA-induced suppression of oxidative stress in osteoblasts, the
present study first analyzed the activity of NF-κB in osteocytes.
The results (Fig. 5A) showed that
Tan-IIA treatment inhibited the activity of NF-κB in osteocytes.
Tan-IIA treatment also suppressed the activation of NF-κB
phosphorylation in the osteoblasts (Fig. 5B). The levels of NF-κB target genes
(TNF-α, iNOS and COX2) were decreased (Fig. 5C-E), and the levels of TRAF-1 and
IAP were increased (Fig. 5F and G)
in osteoblasts. The results of the western blot analysis revealed
that the levels of p65, inhibitor of NF-κB (IκB)α and IκB kinase
(IKK)-β were downregulated following treatment with Tan-IIA
(Fig. 5H). Collectively, these
results indicated that Tan-IIA improved osteoporosis by regulating
the NF-κB signaling pathway.
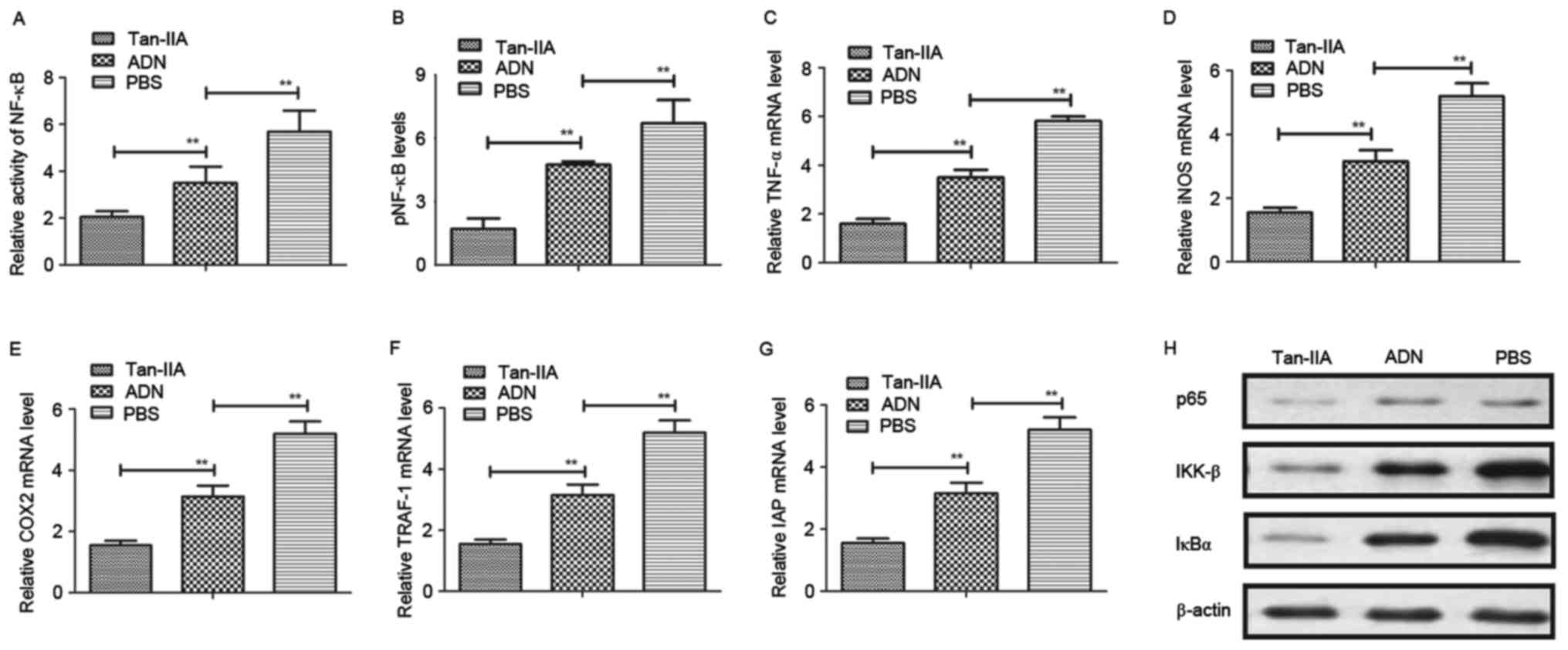 | Figure 5.Tan-IIA improves osteoporosis through
the NF-κB signaling pathway. (A) Evaluation of the activity of
NF-κB in osteoblasts from experimental mice. (B) Activation of
NF-κB phosphorylation in osteoblasts from experimental mice
following treatment with Tan-IIA, ADN and PBS. Expression levels of
(C) TNF-α, (D) iNOS and (E) COX2 in osteoblasts from experimental
mice treated with Tan-IIA, ADN and PBS. Expression levels of (F)
TRAF-1 and (G) IAP in osteoblasts treated with Tan-IIA, ADN and
PBS. (H) Expression levels of p65, IKK-β and IκBα in osteoblasts
following treatment with Tan-IIA, ADN and PBS. Data are presented
as the mean ± standard error of the mean. **P<0.01. ADN,
alendronate; Tan-IIA, tanshinone IIA; NF-κB, nuclear factor-κB;
pNF-κB, phosphorylated NF-κB; TNF-α, tumor necrosis factor-α; iNOS,
inducible nitric oxide synthase; COX2, cyclooxygenase 2; TRAF-1,
tumor necrosis factor receptor-associated factor 1; IAP, inhibitor
of apoptosis protein, IκBα, inhibitor of NF-κBα; IKK-β, IκB
kinase-β. |
Discussion
Osteoporosis is a comprehensive disease occurring in
skeletal tissues, which is increasing in the aging population
(32). The clinical consequences
of osteoporosis include fractures of the upper extremities, hip and
even spine, resulting in loss of function and independence,
impaired quality of life, and increasing morbidity and mortality
rates (33). Therefore,
understanding the molecular mechanism of osteoporosis and
identifying more efficient drugs for the treatment of osteoporosis
are important for the clinical treatment of osteoporosis. In the
present study, the efficacy and mechanism of Tan-IIA on osteocytes
were investigated using a mice model of osteoporosis. In addition,
osteocyte viability and osteoclast differentiation were examined to
identify the therapeutic effects of Tan-IIA on osteoporosis. The
effects of Tan-IIA on oxidative stress were investigated in
vitro and in vivo. Notably, the mechanism underlying the
Tan-IIA-mediated signaling pathway was analyzed. The data obtained
suggested that Tan-IIA exhibited protective effects against
oxidative stress in osteoblast differentiation in mice with
osteoporosis via regulation of the NF-κB signaling pathway. The
results also indicated that oxidative stress was important in the
progression of osteoporosis and may be an efficient target,
consistent with previous reports (34,35).
Previous reports have shown that the excessive
accumulation of ROS and the subsequent activation of oxidative
stress act as contributory functions in the development and
progression of osteoporosis in the skeletal system (36,37).
The involvement of the production of ROS in age-related
osteoporosis and glucocorticoid-induced osteoporosis have been well
documented (23). In addition,
evidence has indicated that oxidative stress can damage various
cellular components of osteoblasts, and contribute to the
aggravation of osteoporosis (22,38).
Oxidative stress is also considered an important pathogenic factor
on bone mineral density and bone loss (20). Increasing evidence has shown that
cell autophagy is important in the response to oxidative stress
(21), and oxidative damage to
osteoblasts can be considered a target for alleviating the
condition of osteoporosis through the endoplasmic reticulum stress
pathway (39). In the present
study, the efficacy of Tan-IIA on the improvement of osteoblasts
was confirmed, and it was also shown that Tan-IIA promoted
osteoblastic differentiation through oxidative stress, which may be
associated with changes in bone matrix formation and bone
mineralization in the progression bone rarefaction.
Previous investigations have been performed to
understand the Tan-IIA-associated therapeutic regimen via targeting
specific molecules and disrupting the dopaminergic system, leading
to various symptoms of cardiovascular diseases (40,41).
However, the molecular mechanism of Tan-IIA in mediating the
treatment of osteoporosis has not been investigated previously. Lee
et al (42) suggested that
the activation of NF-κB was associated with osteoclast
differentiation, and their investigations indicated that the
downregulation of RANKLinduced osteoclast differentiation assisted
in the recovery of osteoporosis through inhibiting IκB degradation.
In the present study, the findings indicated that Tan-IIA treatment
may be a potential drug candidate for the treatment of
osteoporosis. The results suggested that Tan-IIA inhibited the
deleterious outcomes triggered by oxidative stress. In addition,
Tan-IIA inhibited the activation of NF-κB and its target genes,
TNF-α, iNOS and COX2, and increased the expression of TRAF-1 and
IAP1/IAP2 in osteocytes.
In conclusion, the present study aimed to elucidate
the underlying mechanisms responsible for Tan-IIA-mediated
anti-apoptotic effects, and its effects on oxidative stress and
signaling pathways. The investigation was extended to understand
the Tan-IIA-mediated improvement of osteoporosis in a mouse model.
The results confirmed the protective effects of Tan-IIA against
oxidative stress and its beneficial effect on osteoblast
differentiation. Extensive evidence has indicated oxidative stress
as a novel mechanism, which contributes to the development and
progression of osteoporosis, with its increase leading to
degenerative disease in osteoporosis (43,44).
The results of the present study confirmed those of previous
reports, demonstrating that Tan-IIA ameliorated the apoptosis of
osteoblasts and improved osteoblast function through the NF-κB
signaling pathway. Future investigations of Tan-IIA may clarify the
most promising outcomes to be investigated as a potential agent for
osteoporosis.
Acknowledgements
Not applicable.
Funding
This study was supported by grant from the Natural
Science Foundation of Tianjin (grant no. 2011TJH1104PU).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
SZ is the executor for this paper and performed most
of the experiments. WW designed the experiments. ZL, YY and HJ
prepared the figures and conducted the data statistics.
Ethics approval and consent to
participate
All experimental protocols on animals were performed
in accordance with the National Institutes of Health (44) and approved by the Committee on the
Ethics of Animal Experiments Defence Research of Tianjin Medical
University General Hospital (Tianjin, China). All surgery and
euthanasia were made to minimize suffering.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cornelius C, Koverech G, Crupi R, Di Paola
R, Koverech A, Lodato F, Scuto M, Salinaro AT, Cuzzocrea S,
Calabrese EJ and Calabrese V: Osteoporosis and alzheimer pathology:
Role of cellular stress response and hormetic redox signaling in
aging and bone remodeling. Front Pharmacol. 5:1202014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kondo T, Endo I and Matsumoto T: The
pathology, diagnosis and therapy on osteoporosis. Nihon Rinsho.
72:1774–1779. 2014.(In Japanese). PubMed/NCBI
|
|
3
|
Scott LJ: Denosumab: A review of its use
in postmenopausal women with osteoporosis. Drugs Aging. 31:555–576.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu S, Liu F, Cheng Z and Wang Q:
Association between osteoporosis and benign paroxysmal positional
vertigo: A systematic review. BMC Neurol. 14:1102014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Henriksen K, Bollerslev J, Everts V and
Karsdal MA: Osteoclast activity and subtypes as a function of
physiology and pathology-implications for future treatments of
osteoporosis. Endocr Rev. 32:31–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sadat-Ali M, Al-Omran A, Al-Bakr W, Azam
MQ, Tantawy A and Al-Othman A: Established osteoporosis and gaps in
the management: Review from a teaching hospital. Ann Med Health Sci
Res. 4:198–201. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miyazaki T, Tokimura F and Tanaka S: A
review of denosumab for the treatment of osteoporosis. Patient
Prefer Adherence. 8:463–471. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li F, Han G and Wu K: Tanshinone IIA
alleviates the AD phenotypes in APP and PS1 transgenic mice. Biomed
Res Int. 2016:76318012016.PubMed/NCBI
|
|
9
|
Kim EO, Kang SE, Im CR, Lee JH, Ahn KS,
Yang WM, Um JY, Lee SG and Yun M: Tanshinone IIA induces TRAIL
sensitization of human lung cancer cells through selective ER
stress induction. Int J Oncol. 48:2205–2212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Su CC and Chiu TL: Tanshinone IIA
decreases the protein expression of EGFR and IGFR blocking the
PI3K/Akt/mTOR pathway in gastric carcinoma AGS cells both in
vitro and in vivo. Oncol Rep. 36:1173–1179. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang Q, Lu W, Yang K, Hadadi C, Fu X,
Chen Y, Yun X, Zhang J, Li M, Xu L, et al: Sodium tanshinone IIA
sulfonate inhibits hypoxia-induced enhancement of SOCE in pulmonary
arterial smooth muscle cells via the PKG-PPAR-γ signaling axis. Am
J Physiol Cell Physiol. 311:C136–C149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mao C, Zhang Y, Cao L, Shao H, Wang L, Zhu
L and Xu Z: The effect of tanshinone IIA on the cardiovascular
system in ovine fetus in utero. AM J Chin Med. 37:1031–1044. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma S, Liu W, Liu P, Liu J, Chen L and Qin
C: Tanshinone IIA treatment alleviated the rat gingival connective
tissue overgrowth induced by cyclosporine A. J Periodontal Res.
51:567–576. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li DC, Bao XQ, Sun H and Zhang D: Research
progress in the study of protective effect of tanshinone IIA on
cerebral ischemic stroke. Yao Xue Xue Bao. 50:635–639. 2015.(In
Chinese). PubMed/NCBI
|
|
15
|
Ouyang DS, Huang WH, Chen D, Zhang W, Tan
ZR, Peng JB, Wang YC, Guo Y, Hu DL, Xiao J and Chen Y: Kinetics of
cytochrome P450 enzymes for metabolism of sodium tanshinone IIA
sulfonate in vitro. Chin Med. 11:112016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bai Y, Zhang L, Fang X and Yang Y:
Tanshinone IIA enhances chemosensitivity of colon cancer cells by
suppressing nuclear factor-κB. Exp Ther Med. 11:1085–1089. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kurian GA, Rajagopal R, Vedantham S and
Rajesh M: The role of oxidative stress in myocardial ischemia and
reperfusion injury and remodeling: Revisited. Oxid Med Cell Longev.
2016:16564502016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saito M: On ‘2015 guidelines for
prevention and treatment of osteoporosis’. The mechanism of bone
fragility. Clin Calcium. 25:1301–1306. 2015.(In Japanese).
PubMed/NCBI
|
|
19
|
Koga T and Takayanagi H: On ‘2015
guidelines for prevention and treatment of osteoporosis’. Cellular
mechanism and etiology of osteoporosis. Clin Calcium. 25:1293–1300.
2015.(In Japanese). PubMed/NCBI
|
|
20
|
Wu Q, Zhong ZM, Pan Y, Zeng JH, Zheng S,
Zhu SY and Chen J: Advanced oxidation protein products as a novel
marker of oxidative stress in postmenopausal osteoporosis. Med Sci.
21:2428–2432. 2015.
|
|
21
|
Yang YH, Li B, Zheng XF, Chen JW, Chen K,
Jiang SD and Jiang LS: Oxidative damage to osteoblasts can be
alleviated by early autophagy through the endoplasmic reticulum
stress pathway-implications for the treatment of osteoporosis. Free
Radic Biol Med. 77:10–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Spilmont M, Leotoing L, Davicco MJ,
Lebecque P, Mercier S, Miot-Noirault E, Pilet P, Rios L, Wittrant Y
and Coxam V: Pomegranate and its derivatives can improve bone
health through decreased inflammation and oxidative stress in an
animal model of postmenopausal osteoporosis. Eur J Nutr.
53:1155–1164. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wilson C: Bone: Oxidative stress and
osteoporosis. Nat Rev Endocrinol. 10:32014. View Article : Google Scholar
|
|
24
|
Couzin-Frankel J: National Institutes of
Health. Needed: More females in animal and cell studies. Science.
344:6792014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kushwaha P, Khedgikar V, Ahmad N, Karvande
A, Gautam J, Kumar P, Maurya R and Trivedi R: A neoflavonoid
dalsissooal isolated from heartwood of Dalbergia sissoo Roxb. has
bone forming effects in mice model for osteoporosis. Eur J
Pharmacol. 788:65–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shum LC, White NS, Nadtochiy SM, Bentley
KL, Brookes PS, Jonason JH and Eliseev RA: Cyclophilin D knock-out
mice show enhanced resistance to osteoporosis and to metabolic
changes observed in aging bone. PLoS One. 11:e01557092016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gartland A, Rumney RM, Dillon JP and
Gallagher JA: Isolation and culture of human osteoblasts. Methods
Mol Biol. 806:337–355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wai-Hoe L, Wing-Seng L, Ismail Z and
Lay-Harn G: SDS-PAGE-based quantitative assay for screening of
kidney stone disease. Biol Proced Online. 11:pp. 145–160. 2009;
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Williams EL, White K and Oreffo RO:
Isolation and enrichment of Stro-1 immunoselected mesenchymal stem
cells from adult human bone marrow. Methods Mol Biol. 1035:67–73.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Elsafadi M, Manikandan M, Dawud RA, Alajez
NM, Hamam R, Alfayez M, Kassem M, Aldahmash A and Mahmood A:
Transgelin is a TGFβ-inducible gene that regulates osteoblastic and
adipogenic differentiation of human skeletal stem cells through
actin cytoskeleston organization. Cell Death Dis. 7:e23212016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lambert MF, Cook JV, Roelant E, Bradshaw
C, Foy R and Eccles MP: Feasibility of applying review criteria for
depression and osteoporosis national guidance in primary care. Prim
Health Care Res Dev. 15:396–405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ghosh M and Majumdar SR: Antihypertensive
medications, bone mineral density and fractures: A review of old
cardiac drugs that provides new insights into osteoporosis.
Endocrine. 46:397–405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sanchez-Rodriguez MA, Ruiz-Ramos M,
Correa-Muñoz E and Mendoza-Núñez VM: Oxidative stress as a risk
factor for osteoporosis in elderly Mexicans as characterized by
antioxidant enzymes. BMC Musculoskelet Disord. 8:1242007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chavan SN, More U, Mulgund S, Saxena V and
Sontakke AN: Effect of supplementation of vitamin C and E on
oxidative stress in osteoporosis. Indian J Clin Biochem.
22:101–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yin H, Shi ZG, Yu YS, Hu J, Wang R, Luan
ZP and Guo DH: Protection against osteoporosis by statins is linked
to a reduction of oxidative stress and restoration of nitric oxide
formation in aged and ovariectomized rats. Eur J Pharmacol.
674:200–206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baek KH, Oh KW, Lee WY, Lee SS, Kim MK,
Kwon HS, Rhee EJ, Han JH, Song KH, Cha BY, et al: Association of
oxidative stress with postmenopausal osteoporosis and the effects
of hydrogen peroxide on osteoclast formation in human bone marrow
cell cultures. Calcif Tissue Int. 87:226–235. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cervellati C, Bonaccorsi G, Cremonini E,
Romani A, Fila E, Castaldini MC, Ferrazzini S, Giganti M and
Massari L: Oxidative stress and bone resorption interplay as a
possible trigger for postmenopausal osteoporosis. Biomed Res Int.
2014:5695632014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yilmaz N and Eren E: Homocysteine
oxidative stress and relation to bone mineral density in
post-menopausal osteoporosis. Aging Clin Exp Res. 21:353–357. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Puntmann VO, Taylor PC and Mayr M:
Coupling vascular and myocardial inflammatory injury into a common
phenotype of cardiovascular dysfunction: Systemic inflammation and
aging-a mini-review. Gerontology. 57:295–303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ramachandran A, Jha S and Lefer DJ: Review
paper: Pathophysiology of myocardial reperfusion injury: The role
of genetically engineered mouse models. Vet Pathol. 45:698–706.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee WS, Lee EG, Sung MS, Choi YJ and Yoo
WH: Atorvastatin inhibits osteoclast differentiation by suppressing
NF-κB and MAPK signaling during IL-1β-induced osteoclastogenesis.
Korean J Intern Med. Mar 28–2017.(Epub ahead of print).
|
|
43
|
Handzlik-Orlik G, Holecki M, Wilczynski K
and Dulawa J: Osteoporosis in liver disease: Pathogenesis and
management. Ther Adv Endocrinol Metab. 7:128–135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim IH, Chung MY, Shin JY and Han D:
Protective effects of black hoof medicinal mushroom from Κorea,
Phellinus linteus (higher basidiomycetes), on osteoporosis
in vitro and in vivo. Int J Med Mushrooms. 18:39–47. 2016.
View Article : Google Scholar : PubMed/NCBI
|