Introduction
β2-GP I, additionally termed apolipoprotein H, is a
glycoprotein present in human plasma. It combines with fat and
results in formation of chylomicron, very low-density lipoprotein
and high-density lipoprotein (1).
β2-GP I is primarily synthesized in the liver and in the placenta
during pregnancy. Previous studies have demonstrated that β2-GP I
binds to phosphatidylserine on the surface of apoptotic cells,
competitively with Annexin A5, to enhance the phagocytic rate of
phagocytes (2,3). It has been demonstrated that in the
hepatocellular carcinoma cell line SMMC-7721, β2-GP I mediates the
process of hepatitis B viral invasion through interaction with cell
membrane receptor annexin A2II (4). β2-GP I has a broad biological
significance, and research remains in progress.
Antiphospholipid syndrome (APS) is an autoimmune
disease that results from upregulation of antiphospholipid antibody
(aPL) in serum. Anticardiolipin antibody (aCL), lupus anticoagulant
(LA) and aβ2-GP I are three forms of aPL, and each may lead to the
disease (5). In patients with APS,
aPL persists in the serum and results in thrombocytopenia,
recurrent abortion, and arterial and venous thrombosis (6–8).
Particularly, aβ2-GPI is a structural domain of aCL and LA, acting
as an antibody to β2-GP I (9). The
mechanism underlying the regulation of expression of aβ2-GPI
remains to be elucidated, however it has been reported to be
associated with viral infection (10). In a study by Raschi et al
(11), lipopolysaccharide promotes
the binding of aβ2-GP I to β2-GP I which induces an effect on
pathogenesis of APS through signal transduction by cell membrane
toll-like receptor 4 (TLR4). Immunization of β2-GP I+/+
and β2-GP I−/− mice with aβ2-GP I antibody revealed that
only β2-GP I+/+ mice with fetal loss may be detected
(12). The aforementioned studies
suggest that aβ2-G I/β2-GP I complex may serve a role in the
pathogenesis of APS. However, further evidence is necessary to
support this hypothesis.
The present study determined the effect of aβ2-GP
I/β2-GP I complex on JEG-3 cell proliferation, migration and
invasion and the resulting molecular alterations of the nuclear
factor (NF)-κB signaling pathway. Recombinant human (rh)β2-GP I was
expressed in a prokaryotic expression system and human aβ2-GP I was
purified from serum of patients with recurrent pregnancy loss.
Subsequently, cell counting kit-8 (CCK-8), cell cycle and transwell
assays, and EdU staining were carried out to detect the effect of
aβ2-GP I/rhβ2-GP I complex on JEG-3 cells. Furthermore, effect of
alterations of the aβ2-GP I/rhβ2-GP I complex on the NF-κB
signaling pathway were investigated. The results demonstrated a
potential role of the aβ2-GP I/rhβ2-GP I complex in the
aforementioned processes.
Materials and methods
Cell culture
Human choriocarcinoma cell line JEG-3 and human
hepatocarcinoma cell line Huh-7 was purchased from the BeNa Culture
Collection (Shanghai, China). Cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% (vol/vol) FBS (Gibco;
Thermo Fisher Scientific, Inc.) and 100 nM penicillin/streptomycin
in a 5% CO2 incubator at 37°C.
Stably transfect cell lines
The PGMLV-NF-κB-Lu vector was purchased from
GenomeDitech Co., Ltd. (Shanghai, China). JEG-3 cell line was
transfected using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. For 12-well plates, 1 µg PGMLV-NF-κB-Lu vector and 5 µl
Lipofectamine® 2000 was added to each well. Positive
clones were screened using 1 µg/ml of puromycin. For amplification,
10 positive clones were selected randomly and cultured in 12-well
plates independently. Further screening was performed using tumor
necrosis factor-α (TNF-α, Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) at 0, 5 and 10 ng/ml, and detected fluorescein expression
level by a luciferase assay kit (Pierce; Thermo Fisher Scientific,
Inc.). Positive clones with elevated expression level of
fluorescein at 5 ng/ml TNF-α were selected and two cell clones with
high fluorescein signal were selected, and called JEG-3-NFkB-Luc1
cells and JEG-3-NFkB-Luc2 cells, respectively.
Plasmid encoding β2-GP I
Based on the sequence information from the National
Center for Biotechnology Information database (13), the β2-GP I gene (GenBank: X57847.1)
was amplified using PCR with primers designed to generate Hind III
and BamH I restriction sites at the 5′ and 3′ends of the amplified
fragments, respectively. The cDNA from Huh-7 cells was used as
template and the following primers were used:
5′-CGCGGATCCATGATTTCTCCAGT-3′ and 5′-CCCAAGCTTTTAGCATGGCTTTAC-3′;
RNA extraction was conducted as described below. The following
thermocycling conditions were used for the PCR: Initial
denaturation for 1 min at 95°C; 30 cycles of 95°C for 10 sec, 55°C
for 10 sec and 72°C for 60 sec, using 2X PCR Solution Premix Taq™
(Takara Bio, Inc., Otsu, Japan). The amplified β2-GP I products
were digested with HindIII and BamHI restriction
enzymes (Takara Bio, Inc.) and cloned into multiple clone sites of
pET-44a (+) (Novagen; Merck KGaA).
Purification of the His-tagged
protein
In order to construct the prokaryotic expression
vector, the structural gene of human β2-GP I protein was amplified
by PCR and inserted into the pET-44a (+) plasmid. Following the
aforementioned procedure, 1 ng of constructed vector transfected
into E. coli BL21 (DE3, DE3:T7 RNA polymerase gene) cells
(Vazyme, Piscataway, NJ, USA) and one single clone from the solid
agar Petri dish (containing 100 ug/ml ampicillin), was amplified in
liquid agar medium (containing 100 ug/ml ampicillin). The vector of
pET-44a(+) encoding the His-tag sequence and the expression of
fusion protein rhβ2-GP I was induced by 0.1 mM isopropyl
β-D-1-thiogalactopyranoside (IPTG; Sigma-Aldrich; Merck KGaA); the
bacteria were incubated at 25°C, 100 rpm/min for 2, 4 or 6 h.
Subsequently, the bacteria was lysed with lysis buffer (50 mM
NaH2PO4, 300 mM NaCl, 5 mM imidazole and 1
mg/ml lysozyme, pH=8.0) then sonicated 6 times at 200 W in 10 sec
bursts with a 10 sec cooling period between each burst. The induced
protein in the lysate was further purified by Ni-NTA agarose
(Qiagen, Germantown, MD, USA) and identified by western blotting as
described below. Anti-6X His tag antibody (cat: ab213204, 1:5,000
dilution) and horseradish peroxidase (HRP)-conjugated goat
anti-rabbit immunoglobulin (Ig)G (H+L) secondary antibody (1:5,000
dilution) were obtained from Abcam (Cambridge, MA, USA) and used to
detected the His-tagged protein. Protein concentration was
determined using Bicinchoninic Acid assay kit (Pierce; Thermo
Fisher Scientific, Inc.). Coomassie staining was performed to
determine protein induction and purification. Following induction
with IPTG at 2, 4 or 6 h, bacteria from 1 ml bacterial suspension
was collected and lysed with 200 µl SDS-PAGE loading buffer
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China). Following boiling for 10 min, 10 µl of each sample was
separated by 8–10% SDS-PAGE. Then, the gel was stained with
Coomassie staining buffer (Beijing Solarbio Science &
Technology Co., Ltd.) at 37°C for 30 min, and decolored with 10%
acetic acid at 37°C for 1 h. The SDS-PAGE gel images were captured
with an Epson 4990 scanner (Seiko Epson Corporation, Suwa,
Japan).
aβ2-GPI antibody purification
The aβ2-GP I antibody was purified from the serum of
10 patients with fetal loss with elevated levels of the aβ2-GP I
antibody. Mixing of the serum and purification of the aβ2-GPI
antibody were carried out using protein A agarose beads
(Sigma-Aldrich; Merck KGaA). The IgG control antibody was purified
from the serum of 10 normal pregnant women and the purification
method was the same as for the aβ2-GP I antibody. All of the 20
women volunteers were enrolled in the present study from The Second
Affiliated Hospital of Harbin Medical University (Heilongjiang,
China) and aged from 22 to 35; samples were collected during March
2016 to May 2016. Subsequently, filtration with a 0.22 µm filter
(EMD Millipore, Billerica, MA, USA) was carried out and endotoxin
contamination was detected by limulus amoebocyte lysate gel-clot
test (limulus reagent; Xiamen Bioendo Technology Co., Ltd; Xiamen;
China) according to the manufacture's protocol. Antibody
concentration was determined by spectrophotometry. All human
studies were approved by the Medical Ethics Board of The Second
Affiliated Hospital of Harbin Medical University (Harbin, China).
All participants signed written informed consent forms prior to the
study.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
TRIzol® reagent (Thermo Fisher
Scientific, Inc.) was used to isolate total RNA from JEG-3 or Huh-7
cells, according to the manufacturer's protocol. Furthermore, a
total of 500 ng RNA sample was de-folded at 70°C for 5 min, then
reverse-transcribed into cDNA at 42°C for 60 min in a 12.5 µl
reaction volume using the M-MLV first stand cDNA synthesis kit
(Omega Bio-Tek, Inc., Norcross, GA, USA), and the qPCR assay was
performed with 0.25 µl cDNA using 2X Supermix (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) using the CFX96 Touch System
(Bio-Rad Laboratories, Inc.). The data was analyzed using CFX
Manager software (version 3.0; Bio-Rad Laboratories, Inc.) and
β-actin gene expression was used as an internal reference. Data was
analyzed using the 2−ΔΔCq method (14). The following primer sequences were
used for the PCR: NF-κB, forward, 5′-GCTTAGGAGGGAGAGCCCAC-3′ and
reverse, 5′-AGGTATGGGCCATCTGCTGT-3′; inhibitor of nuclear factor
kappa-B kinase subunit (IKKβ), forward, 5′-GCTGCAACTGATGCTGATGT-3′
and reverse, 5′-TGTCACAGGGTAGGTGTGGA-3′; IκBα, forward,
5′-AAGTGATCCGCCAGGTGAAG-3′ and reverse, 5′-CTGCTCACAGGCAAGGTGTA-3′;
myeloid differentiation primary response protein MyD88 (MyD88),
forward, 5′-AGCATTGAGGAGGATTGCCA-3′ and reverse,
5′-GCTGGGGCAATAGCAGATGA-3′, and β-actin, forward
5′-CTCCATCCTGGCCTCGCTGT-3′ and reverse 5′-GCTGTCACCTTCACCGTTCC-3′.
The following PCR conditions were used for PCR: Initial
denaturation for 1 min at 95°C; 40 cycles of 95°C for 10 sec, 60°C
for 30 sec and 72°C for 20 sec.
Western blotting
Following incubation of rhβ2-GP I, aβ2-GP I and
negative control IgG at 50 µg/ml, 37°C for 24 h, all five groups
(rhβ2-GP I, aβ2-GP I, IgG, aβ2-GP I/rhβ2-GP I and IgG/rhβ2-GP I) of
JEG-3 cells were washed once with PBS then directly lysed with
SDS-PAGE loading buffer (Beijing Solarbio Science & Technology
Co., Ltd.) for total protein extraction. For each well of the
6-well plate, 100 µl SDS-PAGE loading buffer was applied. After
boiled for 5 min, 10 µl of each sample was separated by 8–10%
SDS-PAGE, and transferred onto polyvinylidene fluoride membranes
(Millipore, Bedford, MA, USA). The membranes were blocked with 5%
skimmed milk for 1 h at room temperature and incubated with
anti-N-cadherin (cat. no. ab76011), anti-E-cadherin (cat. no.
ab40772), anti-γ-catenin (cat. no. ab184919), anti-MyD88 (cat. no.
ab133739), anti-IκBα (cat. no. ab32518), anti-p-IκBα (cat. no.
ab133462) and anti-β-actin (cat. no. ab8227) antibodies,
respectively, at 4°C overnight. All primary antibodies used in the
present study were from Abcam at a 1:1,000 dilution. Subsequently,
the membranes were incubated with HRP-conjugated goat anti-rabbit
IgG (H+L) secondary antibody (1:5,000; Thermo Fisher Scientific,
Inc.) for 60 min at room temperature, followed by visualization of
the target protein using Enhanced Chemiluminescence reagent
(Pierce; Thermo Fisher Scientific, Inc.). Quantitative analysis was
performed using the Image Pro Plus software (version 6.0, Media
Cybernetics, Inc., Rockville, MD, USA).
Cell proliferation
In each well of a 96-well plate, 2×103
JEG-3 cells were cultured as attached monolayers at 37°C in DMEM
with 10% (V/V) FBS. rhβ2-GP I, aβ2-GP I and negative control IgG
were added to cell cultures at 50 µg/ml for 24 h. For the CCK-8
assay (Dojindo Molecular Technologies, Inc., Kumamoto, Japan), 10
µl CCK-8 solution was added to each well. Following 1 h of
incubation, the absorbance was measured at a wavelength of 450 nm.
For the EdU assay, EdU labeling (Hoechst staining) was performed
with EdU labeling kit purchased from Guangzhou RiboBio Co., Ltd.
(Guangzhou, China); 50 µM EdU medium was added to each well and
cells were cultured in an incubator for 1–2 h at 37°C. Subsequent
steps of EdU labeling were conducted according to manufacturer's
protocol. Then, analysis was conducted using an inverted
fluorescence microscope (Olympus IX71, Olympus Corporation, Tokyo,
Japan) with 460–550 nm excitation wavelength. Quantitative analysis
was performed using the Image Pro Plus software (version 6.0, Media
Cybernetics, Inc.).
Transwell invasion and migration
assays
For the Matrigel-based invasion assay, a total of
100 µl Matrigel was added to the upper chamber of the transwell
inserts. For the migration assay, Matrigel was not added to the
upper chamber. JEG-3 cells in logarithmic phase were digested with
trypsin and cell concentration was adjusted to 5×105
cells/ml with serum-free medium. In the upper chamber of the
transwell insert, 100 µl of cell suspension was added to each well.
A total of 800 µl DMEM supplemented with 10% FBS and 50 µg/ml
rhβ2-GP, aβ2-GP I, negative control IgG, aβ2-GP I/rhβ2-GP I or
IgG/rhβ2-GP I were added to the lower chamber and incubated for 24
h. Subsequently, the upper chamber was removed and cells wiped off
the upper surface of the upper chamber. The cells on the lower
surface of the upper chamber were fixed with 4% paraformaldehyde
for 30 min and stained for additional 30 min at room temperature
with 0.2% crystal violet (Sigma-Aldrich; Merck KGaA). Then,
analysis was conducted using an inverted microscope (Olympus IX71,
Olympus Corporation); quantitative analysis was investigated using
the Image Pro Plus software (version 6.0, Media Cybernetics,
USA).
Flow cytometry to detect cell
cycle
Following incubation of 50 µg/ml rhβ2-GP I, aβ2-GP I
and negative control IgG at 37°C for 24 h, all five groups (rhβ2-GP
I, aβ2-GP I, IgG, aβ2-GP I/rhβ2-GP I and IgG/rhβ2-GP I) of JEG-3
cells were washed once with PBS, fixed with 70% ethanol at 4°C for
24 h, stained with propidium iodide/RNase staining buffer solution
(BD Biosciences, San Jose, CA, USA) and analyzed with a flow
cytometer (Beckman Coulter, Inc., Brea, CA, USA) and ModFit
software 3.2.1 (Verity Software House, Inc., Topsham, ME, USA).
Luciferase assay for NF-κB
activation
Either JEG-3-NFkB-Luc1 cells or JEG-3-NFkB-Luc2
cells were plated in 96-well plates at a density of
1×104 cells/well and treated with 50 µg/ml rhβ2-GP I,
aβ2-GP I, negative control IgG, aβ2-GP I/rhβ2-GP I or IgG/rhβ2-GP I
for 24 h. Subsequently cells were washed with PBS and lysed with 50
µl passive lysis buffer. 20 µl lysates were transferred into
96-well white plates and 50 µl D-luciferin working solution was
added to assess luciferase activity with a microplate reader
(Bio-Rad Laboratories, Hercules, CA, USA). The reagent used for
luciferase assay was performed with Firefly Luciferase Glow Assay
kit (Pierce; Thermo Fisher Scientific, Inc.).
Statistical analysis
All results are presented as the mean ± standard
error of the mean. Statistical analysis was performed using SPSS
15.0 software (SPSS, Inc., Chicago, IL, USA). Differences were
analyzed using one-way analysis of variance followed by the Tukey
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
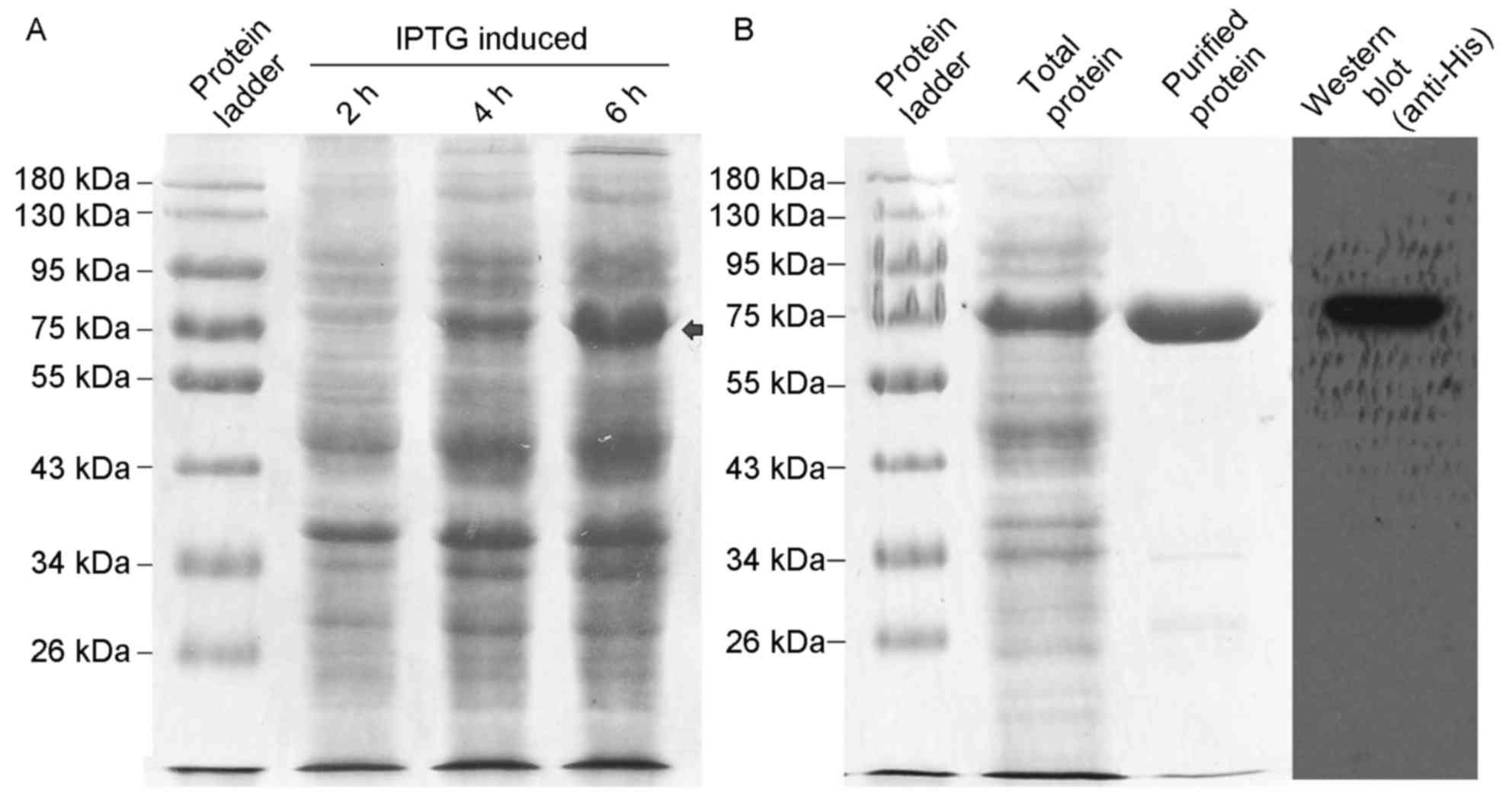
Expression and purification of rhβ2-GP
I
Human β2-GP I is a glycosylated protein present in
plasma. It has been reported that the affinity of aβ2-GP I against
partially glycosylated β2-GP I and completely de-glycosylated
β2-GPI is increased compared with native β2-GP I (15). Based on these results, the present
study used a prokaryotic expression system for obtaining β2-GP I
protein to ensure lack of glycosylation. β2-GP I cDNA was obtained
by PCR amplification and cloned into the multiple cloning sites of
pET-44a(+). The His-tagged fusion protein rhβ2-GP I was
overexpressed in E. coli BL21 (DE3) by induction with
isopropyl β-D-1-thiogalactopyranoside. After 2, 4 and 6 h
incubation with IPTG, the level of induced rhβ2-GP I fusion protein
was increased over time (Fig. 1A,
10% acrylamide). His-tag rhβ2-GP I fusion protein was successfully
purified by the Ni-NTA system and detected by western blot analysis
(Fig. 1B, 10% acrylamide, 1 µg
purified rhβ2-GP I protein was loaded per lane).
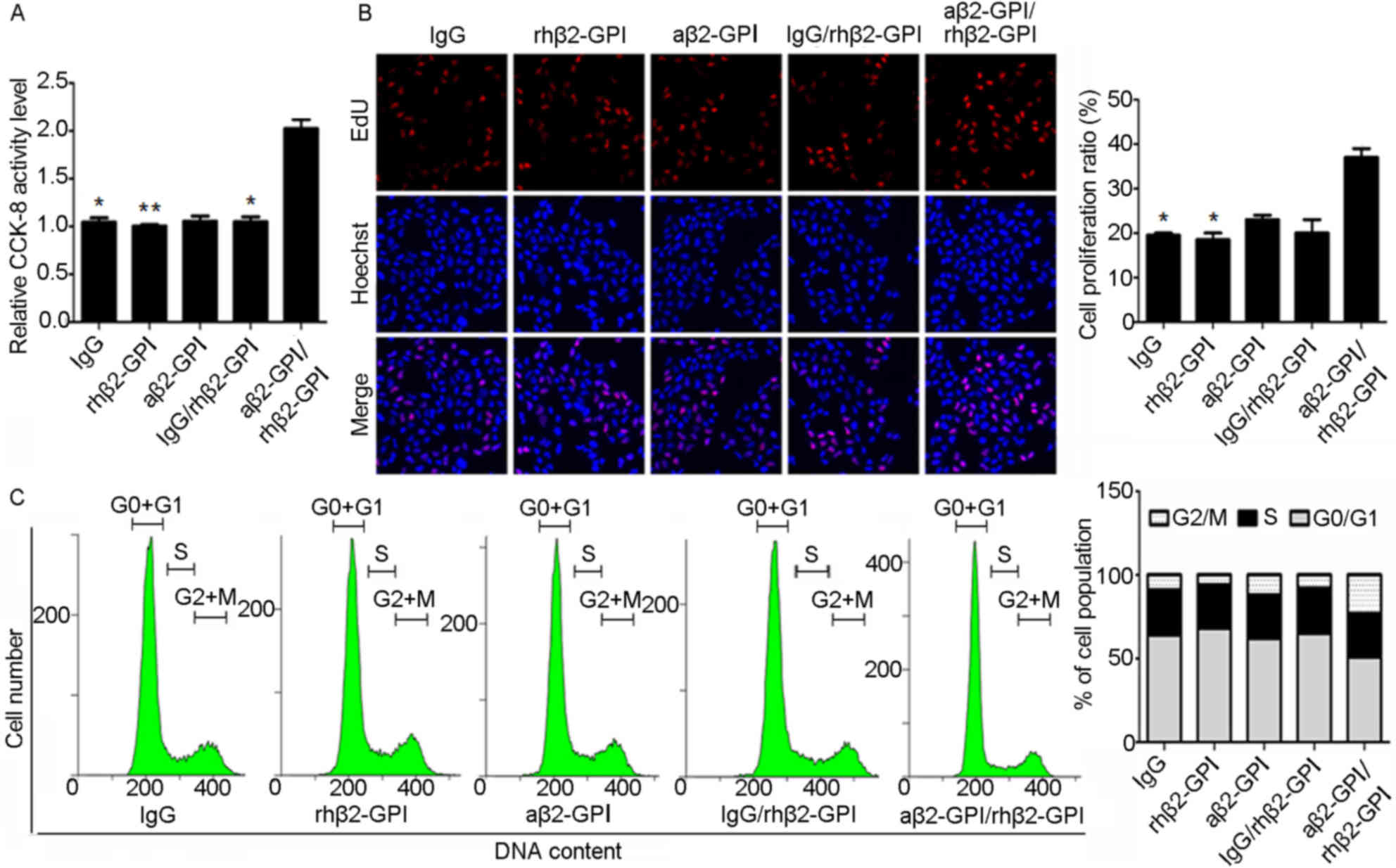
Effect of aβ2-GP I/rhβ2-GP I complex
on JEG-3 cell proliferation
JEG-3 cells are typically used to model
proliferation and migration of human choriocarcinoma, and to
estimate the degree of alteration of tumor malignancy following
experimental treatment. The effect of aβ2-GP I/rhβ2-GP I complex on
JEG-3 cell proliferation was detected by CCK-8 assay and EdU
staining. Compared with the IgG group, CCK-8 activity (Fig. 2A) and the number of EdU-positive
cells (Fig. 2B) increased in the
aβ2-GP I/rhβ2-GP I induced JEG-3 cell group, indicating that aβ2-GP
I/rhβ2-GP I complexes may promote JEG-3 cell proliferation.
Furthermore, cell cycle was analyzed using PI/RNase stain flow
cytometry assay. Compared with the IgG group, the proportion of
cells in the G0/G1 phase was reduced in the
aβ2-GP I/rhβ2-GP I group (Fig.
2C), which suggests that aβ2-GP I/rhβ2-GP I complexes may
promote cell division. The aforementioned results indicated that
the aβ2-GP I/rhβ2-GP I complex may promote JEG-3 cell
proliferation.
Effects of aβ2-GP I/rhβ2-GP I complex
on JEG-3 cell migration and invasion
As previously mentioned, cell migration and invasion
abilities are the characteristics used to estimate the degree of
tumor malignancy. A Matrigel-based transwell assay was used to
determine cell invasion and a transwell assay without matrigel was
used to study cell migration. The number of invaded cells
significantly increased in the aβ2-GP I/rhβ2-GP I group compared
with the all other groups (all P<0.01; Fig. 3A and B). Furthermore, protein
levels of tumor metastasis- and diffusion-associated factors
N-cadherin, E-cadherin and γ-catenin were tested by western blot
analysis (Fig. 3C and D, 8%
acrylamide). In the aβ2-GP I/rhβ2-GP I group, an increase in
E-cadherin (P<0.01) and γ-catenin (P<0.01), and decrease in
N-cadherin (P<0.05) expression levels were observed, compared
with the IgG group. These observations suggest that the aβ2-GP
I/rhβ2-GP I complex increases JEG-3 cell migration and invasion
abilities.
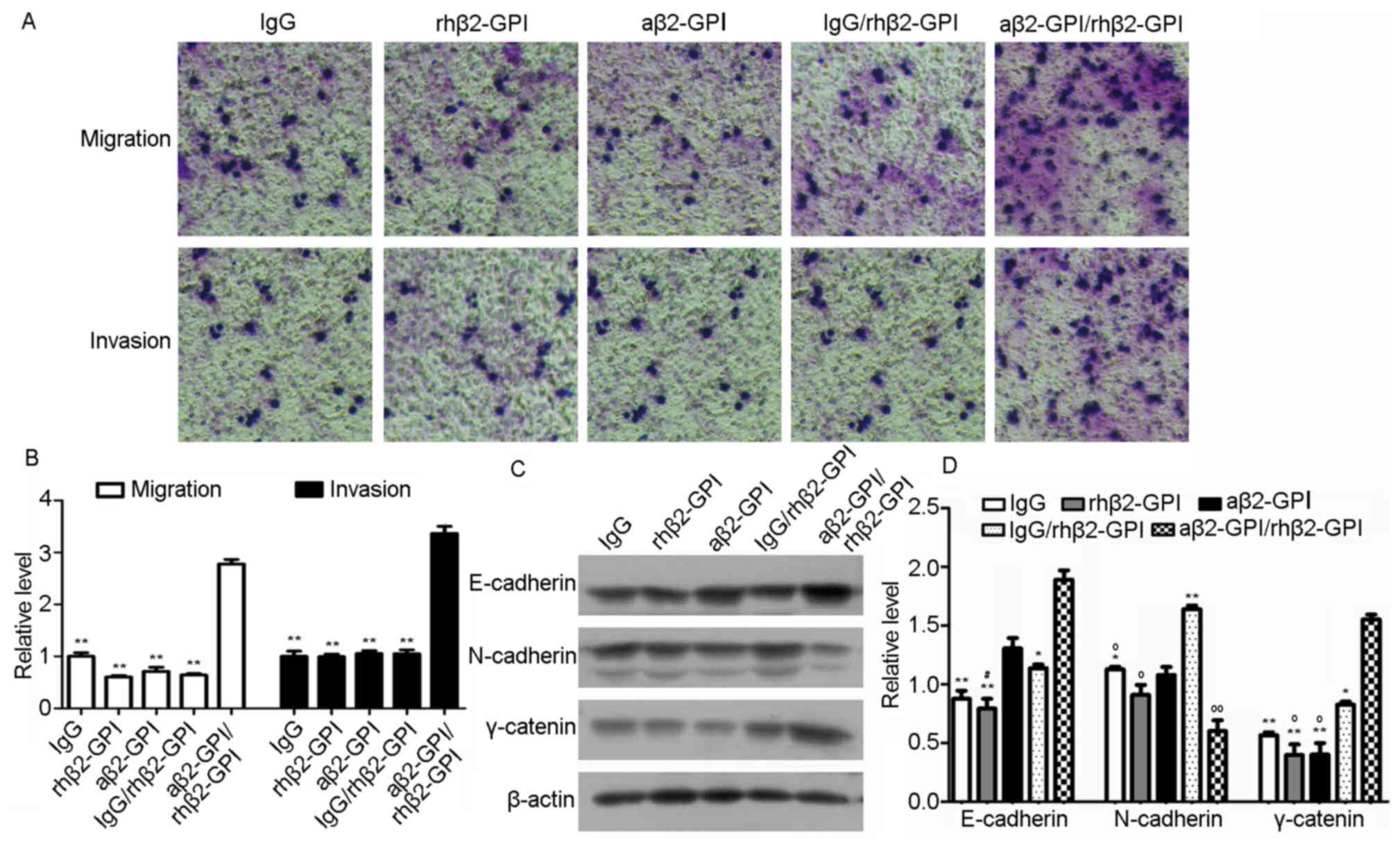 | Figure 3.Effects of the aβ2-GP I/rhβ2-GP I
complex on JEG-3 cell migration and invasion, and protein
expression levels of tumor metastasis- and diffusion-associated
factors. (A and B) Cell migration measured by transwell assay
(magnification, ×200). Statistical analysis of the results of the
transwell assay was performed using Image-Pro Plus 6.0 software. (C
and D) Protein levels of N-cadherin, E-cadherin and γ-caterin were
detected by western blotting. Statistical analysis was performed by
Image-Pro Plus 6.0 software. IgG, rhβ2-GP I, aβ2-GPIand IgG/rhβ2-GP
I were regarded as controls. *P<0.05 and **P<0.01 vs. the
aβ2-GP I/rhβ2-GP I group, oP<0.05 and
ooP<0.01 vs. the IgG/rhβ2-GP I group,
#P<0.05 vs. the aβ2-GP I group. aβ2-GP I,
anti-β2-glycoprotein I antibody; rh, recombinant human; Ig,
immunoglobulin. |
Induction of NF-κB activity in JEG-3
cells stimulated with the aβ2-GP I/rhβ2-GP I complex
To study the effects of the aβ2-GP I/rhβ2-GP I
complex on proliferation, migration and invasion of JEG-3 cells,
alterations in the associated signaling pathways were investigated.
The MyD88 signaling pathway is activated by stimulation of
toll-like receptors (TLRs), which transmit the signal to MyD88
adaptors, which, via downstream biochemical cascades of
protein-protein interactions, leads to induction of NF-κB activity
(16). The activity of NF-κB is
associated with tumor cell proliferation, migration and invasion
(17); TLR2/TLR4 may act as an
endothelial receptor for aβ2-GP I/rhβ2-GP I (18). The aforementioned data suggests
that the aβ2-GP I/rhβ2-GP I complex may be involved in inducing the
MyD88 signaling pathway. RT-qPCR results demonstrated that the
aβ2-GPI/rhβ2-GPI complex increased the expression level of MyD88
(P<0.05), IKKβ (P<0.05), NF-κB (P<0.01) and IκBα
(P<0.05) (Fig. 4A).
Furthermore, western blot analysis was performed to detect the
alterations in MyD88, IκBα and p-IκBα expression levels, and
luciferase assay was used to detect NF-κB activity in JEG-3 cells.
Results of the present study demonstrated that, compared with the
IgG group, the inhibitory effect of IκBα on NF-κB was attenuated by
phosphorylation of IκBα in the aβ2-GP I/rhβ2-GP I treatment group
(Fig. 4B, P<0.01, 10%
acrylamide). A corresponding increase of luciferase activity can be
observed in the aβ2-GP I/rhβ2-GP I group, compared with the IgG,
rhβ2-GP I and IgG/rhβ2-GP I groups (Fig. 4C; all P<0.05). No significant
alterations were detected between the aβ2-GP I/rhβ2-GP I and aβ2-GP
I groups, as aβ2-GP I may exhibit similar effects to that of the
aβ2-GP I on β2-GP I basal expression within JEG-3 cells. The
aforementioned results indicate that the expression of NF-κB may be
increased by the aβ2-GP I/rhβ2-GP I complex.
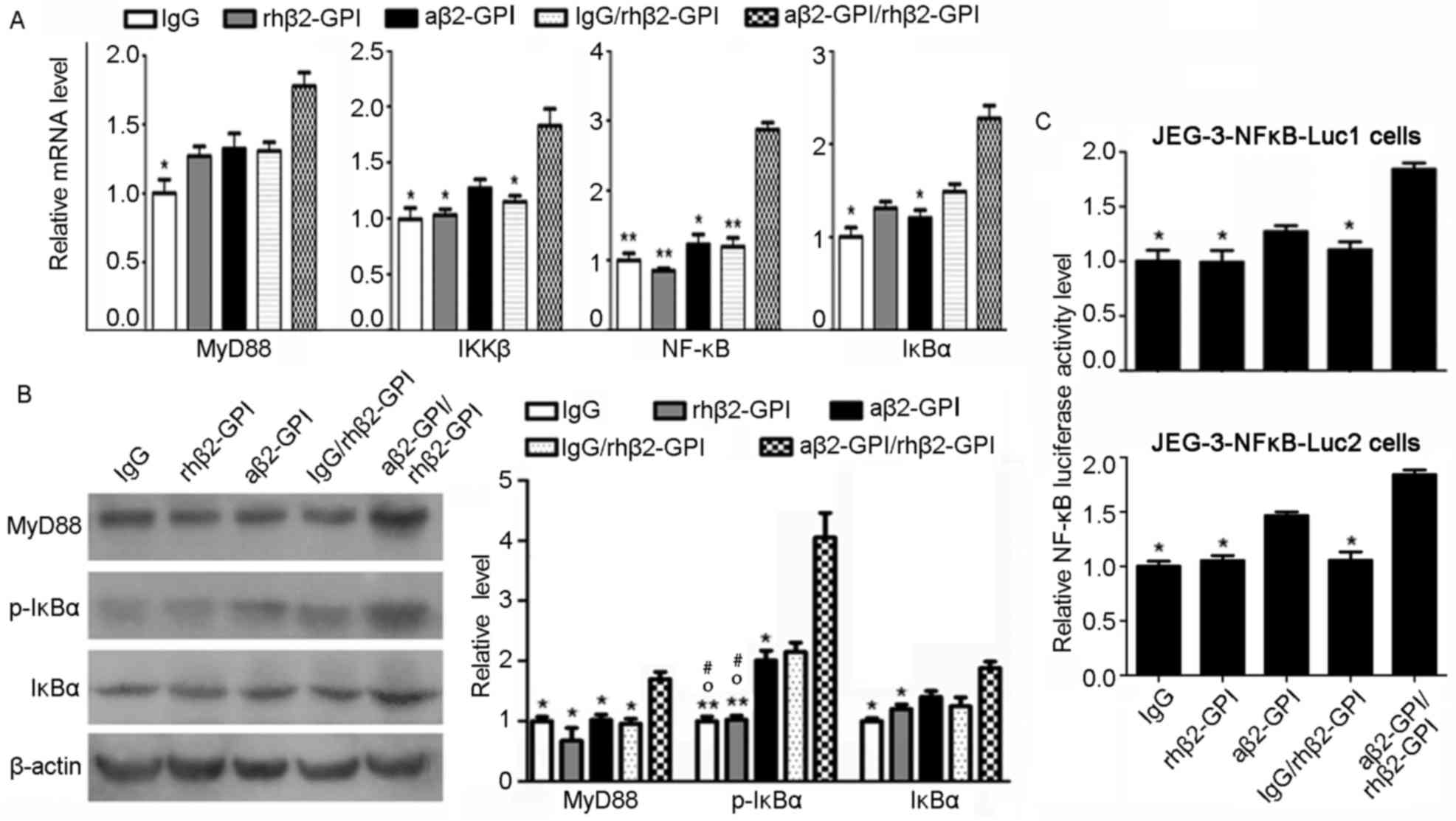 | Figure 4.aβ2-GP I/rhβ2-GP I complex induces the
activity of NF-κB in JEG-3 cells. (A) Relative mRNA expressions of
MyD88, IKKβ, NF-κB and IκBα in JEG-3 cell line. (B) The protein
level of MyD88, IκBα and p-IκBα in stimulated JEG-3 cells detected
by western blotting. The data statistics of western blot test was
performed by Image-Pro Plus 6.0 software. (C) Luciferase activity
was measured to assess NF-κB expression. IgG, rhβ2-GP I, aβ2-GP I
and IgG/rhβ2-GPI were regarded as controls. *P<0.05 and
**P<0.01 vs. the aβ2-GP I/rhβ2-GPI group. oP<0.05
vs. the IgG/rhβ2-GPIgroup. #P<0.05 vs. the aβ2-GP I
group. NF-κB, nuclear factor-κB; aβ2-GP I, anti-β2-glycoprotein I
antibody; rh, human recombinant; MyD88, myeloid differentiation
primary response protein MyD88; IKKβ, inhibitor of nuclear factor
κ-B kinase subunit; Ig, immunoglobulin; IκBα, NF-κB inhibitor
α. |
Discussion
Recurrent pregnancy loss is an APS disease
characterized by persistent presence of aβ2-GP I in serum. β2-GP I
is a apolipoprotein present in plasma composed of five domains,
which is belongs to complement control protein (CCP) superfamily.
The domains I–IV are normal as other family members, conserved and
located at the N-terminus; however, domain V is abnormal and
located at the C-terminus (19,20).
β2-GP I may exist in at least two conformations. In plasma, it is
present as a circular protein in which domain I interacts with
domain V and the epitope is hidden inside the circular structure.
This closed structure is altered to form an open structure when
domain V of β2-GP binds to a lipid layer and domains I–IV expose
the potential binding site for aβ2-GP I (21). Fetal loss may occur when β2-GP I
binds to its antibody aβ2-GP I, which results in thrombogenesis,
hypercoagulability and inhibition of growth, differentiation,
invasion and migration of trophoblast cells (22–24).
However, the mechanism underlying these reactions remains to be
elucidated. Previous studies demonstrated that TLR4 and annexin A2
are the co-receptors of the aβ2-GP I/rhβ2-GPI complex, which
mediates the expression of tissue factor, MyD88, TIR-domain
containing adaptor-inducing interferon-β and tumor necrosis
factor-, resulting in inflammation, thrombus formation, and
pathology of APS (18,25,26).
NF-κB signaling pathway is a signal transduction
pathway in cells that serves a role in inflammatory response, cell
differentiation, apoptosis and tumorigenesis (27). In the present study, the aβ2-GP
I/rhβ2-GP I complex enhanced JEG-3 cell proliferation, migration
and invasion. However, inhibition of cell growth, differentiation,
invasion and migration of trophoblast cells is the primary
demonstration of aβ2-GPI/rhβ2-GP I complex-mediated fetal loss;
opposing results were observed in the present study and may be due
to the tumorigenic nature of trophoblastic JEG-3 cells. Further
analysis on the molecular level, revealed that mRNA levels of
MyD88, IKKβ, IκBα and NF-κB were elevated in the aβ2-GP I/rhβ2-GP I
complex-stimulated JEG-3 cells. Protein levels of MyD88 and p-IκBα
also increased following stimulation with aβ2-GP I/rhβ2-GP,
suggesting that the complex results in phosphorylation of IκBα and
activation of NF-κB. These results are consistent with the
aforementioned results of the luciferase assay performed in the
present study, which demonstrated that NF-κB-luciferase activity is
enhanced in aβ2-GP I/rhβ2-GP I complex-stimulated JEG-3-NFkB-Luc1
and JEG-3-NFkB-Luc2 cells. The results of the present study
indicated that the aβ2-GP I/rhβ2-GP I complex induces the activity
of NF-κB and alters JEG-3 cell proliferation, migration and
invasion.
Trophoblast cells serve a role in pregnancy.
Abnormal myometrial trophoblast invasion and spiral artery
transformation may lead to trophoblast cell damage and apoptosis
(28,29). The abnormality may result in
pre-eclampsia, fetal growth restriction and premature labor with
and without pre-labor rupture of membranes (30–32).
However, the loss of control of cell proliferation, migration and
invasion achieved by trophoblast cells has been associated with
hydatidiform mole and choriocarcinoma (33). The present study demonstrated that
the activity of NF-κB was induced and the ability of cell
proliferation, migration and invasion were enhanced by the aβ2-GP
I/rhβ2-GP I complex in JEG-3 choriocarcinoma cells. A present,
β2-GP I has been considered to be one of the important clinical
indicators of pregnancy loss, but may be regarded as a signal of
choriocarcinoma malignance transformation in future.
In conclusion, the present study demonstrated that
the aβ2-GP I/rhβ2-GP I complex activates the expression of NF-κB by
activation of the MyD88 pathway, which promotes JEG-3 cell
proliferation, migration and invasion. These alterations lead to
enhancement of the degree of tumor malignancy and may represent the
mechanism underlying the disease caused by aPL.
Acknowledgements
The authors of the present study would like to thank
Dr Jiao Lv (Central Laboratory, the Teaching Hospital of Chengdu
University of Traditional Chinese Medicine, Chengdu, Sichuan
610072, P.R. China) for suggestions throughout the execution of
this project and English language editing.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL contributed to all aspects of this paper. XL
contributed in most of the experimental design and operations, as
well as manuscript writing and review. LR and WZ were responsible
for the recruitment of the women blood samples and purification of
IgG and aβ2-GP I from the blood samples, as well as manuscript
writing and review. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by The Second
Affiliated Hospital of Harbin Medical University Ethics Committee;
approval number: SCILLSC-2016-03. Participants provided consent to
participate in this research.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
References
|
1
|
Lee NS, Brewer HB Jr and Osborne JC Jr:
Beta 2-Glycoprotien I: Molecular properties of an unusual
apolipoprotein H. J Biol Chem. 258:4765–4770. 1983.PubMed/NCBI
|
|
2
|
Rand JH, Wu XX, Andree HA, Ross JB,
Rusinova E, Gascon-Lema MG, Calandri C and Harpel PC:
Antiphospholipid antibodies accelerate plasma coagulation by
inhibiting annexin-V binding to phospholipids: A ‘Lupus
Procoagulant’ phenomenon. Blood. 92:1652–1660. 1998.PubMed/NCBI
|
|
3
|
Pittoni V, Ravirajan CT, Donohoe S, Machin
SJ, Lydyard PM and Isenberg DA: Human monoclonal anti-phospholipid
antibodies selectively bind to membrane phospholipid and
beta2-glycoprotein I (beta2-GPI) on apoptotic cells. Clin Exp
Immunol. 119:533–543. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gao PJ, Shi Y, Gao YH, Liu YW and Tan Y:
The receptor for beta(2)GP I on membrane of hepatocellular
carcinoma cell line SMMC-7721 is annexin II. World J Gastroenterol.
13:3364–3368. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miyakis S, Lockshin MD, Atsumi T, Branch
DW, Brey RL, Cervera R, Derksen RH, DE Groot PG, Koike T, Meroni
PL, et al: International consensus statement on an update of the
classification criteria for definite antiphospholipid syndrome
(APS). J Thromb Haemost. 4:295–306. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pablo Demetrio R, Muñoz P, Hoyos López M,
Calvo V, Riancho L and Taboada Martinez VM: Thrombocytopenia as a
thrombotic risk factor in patients with antiphospholipid antibodies
without disease criteria. Med Clin (Barc). 148:394–400.
2017.PubMed/NCBI
|
|
7
|
Cris JC, Bouvier S, Nouvellon E,
Lissalde-Lavigne G, Mercier E, Balducchi JP and Marès P:
Antiphospholid antibodies and the risk of pregnancy complication.
Thromb Res. 151 Suppl 1:S34–S37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chighizola CB, Raimondo MG and Meroni PL:
Management of thrombotic antiphospholipid syndrome. Semin Thromb
Hemost. Mar 9–2017.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kandiah DA, Sheng YH and Krilis SA: beta
2-Glycoprotein I: Target antigen for autoantibodies in the
‘antiphospholipid syndrome. Lupus. 5:381–385. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dalekos GN, Zachou K and Liaskos C: The
antiphospholipid syndrome and infection. Curr Rheumatol Rep.
3:277–285. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Raschi E, Chighizola CB, Grossi C, Ronda
N, Gatti R, Meroni PL and Borghi MO: β2-glycoprotein I,
lipopolysaccharide and endothelial TLR4: Three players in the two
hit theory for anti-phospholipid-mediated thrombosis. J Autoimmun.
55:42–50. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Robertson SA, Roberts CT, van Beijering E,
Pensa K, Sheng Y, Shi T and Krilis SA: Effect of beta2-glycoprotein
I null mutation on reproductive outcome and antiphospholipid
antibody-mediated pregnancy pathology in mice. Mol Hum Reprod.
10:409–416. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Steinkasserer A, Cockburn DJ, Black DM,
Boyd Y, Solomon E and Sim RB: Assignment of apolipoprotein H (APOH:
beta-2-glycoprotein I) to human chromosome 17q23----qter;
determination of the major expression site. Cytogenet Cell Genet.
60:31–33. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hernández-Ramirez DF, Olivares-Martinez E,
Núñez-Álvarez CA, Chavelas EA, Garcia-Hernandez E, Gómez-Hernandez
G, Llorente L and Cabral AR: The role of β2-glycoprotein I (β2GPI)
carbohydrate chains in the reactivity of anti-β2GPI antibodies from
patients with primary antiphospholipid syndrome and in the
activation and differentiation of U937 cells. Biochem Biophys Res
Commun. 453:94–100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Muzio M, Polentarutti N, Bosisio D, Kumar
Manoj PP and Mantovani A: Toll-like receptor family and signalling
pathway. Biochem Soc Trans. 28:563–566. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gyrd-Hansen M and Meier P: IAPs: From
caspase inhibitors to modulators of NF-kappaB, inflammation and
cancer. Nat Rev Cancer. 10:561–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie H, Zhou H, Wang H, Chen D, Xia L, Wang
T and Yan J: Anti-β(2)GPI/β(2)GPI induced TF and TNF-α expression
in monocytes involving both TLR4/MyD88 and TLR4/TRIF signaling
pathways. Mol Immunol. 53:246–254. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
De Laat B, Derksen RH, van Lummel M,
Pennings MT and de Groot PG: Pathogenic anti-beta2-glycoprotein I
antibodies recognize domain I of beta2-glycoprotein I only after a
conformational change. Blood. 107:1916–1924. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lutters BC, Derksen RH, Tekelenburg WL,
Lenting PJ, Arnout J and de Groot PG: Dimers of beta 2-glycoprotein
I increase platelet deposition to collagen via interaction with
phospholipids and the apolipoprotein E receptor 2. J Biol Chem.
278:33831–33838. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
De Groot PG and Meijers JC:
β(2)-glycoprotein I: Evolution, structure and function. J Thromb
Haemost. 9:1275–1284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ailus K, Tulppala M, Palosuo T, Ylikorkala
O and Vaarala O: Antibodies to beta 2-glycoprotein I and
prothrombin in habitual abortion. Fertil Steril. 66:937–941. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vinatier D, Dufour P, Cosson M and Houpeau
JL: Antiphospholipid syndrome and recurrent miscarriages. Eur
Obstet Gynecol Reproduct Biol. 96:37–50. 2001. View Article : Google Scholar
|
|
24
|
Sada PR, Cohen H and Isenberg D: The
pathophysiology of antiphospholipid syndrome. Open Urol Nephrol J.
8:2–9. 2015. View Article : Google Scholar
|
|
25
|
Alijotas-Reig J, Esteve-Valverde E,
Ferrer-Oliveras R, Llurba E and Gris JM: Tumor necrosis
factor-alpha and pregnancy: Focus on biologics. An updated and
comprehensive review. Clin Rev Allergy Immunol. 53:40–53. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Weiss R, Bitton A, Nahary L, Arango MT,
Benhar I, Blank M, Shoenfeld Y and Chapman J: Cross-reactivity
between annexin A2 and Beta-2-glycoprotein I in animal models of
antiphospholipid syndrome. Immunol Res. 65:355–362. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yoshimura A: Signal transduction of
inflammatory cytokines and tumor development. Cancer Sci.
97:439–447. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ball E, Bulmer JN, Ayis S, Lyall F and
Robson SC: Late sporadic miscarriage is associated with
abnormalities in spiral artery transformation and trophoblast
invasion. J Pathol. 208:535–542. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ferretti C, Bruni L, Dangles-Marie V,
Pecking AP and Bellet D: Molecular circuits shared by placental and
cancer cells, and their implications in the proliferative, invasive
and migratory capacities of trophoblasts. Hum Reprod Update.
13:121–141. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aardema MW, Oosterhof H, Timmer A, van
Rooy I and Aarnoudse JG: Uterine artery Doppler flow and
uteroplacental vascular pathology in normal pregnancies and
pregnancies complicated by pre-eclampsia and small for gestational
age fetuses. Placenta. 22:405–411. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim YM, Chaiworapongsa T, Gomez R, Bujold
E, Yoon BH, Rotmensch S, Thaler HT and Romero R: Failure of
physiologic transformation of the spiral arteries in the placental
bed in preterm premature rupture of membranes. Am J Obstet Gynecol.
187:1137–1142. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim YM, Bujold E, Chaiworapongsa T, Gomez
R, Yoon BH, Thaler HT, Rotmensch S and Romero R: Failure of
physiologic transformation of the spiral arteries in patients with
preterm labor and intact membranes. Am J Obstet Gynecol.
189:1063–1069. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cui JQ, Shi YF, Zhou HJ and Li JQ: Changes
in gene expression profiles of hydatidiform mole and
choriocarcinoma as compared with trophoblast hyperplasia. Zhonghua
Zhong Liu Za Zhi. 26:727–731. 2004.(In Chinese). PubMed/NCBI
|