Introduction
Hodgkin's lymphoma (HL) is a common malignant tumor
among adolescents (1) that is
divided into classical and nodular lymphocyte predominant variants
based on the 2008 World Health Organization classification
(2). Using chemotherapy and
radiotherapy, alone or in combination, the durable remission rate
for HL is ~60–80% (3,4). However, survivors have an increased
risk of long-term sequelae, including cardiovascular disease and
secondary malignancy (5,6). Therefore, understanding the
underlying molecular mechanisms of HL development is essential for
the discovery of novel therapeutic targets for HL.
Previous studies have demonstrated that the
development and maintenance of various solid tumors and lymphoid
malignancies is driven by nuclear factor-κB (NF-κB) (7,8).
Constitutive NF-κB activation has been identified in classical HL
(9). A recent study by de Oliveira
et al (10) investigated
the functional roles and specific transcriptomes of NF-κB dimers
with distinct subunit compositions by knockdown of NF-kB subunit 2
(NFKB2) in a HL cell line; it was reported that NFKB2 knockdown
upregulated genes associated with hematopoietic and lymphoid organ
development. NFKB2 is a member of the transcription factor complex
NF-κB family, which promotes tumor cell proliferation and inhibits
apoptosis (11). Truncations in
the C-terminal region of NKFB2 are closely associated with
tumorigenesis in various hematopoietic tumors, including multiple
myeloma, chronic lymphocytic leukemia and cutaneous T-cell lymphoma
(12,13). A clear understanding of the
regulatory role of NFKB2 in HL may contribute to the development of
HL therapies.
de Oliveira et al (10) only analyzed the differentially
expressed genes (DEGs) associated with NFKB2 knockdown in a HL cell
line, and the regulatory mechanism of NFKB2 in HL remains poorly
understood. Therefore, in the present study, the regulatory
mechanism of NFKB2 in the tumorigenesis of HL was investigated by
downloading the gene expression data (GSE64234) deposited by de
Oliveira et al (10). DEGs
between the NFKB2 knockdown and control groups were screened for
prior to functional enrichment analysis of the identified DEGs,
followed by protein-protein interaction (PPI) network and module
analyses. Furthermore, microRNA (miRNA/miR) targets were predicted
for the identified DEGs and a miRNA-target regulatory network was
constructed.
Materials and methods
Affymetrix microarray data
The GSE64234 gene expression profile dataset
deposited by de Oliveira et al (10) was downloaded from the Gene
Expression Omnibus (https://www.ncbi.nlm.nih.gov/geo/) database, which was
based on the GPL6244 Affymetrix Human Gene 1.0 ST Array (Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Data on 12 HL cell line
specimens were available in GSE64234, including 6 treated with
non-targeting small interfering (si)RNA (control group) and 6 with
NFKB2-targeted siRNA (knockdown group).
Data preprocessing
Probe data in CEL files were transformed into
expression profiles. If numerous probes corresponded to the same
gene, the mean expression value of the probes was used as the gene
expression value. The oligo package (version 1.34.0) (14) in R (version 3.3.2) (15) was used to perform raw data format
conversion, missing value imputation and background correction
normalization for expression profile data. Quartiles were used for
data normalization.
DEG screening
The gene expression matrix derived from the
knockdown and control groups was analyzed to identify the DEGs.
P-values for genes were calculated with the non-paired t-test using
the limma package (version 3.10.3) (16) in R and subsequently adjusted using
the Benjamin and Hochberg method. An adjusted P-value of <0.05
was set as the threshold value. To directly assess DEG expression
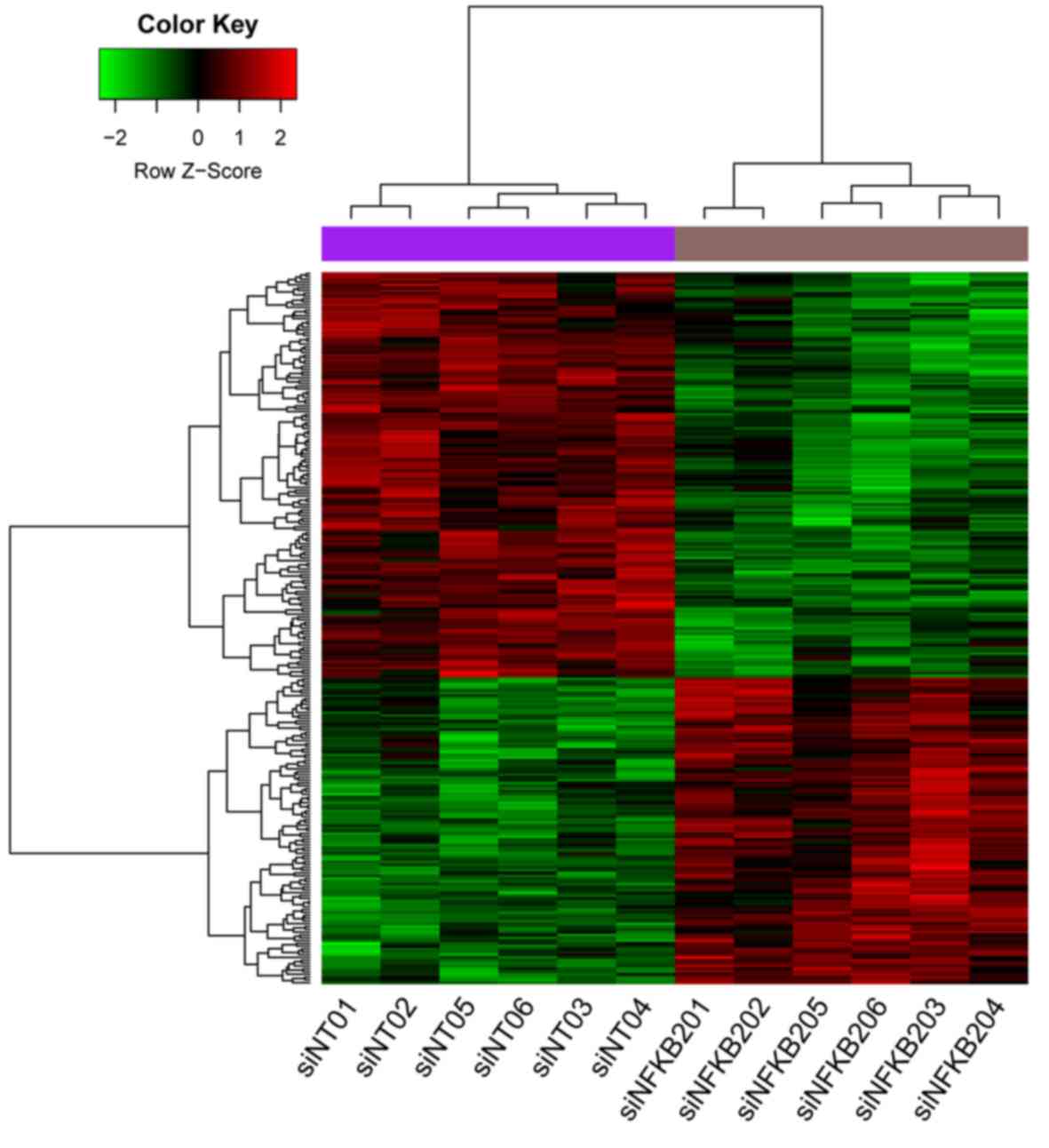
in these samples and confirm the classification, a DEG heat map was
drawn using the pheatmap package (version 1.0.8) (17) in R.
Functional enrichment analyses
Upregulated and downregulated DEGs were evaluated
based on Gene Ontology (GO) (18)
and Kyoto Encyclopedia of Genes Genomes (KEGG) (19) pathway enrichment analyses, using
the ‘gene set function-functional enrichment-mRNA enrichment’
module in the online Multifaceted Analysis Tool for Human
Transcriptome (www.biocloudservice.com). Functional enrichment
analyses were based on Fisher's exact test and P<0.05 was
selected as the threshold value.
PPI network and subnetwork module
analyses
PPI pairs were predicted by the Search Tool for the
Retrieval of Interacting Genes database (version 10.0; http://www.string-db.org/) (20) with a confidence score of >0.4.
Using the obtained PPI pairs, the PPI network was constructed with
Cytoscape version 3.2.0 (21). All
nodes in the PPI network were DEGs. In the PPI network, the node
score was calculated based on degree centrality. Nodes with a high
score, which likely have an important role in the PPI network, were
referred to as hub proteins. Proteins typically interact with other
proteins in a module. Therefore, following PPI network analysis,
significant modules from the PPI network were analyzed using the
MultiContrast Delayed Enhancement (MCODE) version 1.4.2 (22) plugin in Cytoscape (23).
miRNA-target regulatory network
analysis
miRNAs have a major role in the regulation of
biological processes in organisms (24). Therefore, miRNAs that may target
the identified DEGs were predicted using the ‘gene-miRNA
interaction information retrieval system’ in miRWalk 2.0 (25) (http://mirwalk.uni-hd.de/), with a minimum seed length
of seven and statistical significance set at P<0.05. miRWalk is
a comprehensive database that not only documents miRNA binding
sites within the complete sequence of a gene, but also combines
this information with a comparison of binding sites resulting from
12 existing miRNA-target prediction programs (25). In this study, the identified
miRNA-target gene pairs required prediction by miRWalk, which
included miRanda, miRDB 4.0, miRmap and TargetScan 6.2 databases.
Predicted miRNAs were reviewed using the ‘holistic view of
validated disease-miRNA interactions’ system in miRWalk 2.0 to
select miRNAs that were confirmed to be associated with HL. These
verified miRNA-target pairs were used to construct the regulatory
network in Cytoscape (21).
Results
DEG screening
A total of 253 DEGs were identified following data
preprocessing, which included 109 upregulated DEGs and 144
downregulated DEGs. The heat map clearly indicated the distinct
separation of the knockdown and control groups (Fig. 1).
Functional enrichment analyses
The upregulated DEGs were significantly enriched in
‘platelet activation’, ‘osteoclast differentiation’ and ‘long-term
depression’ KEGG pathways, while downregulated DEGs were
predominantly enriched in ‘intestinal immune network for IgA
production’, ‘HTLV-I infection’ and ‘NF-κB signaling pathway’ KEGG
pathways (Table I).
 | Table I.Kyoto Encyclopedia of Genes and
Genomes pathway enrichment analysis for upregulated and
downregulated DEGs. |
Table I.
Kyoto Encyclopedia of Genes and
Genomes pathway enrichment analysis for upregulated and
downregulated DEGs.
| A, Upregulated
DEGs |
|---|
|
|---|
| ID | Name | P-value | Genes |
|---|
| hsa04611 | Platelet
activation | 0.03878 | PLA2G4F, FCGR2A,
PRKG2, PRKG1 |
| hsa04380 | Osteoclast
differentiation | 0.03953 | TYK2, GAB2, FCGR2A,
MAPK10 |
| hsa04730 | Long-term
depression | 0.04684 | PLA2G4F, PRKG2,
PRKG1 |
|
| B, Downregulated
DEGs |
|
| ID | Name | P-value | Genes |
|
| hsa04940 | Type I diabetes
mellitus | 0.0003212 | CD80, HLA-DRB5,
IL12B, HLA-DPB1, LTA |
| hsa04672 | Intestinal immune
network for IgA production | 0.0004971 | CD80, HLA-DRB5,
TNFRSF17, AICDA, HLA-DPB1 |
| hsa05330 | Allograft
rejection | 0.003062 | CD80, HLA-DRB5,
IL12B, HLA-DPB1 |
| hsa05166 | HTLV-I
infection | 0.003913 | WNT5A, CSF2, TLN1,
HLA-DRB5, NFKB2, BCL2L1, HLA-DPB1, LTA |
| hsa04064 | NF-κB signaling
pathway | 0.004905 | NFKB2, BCL2L1,
BIRC2, PLAU, LTA |
| hsa04060 | Cytokine-cytokine
receptor interaction | 0.009454 | CSF2, CCR4,
TNFRSF17, TNFRSF8, TNFRSF14, IL12B, LTA |
| hsa05145 | Toxoplasmosis | 0.01411 | HLA-DRB5, BCL2L1,
IL12B, HLA-DPB1, BIRC2 |
| hsa05168 | Herpes simplex
infection | 0.01472 | NECTIN1, HLA-DRB5,
TNFRSF14, IL12B, HLA-DPB1, LTA |
| hsa04062 | Chemokine signaling
pathway | 0.0157 | CCL22, CCR4, PTK2B,
HCK, GRK2, PRKCD |
| hsa04514 | Cell adhesion
molecules (CAMs) | 0.02597 | ALCAM, CD80,
NECTIN1, HLA-DRB5, HLA-DPB1 |
| hsa05332 | Graft-versus-host
disease | 0.02806 | CD80, HLA-DRB5,
HLA-DPB1 |
| hsa05323 | Rheumatoid
arthritis | 0.03255 | CSF2, CD80,
HLA-DRB5, HLA-DPB1 |
| hsa05202 | Transcriptional
misregulation in cancer | 0.04407 | CSF2, REL, RARA,
BCL2L1, PLAU |
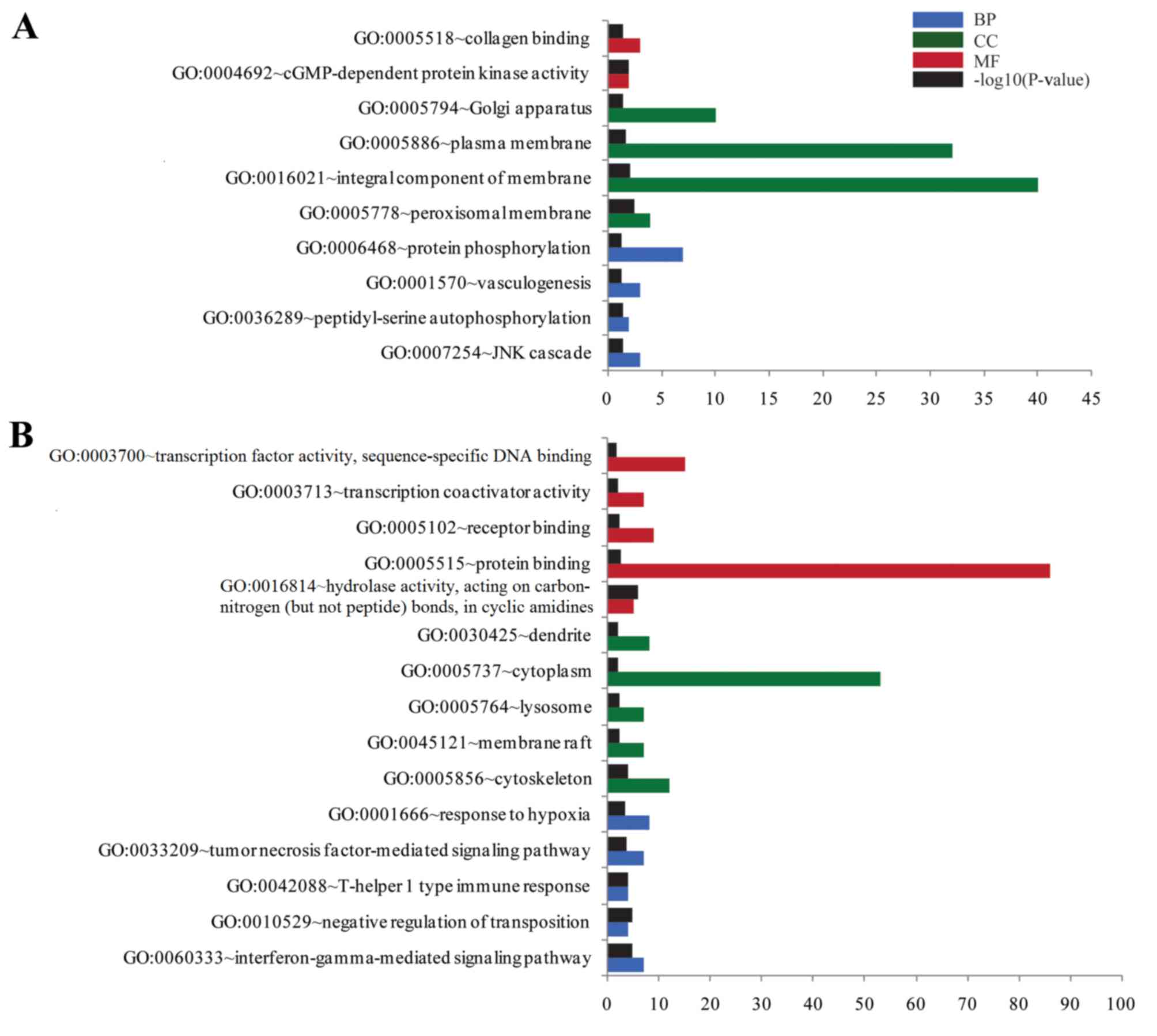
Additionally, upregulated DEGs were significantly
enriched in 10 GO terms, including ‘integral component of
membrane’, ‘plasma membrane’ and ‘Golgi apparatus’ terms (Fig. 2A). Downregulated DEGs were
significantly enriched in 53 GO terms, including ‘protein binding’,
‘cytoplasm’, ‘transcription factor activity’ and ‘sequence-specific
DNA binding’. The top 15 downregulated GO terms are presented in
Fig. 2B.
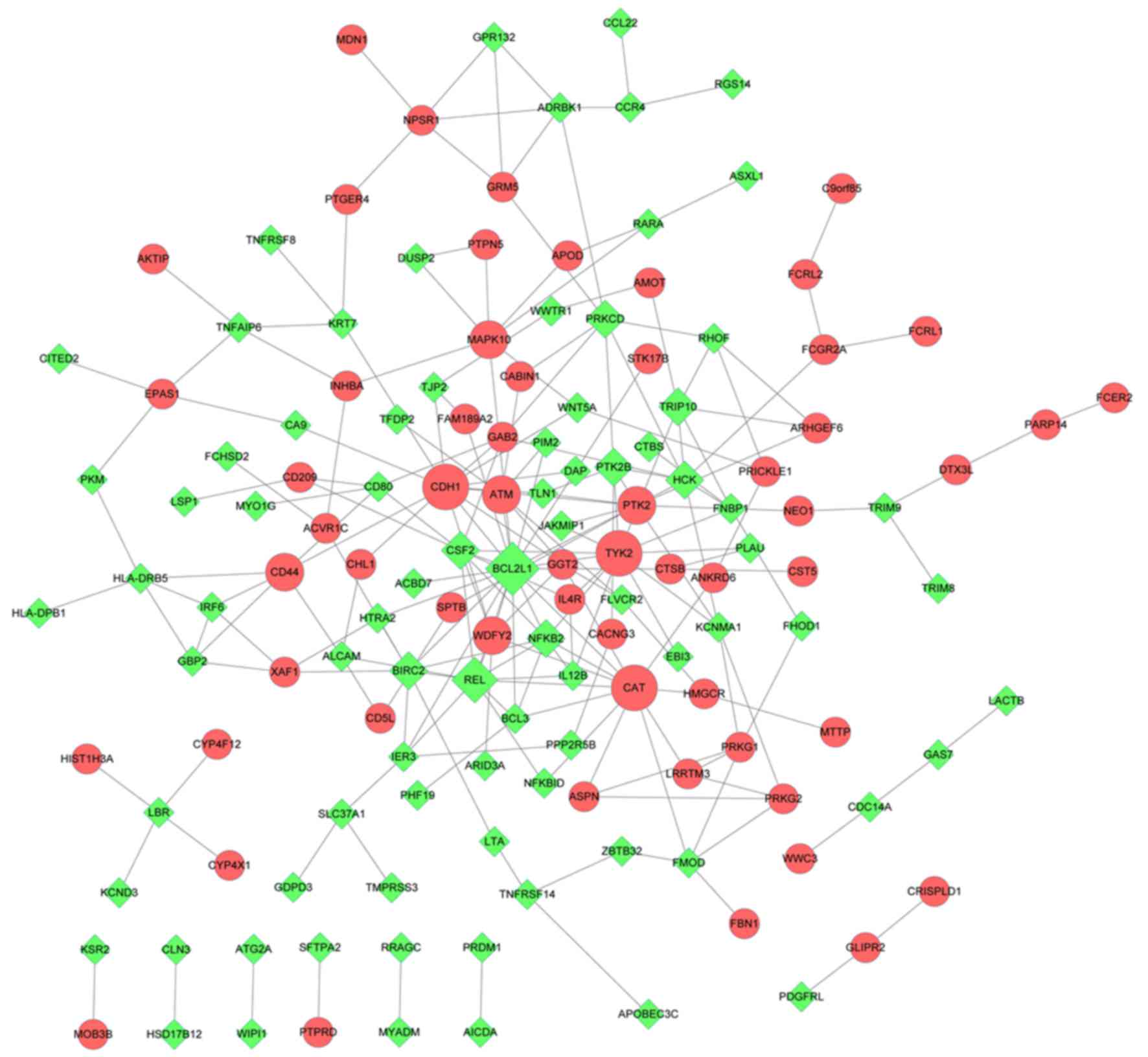
PPI network analysis
The PPI network included 142 nodes and 220 PPI pairs
(Fig. 3). Based on degree
centrality analysis, 8 upregulated and 12 downregulated DEGs were
determined to be hub genes (Table
II). KEGG pathway enrichment analysis was subsequently
performed to investigate the function of the 20 identified hub
genes. As presented in Table
III, these hub genes were significantly enriched in 14 KEGG
pathways. For instance, B-cell lymphoma 2-like 1 (BCL2L1) was
significantly enriched in ‘NF-κB signaling pathway’ and ‘Jak-STAT
signaling pathway’. Colony-stimulating factor 2 (CSF2) was
significantly enriched in ‘transcriptional misregulation in
cancer’, ‘TNF signaling pathway’ and ‘Jak-STAT signaling pathway’.
Tyrosine kinase 2 was significantly enriched in ‘Jak-STAT signaling
pathway’, ‘toxoplasmosis’, ‘Epstein-Barr virus infection’ and
‘osteoclast differentiation’. CD44 antigen was enriched in
‘Epstein-Barr virus infection’.
| Table II.Degree centrality analysis for 20 hub
genes. |
Table II.
Degree centrality analysis for 20 hub
genes.
| Node | Description | Degree |
|---|
| BCL2L1 | Downregulated | 19 |
| CDH1 | Upregulated | 13 |
| CAT | Upregulated | 12 |
| TYK2 | Upregulated | 12 |
| REL | Downregulated | 11 |
| PTK2 | Upregulated | 10 |
| CSF2 | Downregulated | 9 |
| HCK | Downregulated | 8 |
| BIRC2 | Downregulated | 8 |
| NFKB2 | Downregulated | 8 |
| WDFY2 | Upregulated | 7 |
| MAPK10 | Upregulated | 7 |
| PRKCD | Downregulated | 7 |
| ATM | Upregulated | 6 |
| TRIP10 | Downregulated | 6 |
| CD44 | Upregulated | 6 |
| PTK2B | Downregulated | 6 |
| FMOD | Downregulated | 5 |
| FNBP1 | Downregulated | 5 |
| IER3 | Downregulated | 5 |
| Table III.Kyoto Encyclopedia of Genes and
Genomes pathway enrichment analysis for the 20 hub genes. |
Table III.
Kyoto Encyclopedia of Genes and
Genomes pathway enrichment analysis for the 20 hub genes.
| ID | Name | P-value | Genes |
|---|
| hsa05202 | Transcriptional
misregulation in cancer | 0.0003731 | CSF2, PTK2, REL,
BCL2L1, ATM |
| hsa04064 | NF-κB signaling
pathway | 0.0007863 | NFKB2, BCL2L1,
BIRC2, ATM |
| hsa05200 | Pathways in
cancer | 0.001083 | PTK2, CDH1, NFKB2,
BCL2L1, MAPK10, BIRC2 |
| hsa05145 | Toxoplasmosis | 0.001901 | TYK2, BCL2L1,
MAPK10, BIRC2 |
| hsa04062 | Chemokine signaling
pathway | 0.006877 | PTK2, PTK2B, HCK,
PRKCD |
| hsa05169 | Epstein-Barr virus
infection | 0.007294 | TYK2, CD44, NFKB2,
MAPK10 |
| hsa04210 | Apoptosis | 0.007715 | BCL2L1, BIRC2,
ATM |
| hsa05222 | Small cell lung
cancer | 0.01415 | PTK2, BCL2L1,
BIRC2 |
| hsa04912 | GnRH signaling
pathway | 0.01611 | PTK2B, MAPK10,
PRKCD |
| hsa05166 | HTLV-I
infection | 0.01643 | CSF2, NFKB2,
BCL2L1, ATM |
| hsa04668 | TNF signaling
pathway | 0.02149 | CSF2, MAPK10,
BIRC2 |
| hsa04380 | Osteoclast
differentiation | 0.03187 | TYK2, NFKB2,
MAPK10 |
| hsa04068 | FoxO signaling
pathway | 0.03323 | CAT, MAPK10,
ATM |
| hsa04630 | Jak-STAT signaling
pathway | 0.03840 | TYK2, CSF2,
BCL2L1 |
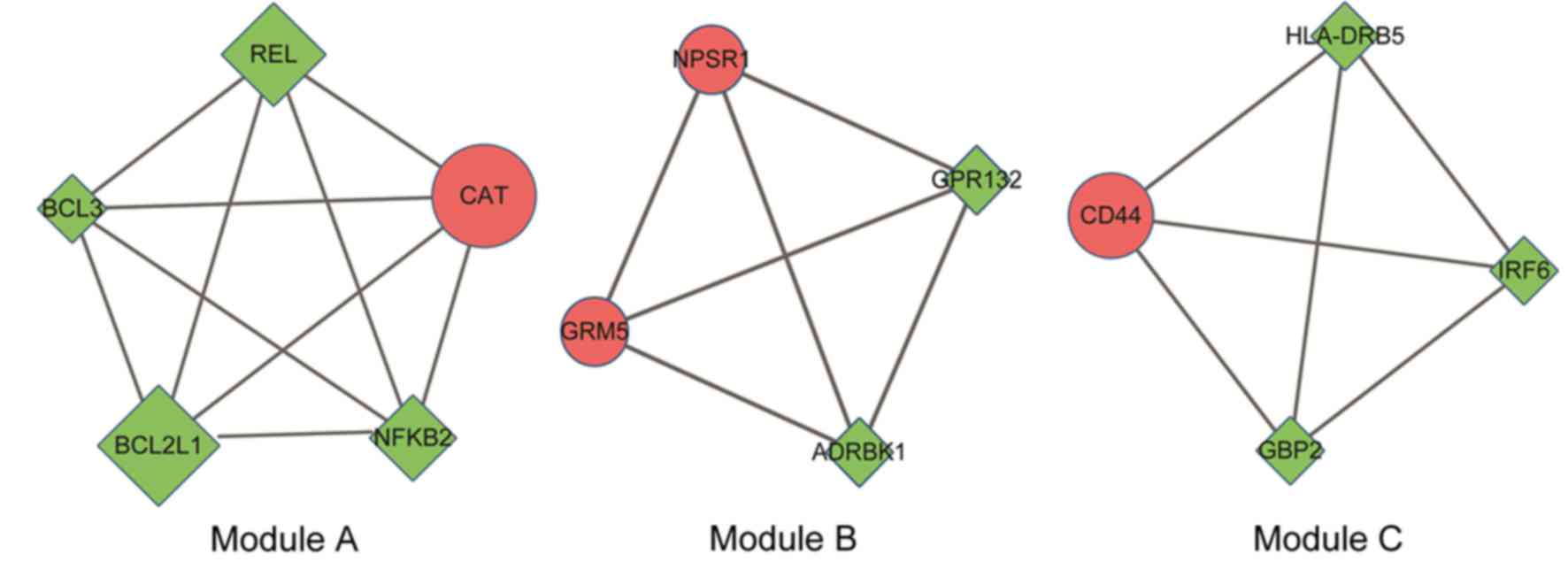
Subnetwork module analysis
Module analysis identified three modules (termed A,
B and C; Fig. 4). Module A was
comprised of 5 nodes, 10 interaction pairs and was enriched in
‘amyotrophic lateral sclerosis (ALS)’ and ‘NF-κB signaling pathway’
KEGG pathways. There were 4 nodes and 6 interaction pairs in module
B, without KEGG pathway enrichment. Module C contained 4 nodes and
6 interaction pairs and was enriched in the ‘hematopoietic cell
lineage’ KEGG pathway.
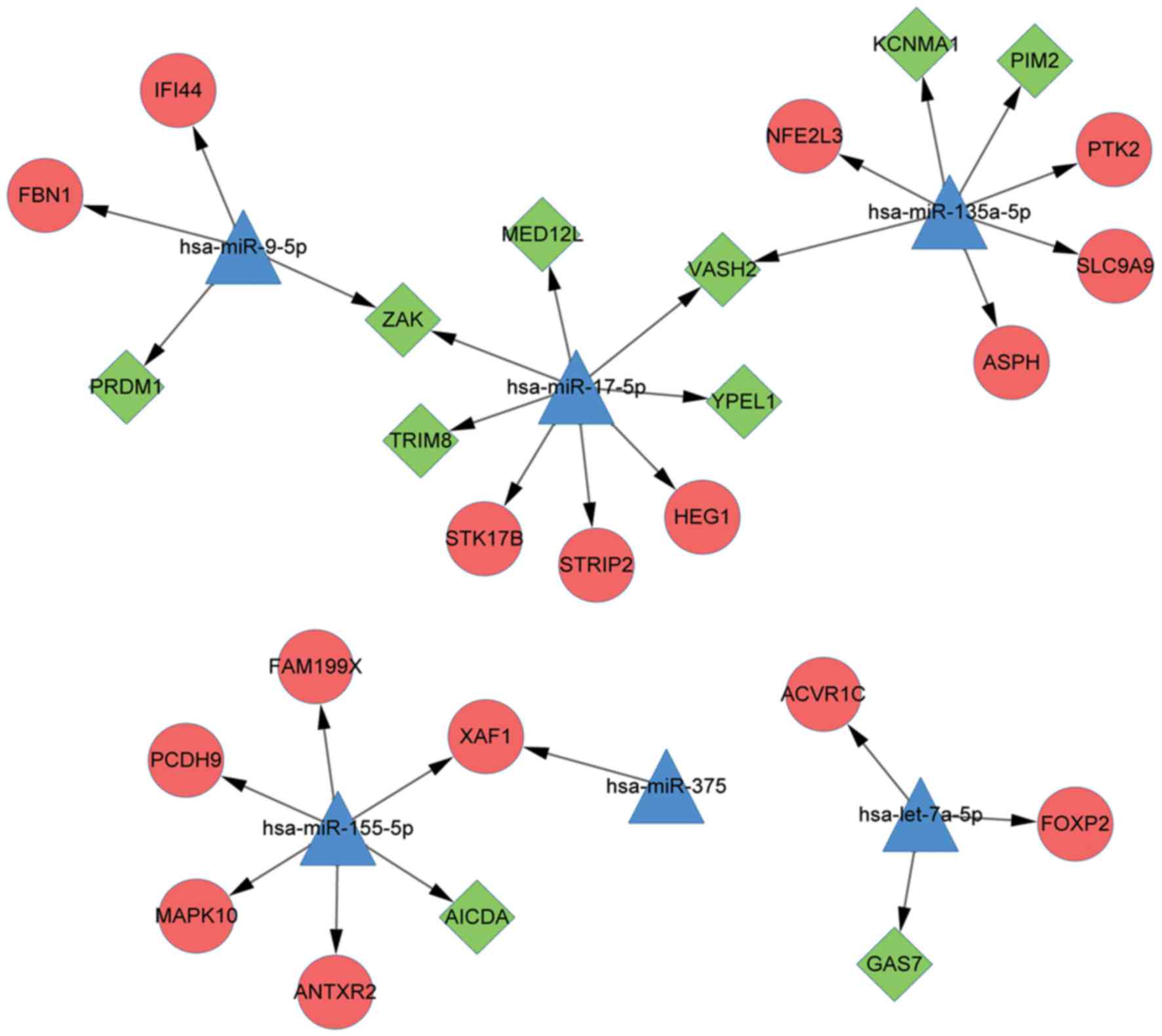
miRNA-target regulatory network
analysis
The miRNA-target regulatory network was constructed
with 32 nodes and 29 PPI pairs (Fig.
5). Among the 32 nodes, there were 16 upregulated DEGs, 10
downregulated DEGs and 6 miRNAs, which included let-7a-5p,
miR-9-5p, miR-155-5p, miR-135a-5p, miR-17-5p and miR-375.
Discussion
In the present study, a total of 109 upregulated and
144 downregulated DEGs were identified between the NFKB2 knockdown
and control groups. Pathway enrichment analysis revealed that
BCL2L1 was significantly enriched in the NF-κB signaling pathway,
and CSF2 and BCL2L1 were enriched in the Jak-STAT signaling
pathway. Additionally, BCL2L1 and CSF2 were hub genes in the PPI
network. Prediction of miRNA-target interactions identified 29
miRNA-target pairs involving in 6 miRNAs (let-7a-5p, miR-9-5p,
miR-155-5p, miR-135a-5p, miR-17-5p and miR-375). The findings of
the present study may provide guidelines for understanding the
regulatory mechanism of NFKB2 and contribute to the identification
of therapeutic targets for HL.
BCL2L1 was demonstrated to be downregulated in HL
cell lines following NFKB2 knockdown. This gene is a member of the
Bcl-2 family and is a vital apoptosis-mediating gene that encodes
both an antiapoptotic and proapoptotic splice variant (26). Additionally, in the present study,
module analysis revealed that BCL2L1 interacts with NFKB2 in module
A, which indicated that knockdown of NFKB2 may suppress BCL2L1
expression and subsequently regulate HL cell apoptosis. Notably,
this result was consistent with the findings of de Oliveira et
al (10), who reported that
NFKB2 knockdown may downregulate BCL2L1 expression. Furthermore,
BCL2L1 was enriched in the NF-κB signaling pathway in the present
study. The NF-κB signaling pathway has been associated with the
progression of several tumors, including HL (7,11).
Therefore, it was speculated that BCL2L1 may be involved in
NF-κB-dependent control of HL cell survival.
In addition to BCL2L1, CSF2 was also downregulated
in the NFKB2 knockdown group. CSF2 encodes a cytokine that has been
reported to be involved in the pathogenesis of HL (27) and is highly expressed in HL cells
(28). In accordance with these
findings, the present study demonstrated that CSF2 was upregulated
in HL control cells compared with HL cells with NFKB2 knockdown,
indicating that NFKB2 may be involved in HL tumorigenesis through
the upregulation of CSF2. KEGG pathway enrichment analysis revealed
that CSF2 was enriched in the Jak-STAT signaling pathway. The
Jak-STAT signaling pathway is regulated by a large array of
cytokines and growth factors that induce proliferation or
differentiation (29). Activation
of the Jak-STAT signaling pathway is a hallmark of various B-cell
lymphomas, including classical HL (30). Taken together, these data indicate
that NFKB2 may be implicated HL development by regulating CSF2
expression, which in turn activates the Jak-STAT signaling
pathway.
miRNAs regulate gene expression and cellular
processes (24). They are also
potential diagnostic and prognostic molecular markers in cancer, as
well as targets for the development of highly specific therapies
(31). Based on the DEGs
identified in the present study, several miRNAs were predicted.
Among these, miR-135a-5p has been previously reported to have
critical involvement in HL cell survival by targeting Jak2
(32). Additionally, miR-155-5p
has been recommended as a diagnostic marker in hematological tumors
(33). Notably, dysregulated
expression of miR-155 has been reported in various types of
lymphoma (33). Differential
expression of miR-9-5p has been demonstrated in hematological cell
lines compared with normal lymphocyte populations (34). Furthermore, a target gene of
miR-9-5p, PR/SET domain 1, was reported to be downregulated in HL
cells (35), further indicating a
potential role for miR-9-5p in HL. Therefore, NFKB2 may be
implicated in HL development through interaction with these
miRNAs.
In conclusion, the present study demonstrated that
NFKB2 may be involved in the development of HL by interacting with
several genes and miRNAs, including BCL2L1, CSF2, miR-135a-5p,
miR-155-5p and miR-9-5p. Additionally, NFKB2-mediated modulation of
the Jak-STAT and NF-κB signaling pathways may have a role in HL
tumorigenesis. The genes, miRNAs and signaling pathways identified
in the present study may serve as promising therapeutic targets for
HL. Future experimental verification in vivo is required to
confirm the findings of the present study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and LW conceptualized the study design. LT and SC
acquired the data. LT, SC and WW performed the statistical
analysis. YZ drafted the manuscript. LW revised the manuscript. All
author read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wethal T, Lund MB, Edvardsen T, Fosså SD,
Pripp AH, Holte H, Kjekshus J and Fosså A: Valvular dysfunction and
left ventricular changes in Hodgkin's lymphoma survivors. A
longitudinal study. Br J Cancer. 101:575–581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Swerdlow SH, Campo E, Harris NL, Jaffe ES,
Pileri SA, Stein H, Thiele J and Vardiman JW: WHO classification of
tumours of haematopoietic and lymphoid tissues. 4th edition. IARC
Press; Lyon: 2008
|
|
3
|
Connors JM: State-of-the-art therapeutics:
Hodgkin's lymphoma. J Clin Oncol. 23:6400–6408. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Diehl V, Franklin J, Pfreundschuh M,
Lathan B, Paulus U, Hasenclever D, Tesch H, Herrmann R, Dörken B,
Müller-Hermelink HK, et al: Standard and increased-dose BEACOPP
chemotherapy compared with COPP-ABVD for advanced Hodgkin's
disease. N Engl J Med. 348:2386–2395. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Steidl C, Lee T, Shah SP, Farinha P, Han
G, Nayar T, Delaney A, Jones SJ, Iqbal J, Weisenburger DD, et al:
Tumor-associated macrophages and survival in classic Hodgkin's
lymphoma. N Engl J Med. 362:875–885. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Heidenreich PA, Schnittger I, Strauss HW,
Vagelos RH, Lee BK, Mariscal CS, Tate DJ, Horning SJ, Hoppe RT and
Hancock SL: Screening for coronary artery disease after mediastinal
irradiation for Hodgkin's disease. J Clin Oncol. 25:43–49. 2006.
View Article : Google Scholar
|
|
7
|
Lim KH, Yang Y and Staudt LM: Pathogenetic
importance and therapeutic implications of NF-κB in lymphoid
malignancies. Immunol Rev. 246:359–378. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Didonato JA, Mercurio F and Karin M: NF-κB
and the link between inflammation and cancer. Immunol Rev.
246:379–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jost PJ and Ruland J: Aberrant NF-kappaB
signaling in lymphoma: Mechanisms, consequences, and therapeutic
implications. Blood. 109:2700–2707. 2007.PubMed/NCBI
|
|
10
|
de Oliveira KA, Kaergel E, Heinig M,
Fontaine JF, Patone G, Muro EM, Mathas S, Hummel M, Andrade-Navarro
MA, Hübner N and Scheidereit C: A roadmap of constitutive NF-κB
activity in Hodgkin lymphoma: Dominant roles of p50 and p52
revealed by genome-wide analyses. Genome Med. 8:282016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Khan G: Epstein-Barr virus, cytokines, and
inflammation: A cocktail for the pathogenesis of Hodgkin's
lymphoma? Exp Hematol. 34:399–406. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Neri A, Fracchiolla NS, Migliazza A,
Trecca D and Lombardi L: The involvement of the candidate
proto-oncogene NFKB2/lyt-10 in lymphoid malignancies. Leuk
Lymphoma. 23:43–48. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Neri A, Chang CC, Lombardi L, Salina M,
Corradini P, Maiolo AT, Chaganti RS and Dalla-Favera R: B cell
lymphoma-associated chromosomal translocation involves candidate
oncogene lyt-10, homologous to NF-kappa B p50. Cell. 67:1075–1087.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ihaka R and Gentleman R: R: A language for
data analysis and graphics. J Comput Graphical Stat. 5:299–314.
1996. View Article : Google Scholar
|
|
16
|
Smyth GK: Limma: Linear models for
microarray data. Bioinformatics Comput Biol Solut Using R
Bioconductor. 397–420. 2005. View Article : Google Scholar
|
|
17
|
Wang L, Cao C, Ma Q, Zeng Q, Wang H, Cheng
Z, Zhu G, Qi J, Ma H, Nian H and Wang Y: RNA-seq analyses of
multiple meristems of soybean: Novel and alternative transcripts,
evolutionary and functional implications. BMC Plant Biol.
14:1692014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
von Mering C, Huynen M, Jaeggi D, Schmidt
S, Bork P and Snel B: STRING: A database of predicted functional
associations between proteins. Nucleic Acids Res. 31:258–261. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bandettini WP, Kellman P, Mancini C,
Booker OJ, Vasu S, Leung SW, Wilson JR, Shanbhag SM, Chen MY and
Arai AE: MultiContrast Delayed Enhancement (MCODE) improves
detection of subendocardial myocardial infarction by late
gadolinium enhancement cardiovascular magnetic resonance: A
clinical validation study. J Cardiovasc Magn Reson. 14:832012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hobert O: Gene regulation by transcription
factors and microRNAs. Science. 319:1785–1786. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dweep H and Gretz N: miRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sillars-Hardebol AH, Carvalho B, Beliën
JA, de Wit M, Delis-van Diemen PM, Tijssen M, van de Wiel MA,
Pontén F, Fijneman RJ and Meijer GA: BCL2L1 has a functional role
in colorectal cancer and its protein expression is associated with
chromosome 20q gain. J Pathol. 226:442–450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Steidl C, Connors JM and Gascoyne RD:
Molecular pathogenesis of Hodgkin's lymphoma: Increasing evidence
of the importance of the microenvironment. J Clin Oncol.
29:1812–1826. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brase JC, Johannes M, Schlomm T, Fälth M,
Haese A, Steuber T, Beissbarth T, Kuner R and Sültmann H:
Circulating miRNAs are correlated with tumor progression in
prostate cancer. Int J Cancer. 128:608–616. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Harrison DA, Mccoon PE, Binari R, Gilman M
and Perrimon N: Drosophila unpaired encodes a secreted protein that
activates the JAK signaling pathway. Genes Dev. 12:3252–3263. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zahn M, Marienfeld R, Melzner I, Heinrich
J, Renner B, Wegener S, Mießner A, Barth TF, Dorsch K, Brüderlein S
and Möller P: A novel PTPN1 splice variant upregulates JAK/STAT
activity in classical Hodgkin lymphoma cells. Blood. 129:1480–1490.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Navarro A, Diaz T, Martinez A, Gaya A,
Pons A, Gel B, Codony C, Ferrer G, Martinez C, Montserrat E and
Monzo M: Regulation of JAK2 by miR-135a: Prognostic impact in
classic Hodgkin lymphoma. Blood. 114:2945–2951. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jurkovicova D, Magyerkova M, Kulcsar L,
Krivjanska M, Krivjansky V, Gibadulinova A, Oveckova I and Chovanec
M: miR-155 as a diagnostic and prognostic marker in hematological
and solid malignancies. Neoplasma. 61:241–251. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lawrie CH, Saunders NJ, Soneji S, Palazzo
S, Dunlop HM, Cooper CD, Brown PJ, Troussard X, Mossafa H, Enver T,
et al: MicroRNA expression in lymphocyte development and
malignancy. Leukemia. 22:1440–1446. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nie K, Gomez M, Landgraf P, Garcia JF, Liu
Y, Tan LH, Chadburn A, Tuschl T, Knowles DM and Tam W:
MicroRNA-mediated down-regulation of PRDM1/Blimp-1 in
Hodgkin/Reed-Sternberg cells: A potential pathogenetic lesion in
Hodgkin lymphomas. Am J Pathol. 173:242–252. 2008. View Article : Google Scholar : PubMed/NCBI
|