Introduction
Shigella is a genus of pathogens responsible
for acute bacterial diarrhea, chronic bacteritic dysentery and
toxic dysentery. Certain Shigella strains can cause
hemolytic uremic syndrome and Reiter's chronic arthritis syndrome,
and with the extensive use of antibacterial agents, bacterial
drug-resistance is becoming a serious concern. Mobile genetic
elements, including plasmids, transposons and integrons are
important in the spread of antibiotic resistance (1). Among these elements, class 1, 2 and 3
integrons are associated with drug resistance. A previous study
showed that the involvement of the active efflux pump is
significant in bacterial multi-drug resistance (2). Previous studies have mainly focused
on the active efflux pump of Escherichia coli, AcrAB-TolC,
which is also expressed in Shigella striains (3). The regulation of gene mutation may be
important in the multi-drug resistance mediated by the gene
(4). In the present study, the
characteristics of integron genes were analyzed in clinical
Shigella isolates from children, mutations of the active
effiux pump gene (acrAB-tolC) and regulatory genes were
examined, and the association between these genes and the
drug-resistance of Shigella was investigated. This may
provide evidence for the clinical treatment of dysentery and the
control of multi-drug resistance of Shigella.
Materials and methods
Location of isolated strains
A total of 158 isolates of Shigella flexneri were
isolated from the stool samples of 94 male cases and 64 female
cases aged from 6 months to 14 years old between May 2012 and
October 2015, in which there were 106 cases aged less than 5 years
old. The isolates were identified using ID32E identificational
strips and an ATB expression instrument, and were serotyped using
diagnostic-serum of Shigella. Escherichia coli (cat. no. 25922;
American Type Culture Collection) was preserved in the Clinical
Microbiology Laboratory of The Fourth Hospital of Jinan City
(Jinan, China) and it was the quality control strain for
antimicrobial susceptibility assessment. The present study was
approved by the Ethics Committee of Jinan Infectious Disease
Hospital Affiliated To Shandong University (Shandong, China).
Written informed consent was obtained from all patients or patient
guardians for the present study.
Reagents and equipment
The following were obtained for use in the present
study: Shigella diagnostic serum (Lanzhou Institute of
Biological Products Co. Ltd., Lanzhou, China); antibotic discs of
ampicillin, chloramphenicol, tetracycline, streptomycin,
trimethoprim, sulfamethoxazole, ciprofloxacin, cefotaxime,
ceftazidime, cefoperazone/sulbactam and imipenem, and M-H agar
(Oxoid; Thermo Fisher Scientific, Inc., Waltham, MA, USA); 100 bp
DNA Ladder marker and D15000+2000 DNA marker (Tiangen Biotech Co.,
Ltd., Beijing, China); Takara Taq kit, DL2000TMDNA marker
and restriction enzyme (Takara Biotechnology Co., Ltd., Dalian,
China), agarose (Invitrogen; Thermo Fisher Scientific, Inc.);
bacterial identification instrument (ATB Expression; BioMérieux,
Marcy l'Etoile, France); DNA amplifier (Biometra GmbH, Göttingen,
Germany); electrophoresis apparatus (10C type; Beijing Liuyi
Biotechnology Co., Ltd., Beijing, China); biosafety cabinets
(1200IIA2; Shanghai Lishen Biotechnology Co., Ltd.). Casein
hydrolysate acid (M-H) agar (Shanghai Lishen Biological Technology
Co., Ltd.).
Strain identification and
antimicrobial susceptibility assessment
The preserved strains were removed from the −86°C
ultra-low temperature freezer, thawed at room temprature and
re-identified. Subsequently, the strains underwent an agglutination
test with Shigella diagnostic serum for the serotype
identification. In total, 3–4 bacterial colonies of S.
flexneri were detected following culture of the stains for
16–18 h at 35°C, followed by adjustment of the turbidity instrument
to 0.5 McF with normal saline. Sterile cotton swabs were dipped
into the bacteria solution and excess liquid was squeezed out
against the tube wall, followed by application of the bacteria onto
the M-H plate. The bacteria were applied ontp the M-H plate three
times with a 60° rotation; the bacteria were also applied along the
edge for entire 360° rotation. Following 5 min, the antimicrobial
susceptibility disks were placed on the colonies and cultured for
16–18 h at 35°C. Following these procedures, the inhibition zone
diameter was detected with vernier calipers and the data were input
into WHONET 5.4 software (http://www.whonet.org/). According to the 2010 version
of Performance Standards for Antimicrobial Susceptibility Testing
of the National Committee for Clinical Standards Laboratory
Institute) (5), the quality
control strains was Escherichia coli (cat. no. 25922).
Assessment and sequencing for
resistance gene cassettes of integrons and variable regions
The integron and plasmid DNA templates were prepared
using a boiling method for 5 min, and they were preserved in a
−20°C refrigerator. The primers and primer sequences were
synthesized in the accordance with relative reports (6–9), as
in Table I. The class 1, 2 and 3
integrons, integrase primers and variable region primers were
synthesized by Takara Biotechnology Co., Ltd. The reaction system
was as follows: 10X buffer containing Mg2+ (5 µl), dNTP
(4 µl; 2.5 mmol/l), downstream and upstream primers (2.5 µl; 10
µmol/l of each), Taq eznzyme (0.25 µl; 5 U/µl) and DNA templates (5
µl). Deionized water was added to the reaction system to 50 µl. The
amplification parameters for integrase were as follows:
Pre-denaturation at 94°C for 5 min, followed by 94°C for 30 sec,
annealing for 30 sec (annealing temperature shown in Table I) and 72°C for 40 sec, for 35
cycles, with final extension at 72°C for 5 min. The amplification
parameters for variable regions were as follows: Pre-denaturation
at 94°C for 5 min; touchdown polyermase chain reaction (PCR) for 10
cycles, annealing temperature reduction from 60 to 50°C
(temperature decrease by 1.4°C each cycle), followed by 94°C for 30
sec, 52°C for 30 sec and 72°C for 180 sec, for 25 cycles with final
extension at 72°C for 8 min. For product analysis, the
amplification product (10 µl) was used for 1% agarose
electrophoresis under 120 V for 20 min. Following EB staining, the
products were observed under the UV absorption spectra analyzer.
Images were captured and the results recorded. The amplification
products were sent to Takara Biotechnology Co., Ltd. for gene
sequencing and the sequences were compared with NCBI/BLAST
(https://blast.ncbi.nlm.nih.gov/Blast.cgi).
 | Table I.Primers of genes associated with
class 1 integron. |
Table I.
Primers of genes associated with
class 1 integron.
| Primer | Sequence
(5′-3′) | Primer sequence
site |
|---|
| intI1 F |
ACATGTGATGGCGACGCACGA | intI1 |
| intI1 R |
ATTTCTGTCCTGGCTGGCGA | intI1 |
| In F |
GGCATCCAAGCAGCAAGC | 5′-conserved
segment in the class 1 integrons |
| In R |
AAGCAGACTTGACCTGAT | 3′-conserved
segment in the class 1 integrons |
| qacEΔ1 |
ATCGCAATAGTTGGCGAAGT | qacEΔ1 |
| sul1 |
GCAAGGCGGAAACCCGCGCC | sul1 |
| intI1ca |
CGTAGAAGAACAGCAAGG | intI1 |
| IS1ca |
AGTGAGAGCAGAGATAGC | IS1 |
| intI2 F |
GTAGCAAACGAGTGACGAAATG | intI2 |
| intI2 R |
CACGGATATGCGACAAAAAGGT | intI2 |
| intI2ca F |
CGGGATCCCGGACGGCATGCACGATTTGTA | intI2 |
| intI2ca R |
GATGCCATCGCAAGTACGAG | 3′-conserved
segment in the class 2 integrons |
| intI3 F |
GCCTCCGGCAGCGACTTTCAG | intI3 |
| intI3 R |
ACGGATCTGCCAAACCTGACT | intI3 |
BamHI restriction enzyme
digestion
Enzyme digestion was performed in the variable
regions of the atypical class 1,2 integrons with the restriction
enzyme HindIII and Hinf1. The specific steps were as
follows: 10 µl enzyme buffer, 40 µl PCR products and 5 µl
restriction enzyme were added to deionized water to 100 µl. The
solution was placed at 37°C for the isothermal reaction for ~1 h.
Subsequently, 10X loading buffer was added to terminate the
reaction. The 10-µl samples were used for 2.5% agarose
electrophoresis under 120 V for 20 min. Following EB staining, the
products were observed under the UV absorption spectra analyzer.
Images were captured and the results recorded.
Analysis of acrAB-tolC and its
associated regulatory genes
The multi-drug resistant Shigella strains
(resistant to five or more antibiotics) and sensitive strains were
used for assessment. The PCR amplication and sequencing methods
were applied. The specific steps were in accordance with the above
methods. According to previous reports (10,11),
the experimental primers were synthesized by Takara Biotechnology
Co., Ltd. (Table II). The
sequences were compared with NCBI/BLAST to investigate the numbers
of mutations and mutational sites.
| Table II.Active efflux pump acrAB-tolC
and its regulatory genes. |
Table II.
Active efflux pump acrAB-tolC
and its regulatory genes.
| Primer | Sequence
(5′-3′) | Length (bp) | Annealing
temperature (°C) |
|---|
| gyrA |
TACACCGGTCAACATTGACG | 648 | 50 |
|
|
TTAATGTTGCCGCCGTCGG |
|
|
| parC |
GCGTTGCCGTTTATTGGTGAT | 469 | 52 |
|
|
TGGACATCGTCATACCTCT |
|
|
| acrA |
TGCGGCTTGCTGGTTATT | 1,131 | 52 |
|
|
GCGGTCGTTCTGATGCTC |
|
|
| acrB |
GATTCCGACCATTGCCGTAC | 510 | 52 |
|
|
GCCAGAATACCGCCTACGC |
|
|
| marOR |
CACTCTTTAGCTAGCCTTG | 604 | 51 |
|
|
TGGACATCGTCATACCTCT |
|
|
| acrR |
AAACCCATTGCTGCGTTTAT | 800 | 55 |
|
|
AAACCGCAAGAATATCACGA |
|
|
Results
Results of drug sensitivity tests
A total of 91.1% of the 158 strains were
multi-resistant to ampicillin, chloramphenicol, tetracycline and
streptomycin, and 69.6% of the strains were multi-resistant to
sulfamethoxazole/trimethoprim. The resistance rate to ceftazidime,
ciprofloxacin and levofloxacin was <32.9%. All (100%) of the
strains were sensitive to cefoxitin, cefoperazone/sulbactam and
imipenem. The frequent resistant pattern
(ampicillin-tetracycline-chloromycetin-streptomycin) accounted for
91.1% of strains (144/158), and 77.2% (122/158) of the strains were
resistant to more than five antibiotics. Multi-drug resistance was
a serious problem. Only 14 strains were sensitive strains, which
were not drug-resistant (Table
III).
| Table III.Results of class 1 and class 2
integron detection in 79 strains of Shigella flexneri. |
Table III.
Results of class 1 and class 2
integron detection in 79 strains of Shigella flexneri.
| Serotype | Typical class 1
integron (intI1, in, qacEΔ1-sul1-positive) | Atypical class 1
integron (intI1, intI1-IS1-positive) | Class 2 integron
(intI2, intI2ca-positive) |
|---|
| S.
flexneri 1a | 0 | 16 | 14 |
| S.
flexneri 2a | 20 | 70 | 74 |
| S.
flexneri 2b | 2 | 10 | 10 |
| S.
flexneri 4a | 4 | 34 | 34 |
| S.
flexneri × variant | 0 | 4 | 4 |
| Total | 26 | 134 | 136 |
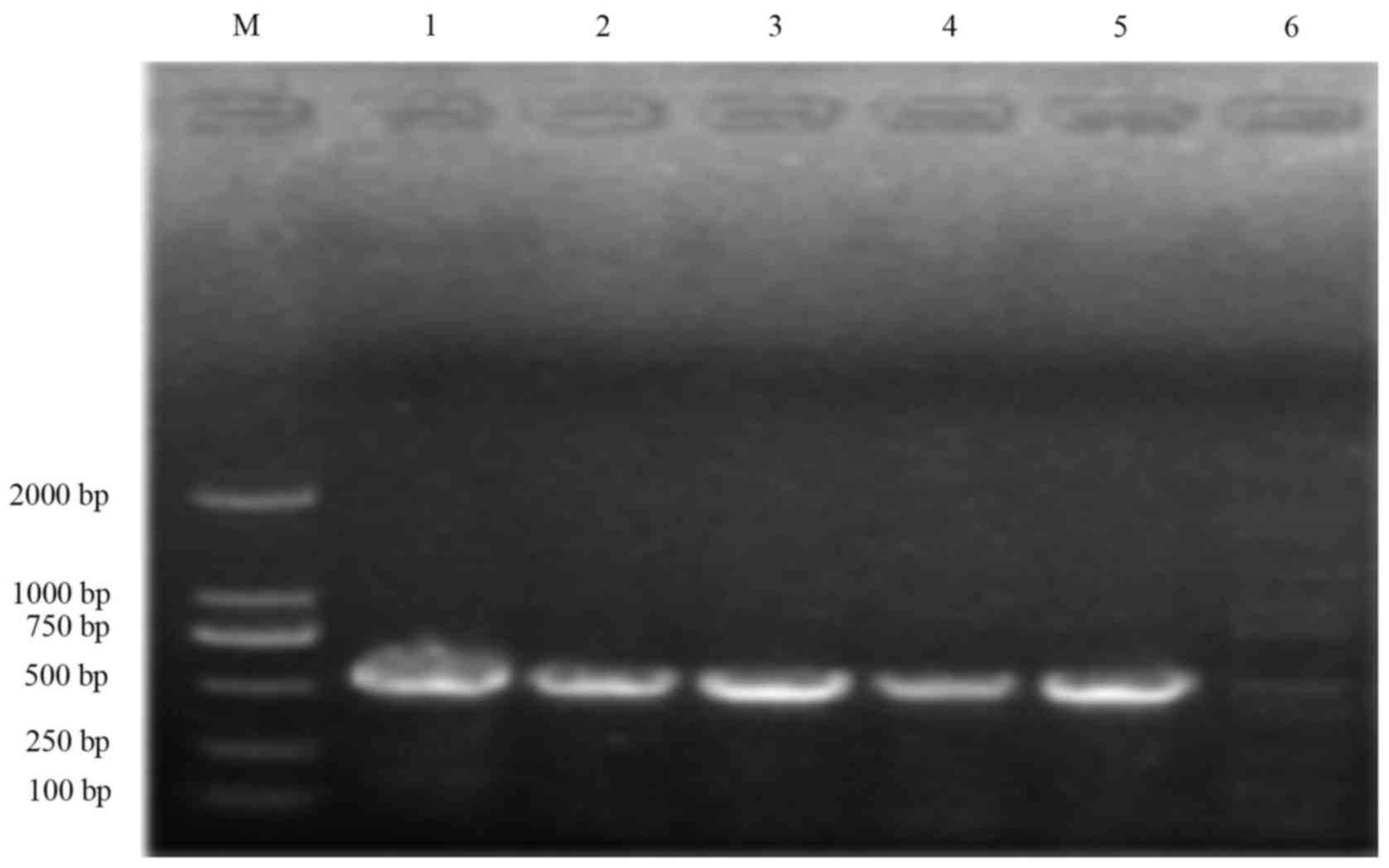
Integration assessment results
Among the 158 S. flexneri strains, 144
strains were intI1-positive and 136 strains were intI
2-positive. The positive rates were 91.1% (144/158) and 86.1%
(136/158), respectively (Figs. 1
and 2). All strains were
intI3-positive. In total, 10 strains were single
ntI1-positive, accounting for 6.3% (10/158), and 134 strains
were intI1-intI2 positive, accounting for 84.8% (134/158).
Among the intI1-positive strains, the 3′-conserved segment
of 16.5% of the strains (26/156) were positive. They were typical
class 1 integrons [intI1, (conservative primer of class 1
integrons) in and qacEΔ1-sul1-positive; Fig. 3]. The 3′-conserved segment of 84.8%
(134/158) of the strains were negative. They were atypical class 1
integrons (intI1 and intI1-IS1-positive; Fig. 4. In addition, 10.1% (16/158)
strains (S. flexneri 2A) possessed typical and atypical
class 1 integron class 1 integrons. The integron assessment results
are shown in Table III.
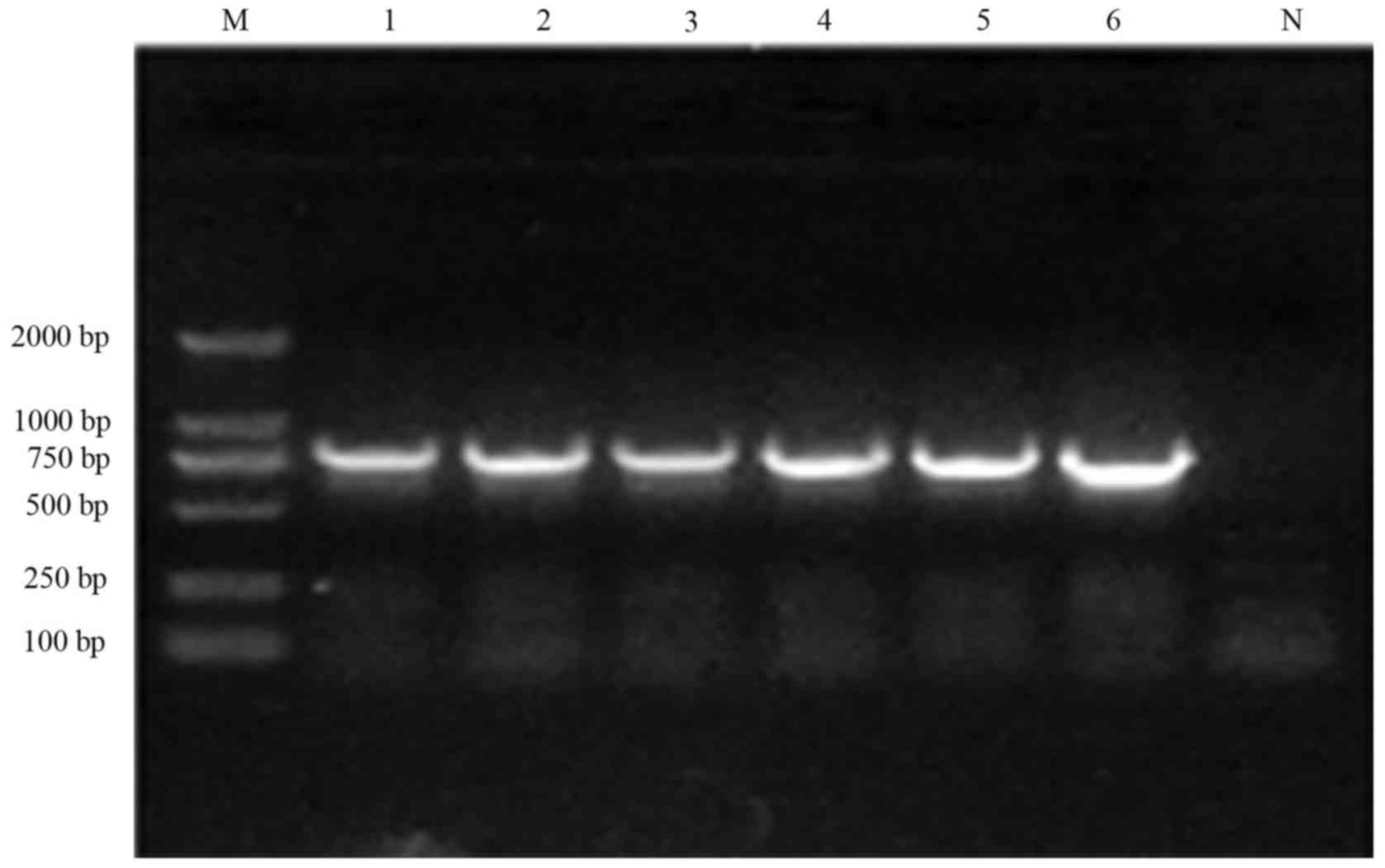
Assessment and sequencing for
resistance gene cassettes of integrons and variable regions
Among the 26 strains with typical class 1 integrons,
PCR amplification of the variable regions in 22 strains yielded two
types of fragment. The sizes were ~1.6 kb and 0.7 KB, respectively
(Fig. 3). Confirmed by the gene
sequencing, four S. flexneri 4a strains carried the
dfrA17-aadA5 (1,624 bp) drug-resistant gene cassette, which
was the dihydrofolate reductase gene, and aminoglycoside
acyltransferase gene. A total of 18 S. flexneri 2a strains
carried the dfrV (729 bp) drug-resistant gene cassette,
which was the dihydrofolate reductase gene and aminoglycoside
acyltransferase gene. The PCR amplification of four stains yielded
nothing suggest an empty cassette. The assessment of antibiotic
susceptibility showed that the drug resistance of pathogens to
ampicillin and other drugs was severe, suggesting that the drug
resistance was associated with the insertion of a 3′ conservative
region into the C-terminal conservative region (9). The variable regions of 134 S.
flexneri strains were amplified with the intI1ca-IS1ca
primer. A total of 134 strains yielded 2.4 kb fragments, as shown
in Fig. 4. The products had the
same enzymatic maps following HindIII enzyme digestion. Four
strains were randomly selected for sequencing. The carried drug
resistant gene cassette was bla0XA-30-aadA1 (2,453 bp),
which was the β-lactamase and aminoglycoside adenosine
acyltransferase gene. A total of 134 strains with atypical class 1
integron were resistant to ampicillin, streptomycin, tetracycline
and chloromycetin. Only 10 of the 26 strains without atypical class
1 integron were resistant to more than four antibiotics. There was
a significant difference between two groups (χ2=35.96,
P<0.01). The detailed results of the resistance gene in class 1
integron variable regions are shown in Table III.
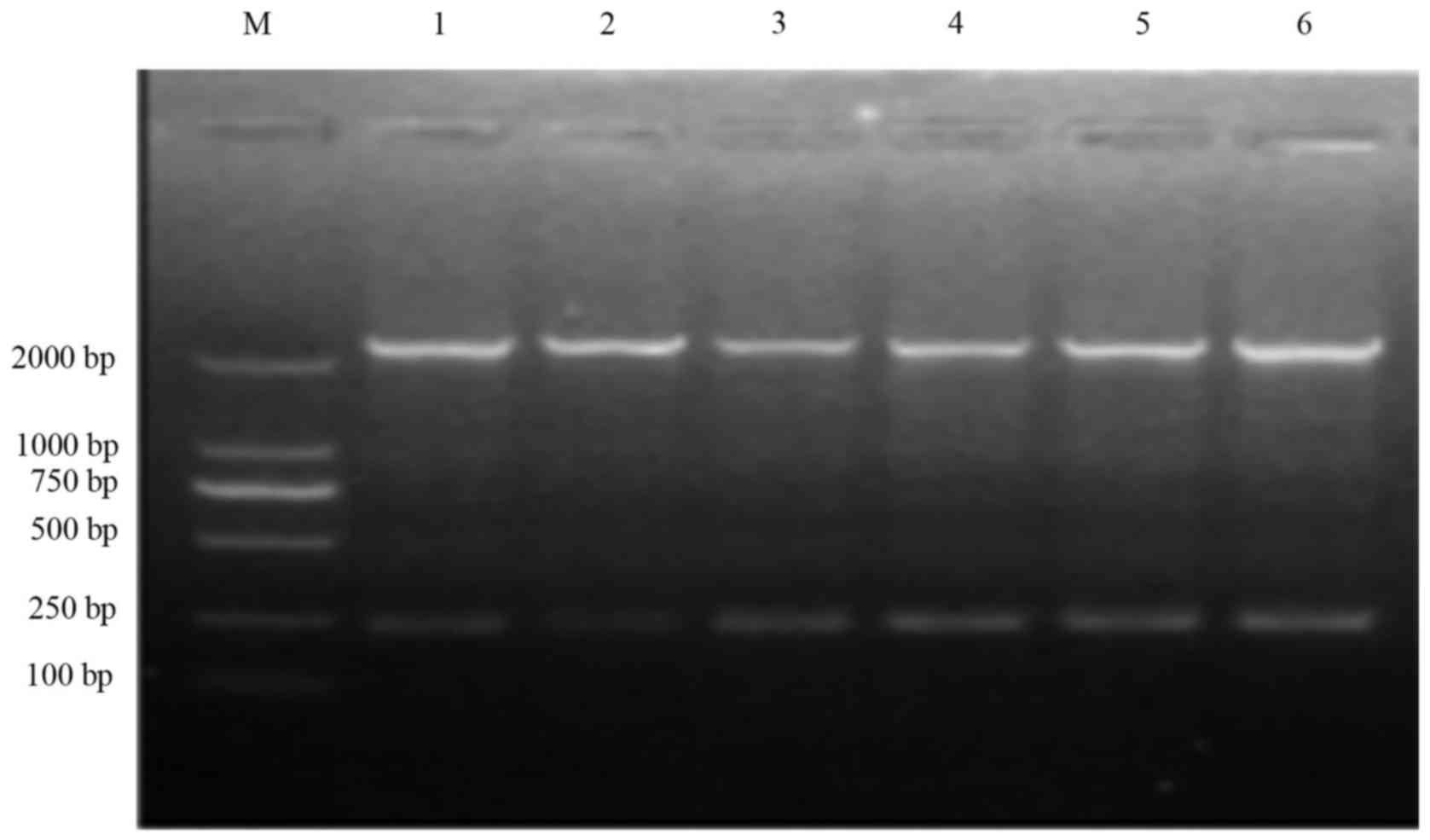
Resistance gene cassette assessment
and sequencing in variable regions of integrons
The variable region of integrons in the 136 strains
was amplified by integron PCR. Following electrophoresis, all the
strains yielded 2.2 kb fragments (Fig.
5). Following hinf1 digestion, the amplification
products manifested the same characteristic bands in
electrophoresis. Verified by gene sequencing, the drug-resistant
gene carried by bacteria was the dfrA1-sat1-aadA1 gene
(2,224 bp), which was responsible for the drug resistance to
trimethoprim, streptomycin and aminoglycoside. The results of the
resistance gene in class 2 integron variable regions are shown in
Table IV.
| Table IV.Drug-resistant phenotype integrons
and gene cassette of 56 Shigella flexneri strains. |
Table IV.
Drug-resistant phenotype integrons
and gene cassette of 56 Shigella flexneri strains.
|
| Class 1 integron
and gene cassettes | Class 2 integron
and gene cassettes |
|---|
|
|
|
|
|---|
| Drug-resistant
phenotype (number of strains) | Class 1 integrase
gene | Gene cassettes | Class 2 integrase
gene | Gene cassettes |
|---|
| ATCR (2) | intI1
(n=2) |
blaOXA-30-aadA1 | intI2
(n=0) | – |
|
| intI1
(n=4) | – | intI2
(n=2) |
dfrA1-sat1-aadA1 |
|
| intI1
(n=16) |
blaOXA-30-aadA1 | intI2
(n=16) |
dfrA1-sat1-aadA1 |
| ATCRS (40) | intI1
(n=12) |
blaOXA-30-aadA1 | intI2
(n=12) |
dfrA1-sat1-aadA1 |
|
|
| dfr V |
|
|
|
| intI1
(n=6) | dfr V | intI2
(n=6) |
dfrA1-sat1-aadA1 |
|
| intI1
(n=4) |
blaOXA-30-aadA1 | intI2
(n=0) | – |
|
| intI1
(n=18) |
blaOXA-30-aadA1 | intI2
(n=18) |
dfrA1-sat1-aadA1 |
| ATCRP (2) | intI1
(n=2) |
blaOXA-30-aadA1 | intI2
(n=2) |
dfrA1-sat1-aadA1 |
| ATCRPL (10) | intI1
(n=10) |
blaOXA-30-aadA1 | intI2
(n=10) |
dfrA1-sat1-aadA1 |
| ATCRSPL (22) | intI1
(n=18) |
blaOXA-30-aadA1 | intI2
(n=18) |
dfrA1-sat1-aadA1 |
|
| intI1
(n=4) |
blaOXA-30-aadA1
dfrA17-aadA5 | intI2
(n=4) |
dfrA1-sat1-aadA1 |
| ATCRSPLX (12) | intI1
(n=12) |
blaOXA-30-aadA1 | intI2
(n=12) |
dfrA1-sat1-aadA1 |
| ATCRSXZ (30) | intI1
(n=30) |
blaOXA-30-aadA1 | intI2
(n=30) |
dfrA1-sat1-aadA1 |
| ATCRSPLXZ (6) | intI1
(n=6) |
blaOXA-30-aadA1 | intI2
(n=6) |
dfrA1-sat1-aadA1 |
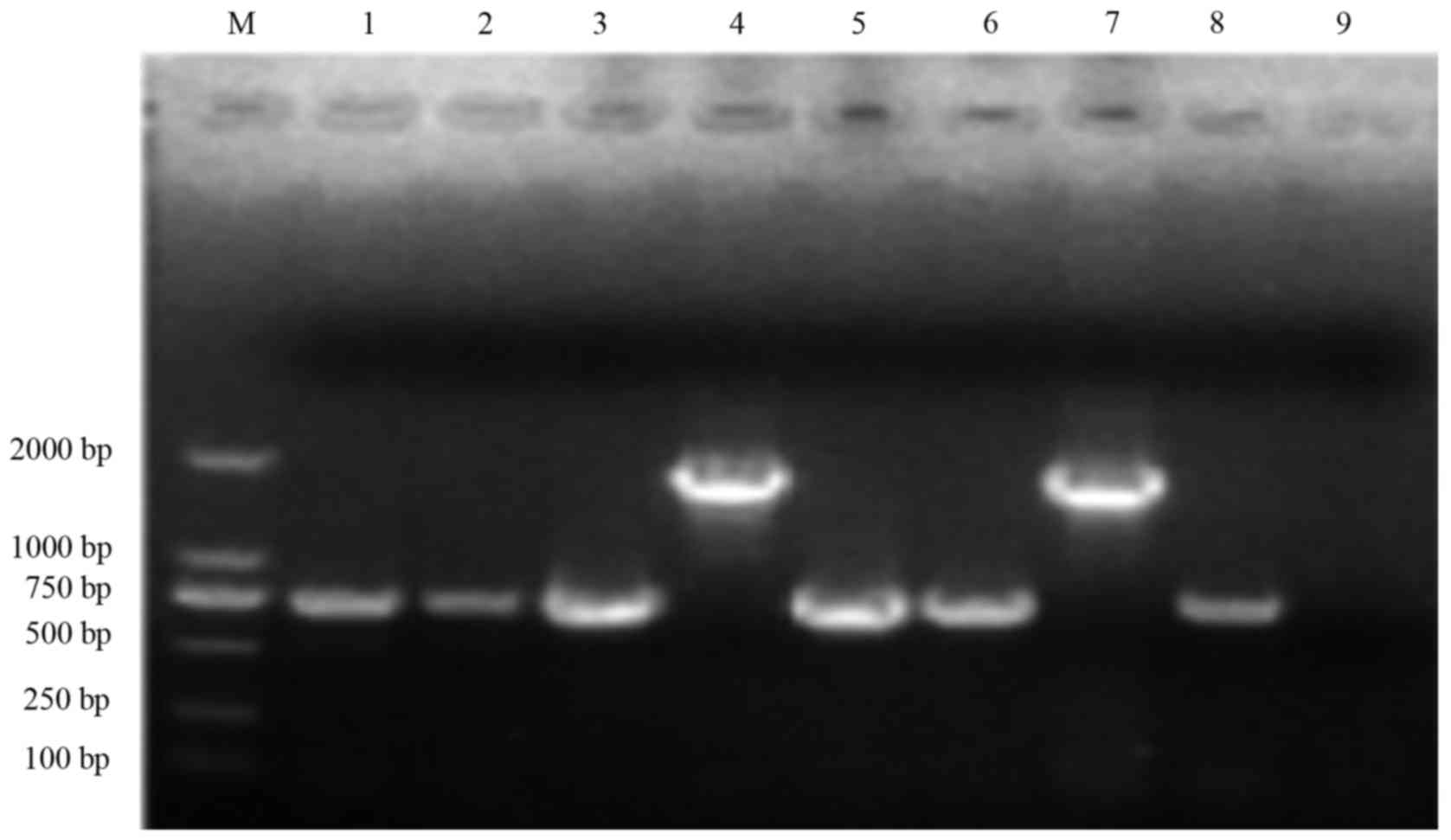
Analysis of acrAB-tolC and its
associated regulatory genes in multi-drug resistant Shigella
There were 12 strains with gene deletions among the
122 multi-drug resistant strains and two strains with gene
deletions in 17 sensitive strains. Therefore 110 multi-drug
resistant strains and 15 sensitive strains yeilded 131, 510, 604,
1,100 and 800 bp sequences (Fig.
6-D). Verified by gene sequencing, these fragments were the PCR
amplification products of acrA, acrB, marOR, soxS and
acrB. There were 110 cases in drug resistant group and four
cases in the sensitive group. The gene analysis showed a low
mutation rate in the acrA, acrB, acrR and soxS genes.
In the multi-drug resistant strains, a base CATT deletion in the
36, 37, 38 and 39 sites of marOR was detected. However, no
marOR mutation was found in sensitive strains.
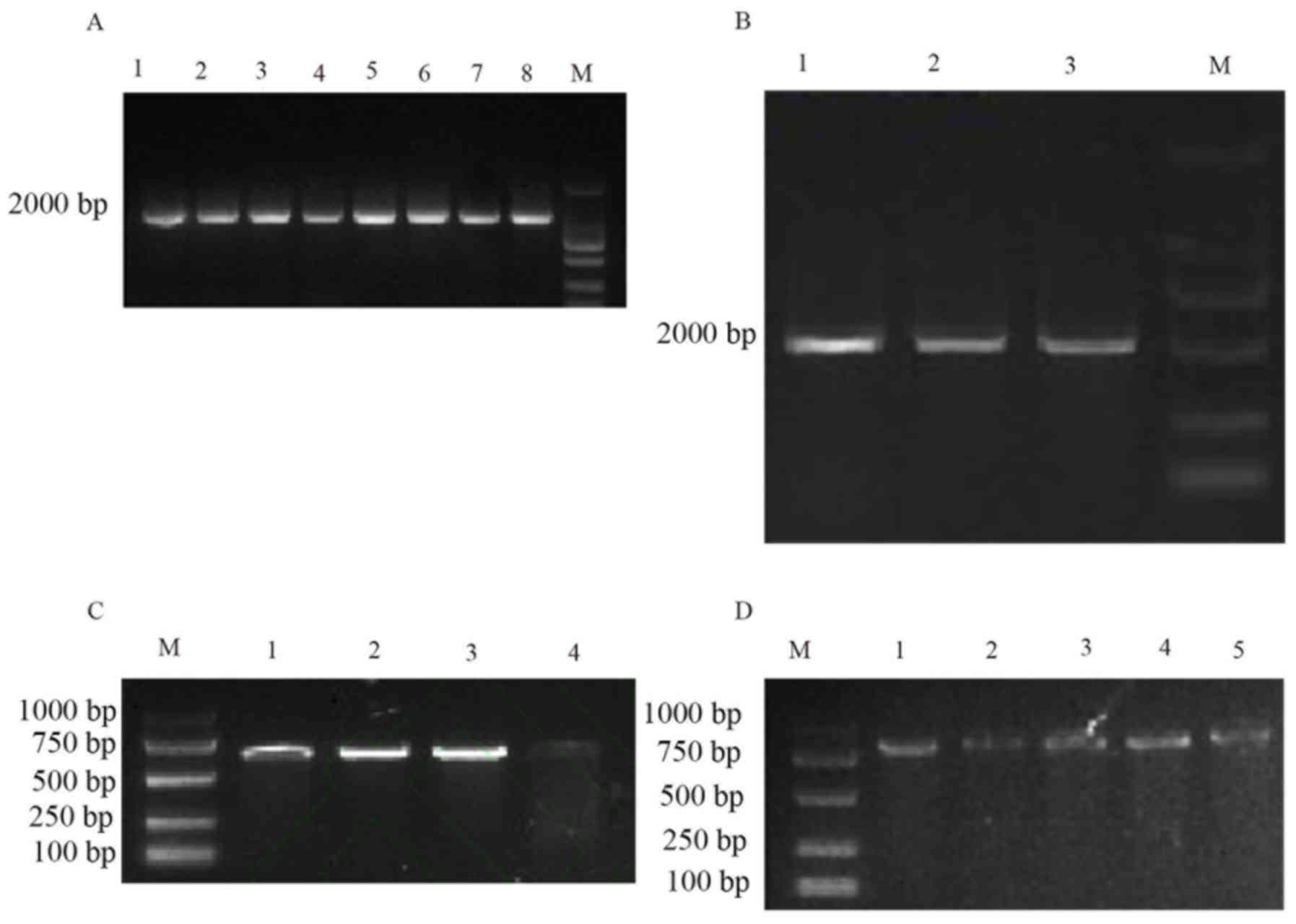 | Figure 6.Polymerase chain reaction
amplification and electrophoretogram of multi-drug resistance gene
acrAB-tolC and its regulatory genes (A) multi-drug resistance gene
acrA, (B) multi-drug resistance gene acrB, (C) multi-drug
resistance gene marOR and (D) multi-drug resistance gene acrR. 1,
368 strains; 2, 157 strains; 3, 3,171 strains; 4, 187 strains; 5,
4,536 strains; 6, 1,113 strains; 7, 22 strains; 8, N8; M, DNA
marker (DL2000). Upper to lower progressively 2,000, 1,000, 750,
500, 250 and 100 bp. M, DNA marker; 1–8 are the specimen
numbers. |
Discussion
Bacillary dysentery caused by Shigella is a
serious intestinal infectious disease and the incidence ranks third
among the national statutory B infectious diseases. In addition to
transient immunity following infection, the absence of
cross-immunoreactivity between subtypes and problems in the immune
system of children, the increased drug-resistance of
Shigella poses a serious threat to childrens' health. A
previous study showed that Shigella had the AcrAB-TolC
active efflux pump, which caused multi-drug resistance to organic
solvents, dyes, detergents and virious antibiotics drugs (12). In another study (13), the high expresion level of the
active efflux pump can enhance the resistance of bacteria to
organic solvents. It also been reported that the resistance of
Escherichia coli to organic solvents was affected by certain
inherited traits, including the imp/ostA gene, which coded a
protein with a molecular mass of 87,000. The protein was attached
to the outer membranes. The low expression of the gene increased
the sensitivity to N-hexane (14).
Certain cell elements that do not belong to the antivie efflux
pump, for example the derivatives of the O-sidechain of
lipopolysaccharides or the isomers of fatty acid membranes, may
have an effect on the resistance of Pseudomonas to organic
solvents (15).
The results of the present study showed that 91.1%
S. flexneri strains were multi-drug resistant. According to
the shigella treatment recommendations of the American
Clinical and Laboratory Standards Institute, ampicillin,
sulfamethoxazole/trimethoprim were not suitable for the empirical
clinical medication. Although the drug resistance rate of
ciprofloxacin and levofloxacin was 32.9%, the fluroquinolones were
not suitable for pediatric patients due to the potential damage to
cartilage in infants and children. The drug-resistance mechanism
was associated with multiple-site mutations of gene gyrA
and/or parC. The drug-resistance rate to cefotaxime and
cefotaxime was <30.4%, which meant that these drugs were
suitable for use in the treatment of bacillary dysentery in
children. It is noteworthy that the drug resistance was
significantly higher than that reported previously In China and
elsewhere (16,17), sugesting that, in case of treatment
failure with third generation cephalosporins or in case of
pediatric dental patents, cefoxitin, cefoperazone/sulbactam and
imipenem may be applied in the treatment.
The integron-mediated muhidrug-resistance has been
verified previously. Class 1 integron was detected in 91.1% of 158
S. flexneri strains. Class 2 integron was detected in 86.1%
of 158 S. flexneri strains. The propotions were
significantly higher, compared with those reported in South Korea
and Iran (18,19). The islolating regions and epidemic
strains may accout for this. The typical class 1 integron gene was
present in 15.2% of the isolates screened. The drug resistance gene
cassette was frV and dfrA17-aadA5. The atypical class
1 integron accounted for 86.1% strains. The drug resistance gene
cassette was blaOXA30-aadA1. The typical and atypical
class 1 integrons were present in 10.1% of isolates screened,
similar with the relative reports (20,21).
The typical class 1 integron was present in 11.4% of the S.
flexneri isolates screened. This result has not been reported
previously, to the best of our knowledge. In individual studies,
the dfrv gene was present in Escherichia coli,
Salmonella bacteria and Gram-negative bacilli in waste water
(20–22). Whether the resistance genes were
transfered from these bacteria requires further investigation.
A total of 134 strains with atypical class 1
integron were resistant to ampicillin, streptomycin, tetracycline
and chloromycetin. Only 10 of the 26 strains without the atypical
class 1 integron were resistant to more than four antibiotics.
There was a significant difference between them
(χ2=35.96, P<0.01). This difference is associated
with a situation that the blaOXA-30-aadA1 gene
cassette in the atypical class 1 integron variable region was
adjacent to the chloramphenicol- and tetracycline-resistant
determinants (23). Those strains
with no atypical class 1 integrons, which were resistant to the
four antibiotics may have other drug resistance mechanisms. There
were 10.1% strains (S. flexneri 2A) possessing typical and
atypical class 1 integron class 1 integrons (24), which deteriorate the drug
resistance of S. flexneri. The typical class 1 integron was
found in conjugative plasmids and can mediate the horizontal gene
transfer. In the present study, 86.1% S. flexneri isolates
carried the class 2 integron and the gene cassette was
dfrA1-sat1-aadA1, which was consistant with other reports
(25,26). The gene cassettes reported
currently comprise dfrAl-sat1, dfrAl-sat2-aadA1 and
sat2-aadA1 (27–29). As the class 2 integron integrase is
inadequate for integrating novel resistance genes from externally
or to resect the resistance genes from its own variable region, the
drug resistance gene cassettes of class 2 integron variable regions
are relatively conservative.
The drug resistance rate to
sulfamethoxazole/trimethoprim was 69.6%. The strains resistant to
sulfamethoxazole/trimethoprim were compared with the sull
gene carried in the atypical class 1 integron, the dfrAl
gene carried in the class 2 integron, and the two together,
suggesting that the two genes were not associated with the drug
resistance. The two genes may be nonfunctional structural genes.
The mechanism undferlying the drug resistance to
sulfamethoxazole/trimethoprim requires further investigation.
The single-stranded conformation polymorphism
suggested a low mutation rate of acrA, acrB and tolC
in drug-resistant Shigella. The expression of AcrA and AcrB
are regulated by several regulators. Of all the regulators the
acrR suppressors and marOR operon are important. The
results demonstrated a low mutation rate of marOR in
drug-resistant Shigella, suggesting that the mutaion of the
reglatory gene contributed more to the drug resistance of
Shigella than the mutaion of a pump gene. Further
investigations aim to examine the association between the mutation
of regulatory gene marOR and the expression of acrA,
acrB and tolC, which may provide a theoretical basis for
the drug-resistant mechanism mediated by the active efflux pump of
Shigella.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
QPM was responsible for specimen collection,
bacterial identification, and experimental procedures. MXY was
responsible for experimental operations and data collection. LS and
JWL were responsible for data collection and statistical analysis.
GYY was responsible for experimental design and data review.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Jinan Infectious Disease Hospital Affiliated To Shandong University
(Shandong, China).
Consent for publication
Written informed consent was obtained from all
patients or patient guardians for the present study.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Partridge SR: Analysis of antibiotic
resistance regions in Gram-negative bacteria. FEMS Microbiol Rev.
35:820–855. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Poole K: Efflux-mediated multiresistance
in Gram-negative bacteria. Clin Microbiol Infect. 10:12–26. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang HY, Duan GC and Xi YL: Distribution
and expression of active efflux pump acrAB in clinical isolates of
Shigella. Chin J Public Health. 21:685–687. 2005.
|
|
4
|
Lv RL, Duan G, Ca and Song CH: Analysis on
acrR, marOR multi-drug regulated gene mutations in clinical
isolates of Shigella. Chin J Public Health. 24:865–965.
2008.
|
|
5
|
Cockerill FR, Cockerill F, Adler J, Dudley
MN, Patel JB, Eliopoulos GM and Bradford PA: Performance standards
for antimicrobial susceptibility testing: Twenty-first
informational supplement. Clin Lab Stand Inst. 31:21–42. 2011.
|
|
6
|
White PA, McIver CJ and Rawlinson WD:
Integrons and gene cassettes in the enterobacteiaceae. Antimicrob
Agents Chemother. 45:2658–2661. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ploy MC, Denis F, Courvalin P and Lambert
T: Molecular characterization of integrons in Acinetobacter
baumannii: Description of a hybrid class 2 integron. Antimicrob
Agents Chemother. 44:2684–2688. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dalsgaard A, Forslund A, Serichantalergs O
and Sandvang D: Distribution and content of class 1 integrons in
different Vibrio cholerae O-serotype strains isolated in Thailand.
Antimicrob Agents Chemother. 44:1315–1321. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pan JC, Ye R, Meng DM, Zhang W, Wang HQ
and Liu KZ: Molecular characteristics of class 1 and class 2
integrons and their relationships to antibiotic resistance in
clinical isolates of Shigella sonnei and Shigella
flexneri. J Antimicrob Chemother. 58:288–296. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, et al: Relation of acrAB-tolC efflux pump and
marOR regulatory gene mutation with antimicrobial resistance in
Shigella. spp. J Jilin Univ. 36:45–48. 2010.
|
|
11
|
Rahman M, Mauff G, Levy J, Couturier M,
Pulverer G, Glasdorff N and Butzler JP: Detection of 4-quinolone
resistance mutation in gyrA gene of Shigella dysenteriae
type 1 by PCR. Antimicrob Agents Chemother. 38:2488–2491. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abe S, Okutsu T, Nakajima H, Kakuda N,
Ohtsu I and Aono R: n-Hexane sensitivity of Escherichia coli
due to low expression of imp/ostA encoding an 87 kDa minor protein
associated with the outer membrane. Microbiology. 149:1265–1273.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sikkema J, de Bont JA and Poolman B:
Mechanism of membrane toxicity of hydrocarbons. Microbiol Rev.
59:201–222. 1995.PubMed/NCBI
|
|
14
|
Wang XN, Wei DJ, Zhang JL and Li WS: Study
on extended-spectrum β-lactamases and their genotypes of
Shigella isolated from Tianjin. Chin J Infect Dis. 30:71–75.
2012.
|
|
15
|
Varghese SR and Aggarwal A: Extended
spectrum beta-lactamase production in Shigella isolates-a
matter of concern. Indian J Med Microbiol. 29:76–78. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hong S, Choi YH, Choo YA, Choi Y, Choi SY,
Kim DW, Lee BK and Park MS: Genetic characterization of atypical
Shigella flexneri isolated in Korea. J Microbiol Biotechnol.
20:1457–1462. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Eftekhari N, Bakhshi B, Pourshafie MR,
Zarbakhsh B, Rahbar M, Hajia M and Ghazvini K: Genetic diversity of
Shigella spp. and their integron content. Foodborne Pathog
Dis. 10:237–242. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schaumburg F, Alabi AS, Kaba H, Lell B,
Becker K, Grobusch MP, Kremsner PG and Mellmann A: Molecular
characterization of Shigella spp. from patients in Gabon
2011–2013. Trans R Soc Trop Med Hyg. 109:275–279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sow Gassama A, Aïdara-Kane A, Barraud O,
Gatet M, Denis F and Ploy MC: High prevalence of
trimethoprim-resistance cassettes in class 1 and 2 integrons in
Senegalese Shigella spp isolates. J Infect Dev Ctries.
4:207–212. 2010.PubMed/NCBI
|
|
20
|
Kim TE, Jeong YW, Cho SH, Kim SJ and Kwon
HJ: Chronological study of antibiotic resistances and their
relevant genes in Korean avian pathogenic Escherichia coli
isolates. J Clin Microbiol. 45:3309–3315. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nógrády N, Gadó I, Tóth A and Pászti J:
Antibiotic resistance and class 1 integron patterns of
non-typhoidal human Salmonella serotypes isolated in Hungary in
2002 and 2003. Int J Antimicrob Agents. 26:126–132. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rosser SJ and Young HK: Identification and
characterization of class 1 integrons in bacteria from an aquatic
environment. J Antimicrob Chemother. 44:11–18. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luck SN, Turner SA, Rajakumar K,
Sakellaris H and Adler B: Ferric dicitrate transport system (Fec)
of Shigella flexneri 2a YSH6000 is encoded on a novel
pathogenicity island carrying multiple antibiotic resistance genes.
Infect Immun. 69:6012–6021. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu JY, Duan GC, Yang HY, Fan QT and Xi
YL: Atypical class 1 integron coexists with class 1 and class 2
integrons in multi-drug resistant Shigella flexneri isolates
from China. Curr Microbiol. 62:802–806. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang YL, Xu YL, Zhuang L, Qian HM, Gu L,
Gu B, Liu GY and Pan SY: Molecular epidemiology and integron
analysis of Shigella flexneri isolates from Jiangsu province
in 2011. Chi J Clin Infect Dis. 7:34–38. 2011.
|
|
26
|
Wang J, Liang F, Wu XM and Qi W: Transfer
patterns of integron-associated and antibiotic resistance genes in
S. flexneri during different time intervals in Tianjin,
China. Indian J Med Microbiol. 32:256–260. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xia R, Ren Y, Guo X and Xu H: Molecular
diversity of class 2 integrons in antibiotic-resistant
gram-negative bacteria found in wastewater environments in China.
Ecotoxicology. 22:402–414. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang CY, Lu PL, Lin CC, Lee TM, Tsai MY
and Chang LL: Integron types, gene cassettes, antimicrobial
resistance genes and plasmids of Shigella sonnei isolates
from outbreaks and sporadic cases in Taiwan. J Med Microbiol.
60:197–204. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ud-Din AI, Wahid SU, Latif HA, Shahnaij M,
Akter M, Azmi IJ, Hasan TN, Ahmed D, Hossain MA, Faruque AS, et al:
Changing trends in the prevalence of Shigella species:
Emergence of multi-drug resistant Shigella sonnei biotype g
in Bangladesh. PLoS One. 8:e826012013. View Article : Google Scholar : PubMed/NCBI
|