Introduction
Severe burn-induced remote acute lung injury (ALI)
is a universal issue challenging clinical intensive treatment
(1). Severe burns have been
reported to induce marked alterations to the organic internal
environment. Factors including traumatic stress, direct injuries of
heating power and hypoperfusion may induce the upregulation of
expression levels of pro-inflammatory cytokines (2). In addition, these factors may
activate a variety of inflammatory cells, including mononuclear
macrophages, neutrophil granulocytes, lymphocytes, platelets and
endothelial cells (3,4). Furthermore, adhesion molecules of
vascular endothelial cells (VECs) and while blood cells (WBCs) have
been reported to be upregulated (3,4). The
activated WBCs may then induce a respiratory burst and
degranulation as the adhesion intensifies. As a result, metabolites
and proinflammatory cytokines, including fibrinolysin, active
oxygen and arachidonic acid, would be released. This then causes
broader damage to VECs and other tissues (5).
Severe burn-induced remote ALI is characterized by
the pathological features of a uncontrolled inflammatory response,
and it may broadly damage pulmonary VECs and alveolar epithelial
cells (4). Severe-burn induced
remote ALI has been reported to exhibit morbidity and mortality
rates of 20–40% (4). However, no
treatments are currently available within intensive care units
(4). Uncontrollable inflammatory
mediators are closely associated with ALI (4). Macrophages were in patients with ALI
to induce inflammation (6). In
addition, numerous inflammatory cytokines are released, including
interleukin (IL)-6, IL-18, tumor necrosis factor-α (TNF-α) and
interferon-γ. Furthermore, macrophages, neutrophils, granulocytes,
lymphocytes and endothelial cells are activated; oxygen radicals,
proteolytic enzymes and arachidonic acid are also released
(4). Inflammatory responses within
the lungs may induce injury to the alveolar capillary membrane.
Subsequently, the ALI is initiated and the inflammatory response is
induced (7).
Alterations in microRNA (miR) expression have been
associated with immune responses, the inflammatory signaling
pathway and the pathogenesis of inflammatory pulmonary diseases,
such as ALI (8). Therefore, miR
may be promising novel therapeutic targets. miR-induced alterations
in the expression of associated genes are generally moderate
(9); however, its results may
affect the expression of several genes and numerous biological
processes (10). Therefore, miR
may be considered as a potential marker of ALI. The protective
effects of miR-146a on severe burn-induced remote ALI in rats were
examined in the present study. The results suggested that miR-146a
protected against severe burn-induced ALI via the anti-inflammatory
pathway.
Materials and methods
Animals
All experimental protocols were undertaken in
accordance with the Guide for the Care and Use of Laboratory
Animals by the National Institutes of Health (11), with the approval of the Animal
Experimental Ethics Committee of the First Affiliated Hospital of
Nanchang University (Nanchang, China). Healthy male Sprague-Dawley
rats (200–250 g, n=12) were purchased from Laboratory Animal Center
of the Medical Department of Nanchang University (Nanchang, China)
and used in the present study. The animals were housed at 22–24°C,
55–60% humidity, 0.038% CO2, 12 h light/dark cycle and
fed a standard animal diet with food and tap water ad libitum.
Experimental design and burn
procedure
The rats were randomly divided into three groups,
each containing 10 rats: Sham-operated group, ALI group and
oleuropein-treated group. The rats of the ALI and
oleuropein-treated groups were anesthetized with 30 mg/kg of
pentobarbital sodium Sigma-Aldrich (Merck KGaA, Darmstadt, Germany)
and shaved on the dorsal and lateral surfaces. The surface area of
the skin was immersed in 100°C water for 5–10 sec on the dorsal
surface. The skin of all rats were quickly dried, total body
surface area was 30% at all skin area. Sham-operated rats were
anesthetized with 30 mg/kg of pentobarbital and shaved on the
dorsal and lateral surfaces.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol®
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) from lung tissue
samples or cells and ~1 µg total RNA was then used to produce cDNA
using TaqMan® MicroRNA Reverse Transcription kit (Thermo
Fisher Scientific, Inc.) at 37°C for 60 min and 85°C for 1 min by a
TaqMan 7900 (ABI) real-time PCR machine (Invitrogen; Thermo Fisher
Scientific, Inc.). miR-146a, inducible nitric oxide synthase (iNOS)
and cyclooxygenase-2 (COX-2) mRNA expression levels were determined
by RT-qPCR using TransStart™ SYBR-Green qPCR Supermix (Beijing
Transgen Biotech Co., Ltd., Beijing, China). The following
thermocycling conditions were used for the PCR: Initial
denaturation at 95°C for 10 min; 40 cycles of 95°C for 15 sec, 60°C
for 30 sec and 72°C for 30 sec. The primer sequences used were as
follows: miR-146a forward, 5′-TGAGAACTGAATTCCATGGGTT-3′ and
reverse, 5′-TCACCCGTAGAACCGACCTT-3′; iNOS forward,
5′-CCCTTCCGAAGTTTCTGGCAGCAG-3′ and reverse,
5′-GGCTGTCAGAGCCTCGTGGCTTTG-3′; COX-2 forward,
5′-ATGCTCCTGCTTGAGTATGT-3′ and reverse, 5′-CACTACATCCTGACCCACTT-3′;
GAPDH forward, 5′-AACTTTGGCATTGTGGAAGG-3′ and reverse,
5′-ACACATTGGGGGTAGGAACA-3′; and U6 forward,
5′-ATTGGAACGATACAGAGAAGATT-3′ and reverse,
5′-GGAACGCTTCACGAATTTG-3′. The relative levels of the target genes
were normalized using the 2−ΔΔCq method (12).
Cell culture and transfection
16HBE cells (human bronchial epithelial) were
obtained from the Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Shanghai, China) and maintained in RPMI-1640
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) in 5%
CO2 and 95% O2 at 37°C. miR-146a
(5′-UGAGAACUGAAUUCCAUGGGUU-3′ and 5′-CCCAUGGAAUUCAGUUCUCAUU-3′),
anti-miR-146a (5′-AACCCAUGGAAUUCAGUUCUCA-3′ and
5′-GCTGTCAACGATACGCTACGTAACG-3′), and negative mimics
(5′-CCCCCCCCCCCCC-3′ and 5′-CCCCCCCCCCCCCC-3′) were obtained from
Sangon Biotech Co., Ltd., (Shanghai, China). 16HBE cells were
transfected with 200 ng of miR-146a, anti-miR-146a or negative
mimics using Lipofectamine® 2000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). Following transfection for 48 h,
ALI was induced within 16HBE cells using lipopolysaccharide (LPS;
100 ng/ml; Beyotime Institute of Biotechnology, Nanjing, China) for
6 h. Control, 16HBE cells were transfected with negative control
mimics for 48 h and treated with LPS (100 ng/ml) and TAK-242 (2 nM;
MedChemExpress, Monmouth Junction, NJ, USA) for 6 h.
Measurement of nuclear factor (NF)-κB,
IL-6, IL-10 and TNF-α
Bronchoalveolar lavage samples were collected and
centrifuged at 12,000 × g for 10 min at 4°C. The supernatant fluids
were used to measure NF-κB (cat. no. H202), IL-6 (cat. no. H007),
IL-10 (cat. no. H009) and TNF-α (cat. no. H052) levels using ELISA
kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China)
according to the manufacturer's protocols.
Western blot analysis
Protein (50 µg) was excised from lung tissue using
homogenated extraction buffer on ice and centrifuged at 12,000 × g
for 10 min at 4°C in radioimmunoprecipitation assay (Sangon Biotech
Co., Ltd.). Protein concentrations were determined using a
Bicinchoninic Acid protein assay reagent (Sangon Biotech Co.,
Ltd.). Protein extracts (50 µg) were separated via 10% SDS-PAGE
(Sangon Biotech Co., Ltd.) and electrotransferred onto a
nitrocellulose membrane. The membranes were blocked with 5% skimmed
milk in TBS with 0.1% Tween-20 for 1 h at 37°C and immunoblotted
with primary antibodies specific for NF-κB (cat. no. sc-109,
1:3,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
Toll-like receptor 4 (cat. no. sc-10741, TLR4; 1:3,000; Santa Cruz
Biotechnology, Inc.) and GAPDH (cat. no. sc-25778, 1:4,000; Santa
Cruz Biotechnology, Inc.) overnight at 4°C. Following three washes
with Tris-buffered saline-0.1% Tween-20, the membrane was incubated
with anti-rabbit immunoglobulin G (H+L), Biotinylated secondary
antibody (cat. no. 14708, 1:5,000; Cell Signaling Technology, Inc.,
Danvers, MA, USA) for 1 h at 37°C and detected using an enhanced
chemiluminescence plus detection kit (Amersham; GE Healthcare,
Chicago, IL, USA) and exposed to film and analyzed using Image Lab
3.0 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All data were expressed as the mean ± standard error
of the mean using SPSS 17.0 (SPSS, Inc., Chicago, IL, USA). The
results were analyzed statistically using one-way analysis of
variance and Duncan's multiple range test. P<0.05 was considered
to indicate a statistically significant difference.
Results
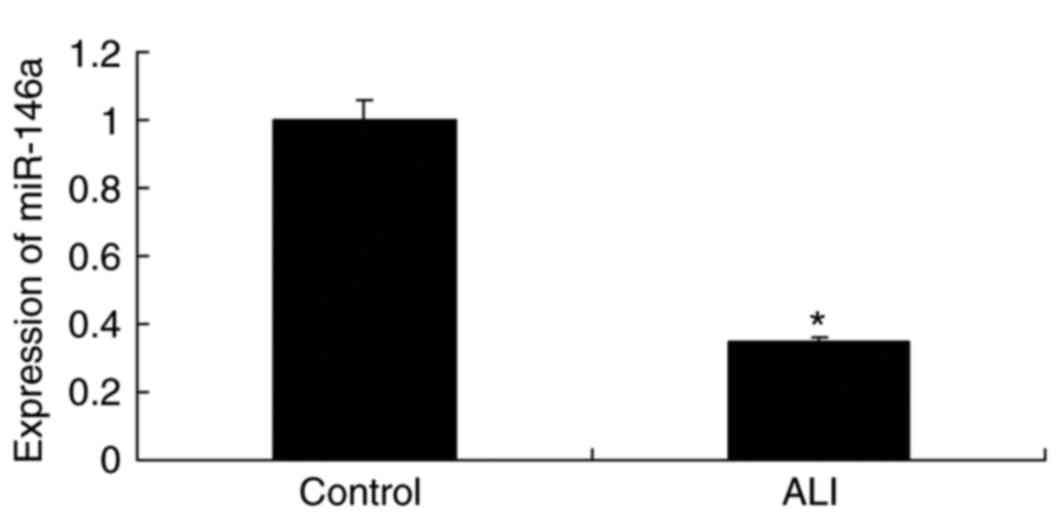
Expression of miR-146a in burn-induced
ALI rat
As presented in Fig.
1, the expression levels of miR-146a were significantly
downregulated in the rat model of burn-induced ALI compared with
the levels in the control group.
Downregulation of miR-146a increases
inflammation in a model of ALI in 16HBE cells
Subsequently, the function of miR-146a in ALI was
explored and miR-146a expression was inhibited using anti-miR-146a
mimics. Levels of NF-κB, IL-6, IL-10 and TNF-α were measured using
ELISA kits. As demonstrated in Fig.
2, NF-κB, IL-6 and TNF-α levels within the model of ALI were
significantly higher in response to miR-146a downregulation than
those in the control group. IL-10 expression levels within the ALI
model were significantly reduced in response to miR-146a
downregulation compared with those in the control group.
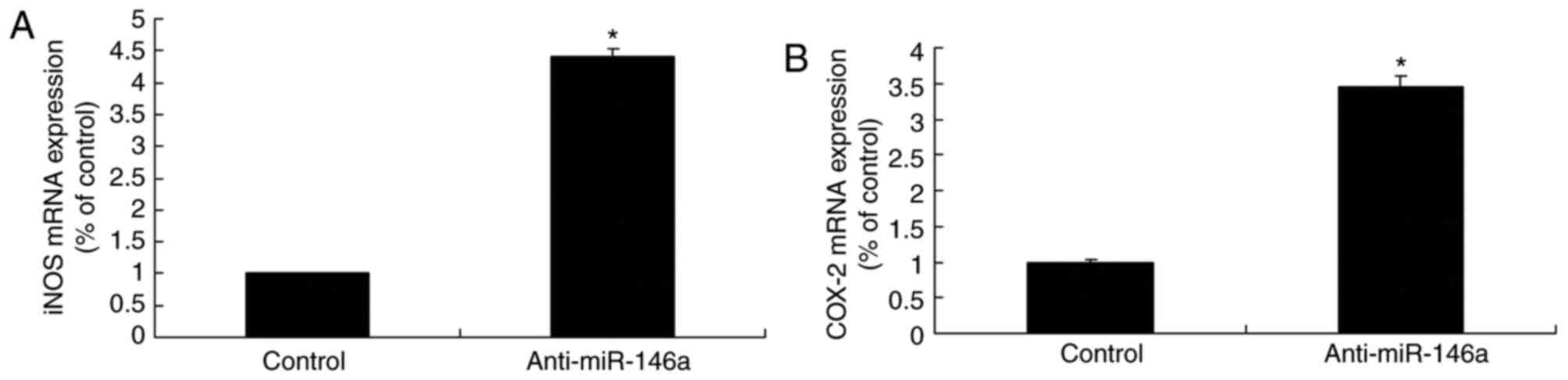
Downregulation of miR-146a increases
iNOS and COX-2 mRNA expression levels in a model of ALI in 16HBE
cells
Downregulation of miR-146a significantly induced
iNOS and COX-2 mRNA expression within the model of ALI compared
with the levels in the control group (Fig. 3). There results demonstrated that
anti-miR-146a regulates iNOS and COX-2 expression to affect the
development of ALI.
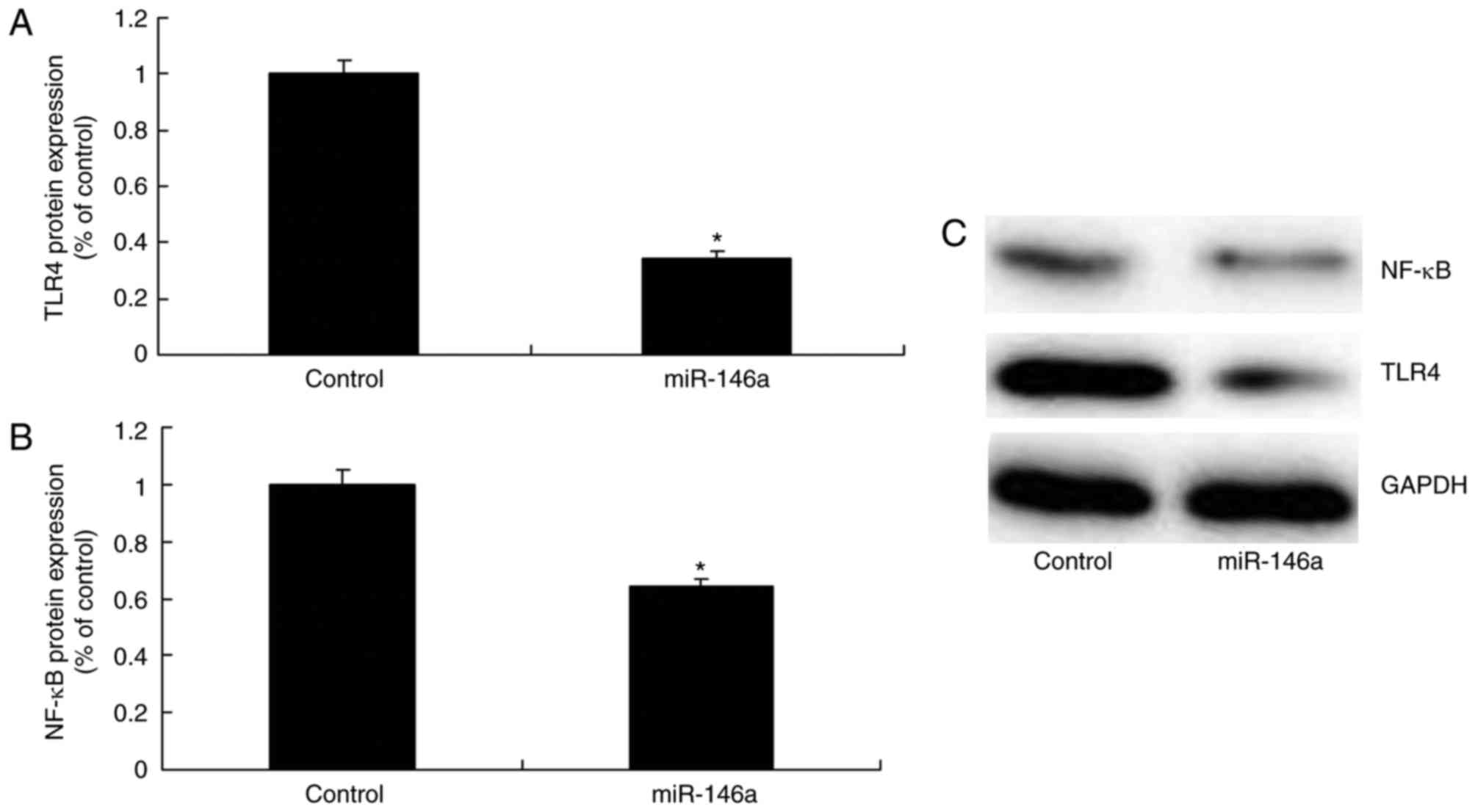
Downregulation of miR-146a upregulates
the TLR4/NF-κB signaling pathway in a model of ALI in 16HBE
cells
The mechanism of the effect of miR-146a on
inflammation within the model of ALI was investigated by analyzing
TLR4 and NF-κB protein expression levels. As presented in Fig. 4, the downregulation of miR-146a
significantly increased TLR4 and NF-κB protein expression levels
within the ALI model compared with the levels in the control
group.
Upregulation of miR-146a decreases
inflammation in a model of ALI in 16HBE cells
miR-146a expression levels were upregulated to
investigate the mechanism of miR-146a on inflammation within the
model of ALI. As demonstrated in Fig.
5, upregulation of miR-146a significantly decreased NF-κB, IL-6
and TNF-α expression levels, and increased IL-10 expression levels
within the model of ALI compared with the levels in the control
group.
Upregulation of miR-146a decreases
iNOS and COX-2 mRNA expression levels in an ALI model in 16HBE
cells
iNOS and COX-2 mRNA expression levels in the ALI
model were significantly reduced by the increased expression of
miR-146a compared with the level in the control group (Fig. 6). Therefore, miR-146a reduced the
expression of iNOS and COX-2 expression to ameliorate ALI.
Upregulation of miR-146a suppresses
the TLR4/NF-κB signaling pathway within a model of ALI in 16HBE
cells
TLR4 and NF-κB protein expression levels were
significantly suppressed within the model of ALI following miR-146a
upregulation compared with the levels in the control group
(Fig. 7).
TLR4 inhibitor reduces the function of
anti-miR-146a on the TLR4/NF-κB signaling pathway within a model of
ALI in 16HBE cells
In order to confirm the role of the TLR4/NF-κB
signaling pathway in the effect of miR-146a on ALI, TAK-242
inhibitor was employed to reduce TLR4 and NF-κB protein expression.
The TLR4 inhibitor suppressed TLR4 and NF-κB protein expression
levels within the model of ALI following anti-miR-146a treatment,
compared with in the levels observed in the anti-miR-146a group
(Fig. 8).
TLR4 inhibitor reduces the function of
anti-miR-146a on inflammation in a model of ALI in 16HBE cells
The induction of NF-κB, IL-6 and TNF-α levels, and
inhibition of IL-10 levels within the model of ALI following
anti-miR-146a treatment were significantly reversed by the
inhibition of TLR4, compared with in the levels in the
anti-miR-146a group (Fig. 9).
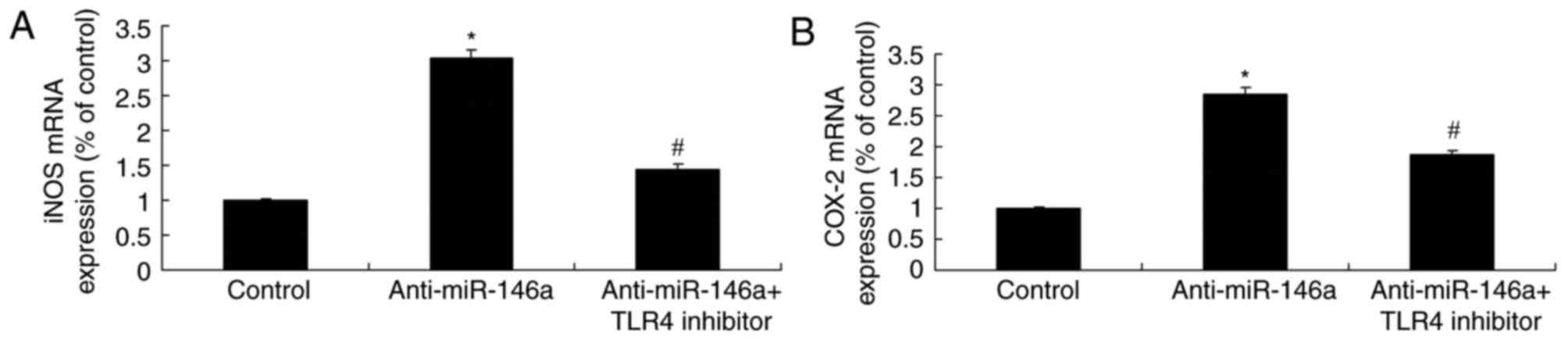
TLR4 inhibitor reduces the function of
anti-miR-146a on iNOS and COX-2 mRNA expression within a model of
ALI in 16HBE cells
The increase of iNOS and COX-2 mRNA expression
levels in the model of ALI induced by anti-miR-146a was
significantly suppressed by the inhibition of TLR4, compared with
the levels observed in the anti-miR-146a group (Fig. 10). MicroRNA-146a/TLR4 signaling
protects against severe burn-induced remote ALI in rats via
suppression of iNOS and COX-2 expression.
Discussion
Various mechanisms are involved in severe
burn-induced remote ALI. Firstly, damage to inflammatory mediators,
including proinflammatory cytokines and alexins, and lung tissues
occurs. Secondly, the increase of cell adhesion molecule expression
levels within neutrophils, granulocytes and endothelial cells
following stimulation by proinflammatory cells occurs, and adhesive
abilities increase (4).
Subsequently, neutrophils and granulocytes invade the inflammatory
lesions and, once activated, these cells undergo a respiratory
burst, degranulation and release metabolites, including
fibrinolysin, active oxygen and arachidonic acid. These molecules
may damage VECs, tissues and other cells (13). Thirdly, the adhesion of WBCs and
VECs, together with the accumulation of WBCs, may block
capillaries, leading to thrombosis, thus resulting in reduced
microcirculatory hemoperfusion (13). Consequently, a disproportional
ratio of ventilation/blood flow may occur, inducing hypoxemia
(14). In addition, the
permeability of pulmonary vasculatures may increase, causing
pulmonary edema. Fourth, the lung has the anatomical and
physiological features of low tension, low resistance and high
flow, and so circulatory inflammatory mediators may easily
accumulate within the lung (4). In
the present study, the expression of miR-146a was significantly
downregulated in a burn-induced rat model of ALI compared with the
levels in the control group and downregulation of miR-146a
increased inflammation in a model of ALI. MiR-146a downregulation
may worsen inflammation within the ALI model. Chen et al
(15) indicated that miR-146a may
regulate glucose-induced inflammation of the retina and kidney in
diabetes or animal/in vitro models.
Endogenous nitric oxide (NO) is a free radical with
high activity, and serves as a signaling molecule and a toxic
molecule with diverse biological functions (4). Excessive or deficient endogenous NO
may damage tissues and cells, and disrupt the equilibrium (4). It has been reported that iNOS itself
possesses the activity of oxygen radicals to induce O2
production (16). NO levels may
decrease once iNOS is inhibited, along with reduced O2
(4). In the present study,
upregulated miR-146a expression levels were associated with
decreased iNOS mRNA expression within the model of ALI.
Furthermore, Li et al (17). suggested that miR-146a may reduce
IL-6, IL-12, TNF-α and iNOS levels in systemic juvenile idiopathic
arthritis.
Expression of inflammatory factors may be initiated
and regulated following NF-κB activation (18). Epoxidase is an important
rate-limiting enzyme during the synthesis of prostaglandin and has
two isozymes, COX-1 and COX-2 (4).
NF-κB activation may induce COX-2 activity, resulting in increased
levels of the immune-suppressor prostaglandin (18), leading to immunosuppression
following infection (19).
Therefore, following immunosuppression, the incidence rates of
infection and mortality are high (18). To the best of our knowledge, the
present study is the first to reveal that upregulated miR-146a may
decrease COX-2 mRNA expression levels in a model of ALI. Sato et
al (20) reported that
miR-146a inhibition increased COX-2 expression in chronic
obstructive pulmonary disease.
Severe burn-induced remote ALI is an infectious lung
disease (21). It is predominantly
caused by bacterial infection-induced septicopyemia (4). LPS, the primary element of
gram-negative bacterium cytoderm, is the major pathogenic factor
(4,22). LPS may activate the signal
transduction system via an interaction with a corresponding
receptor. In addition, it may give rise to the activation of NF-κB,
initiating genetic transcription (4,23).
However, proinflammatory factors are produced to exert toxic
functions (4,23). TLR has been considered to be a
notable receptor that mediates LPS signal transduction (4). In the present study, miR-146a notably
suppressed TLR4/NF-κB signaling within a model of ALI. Ye and
Steinle (24) demonstrated that
miR-146a may attenuate inflammation via TLR4/NF-κB and TNF-α within
high glucose-induced primary human retinal microvascular
endothelial cells. In the present study, TLR4/NF-κB signaling was
analyzed; therefore, further studies into the effects of miR-146a
on NF-κB signaling in ALI must be conducted.
In summary, miR-146a may protect against severe
burn-induced remote ALI in rats. Specifically, miR-146a exerts
anti-inflammatory effects via the TLR4/NF-κB signaling pathway and
inhibits iNOS and COX-2 expression (Fig. 11). However, further investigation
of miR-146 for the treatment of severe burn-induced remote ALI is
required.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
JZ designed the experiment. JL and HY and XC
performed the experiment. JZ analyzed the data and wrote the
manuscript.
Ethics approval and consent to
participate
All experimental protocols were undertaken in
accordance with the Guide for the Care and Use of Laboratory
Animals by the National Institutes of Health, with the approval of
the Animal Experimental Ethics Committee of the First Affiliated
Hospital of Nanchang University (Nanchang, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tianzhu Z and Shumin W: Esculin inhibits
the inflammation of LPS-induced acute lung injury in mice via
regulation of TLR/NF-kappaB pathways. Inflammation. 38:1529–1536.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jiang S, Park DW, Tadie JM, Gregoire M,
Deshane J, Pittet JF, Abraham E and Zmijewski J: Human resistin
promotes neutrophil proinflammatory activation and neutrophil
extracellular trap formation and increases severity of acute lung
injury. J Immunol. 192:4795–4803. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferrando C, Aguilar G, Piqueras L, Soro M,
Moreno J and Belda FJ: Sevoflurane, but not propofol, reduces the
lung inflammatory response and improves oxygenation in an acute
respiratory distress syndrome model: A randomised laboratory study.
Eur J Anaesthesiol. 30:455–463. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bai C, Li T, Sun Q, Xu T, Yu J, Wang Y and
Wei L: Protective effect of baicalin against severe burn-induced
remote acute lung injury in rats. Mol Med Rep. 17:2689–2694.
2018.PubMed/NCBI
|
|
5
|
Fu PK, Wu CL, Tsai TH and Hsieh CL:
Anti-inflammatory and anticoagulative effects of paeonol on
LPS-induced acute lung injury in rats. Evid Based Complement
Alternat Med. 2012:8375132012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang H, Sun T, Liu Z, Zhang J, Wang X and
Liu J: Systemic inflammatory responses and lung injury following
hip fracture surgery increases susceptibility to infection in aged
rats. Mediators Inflamm. 2013:5364352013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shim DW, Han JW, Sun X, Jang CH, Koppula
S, Kim TJ, Kang TB and Lee KH: Lysimachia clethroides Duby extract
attenuates inflammatory response in Raw 264.7 macrophages
stimulated with lipopolysaccharide and in acute lung injury mouse
model. J Ethnopharmacol. 150:1007–1015. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fang Y, Gao F, Hao J and Liu Z:
microRNA-1246 mediates lipopolysaccharide-induced pulmonary
endothelial cell apoptosis and acute lung injury by targeting
angiotensin-converting enzyme 2. Am J Transl Res. 9:1287–1296.
2017.PubMed/NCBI
|
|
9
|
Cao Y, Lyu YI, Tang J and Li Y: MicroRNAs:
Novel regulatory molecules in acute lung injury/acute respiratory
distress syndrome. Biomed Rep. 4:523–527. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rajasekaran S, Pattarayan D, Rajaguru P,
Gandhi Sudhakar PS and Thimmulappa RK: MicroRNA regulation of acute
lung injury and acute respiratory distress syndrome. J Cell
Physiol. 231:2097–2106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bai X, Fan L, He T, Jia W, Yang L, Zhang
J, Liu Y, Shi J, Su L and Hu D: SIRT1 protects rat lung tissue
against severe burn-induced remote ALI by attenuating the apoptosis
of PMVECs via p38 MAPK signaling. Sci Rep. 5:102772015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jernigan PL, Hoehn RS, Blakeman TC, Heyl
J, Robinson BR, Pritts TA and Branson RD: Portable mechanical
ventilation with closed-loop control of inspired fraction of oxygen
maintains oxygenation in the setting of hemorrhage and lung injury.
J Trauma Acute Care Surg. 79:53–59. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raghavendran K, Davidson BA, Huebschmann
JC, Helinski JD, Hutson AD, Dayton MT, Notter RH and Knight PR:
Superimposed gastric aspiration increases the severity of
inflammation and permeability injury in a rat model of lung
contusion. J Surg Res. 155:273–282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen S, Feng B, Thomas AA and Chakrabarti
S: miR-146a regulates glucose induced upregulation of inflammatory
cytokines extracellular matrix proteins in the retina and kidney in
diabetes. PLoS One. 12:e01739182017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li X, Liu Z, Jin H, Fan X, Yang X, Tang W,
Yan J and Liang H: Agmatine protects against zymosan-induced acute
lung injury in mice by inhibiting NF-κB-mediated inflammatory
response. Biomed Res Int. 2014:5837362014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li D, Duan M, Feng Y, Geng L, Li X and
Zhang W: MiR-146a modulates macrophage polarization in systemic
juvenile idiopathic arthritis by targeting INHBA. Mol Immunol.
77:205–212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Quesada AE, Nguyen ND, Rios A and Brown
RE: Morphoproteomics identifies constitutive activation of the
mTORC2/Akt and NF-κB pathways and expressions of IGF-1R, Sirt1,
COX-2, and FASN in peripher lymphomas: Pathogenetic implications
and therapeutic options. Int J Clin Exp Pathol. 7:8732–8739.
2014.PubMed/NCBI
|
|
19
|
Kang SS, Cuendet M, Endringer DC, Croy VL,
Pezzuto JM and Lipton MA: Synthesis and biological evaluation of a
library of resveratrol analogues as inhibitors of COX-1, COX-2 and
NF-kappaB. Bioorg Med Chem. 17:1044–1054. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sato T, Liu X, Nelson A, Nakanishi M,
Kanaji N, Wang X, Kim M, Li Y, Sun J, Michalski J, et al: Reduced
miR-146a increases prostaglandin E2 in chronic obstructive
pulmonary disease fibroblasts. Am J Respir Crit Care Med.
182:1020–1029. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang H, Li Y, Huo P, Li XO, Kong D, Mu W,
Fang W, Li L, Liu N, Fang L, et al: Protective effect of
Jolkinolide B on LPS-induced mouse acute lung injury. Int
Immunopharmacol. 26:119–124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim HJ, Lee HS, Chong YH and Kang JL: p38
Mitogen-activated protein kinase up-regulates LPS-induced NF-kappaB
activation in the development of lung injury and RAW 264.7
macrophages. Toxicology. 225:36–47. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang G, Huang X, Li Y, Guo K, Ning P and
Zhang Y: PARP-1 inhibitor, DPQ, attenuates LPS-induced acute lung
injury through inhibiting NF-κB-mediated inflammatory response.
PLoS One. 8:e797572013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ye EA and Steinle JJ: miR-146a attenuates
inflammatory pathways mediated by TLR4/NF-κB and TNFα to protect
primary human retinal microvascular endothelial cells grown in high
glucose. Mediators Inflamm 2016: Article ID 3958453. 2016.
View Article : Google Scholar
|