Introduction
Although tremendous efforts have been obtained to
heighten early diagnosis and clinical outcomes of patients with
pancreatic cancer (PC), the overall prognosis and survival rate is
still very poor (1). Therefore, it
is urgent to reveal novel biomarkers for PC early diagnosis and
therapy.
MicroRNAs (miRNAs), are short (17–25 nucleotides)
non-coding RNAs that regulate gene expression by binding to
3′-untranslated regions (3′-UTR) sequence of target mRNAs (2). MiRNA-23a is a critical regulator in
carcinogenesis and aberrant miR-23a expression has been detected in
many cancers (3,4). Bioinformatic analysis of breast
cancer tissue-based miRNA panel shows that miR-23a possibly
functions as an oncogenic governor and promotes breast cancer
progression by directly activating transcription of Forkhead box
protein M1 (FOXM1) and Histidine-rich glycoprotein (HRG) (5). However, miR-23a also acts as a
negative regulator of oncogene in several kinds of cancer. It has
been reported that miR-23a is down-regulated in nephroblastomas
(6). In PC, miR-23a is
overexpressed using miRNA microarray-based analysis (7), suggesting that it can be used to as a
potential biomarker for diagnosis and treatment in PC. Although
some studies suggest that miR-23a functions as an oncogenic
regulator in PC (8), the detailed
roles and other molecular mechanisms remain to be revealed.
Polo-like kinase-1 (PLK-1) is a crucial mitotic
protein kinase that regulates multiple intracellular mitotic
processes including DNA replication, G2/M transition and
cytokinesis (9). PLK-1 is
overexpressed in various tumors and negatively correlated with a
wide spectrum of human cancers (10). Numerous studies reveal that there
are a great number of miRNAs which inhibit PLK-1 expression and
further repress oncogenesis, including non-small cell lung cancer
(11,12). Considering the important roles of
miRNAs and PLK-1, it is necessary to screen novel miRNAs targeting
PLK-1for PC diagnosis and treatment.
Herein, we found that PLK-1 was a potential target
of miR-23a in PC cells. Further research showed that miR-23a
restrained cell proliferation, invasion and migration in PC cell
lines, while promoted cell apoptosis possibly through inhibiting
PLK1 expression and its related signaling in vitro and in
vivo. Taken together, these findings might provide new sight
for early diagnosis and developing new therapeutic approaches for
PC.
Materials and methods
Clinical samples, cell lines and cell
transfection
A total of 20 cases of primary PC patients and their
paired adjacent non-tumor tissues were obtained from the Affiliated
Hangzhou Hospital of Nanjing Medical University. All procedures
performed in studies involving human participants were in
accordance with ethical standards of the institutional and/or
national research committee and with the 1964 Declaration of
Helsinki and its later amendments or comparable ethical standards.
Informed consent for publication was obtained from all individual
participants included in the present study. Human PC cell lines
hTERT-HPNE, MIA-PaCa-2, PANC-1, and Aspc-1were purchased from the
Cell Biology Institute of Shanghai (Shanghai, China). All cell
lines were cultured in DMEM medium supplemented with 10% fetal
bovine serum (FBS) and maintained at 37°C in a 5% CO2
atmosphere. To construct overexpression plasmid, we synthesized
sequences of human pre-miR-23a (miR23a-P1 (HpaI):
TATCACATTGCCAGGGATTTCCTTCAGAGAGGAAATCCCTGGCAATGTGATTTTTTTC,
miR23a-P2 (XhoI):
TCGAGAAAAAAATCACATTGCCAGGGATTTCCTCTCTTGAAGGAAATCCCTGGCAATGTGATA),
then inserted it into the linear plasmid PLL3.7. miR-23a inhibitors
and inhibitor NC were purchased from Guangzhou RiboBio Co., Ltd.,
(Guangzhou, China). Before transfection, cells were seeded at
24-well plates and transfected with blank, PLL-3-blank,
PLL-3-miR-23a, miR-23a-inhibitor, inhibitor NC using Lipofectamine™
2000 Transfection Reagent (cat. no. 11668-500; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) applying with 100 pmol
oligonucleotides in each well according to the manufacturer's
instructions.
Quantitative RT-PCR (qPCR)
Total RNA was isolated from the tissues and cells
utilizing TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The primers for qRT-PCR were indicated in Table I. GAPDH or U6 snRNA served as an
internal control for mRNA and miRNA expression, respectively, and
quantified relative to control using the delta-delta CT method.
 | Table I.Quantitative polymerase chain reaction
primer sequences. |
Table I.
Quantitative polymerase chain reaction
primer sequences.
| Gene | Sequences
(5′-3′) |
|---|
| miR-23a forward |
ATTGTATGTGGTCTCCGCTGTTTG |
| miR-23a reverse |
TTTCTTTTGGGTTGAGCCTTTTTT |
| U6 snRNA forward |
CAGCACATATACTAAAATTGGAACG |
| U6 snRNA reverse |
ACGAATTTGCGTGTCATCC |
| PLK-1 forward |
CCCACTGCCCGCCCAACCATTAAC |
| PLK-1 reverse |
CTCCTCTTGCCTGACCAGCCCACG |
| GAPDH forward |
CGGAGTCAACGGATTTGGTCGTAT |
| GAPDH reverse |
AGCCTTCTCCATGGTGGTGAAGAC |
Western blot analysis
Protein extraction from tissues and cells were
performed according to the manufacturer's instructions.
Immunoblotting was conducted using the following primary
antibodies: anti-PLK-1, Bax, Bcl2, cyclinB1, E-cadherin, Vimentin
(Abcam, Cambridge, USA), anti-GAPDH (Abcam), and the second
antibody horseradish peroxidase conjugated against mouse or rabbit
IgG (1:2,000 dilution; ZhongShan, Guangdong, China). Signals were
exposed to a film (FujiFilm, Tokyo, Japan) with ECL Plus (Beyotime
Institute of Biotechnology, Haimen, China) according to the
manufacturer's instructions.
Cell proliferation analysis
CCK8 Kit (YEASEN, Shanghai, China) was used to
analyze cell proliferation rate of cells with different treatments.
In brief, cells were suspended and seeded into 96-well plates,
after culturing for 24, 48 and 72 h. CCK8 was added into the medium
following the manufacturer's instruction. The absorbance was
detected at 450 nm. Cell proliferation rate was presented with the
value relative to control group.
Migration and matrigel invasion
assays
We added suspended and stained (Cell-Tracker-Red;
Invitrogen; Thermo Fisher Scientific, Inc.) MIA-PaCa-2 cells on a
confluent HMC monolayer, which was stained with the fluorescence
dye Cell-Tracker-Green (Invitrogen; Thermo Fisher Scientific,
Inc.). 24 h for migration analysis, and 48 h for invasion
analysis.
Cell apoptosis assay
Cells were grown in 6-well plates to about 50–60%
confluence and transiently transfected with blank, PLL3-blank,
PLL-3-miR-23a, miR-23a-inhibitor and inhibitor NC respectively.
Cells were trypsinized and collected at 48 h post-transfection
before washing with PBS twice, treated by Annexin V-EGFP Apoptosis
Detection kit according to the manufacturer's instructions (Nanjing
KeyGen Biotech., Co., Ltd., Jiangsu, China), and analyzed with a
flow cytometer (FACS calibur; BD Biosciences, Franklin Lakes, NJ,
USA).
Luciferase reporter assay
PLK1-3′UTR-WT and PLK1-3′UTR-mut plasmids were
obtained from GeneChem, Inc., (Daejeon, Korea). Cells were
cotransfected with non-relative control RNA duplex and
PLK1-3′UTR-WT, or cotransfected with non-relative control RNA
duplex and PLK1-3′UTR-mut plasmids were served as control groups.
Cells cotransfected with PLL3-miR23a and PLK1-3′UTR-WT or
PLK1-3′UTR-mut were defined as case groups. The pRL-TK (Promega
Corporation, Madison, WI, USA) was used as a normalization control.
Cells were collected at 24 h after transfection, and luciferase
activity was measured using a dual-luciferase reporter assay kit
(Promega Corporation) and recorded by chemiluminescence meter
(Promega Corporation).
In vivo tumorigenicity assay
All experiments were carried out under the ethical
approval of Ethics Committee for Animal Experimentation of The
Affiliated Hangzhou Hospital of Nanjing Medical University.
Four-week-old female BALB/c nude mice were purchased from ShangHai
and maintained in specific pathogen-free conditions. To build the
subcutaneous human MIA-PaCa-2 tumor xenograft model, MIAPaca-2
cells were subcutaneously inoculated (5×106 cells
suspended in a 1:1 mixture of serum free-Dulbecco's modified
Eagle's medium/Matrigel (BD Biosciences per site); total volume 0.1
ml) bilaterally into the flanks of athymic mice. When tumors were
grown at approximately 100 mm3, mice were injected with
miR-23a agomir or agomir NC (Guangzhou RiboBio Co., Ltd.) following
the manufacturer's recommendation. The animals were monitored for
activity, physical condition, determination of body weight, and
measurement of tumor volume (1/2 × length × width2) were
made twice a week. After 4 weeks of treatment, tumors were
harvested and weighed. To measure the tumors accurately, mice were
rendered unconscious with and digital calipers were used to measure
the tumor size over time.
Statistical analysis
All data were obtained from at least three
independent experiments, and presented as the mean ± standard
deviation. Datasets with only two groups were analyzed using a
Student's t-test. Differences between multiple groups were analyzed
using one-way analysis of variance with the Tukey-Kramer post-hoc
test. P<0.05 was considered to indicate a statistically
significant difference. All of the statistical calculations were
performed using the SPSS software v15.0 (SPSS, Inc., Chicago, IL,
USA).
Results
PLK-1 acts as a potential target of
miR-23a in human PC cells
Sequence analyses of the mature sequence of miR-23a
showed that miR-23a could be the potential functional regulator of
3′UTR of PLK mRNA. To assess whether PLK1 was a potential target of
miR-23a, we performed qPCR to detect expression levels of miR-23a
and PLK-1 in three human PC cell lines (PANC-1, MIA-PaCa-2 and
Aspc-1) and one human pancreatic ductal epithelial cells
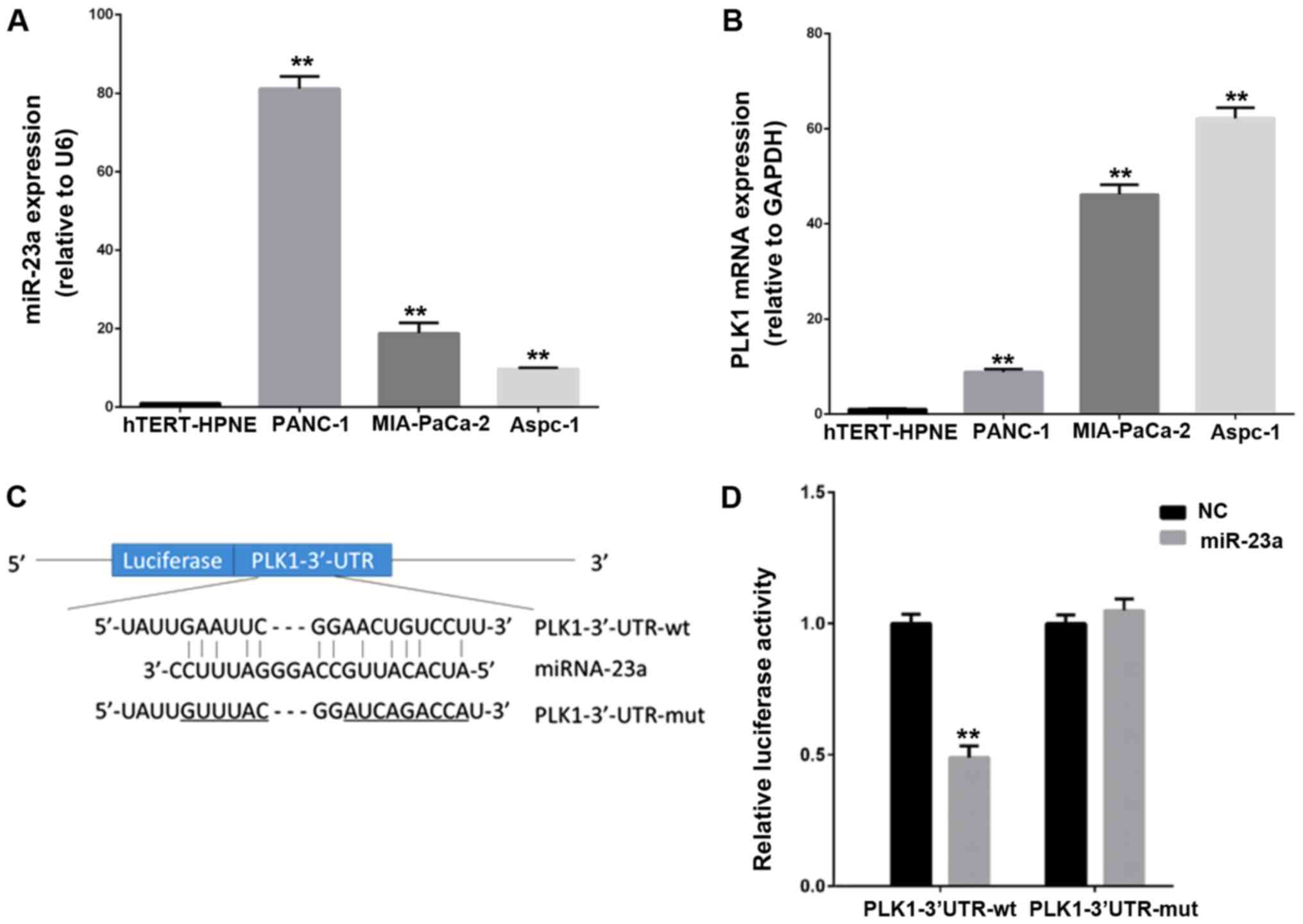
hTERT-HPNE. As shown in Fig. 1A and
B, the expression level of miR-23a and PLK-1 mRNA were
significantly increased in three PC lines compared to hTERT-HPNE
cell. However, the expression pattern of miR-23a and PLK-1 showed a
reverse trend in three PC cell lines. For example, compared with
MIA-PaCa-2 and Aspc-1 cell lines, the highest expression of miR-23a
was shown in PANC-1 cells, while PLK-1 level was lowest in PANC-1
cells. To further verify the direct relationship between miR-23a
and PLK1, we constructed PLK1-3′UTR-WT and PLK1-3′UTR-mut plasmids
to perform double luciferase reporter assay in MIA-PaCa-2 cell
line. Results indicated that up-regulation of miR-23a dramatically
repressed the luciferase activity by 50% when cotransfected with
PLK-1 3′UTR-WT in PC cells, but there was no change in the PLK-1
3′UTR-mut group (Fig. 1C and D).
Therefore, these findings suggest that PLK-1 is a potential target
of miR-23a in PC cells.
Overexpression of miR-23a suppresses
the proliferation, migration, and invasion of MIA-PaCa-2 cells
We further explored whether miR-23a could affect
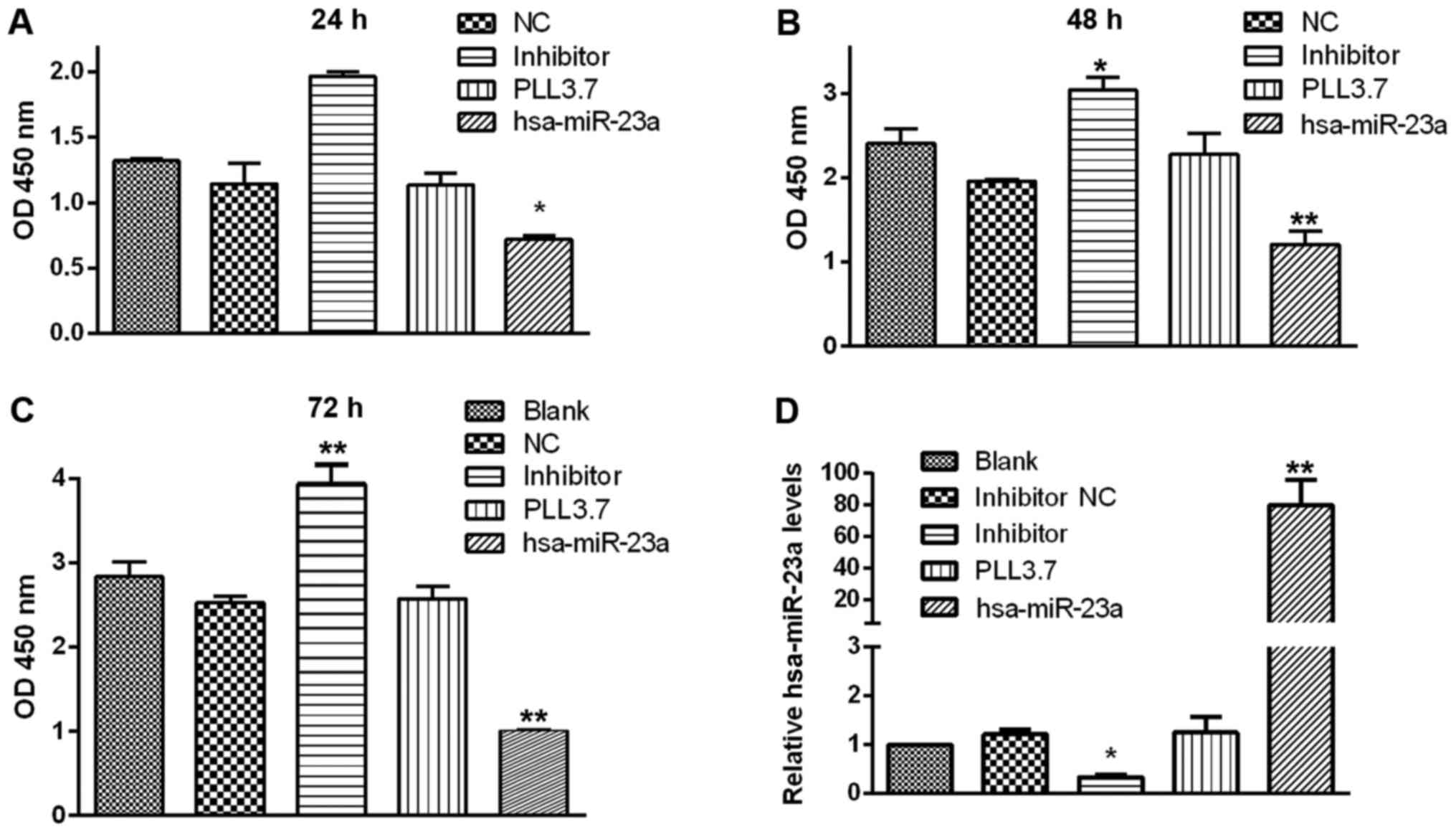
biological functions of PC cells. Cell proliferation assay
suggested that miR-23a overexpression significantly suppressed the
proliferation of PC cell at 24~48 h post transfection, while
transfection with miR-23a inhibitor moderately enhanced the
multiplication capacity of PC cells compared with control groups
(Fig. 2). According to the
statistical analysis indicated that MIA-PaCa-2 cell growth rate was
the lowest in miR-23a transfection group compared with the control
groups, and the highest in group transfected with miR-23a inhibitor
after transfection for 24, 48, and 72 h. However, there was no
significant change among the three control groups at any point in
time (Fig. 3A-C).
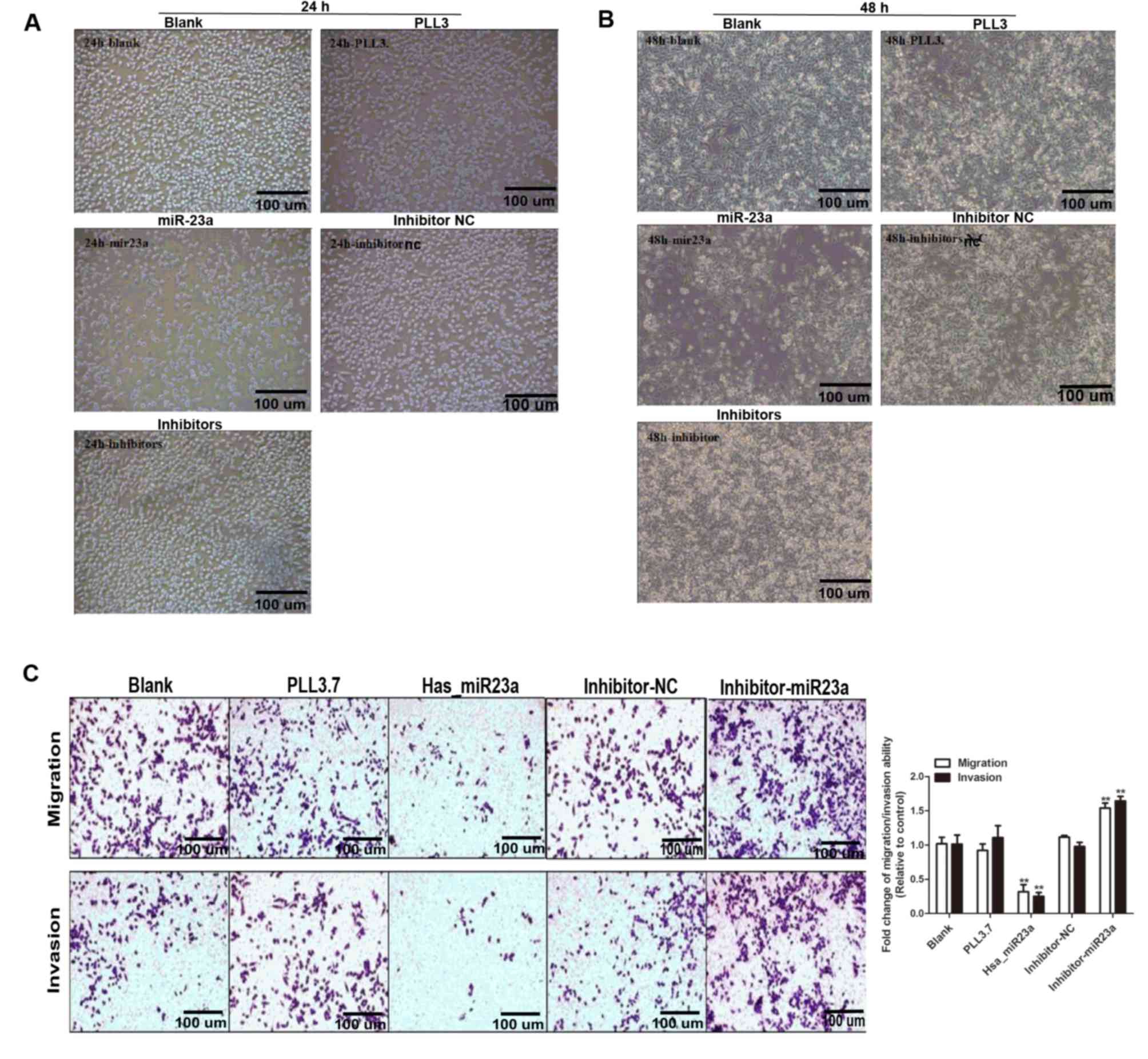
We further investigated the effects of miR-23a on
the capability of migration and invasion in MIA-PaCa-2 cells. As
shown in Fig. 2C, miR-23a
overexpression prominently reduced cell migration compared with
blank and PLL3.7 group, while miR-23a inhibitor increased the
migration ability of MIA-PaCa-2 cells. Similar results were
acquired in cell invasion assay (Fig.
2C, lower panel). Furthermore, transfection efficiency was also
verified through detecting the transcriptional level of has-miR-23a
to support our findings (Fig. 3D).
Taken together, we demonstrate that miR-23a acts as a tumor
suppressor in PC cells.
Increased expression of miR-23a
induces PC cell apoptosis probably through modulating PLK-1
We sought to examine whether miR-23a could inhibit
cell growth by modulating cellular apoptosis. Flow cytometry assays
showed that miR-23a overexpression promoted cell apoptosis
significantly (52.22% vs. 4.92%), while transfection with miR-23a
inhibitor decreased cell apoptotic rate compared with NC group
(4.32% vs. 10.68%) (Fig. 4A). To
investigate the underlying mechanism of proliferation inhibition
and apoptosis promotion induced by miR-23a, the expression level of
several proliferation and apoptosis related proteins, including
oncogene PLK-1, apoptotic activator Bcl-2-associated X protein
(Bax), Bcl-2, tumor suppressor E-cadherin, cell cycle-associated
protein Cyclin B1 and Vimentin were determined, respectively. As
expected, the protein expression of PLK-1 was decreased or
increased in cells transfected with miR-23a mimic or inhibitor,
respectively (Fig. 4B). Moreover,
miR-23a overexpression inhibited Bcl-2, Cyclin B1, and Vimentin
expression, while promoted Bax and E-cadherin expression, while
opposite effects were observed in cells transfected with miR-23a
inhibitor. Therefore, our findings demonstrate that miR-23a
mediates cell apoptosis probably by targeting PLK-1 and regulating
downstream effectors.
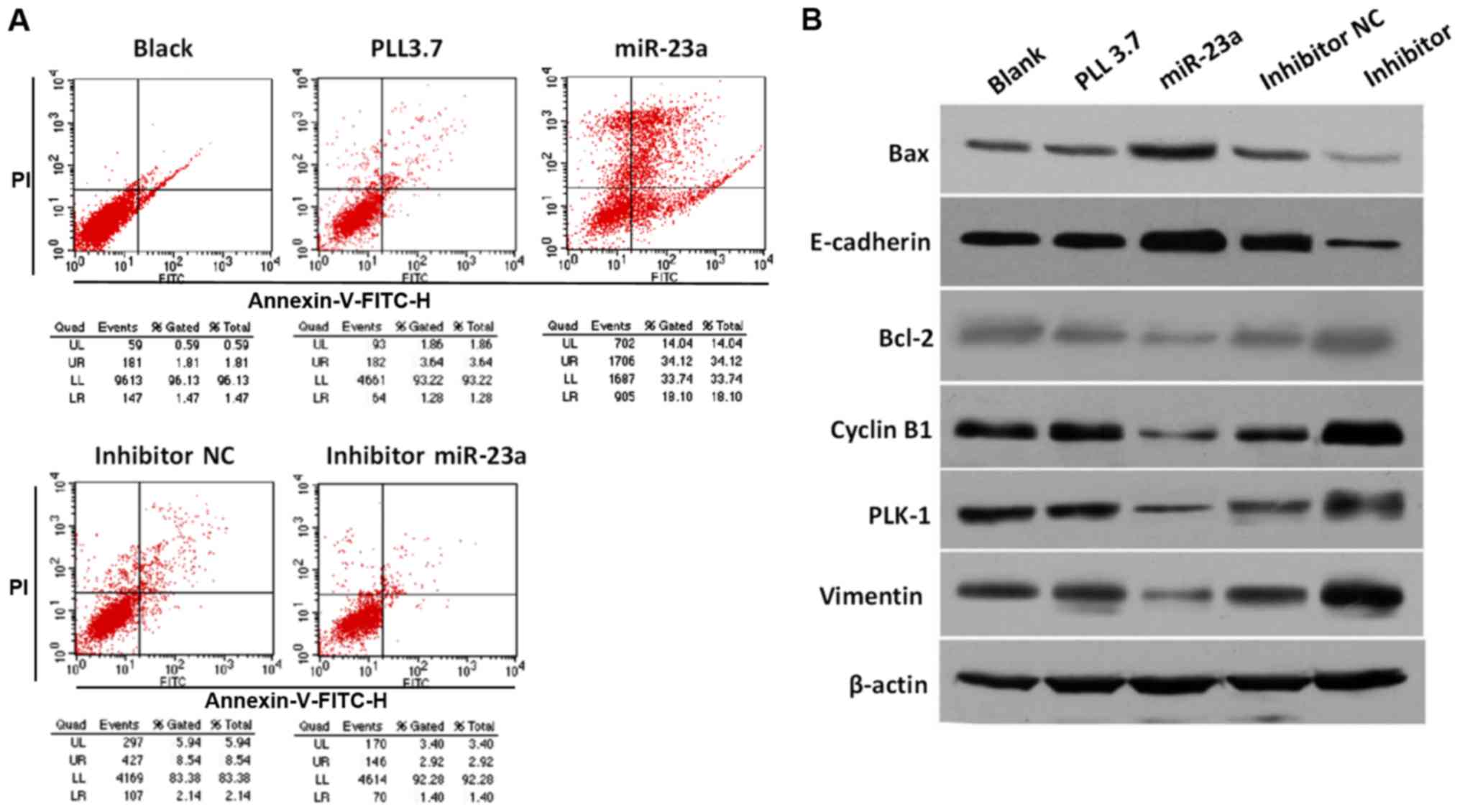 | Figure 4.PC cell apoptosis induced by miR-23a
and multiple signaling molecules expression. (A) Flow cytometry
based apoptosis assay for PC cells with different treatment. (B)
Protein expression of PLK1, Vimentin, Cyclin B1, Bcl-2, E-cadherin,
and Bax proteins in MIA-PaCa-2 cells transfected with different
plasmids were determined by western blotting assay with specific
antibodies. PC, pancreatic cancer; miR, microRNA; Bcl, B-cell
lymphoma; PLK, polo-like kinase-1; NC, negative control. |
MiR-23a inhibits PC tumor growth in
vivo
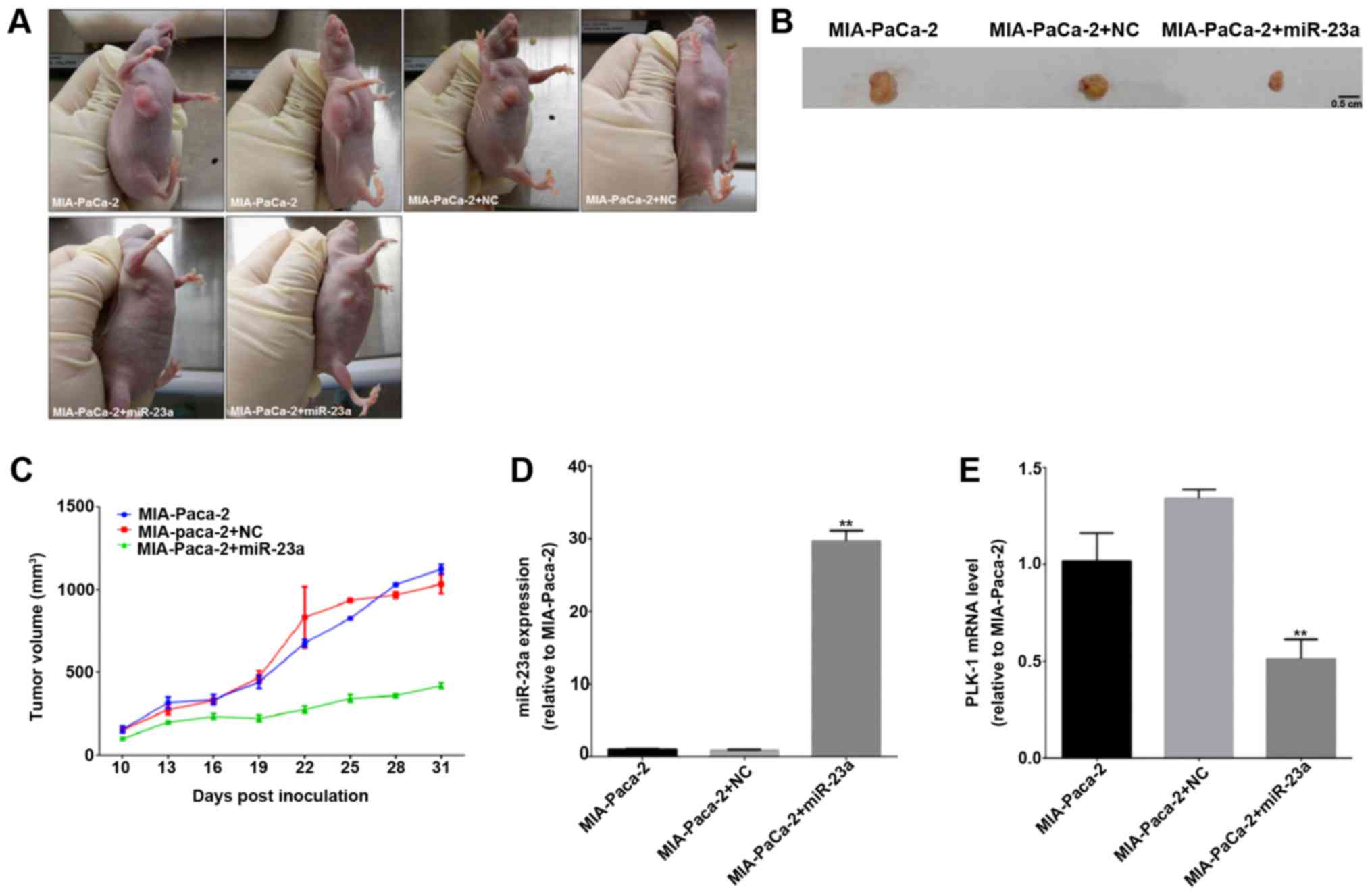
Furthermore, we tried to observe the critical
anti-carcinoma role of miR-23a in vivo. As indicated in
Fig. 5A, the tumor masses
distinctly diminished in group of MIA-PaCa-2 cells injected with
miR-23a agomir compared to control and negative control groups at
24 days after initial injection. We also stripped out the neoplasm
at 31 days post-injection, the result showed that tumor size was
the lowest in miR-23a agomir-treated group than control groups
(Fig. 5B). Moreover, according to
the statistic analyses of tumor growth curve, we could see from
Fig. 5C, control and negative
control group presented a similar increasing tendency, but
up-regulation of miR-23a obviously slowed tumor growth. We further
explored the transcriptional levels of miR-23a and PLK-1 of tumor
mass in each group, and found PLK-1 expression was significantly
suppressed in miR-23a agomir-treated group (Fig. 5D), and these data further verified
the essential inhibition of miR-23a on cell proliferation in
vivo.
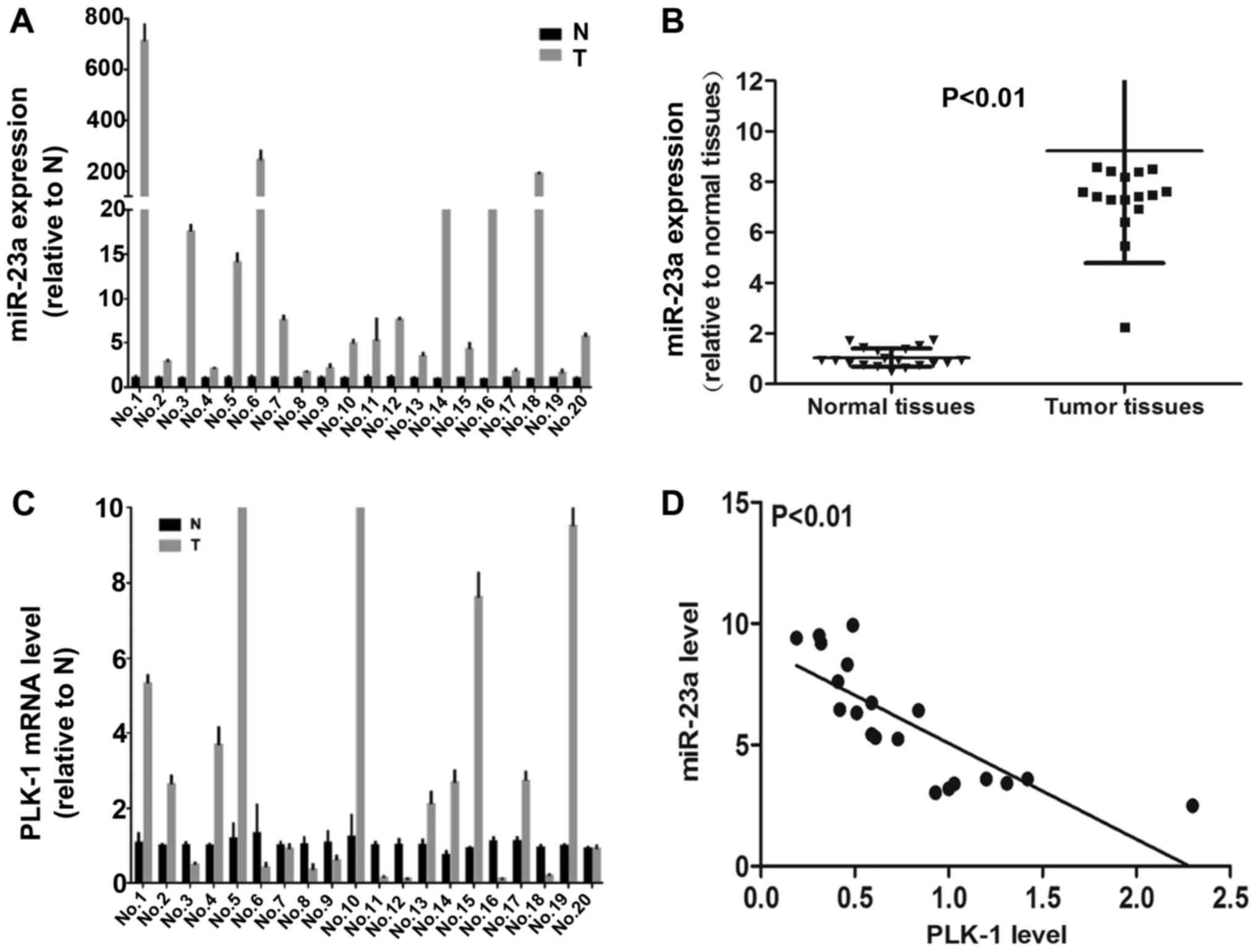
Mir-23a is highly expressed in human
PC tissues
To further determine the correlation between miR23a
and PLK-1in human PC samples, we examined transcriptional levels of
miR-23a and PLK-1 in 20 cases of primary PC patients and their
paired adjacent non-tumor tissues by using qPCR. Among 20 pairs of
PC patient tissue samples, 19 (95%) cases showed remarkable
up-regulation of miR-23a in cancer tissues (Fig. 6A and B). However, miR-23a
expression was negatively correlated with tumor grade, tumor size
and lymph node metastasis (Table
II). As shown in Fig. 6C, 10
(50%) cases showed up-regulation of PLK-1 in cancer tissues
compared with non-tumor tissues, indicating its high expression was
potential correlation with the progression of PC. Conversely, 10
(50%) cases presented down-regulation of PLK-1 in tumor samples
compared to non-tumor tissues. Importantly, pearson's correlation
coefficient analysis indicated that miR-23a and PLK-1 expression
was negatively correlated in PC tissues (Fig. 6D). Thus, these findings in human PC
tissues present some contradictions with our in vitro
data.
| Table II.Association between microRNA-23a
expression and clinical pathological characteristics of 20 patients
with pancreatic cancer. |
Table II.
Association between microRNA-23a
expression and clinical pathological characteristics of 20 patients
with pancreatic cancer.
| Variables | n | Low miR-23a (%) | High miR-23a (%) | P-value |
|---|
| Age |
|
|
| >0.500 |
|
>40 | 12 | 5 | 7 |
|
| ≤40 | 8 | 4 | 4 |
|
| Sex |
|
|
| >0.500 |
| Male | 13 | 8 | 5 |
|
|
Female | 7 | 4 | 3 |
|
| T grade of tumor |
|
|
| 0.012 |
| T1 | 5 | 4 (80.0) | 1 (20.0) |
|
| T2 | 6 | 4 (66.7) | 2 (33.3) |
|
| T3 | 4 | 3 (75.0) | 1 (25.0) |
|
| T4 | 5 | 4 (83.3) | 1 (16.7) |
|
| Tumor size |
|
|
| 0.015 |
| <6
cm | 12 | 9 (75.0) | 3 (25.0) |
|
| >6
cm | 8 | 6 (75.0) | 2 (25.0) |
|
| Lymph node
metastasis |
|
|
| 0.005 |
|
Negative | 9 | 6 (66.7) | 3 (33.3) |
|
|
Positive | 11 | 8 (72.8) | 3 (27.2) |
|
Discussion
Many efforts have been made to define it as a
therapeutic drug target in PC (13). However, due to the lack of
specificity and high toxicity, PLK1-specific inhibitors are limited
in clinical applications (14).
Hence, the essential regulator role of PLK-1 in cancer still drive
scientists to find novel targeted miRNAs which can be taken as a
critical biomarker and targeted for drug discovery.
Here, we verified the expression pattern of miR-23a
and PLK-1, and revealed the negative correlation between them in
human PC cell lines (PANC-1, MIA-PaCa-2 and Aspc-1). Our data
illustrated miR-23a acted as a negative regulator of PLK-1 in PC
cells. Our investigation demonstrated that miR-23a overexpression
in MIA-PaCa-2 suppressed cell growth, as well as increased cell
apoptosis. Xenograft experiment also verified the inhibition of
miR-23a in tumor growth. Even though previous studies have
confirmed that PLK-1 is associated with the development and
progression of pancreatic carcinoma (15), the understanding of upstream and
downstream regulators for PLK-1 are very poor. Moreover, the
underlying mechanism of how PLK1 interaction with cell
proliferation and apoptosis pathway in PC remains to be understood
and needs further investigation. Our findings revealed a novel
regulator of PLK-1 that could ameliorate the oncogenesis of PC
through disturbing PLK-1 expression. Additionally, we found that
up-regulation of miR-23a promoted the expression of
Bcl-2-associated X protein (Bax) and E-Cadherin, suppressed the
expression of Cyclin B1, Vimentin, and Bcl-2. Possibly, our
findings provide a novel regulation mechanism of miR-23a by
possibly targeting PLK-1 targeting PLK-1 in PC progression.
Paradoxically, we and other researchers confirmed
that miR-23a was obviously up-regulation in human PC tissues
compared to normal pancreatic samples (7), indicating that miR-23a might act as
an oncogenic regulator in PC patients. However, it can also
function as a tumor suppressor in a few kinds of tumor. For
example, previous study has suggested that miR-23a could enhance
migration and invasion by down-regulation of PTEN in osteosarcoma
(16). Some researchers indicated
that miR-23a showed lower expression in osteosarcoma specimens and
functioned as a tumor suppressor (17). Our observations demonstrated that
miR-23a was overexpressed in PC patients, but miR-23a
overexpression could inhibit PC progression in vitro. We
thought there were three reasons: 1) The heterogeneity of miR-23a
and PLK-1 expression between human PC samples and PC cells is due
to that the in vivo tumor microenvironment was a highly
complex and situational scene resulting in the diversity of signal
molecule expression. 2) Possibly, miR-23a was indeed up-regulation
in human PC and functioned as an oncogenic regulator by targeting
kinds of tumor suppressor genes. However, the role of miR-23a on
mediating the down-regulation of PLK-1 was covered up by its
oncogenic role in human PC patients. 3) Considering that our
statistical sample was extremely limited, a large number of samples
need to be taken to obtain a meaningful result. Thus, miR-23a might
play different roles and functions based on individual differences.
Probably, miR-23a is an oncogene by modulating various tumor
suppressors in PC. miR-23a functions as anti-oncogene through
probably restraining the expression of oncogene PLK-1 and its
downstream signaling pathway.
In conclusion, we demonstrated the inhibitory roles
of miR-23a possibly by targeting PLK-1 in vitro and in
vivo. We were confident that miR-23a could inhibit PC
progression in some patients or some PC cell lines by affecting the
expression of PLK-1. However, the detailed mechanism and the
possibility of the clinical application of miR-23a in PC still need
to be investigated.
Acknowledgements
Not applicable.
Funding
This work was supported by Special Disease and
Specialty Construction Project of Technology Bureau of Hangzhou
(20150733Q15) and ChiaTai Qingchunbao cancer research projects
(2015ZYC-A02).
Availability of data and materials
All data generated or analysed during this study are
included in this published article.
Authors' contributions
JW and YM designed the study. AZ and LT analyzed the
data. BC, YX, XCL, YPP and JJZ performed the experiments. BC and AZ
wrote the paper.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Review Board of the Affiliated Hangzhou Hospital of Nanjing Medical
University. Written informed consent was obtained from all patients
prior to their inclusion within the study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Subramani R, Gangwani L, Nandy SB,
Arumugam A, Chattopadhyay M and Lakshmanaswamy R: Emerging roles of
microRNAs in pancreatic cancer diagnosis, therapy and prognosis
(Review). Int J Oncol. 47:1203–1210. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yong FL, Wang CW, Roslani AC and Law CW:
The involvement of miR-23a/APAF1 regulation axis in colorectal
cancer. Int J Mol Sci. 15:11713–11729. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma G, Dai W, Sang A, Yang X and Gao C:
Upregulation of microRNA-23a/b promotes tumor progression and
confers poor prognosis in patients with gastric cancer. Int J Clin
Exp Pathol. 7:8833–8840. 2014.PubMed/NCBI
|
|
5
|
Eissa S, Matboli M and Shehata HH: Breast
tissue-based microRNA panel highlights microRNA-23a and selected
target genes as putative biomarkers for breast cancer. Transl Res.
165:417–427. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koller K, Das S, Leuschner I, Korbelius M,
Hoefler G and Guertl B: Identification of the transcription factor
HOXB4 as a novel target of miR-23a. Genes Chromosomes Cancer.
52:709–715. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Piepoli A, Tavano F, Copetti M, Mazza T,
Palumbo O, Panza A, di Mola FF, Pazienza V, Mazzoccoli G, Biscaglia
G, et al: Mirna expression profiles identify drivers in colorectal
and pancreatic cancers. PLoS One. 7:e336632012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu N, Sun YY, Zhang XW, Chen S, Wang Y,
Zhang ZX, Song SW, Qiu GB and Fu WN: Oncogenic miR-23a in
pancreatic ductal adenocarcinogenesis via inhibiting APAF1. Dig Dis
Sci. 60:2000–2008. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu X: Targeting polo-like kinases: A
promising therapeutic approach for cancer treatment. Transl Oncol.
8:185–195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Strebhardt K: Multifaceted polo-like
kinases: Drug targets and antitargets for cancer therapy. Nat Rev
Drug Discov. 9:643–660. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu J, Lu KH, Liu ZL, Sun M, De W and Wang
ZX: MicroRNA-100 is a potential molecular marker of non-small cell
lung cancer and functions as a tumor suppressor by targeting
polo-like kinase 1. BMC Cancer. 12:5192012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu C, Li S, Chen T, Hu H, Ding C, Xu Z,
Chen J, Liu Z, Lei Z, Zhang HT, et al: miR-296-5p suppresses cell
viability by directly targeting PLK1 in non-small cell lung cancer.
Oncol Rep. 35:497–503. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang C, Sun X, Ren Y, Lou Y, Zhou J, Liu
M and Li D: Validation of Polo-like kinase 1 as a therapeutic
target in pancreatic cancer cells. Cancer Biol Ther. 13:1214–1220.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raab M, Kappel S, Krämer A, Sanhaji M,
Matthess Y, Kurunci-Csacsko E, Calzada-Wack J, Rathkolb B, Rozman
J, Adler T, et al: Toxicity modelling of Plk1-targeted therapies in
genetically engineered mice and cultured primary mammalian cells.
Nat Commun. 2:3952011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weichert W, Schmidt M, Jacob J, Gekeler V,
Langrehr J, Neuhaus P, Bahra M, Denkert C, Dietel M and Kristiansen
G: Overexpression of Polo-like kinase 1 is a common and early event
in pancreatic cancer. Pancreatology. 5:259–265. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tian K, Di R and Wang L: MicroRNA-23a
enhances migration and invasion through PTEN in osteosarcoma.
Cancer Gene Ther. 22:351–359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang G, Li B, Fu Y, He M, Wang J, Peng S
and Bai L: miR-23a suppresses proliferation of osteosarcoma cells
by targeting SATB1. Tumour Biol. 36:4715–4721. 2015. View Article : Google Scholar : PubMed/NCBI
|