Introduction
Osteosarcoma is the most common primary malignant
bone tumor among children and adults worldwide and is characterized
by a high metastatic potential to the lung as well as poor
prognosis (1,2). Long non-coding (lnc)RNAs are
sequences of non-coding protein transcripts (>200 nucleotides in
length) with little or no protein-coding capacity (3). Unlike mRNAs, lncRNAs are associated
with numerous biological and cellular processes, including the
regulation of transcription and mRNA post-transcriptional levels
(4,5). Previous studies have revealed that
lncRNAs are responsible for initiation and progression of numerous
cancers due to their involvement in cellular processes (6–8).
Notably, a number of functional lncRNAs have previously been
revealed to be associated with osteosarcoma, including HOX
transcript antisense RNA (AS) (9),
breast cancer anti-estrogen resistance 4 (10), HOXA distal transcript AS (11) and urothelial cancer associated 1
(12). Numerous lncRNAs function
as the antisense partners of their respective protein-coding genes,
including cyclin dependent kinase inhibitor 2B-AS1 (13,14),
C-terminal binding protein 1-AS (15), hepatocyte nuclear factor 1 homeobox
A-AS1 (16) and proliferation
arrest specific 6-AS1 (17).
LncRNA WW domain-containing oxidoreductase
(WWOX)-AS1 is an antisense transcript of WWOX (18). Furthermore, WWOX has been
demonstrated to be strongly associated with numerous cancer types,
including ovarian cancer (19),
hepatocellular carcinoma (20),
colon cancer (21) and bladder
cancer (22). In addition, the
expression levels and functions of antisense lncRNAs are frequently
associated with their corresponding protein-coding genes (13–17).
It has previously been demonstrated that lncRNA WWOX-AS1 is
associated with ovarian cancer and that its overexpression inhibits
the proliferation, migration and invasion of epithelial ovarian
cancer cells (18). Furthermore,
previous studies have also revealed that WWOX expression is
associated with osteosarcoma and that its overexpression inhibits
the progression of osteosarcoma (23,24).
Therefore, it may be suggested that the expression and function of
WWOX-AS1 are associated with the progression and prognosis of
osteosarcoma.
In the present study, a number of experiments were
performed to investigate the functional role of WWOX-AS1 in
osteosarcoma. The expression of WWOX-AS1 in osteosarcoma tissues,
its association with patient prognosis and its correlation with
WWOX expression levels were investigated. Furthermore,
functional assays were performed via the knockdown and
overexpression experiments. In conclusion, the results of the
present study demonstrated the functional role of WWOX-AS1 in
osteosarcoma, which may prove valuable for future osteosarcoma
research.
Materials and methods
Study participants
A total of 70 patients with osteosarcoma were
recruited in the present study (Table
I). The 70 patients with osteosarcoma were classified into two
groups: High-WWOX-AS1 group (n=35; WWOX-AS1 expression value
≥median value) and low-WWOX-AS1 group (n=35; WWOX-AS1 expression
value ≤median value). Staging is based on Enneking system staging
(25). From each patient, an
osteosarcoma tissue sample and an adjacent tissue sample were
collected via appropriate surgical resections. The samples were
collected between March 2011 and December 2014 in the General
Hospital of the People's Liberation Army (Beijing, China). Written
informed consent was obtained from each patient or their guardians.
The present study was approved by the Ethics Committees of the
General Hospital of the People's Liberation Army.
 | Table I.Characteristics of patients with
osteosarcoma participating in the present study. |
Table I.
Characteristics of patients with
osteosarcoma participating in the present study.
| Number | Age (years) | Sex | Stage | Tumor location |
|---|
| 1 | 16 | M | III A | Proximal end of
tibia |
| 2 | 27 | F | III A | Proximal end of
tibia |
| 3 | 41 | F | III B | Proximal end of
humerus |
| 4 | 40 | F | I B | Proximal end of
humerus |
| 5 | 40 | F | II A | Distal end of
humerus |
| 6 | 15 | F | II B | Proximal end of
tibia |
| 7 | 19 | M | II A | Proximal end of
tibia |
| 8 | 42 | F | II B | Proximal end of
tibia |
| 9 | 11 | M | II A | Proximal end of
tibia |
| 10 | 46 | F | I A | Proximal end of
femur |
| 11 | 29 | M | II | Distal end of
humerus |
| 12 | 15 | M | II A | Proximal end of
tibia |
| 13 | 15 | M | II A | Proximal end of
tibia |
| 14 | 10 | M | I A | Distal end of
femur |
| 15 | 12 | F | II B | Proximal end of
tibia |
| 16 | 44 | M | II B | Proximal end of
ulna |
| 17 | 40 | F | II A | Distal end of
femur |
| 18 | 41 | M | I A | Proximal end of
tibia |
| 19 | 43 | F | I B | Distal end of
femur |
| 20 | 19 | F | II B | Proximal end of
femur |
| 21 | 32 | M | I A | Distal end of
femur |
| 22 | 40 | F | II B | Distal end of
femur |
| 23 | 14 | M | II B | Proximal end of
humerus |
| 24 | 41 | M | III A | Proximal end of
humerus |
| 25 | 17 | M | III A | Distal end of
tibia |
| 26 | 12 | M | II B | Proximal end of
tibia |
| 27 | 42 | F | I B | Proximal end of
tibia |
| 28 | 14 | F | I B | Distal end of
femur |
| 29 | 21 | F | I B | Distal end of
femur |
| 30 | 20 | M | II B | Proximal end of
tibia |
| 31 | 19 | F | II B | Proximal end of
femur |
| 32 | 47 | F | III A | Distal end of
femur |
| 33 | 8 | M | I B | Distal end of
femur |
| 34 | 47 | M | III A | Distal end of
femur |
| 35 | 43 | F | II B | Distal end of
ulna |
| 36 | 15 | M | III B | Distal end of
tibia |
| 37 | 41 | F | I A | Proximal end of
humerus |
| 38 | 40 | F | II A | Distal end of
ulna |
| 39 | 41 | F | I B | Distal end of
ulna |
| 40 | 45 | F | III B | Proximal end of
tibia |
| 41 | 55 | M | I A | Proximal end of
tibia |
| 42 | 50 | F | I B | Distal end of
tibia |
| 43 | 12 | M | III A | Distal end of
femur |
| 44 | 18 | F | III A | Proximal end of
tibia |
| 45 | 15 | M | II A | Proximal end of
tibia |
| 46 | 16 | F | I B | Distal end of
femur |
| 47 | 43 | F | II B | Distal end of
tibia |
| 48 | 28 | M | II B | Distal end of
tibia |
| 49 | 40 | F | III A | Proximal end of
femur |
| 50 | 10 | M | I B | Proximal end of
tibia |
| 51 | 48 | M | II A | Proximal end of
femur |
| 52 | 42 | F | I B | Proximal end of
femur |
| 53 | 18 | M | II A | Distal end of
femur |
| 54 | 20 | M | I A | Proximal end of
femur |
| 55 | 45 | F | I B | Proximal end of
tibia |
| 56 | 45 | F | II A | Distal end of
femur |
| 57 | 43 | F | III A | Distal end of
femur |
| 58 | 15 | M | I A | Proximal end of
tibia |
| 59 | 18 | M | III A | Distal end of
femur |
| 60 | 42 | M | III A | Distal end of
femur |
| 61 | 43 | M | I B | Distal end of
femur |
| 62 | 16 | M | III A | Distal end of
tibia |
| 63 | 50 | M | I B | Distal end of
femur |
| 64 | 42 | F | III A | Proximal end of
tibia |
| 65 | 15 | M | I B | Proximal end of
humerus |
| 66 | 15 | F | I B | Distal end of
femur |
| 67 | 16 | F | I A | Distal end of
femur |
| 68 | 18 | M | I B | Distal end of
femur |
| 69 | 49 | M | II A | Proximal end of
tibia |
| 70 | 11 | M | II A | Distal end of
femur |
Cancer cell lines
Three human osteosarcoma cell lines (SAOS2, MG63 and
U2OS) and a normal human osteoblastic cell line (hFOB) were
purchased from the American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's Modified Eagle's Medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal bovine serum (FBS; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and 1%
penicillin/streptomycin solution (Invitrogen, Thermo Fisher
Scientific, Inc.) at 37°C and 5% CO2.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from patients' tissues and
cell lines using TRIzol Total RNA Reagent (Life Technologies;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Complementary (c)DNA was synthesized from 2 µg total RNA
using the RevertAid™ H Minus First Strand cDNA Synthesis kit
(Takara Bio, Inc., Otsu, Japan) and cDNA was stored at −20°C. The
RT reactions were terminated by inactivation at 95°C for 10 min and
stored at 4°C. Primers used were obtained from Chang Jing Bio-Tech,
Ltd. (Changsha, China) and were identical to those used in our
previously published study (18);
they are presented in Table II.
qPCR was performed using the SYBR PrimeScript RT-PCR kit (Takara
Bio, Inc.) in an Applied Biosystems 7500 Fluorescent Quantitative
PCR System (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The thermocycling conditions used were as follows: 95°C for 30 sec;
followed by 40 amplification cycles at 95°C for 5 sec and 60°C for
34 sec. The values were normalized to the internal control products
of β-actin. Quantification of gene expression was performed using
the 2−ΔΔCq method (26), with Cq representing the threshold
cycle. Target gene expression per sample was determined using the
following equation: Target in tumor tissue/target in non-tumorous
tissue. All reactions were performed in triplicate and normalized
to β-actin expression.
| Table II.Primers for quantitative polymerase
chain reaction analysis. |
Table II.
Primers for quantitative polymerase
chain reaction analysis.
|
| Primer
sequence |
|---|
|
|
|
|---|
| Gene name | Forward
(5′→3′) | Reverse
(5′→3′) |
|---|
| β-actin |
CCACTGGCATCGTGATGGA |
CGCTCGGTGAGGATCTTCAT |
| WWOX |
CAAGGGCGAGTGAAGCAGT |
GGCGGAGGGTGGTATTTTGT |
| WWOX-AS1 |
CAGAGTGAGAACCACTGGTGAT |
CGAAGGATGAGTTAAAAAGTTT |
Knockdown of WWOX-AS1 in SAOS2
cells
Small interfering (si)RNA sequences specific to
WWOX-AS1 and negative control (NC) siRNA were obtained from Chang
Jing Bio-Tech, Ltd and termed WWOX-AS1-si-1 and WWOX-AS1-si-2 (NC)
separately. The siRNA sequences used are also presented in Table III, and were identical to those
used in our previously published study (18). A total of 2×103 SAOS2
cells were plated in 6-well plates and then separately transfected
with 50 nmol/l WWOX-AS1-si-1 and 50 nmol/l WWOX-AS1-si-2 (NC) in
vitro using Lipofectamine® RNAiMAX (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. The cells in the wells were exposed to the transfection
mixture for 24 h at 37°C prior to the subsequent experiments.
| Table III.Sequences for small interfering RNA
analysis. |
Table III.
Sequences for small interfering RNA
analysis.
|
| Sequence |
|---|
|
|
|
|---|
| Gene name | Sense (5′→3′) | Antisense
(5′→3′) |
|---|
| WWOX-AS1-si-1 |
AUGGAUAUUUCCAAGCUGGAACCGA |
UCGGUUCCAGCUUGGAAAUAUCCAUGU |
| WWOX-AS1-si-2
(NC) |
AUUGAAAAUGUAAGAACUAUUCUGA |
UCAGAAUAGUUCUUACAUUUUCAAUCA |
Overexpression of WWOX-AS1 in
osteosarcoma cells
WWOX-AS1 was cloned into BamHI-EcoRI sites of
plasmid complementary (pc)DNA3.1 vectors (Promega Corporation,
Madison, WI, USA). The MG63 cells that expressed low levels of
WWOX-AS1 were transfected with 100 ng pcDNA-WWOX-AS1 or empty
vector using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
The cells in the wells were exposed to the transfection mixture for
24 h at 37°C prior to the subsequent experiments.
Cell proliferation assay
Cell proliferation was investigated using a MTS
(3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium)
assay kit (Promega Corporation, Madison, WI, USA) in accordance
with the manufacturer's protocol. MG63 and SAOS2 cells (2,000
cells/well) were separately plated in 96-well plates. MTS reagent
(20 µl) was added to each well containing 100 µl DMEM culture
medium supplemented with 10% FBS. Plates were subsequently
incubated for 2 h at 37°C in a humidified atmosphere containing 5%
CO2. The absorbance was measured at a wavelength of 490
nm using a plate reader. The optical density (OD) values at 24, 48,
72 and 96 h following transfection were determined and normalized
to the OD value at 0 h. Relative cell proliferation was
subsequently determined.
Scratch wound-healing assay
A total of 24 h post-transfection, MG63 and SAOS2
cells were seeded into a 6-well culture plate until a confluent
monolayer was established and then cultured at 37°C. Following
this, uniform wounds were scraped using a sterile 200 µl pipette
tip and each well was washed thrice using 1X PBS to remove floating
cells. The initial gap length (0 h) and the residual gap length (48
h) following wound initiation were calculated from images acquired
using a fluorescence inversion microscope system (Nikon
Corporation, Tokyo, Japan) at a magnification of ×200.
Matrigel invasion assay
Cell invasion assays were performed using modified
Boyden Chambers (BD Biosciences, Franklin Lakes, NJ, USA) according
to the manufacturer's protocol. A total of 2×103 cells
were suspended in 100 µl of DMEM supplemented with 10% FBS in the
lower chamber, and then plated in upper chambers with
Matrigel-coated inserts (pore size, 8 µm) in 24-well culture plates
(BD Biosciences). Invasive cells were stained at 42°C using 0.1%
crystal violet solution for 15 min and counted whilst passing
through the filter. The average invasion rate was calculated using
a fluorescence inversion microscope system (Nikon Corporation) at a
magnification of ×200; the increasing number of invasive cells as a
proportion of the entire cell population after 48 h.
Statistical analysis
Each experiment was repeated at least three times.
The Student's t-test and one-way analysis of variance followed by
the Newman-Keuls method were performed to analyze the statistical
differences between two groups and multiple groups, respectively.
The χ2 test was used to perform correlation analysis of
clinical data. Correlation between gene expression levels was
investigated using Pearson's correlation coefficient. Survival
analysis was performed using the Kaplan-Meier method, and
differences between the survival probabilities of each patient
group were compared using a log-rank test. The values are expressed
as the mean ± standard deviation. Data analyses were performed
using SPSS software (version 18.0; SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
WWOX-AS1 is downregulated in
osteosarcoma and is associated with prognosis of patients
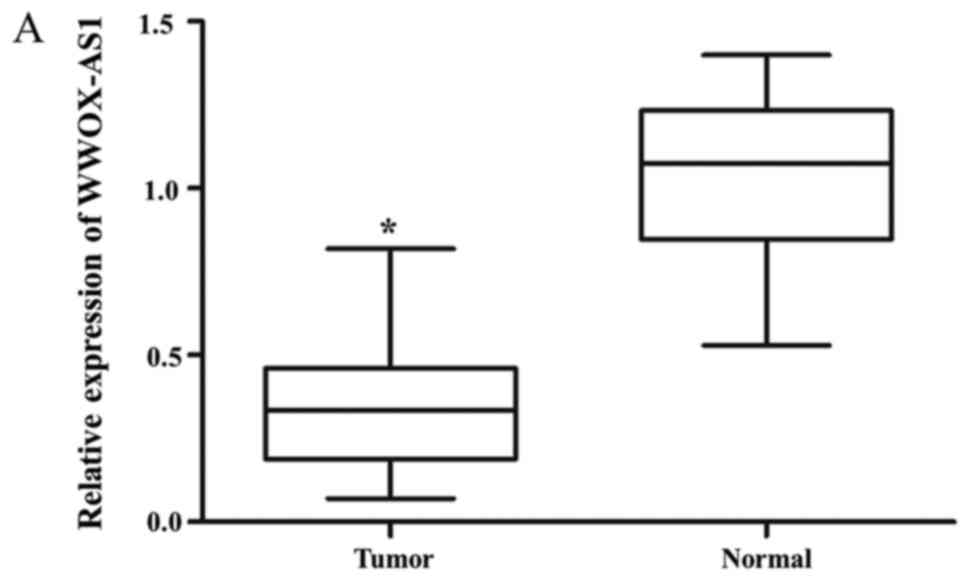
WWOX-AS1 expression in osteosarcoma tissues was
significantly downregulated in tumor tissues compared with adjacent
normal tissues (Fig. 1A).
Furthermore, WWOX-AS1 expression was revealed to be lowest in
patients with World Health Organization (WHO) grade III
osteosarcoma, and highest in patients with WHO grade I osteosarcoma
(Fig. 1B). In osteosarcoma tissue
specimens, the association between WWOX-AS1 expression and the
prognosis of patients with osteosarcoma was investigated using the
Kaplan-Meier and log-rank tests. The 70 patients with osteosarcoma
were classified into two groups: High-WWOX-AS1 group (n=35;
WWOX-AS1 expression value ≥median value) and low-WWOX-AS1 group
(n=35; WWOX-AS1 expression value ≤median value). The results
suggested that the overall survival probability of the
high-WWOX-AS1 group was significantly increased compared with the
low-WWOX-AS1 group (Fig. 1C;
P=0.036), which suggested that downregulation of WWOX-AS1 may serve
an important role in the progression of osteosarcoma. As revealed
in Table IV, significant
differences were observed between WWOX-AS1 expression and pulmonary
metastasis as well as WHO grade in the two different patient groups
(P<0.05). However, no significant associations were revealed
between WWOX-AS1 expression and sex, age, tumor size or
differentiation (Table IV).
| Table IV.Correlation between WW
domain-containing oxidoreductase-antisense RNA 1 expression and
clinicopathological characteristics of patients with
osteosarcoma. |
Table IV.
Correlation between WW
domain-containing oxidoreductase-antisense RNA 1 expression and
clinicopathological characteristics of patients with
osteosarcoma.
|
Characteristics | Low WWOX-AS1
expression | High WWOX-AS1
expression | χ2 | P-value |
|---|
| Sex |
|
| 0.9150 | 0.2430 |
|
Male | 20 | 16 |
|
|
|
Female | 15 | 19 |
|
|
| Age (years) |
|
| 2.8091 | 0.0937 |
|
<40 | 22 | 15 |
|
|
|
≥40 | 13 | 20 |
|
|
| Tumor size
(cm) |
|
| 0.9790 | 0.4212 |
|
<6 | 20 | 24 |
|
|
| ≥6 | 15 | 11 |
|
|
|
Differentiation |
|
| 1.5555 | 0.3096 |
|
Well | 8 | 9 |
|
|
|
Moderate | 17 | 11 |
|
|
|
Poor | 10 | 15 |
|
|
| Pulmonary
metastasis |
|
| 5.1851 | 0.0228 |
|
Yes | 12 | 4 |
|
|
| No | 23 | 31 |
|
|
| WHO grade |
|
| 1.942 | 0.0121 |
| I | 7 | 19 |
|
|
| II | 17 | 10 |
|
|
|
III | 11 | 6 |
|
|
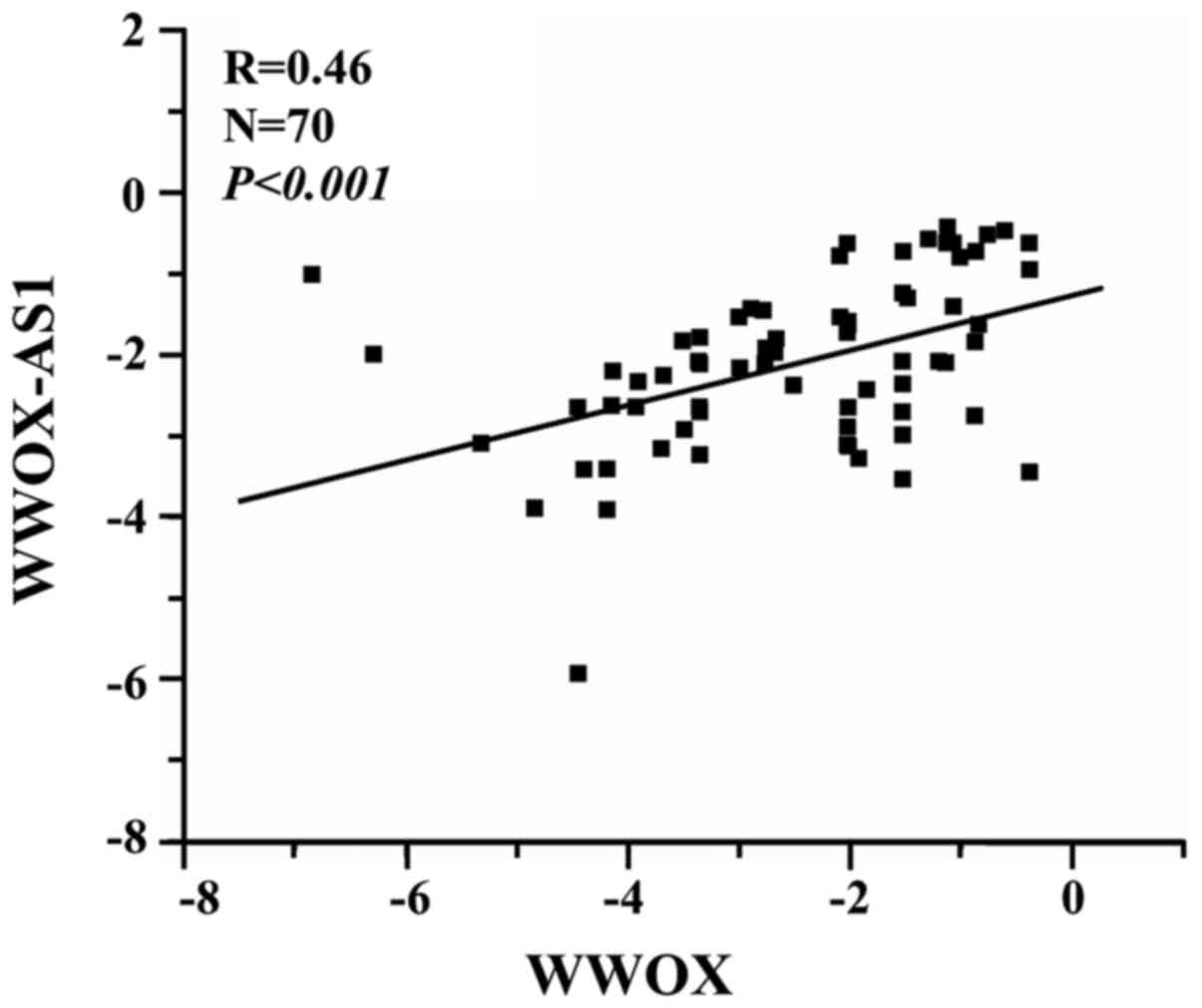
Expression of WWOX-AS1 is positively
correlated with WWOX expression
The association between WWOX-AS1 and WWOX was
investigated using RT-qPCR. The results suggested that WWOX-AS1 and
WWOX expression were positively correlated (R=0.46,
P<0.001; Fig. 2).
Alterations in WWOX expression
correspond with the expression of WWOX-AS1 following WWOX-AS1
overexpression or knockdown in osteosarcoma cell lines
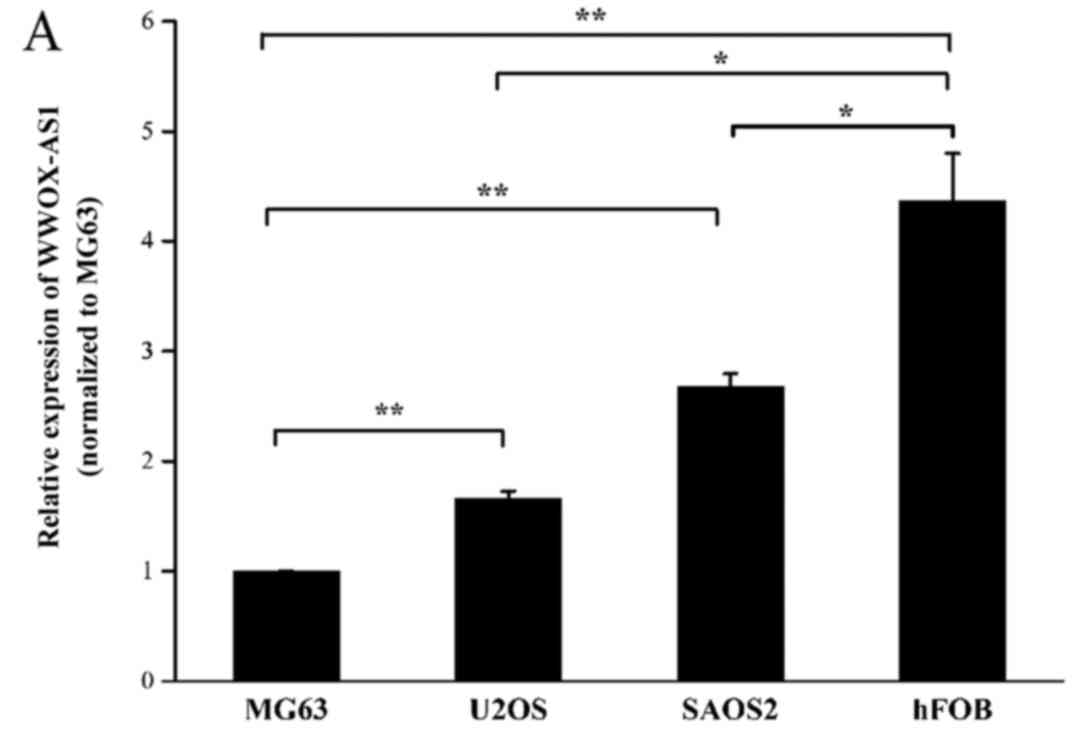
The expression of WWOX-AS1 was investigated in three
human osteosarcoma cell lines (SAOS2, MG63 and U2OS) and one human
normal osteoblastic cell line (hFOB) using RT-qPCR. The results
demonstrated that the expression level of WWOX-AS1 in hFOB was
significantly increased compared with all three human osteosarcoma
cell lines. Furthermore, SAOS2 and U2OS cells were revealed to
exhibit a significantly enhanced expression of WWOX-AS1 compared
with MGC3 cells (Fig. 3A).
In addition, WWOX-AS1 overexpression was analyzed
using RT-qPCR. WWOX-AS1 pcDNA was constructed and transfected into
MG63 cells. The results verified that WWOX-AS1 expression was
significantly enhanced in the pcDNA WWOX-AS1 group compared with
the pcDNA control group. In addition, the result revealed that WWOX
expression was also significantly upregulated in the pcDNA WWOX-AS1
group compared with the pcDNA control group (Fig. 3B).
Furthermore, WWOX-AS1 knockdown was investigated via
RT-qPCR. WWOX-AS1 siRNA was transfected into SAOS2 cells, and the
results demonstrated that both WWOX-AS1 and WWOX expression
levels were significantly downregulated in the WWOX-AS1 siRNA group
compared with the NC group (Fig.
3C). Therefore, the results suggested that WWOX
expression may be regulated by lncRNA WWOX-AS1 in osteosarcoma.
Proliferation, migration and invasion
abilities of MG63 cells are suppressed following WWOX-AS1
overexpression
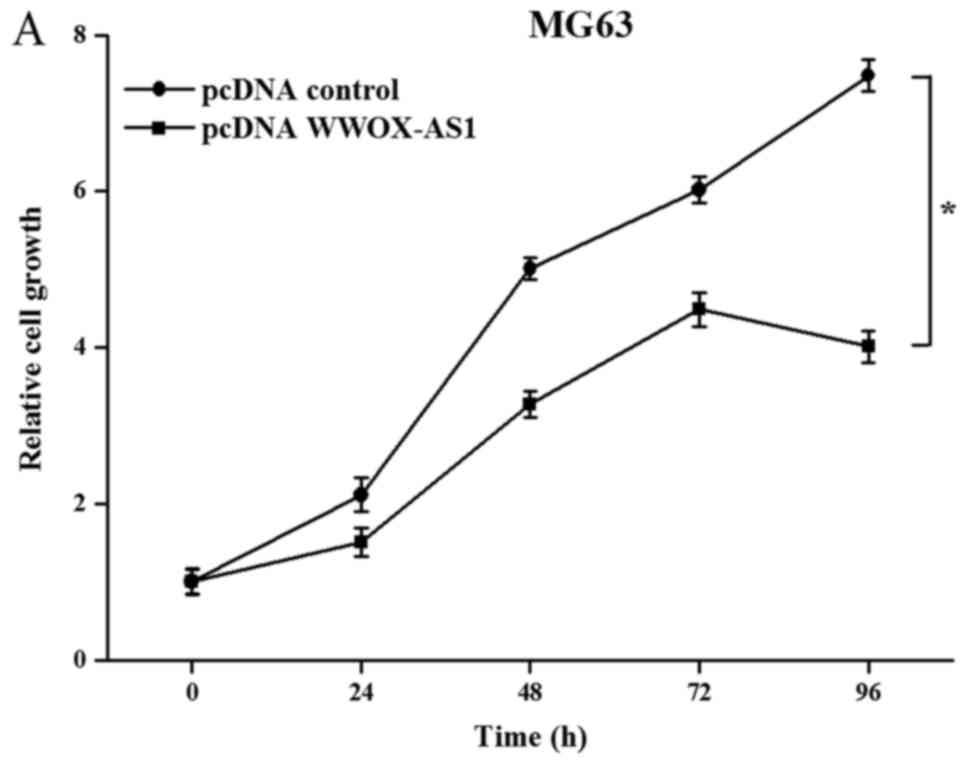
To investigate the role of WWOX-AS1 in osteosarcoma,
WWOX-AS1 overexpression in MG63 cells was performed. The results
demonstrated that the proliferation of MG63 cells transfected with
pcDNA WWOX-AS1 was significantly inhibited compared with the pcDNA
control group with increased time (Fig. 4A). In addition, the results of the
wound healing assay revealed that the migration of MG63 cells
transfected with pcDNA WWOX-AS1 was significantly suppressed
compared with the pcDNA control group (Fig. 4B). Furthermore, the results of the
Matrigel invasion assay demonstrated that the invasive ability of
cells transfected with pcDNA WWOX-AS1 was significantly decreased
compared with the pcDNA control group (Fig. 4C). In conclusion, the results
suggested that the proliferation, migration and invasion of MG63
cells were suppressed following WWOX-AS1 overexpression.
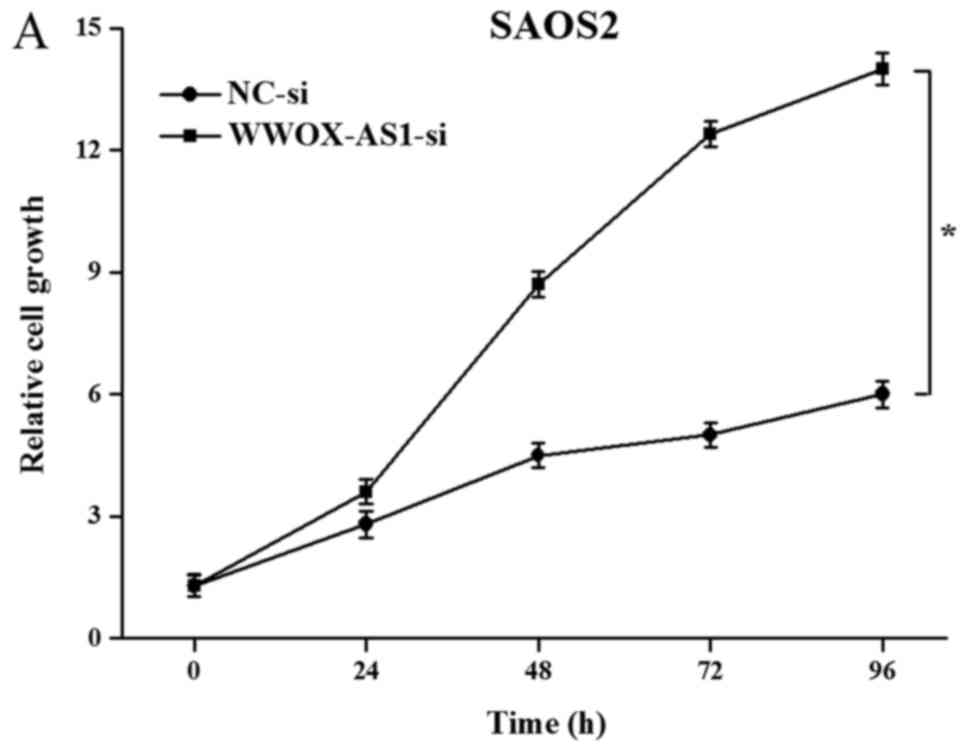
Proliferation, migration and invasion
of SAOS2 cells are enhanced following WWOX-AS1 knockdown
To further investigate the role of WWOX-AS1 in
osteosarcoma, WWOX-AS1 knockdown in SAOS2 cells was performed. The
results demonstrated that proliferation of cells transfected with
WWOX-AS1 siRNA was significantly enhanced compared with the NC
group with increased time (Fig.
5A). The results of the scratch wound-healing assay revealed
that cell migration in the WWOX-AS1 siRNA group was significantly
enhanced compared with the NC group (Fig. 5B). In addition, the results of the
Matrigel invasion assay demonstrated that cell invasion was
significantly increased in the WWOX-AS1 siRNA group compared with
the NC group (Fig. 5C). In
conclusion, the results suggested that the proliferation, migration
and invasion of SAOS2 cells were enhanced following WWOX-AS1
knockdown.
Discussion
Osteosarcoma is the most prevalent primary bone
tumor in children, adolescents and elderly adults (1). In recent years, a number of
therapeutic strategies have been used in the treatment of
osteosarcoma, including surgery, multiagent chemotherapy and
traditional Chinese medical treatments (27–33).
However, current treatments for osteosarcoma are still limited in
their effectiveness, as the molecular mechanisms underlying the
pathogenesis of osteosarcoma have not been fully determined.
Therefore, to improve outcomes for patients, it is important to
elucidate the potential mechanism and underlying regulation process
in osteosarcoma.
Previously, it has been demonstrated that antisense
lncRNAs may regulate their sense transcript in a positive or
negative manner (34–36). Furthermore, numerous antisense
lncRNAs have been revealed to be associated with numerous malignant
tumors (13–17). LncRNA WWOX-AS1 is the antisense
transcript of WWOX and has been demonstrated to regulate the
progression of numerous cancers, including osteosarcoma (23,24),
ovarian cancer (19),
hepatocellular carcinoma (20),
colon cancer (21) and bladder
cancer (22). However, the effect
of WWOX-AS1 on the progression of osteosarcoma remains
undetermined. The present study investigated the expression level
of WWOX-AS1 in human tissues and osteosarcoma cell lines. The
functional role of WWOX-AS1 was also analyzed in osteoblastic cell
lines. The results revealed that the expression of WWOX-AS1 was
downregulated in osteosarcoma tissues and cell lines compared with
healthy tissues and normal human osteoblastic cells, respectively.
The association between prognosis of patients and WWOX-AS1
expression was also determined using Kaplan-Meier and log-rank
tests, which suggested that patients with high WWOX-AS1 expression
exhibited a greater overall survival compared with patients with
low WWOX-AS1 expression. Furthermore, the results of the present
study demonstrated that WWOX-AS1 and WWOX expression levels
were positively correlated in patients. In addition, the expression
levels of WWOX were revealed to be significantly enhanced
and suppressed following WWOX-AS1 overexpression and knockdown in
osteosarcoma cell lines, respectively; which further suggested that
the expression levels of WWOX-AS1 and WWOX are positively
correlated in osteosarcoma. These results suggested that lncRNA
WWOX-AS1 may regulate WWOX expression. Similar results were
also demonstrated in our previous study regarding ovarian cancer
(18).
To date, numerous lncRNAs have been identified as
potent prognostic predictors in the progression of osteosarcoma
(9–12). Therefore, the present study aimed
to determine whether lncRNA WWOX-AS1 could suppress tumor
progression as well as suppress the proliferation, migration and
invasion of cells associated with osteosarcoma. The results
demonstrated that decreased expression of WWOX-AS1 was correlated
with elevated WHO grades, and that patients in the high-WWOX-AS1
group demonstrated a greater overall survival compared with
patients in the low-WWOX-AS1 group. Therefore, it can be suggested
that WWOX-AS1 is associated with tumor development in osteosarcoma.
This may be attributed to the association between WWOX and the
prognosis of osteosarcoma. For example, Zhang et al
(23) demonstrated that
WWOX gene polymorphisms may represent predictive factors for
the assessment of risks and outcomes associated with osteosarcoma.
Kurek et al (37) revealed
the occurrence and frequency of WWOX deletion in
osteosarcoma as well as the prognostic and therapeutic significance
of WWOX in osteosarcoma. The results of the present study
demonstrated that WWOX-AS1 overexpression suppressed the
proliferation, migration and invasion of MG63 cells. In addition,
the results demonstrated that WWOX-AS1 knockdown enhanced the
proliferation, migration and invasion of SAOS2 cells. These results
demonstrated the inhibitory role of WWOX-AS1 on the proliferation,
migration and invasion abilities of osteosarcoma cells. Previous
studies have demonstrated that, as a tumor suppressor gene,
WWOX could suppress the proliferation, migration, invasion
and tumorigenicity of osteosarcoma cells (38), most likely via regulation of runt
related transcription factor 2 (RUNX2) (24). This further suggests that there is
an association between WWOX-AS1 and WWOX. In addition, genes
regulated by WWOX in osteosarcoma may provide important
information regarding the underlying mechanism of WWOX-AS1 in
osteosarcoma. RUNX2 has been previously demonstrated to activate
genes associated with angiogenesis (vascular endothelial growth
factor and osteopontin) (39),
metastasis and invasion [matrix metallopeptidase (MMP)2, MMP9 and
MMP13] (40,41), and survival (survivin) (42). It has been previously demonstrated
that WWOX loss of function is associated with osteosarcoma
progression by inhibition of numerous genes involved in metastasis
via regulation of RUNX2 (24).
Furthermore, a previous study demonstrated that investigation into
the genomic function of RUNX2 in osteosarcoma cells provides
insight into dysregulated molecular pathways in bone cancer
(43). Therefore, it may be
hypothesized that the molecular mechanisms underlying the
involvement of WWOX-AS1 in osteosarcoma may be associated with
RUNX2 or genes activated by RUNX2.
The present study, to the best of the authors
knowledge, is the first to investigate the functional role of
WWOX-AS1 in osteosarcoma. The results of the present study may
provide novel insight into the roles of WWOX-AS1 in osteosarcoma,
and whether WWOX-AS1 may represent a promising therapeutic target
and novel biomarker for the treatment of osteosarcoma.
However, there were certain limitations to the
present study. First, clinical information and samples were only
collected from Chinese patients with osteosarcoma, therefore
limiting the generalizability of the results. Second, in
vivo experiments were performed using si-WWOX-AS1; however, it
failed to achieve stable results (data not shown in this study).
The silencing effect induced by exogenous delivery of synthetic
siRNAs is transient and reactivation of the target gene typically
occurs with a few days' interval (44). siRNA as exogenous RNA is easily
degraded by nuclease in vivo and can therefore lead to
unstable results. In addition, the present study did not
investigate the association between WWOX-AS1 expression and
apoptosis. Furthermore, the exact molecular mechanism of WWOX-AS1
was not determined in the present study. Considering these
limitations, more extensive collection of comprehensive clinical
information and subsequent analyses are required to further verify
the results of the present study. Our future studies will aim to
investigate the mechanisms underlying the association between
WWOX-AS1 and osteosarcoma using viral transfection, and to
establish stable cell lines. Future studies should further
investigate important genes (including RUNX2) and signaling
pathways associated with WWOX-AS1 in osteosarcoma.
Acknowledgements
The authors would like to thank Dr Yao (Department
of Oncology) and Dr Yang (Department of Otorhinolaryngology) at the
161th Hospital of PLA (Wuhan, China) who collected the data and
provided useful suggestions for the manuscript.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GQ, ZM and WT contributed equally to this article.
GQ conceived and designed the study and drafted the manuscript. ZM
and WT analyzed and interpreted the data. JY put forward the
concept of the study and reviewed the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committees of the General Hospital of the People's Liberation
Army.
Consent for publication
Written informed consent was obtained from each
patient or their guardians.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
McKenna WG, Barnes MM, Kinsella TJ,
Rosenberg SA, Lack EE and Glatstein E: Combined modality treatment
of adult soft tissue sarcomas of the head and neck. Int J Radiat
Oncol Biol Phys. 13:1127–1133. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen X, Bahrami A, Pappo A, Easton J,
Dalton J, Hedlund E, Ellison D, Shurtleff S, Wu G, Wei L, et al:
Recurrent somatic structural variations contribute to tumorigenesis
in pediatric osteosarcoma. Cell Rep. 7:104–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee JT: Epigenetic regulation by long
noncoding RNAs. Science. 338:1435–1439. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsai MC, Spitale RC and Chang HY: Long
intergenic noncoding RNAs: New links in cancer progression. Cancer
Res. 71:3–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fan H, Zhu JH and Yao XQ: Knockdown of
long noncoding RNA PVT1 reverses multidrug resistance in colorectal
cancer cells. Mol Med Rep. 2018. View Article : Google Scholar
|
|
8
|
An Q, Zhou L and Xu N: Long noncoding RNA
FOXD2-AS1 accelerates the gemcitabine-resistance of bladder cancer
by sponging miR-143. Biomed Pharmacother. 103:415–420. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou Q, Chen F, Fei Z, Zhao J, Liang Y,
Pan W, Liu X and Zheng D: Genetic variants of lncRNA HOTAIR
contribute to the risk of osteosarcoma. Oncotarget. 7:19928–19934.
2016.PubMed/NCBI
|
|
10
|
Chen F, Mo J and Zhang L: Long noncoding
RNA BCAR4 promotes osteosarcoma progression through activating
GLI2-dependent gene transcription. Tumour Biol. 37:13403–13412.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Z, Zhao L and Wang Q: Overexpression of
long non-coding RNA HOTTIP increases chemoresistance of
osteosarcoma cell by activating the Wnt/β-catenin pathway. Am J
Transl Res. 8:2385–2393. 2016.PubMed/NCBI
|
|
12
|
Li W, Xie P and Ruan WH: Overexpression of
lncRNA UCA1 promotes osteosarcoma progression and correlates with
poor prognosis. J Bone Oncol. 5:80–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pasmant E, Laurendeau I, Heron D, Vidaud
M, Vidaud D and Bieche I: Characterization of a germ-line deletion,
including the entire INK4/ARF locus, in a melanoma-neural system
tumor family: Identification of ANRIL, an antisense noncoding RNA
whose expression coclusters with ARF. Cancer Res. 67:3963–3969.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu W, Gius D, Onyango P, Muldoon-Jacobs K,
Karp J, Feinberg AP and Cui H: Epigenetic silencing of tumour
suppressor gene p15 by its antisense RNA. Nature. 451:202–206.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Takayama K, Horie-Inoue K, Katayama S,
Suzuki T, Tsutsumi S, Ikeda K, Urano T, Fujimura T, Takagi K,
Takahashi S, et al: Androgen-responsive long noncoding RNA CTBP1-AS
promotes prostate cancer. EMBO J. 32:1665–1680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang X, Song JH, Cheng Y, Wu W, Bhagat T,
Yu Y, Abraham JM, Ibrahim S, Ravich W, Roland BC, et al: Long
non-coding RNA HNF1A-AS1 regulates proliferation and migration in
oesophageal adenocarcinoma cells. Gut. 63:881–890. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Han L, Kong R, Yin DD, Zhang EB, Xu TP, De
W and Shu YQ: Low expression of long noncoding RNA GAS6-AS1
predicts a poor prognosis in patients with NSCLC. Med Oncol.
30:6942013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tong W, Yang L, Yu Q, Yao J and He A: A
new tumor suppressor lncRNA RP11-190D6.2 inhibits the
proliferation, migration, and invasion of epithelial ovarian cancer
cells. Onco Targets Ther. 10:1227–1235. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan H, Tong J, Lin X, Han Q and Huang H:
Effect of the WWOX gene on the regulation of the cell cycle and
apoptosis in human ovarian cancer stem cells. Mol Med Rep.
12:1783–1788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xie B, Zen Q, Wang X, He X, Xie Y, Zhang Z
and Li H: ACK1 promotes hepatocellular carcinoma progression via
downregulating WWOX and activating AKT signaling. Int J Oncol.
46:2057–2066. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nowakowska M, Pospiech K, Lewandowska U,
Piastowska-Ciesielska AW and Bednarek AK: Diverse effect of WWOX
overexpression in HT29 and SW480 colon cancer cell lines. Tumour
Biol. 35:9291–9301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li G, Sun L, Mu Z, Huang Y, Fu C and Hu B:
Ectopic WWOX expression inhibits growth of 5637 bladder cancer cell
in vitro and in vivo. Cell Biochem Biophys. 73:417–425. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang N, Jiang Z, Ren W, Yuan L and Zhu Y:
Association of polymorphisms in WWOX gene with risk and outcome of
osteosarcoma in a sample of the young Chinese population. Onco
Targets Ther. 9:807–813. 2016.PubMed/NCBI
|
|
24
|
Del Mare S and Aqeilan RI: Tumor
Suppressor WWOX inhibits osteosarcoma metastasis by modulating
RUNX2 function. Sci Rep. 5:129592015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jawad MU and Scully SP: In brief:
Classifications in brief: Enneking classification: Benign and
malignant tumors of the musculoskeletal system. Clin Orthop Relat
Res. 468:2000–2002. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang T, Li J, Yin F, Lin B, Wang Z, Xu J,
Wang H, Zuo D, Wang G, Hua Y and Cai Z: Toosendanin demonstrates
promising antitumor efficacy in osteosarcoma by targeting STAT3.
Oncogene. 36:6627–6639. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu X, Xiu LJ, Jiao JP, Zhao J, Zhao Y, Lu
Y, Shi J, Li YJ, Ye M, Gu YF, et al: Traditional Chinese medicine
integrated with chemotherapy for stage IV non-surgical gastric
cancer: A retrospective clinical analysis. J Integr Med.
15:469–475. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mondal J, Das J, Shah R and Khuda-Bukhsh
AR: A homeopathic nosode, Hepatitis C 30 demonstrates anticancer
effect against liver cancer cells in vitro by modulating telomerase
and topoisomerase II activities as also by promoting apoptosis via
intrinsic mitochondrial pathway. J Integr Med. 14:209–218. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Adaramoye O, Erguen B, Oyebode O, Nitzsche
B, Höpfner M, Jung K and Rabien A: Antioxidant, antiangiogenic and
antiproliferative activities of root methanol extract of Calliandra
portoricensis in human prostate cancer cells. J Integr Med.
13:185–193. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang X, Wang N, Cheung F, Lao L, Li C and
Feng Y: Chinese medicines for prevention and treatment of human
hepatocellular carcinoma: Current progress on pharmacological
actions and mechanisms. J Integr Med. 13:142–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zang QQ, Zhang L, Gao N and Huang C:
Ophiopogonin D inhibits cell proliferation, causes cell cycle
arrest at G2/M, and induces apoptosis in human breast carcinoma
MCF-7 cells. J Integr Med. 14:51–59. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wahlestedt C: Natural antisense and
noncoding RNA transcripts as potential drug targets. Drug Discov
Today. 11:503–508. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Faghihi MA and Wahlestedt C: Regulatory
roles of natural antisense transcripts. Nat Rev Mol Cell Biol.
10:637–643. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yelin R, Dahary D, Sorek R, Levanon EY,
Goldstein O, Shoshan A, Diber A, Biton S, Tamir Y, Khosravi R, et
al: Widespread occurrence of antisense transcription in the human
genome. Nat Biotechnol. 21:379–386. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kurek KC, Del Mare S, Salah Z, Abdeen S,
Sadiq H, Lee SH, Gaudio E, Zanesi N, Jones KB, DeYoung B, et al:
Frequent attenuation of the WWOX tumor suppressor in osteosarcoma
is associated with increased tumorigenicity and aberrant RUNX2
expression. Cancer Res. 70:5577–5586. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Del Mare S, Kurek KC, Stein GS, Lian JB
and Aqeilan RI: Role of the WWOX tumor suppressor gene in bone
homeostasis and the pathogenesis of osteosarcoma. Am J Cancer Res.
1:585–594. 2011.PubMed/NCBI
|
|
39
|
Zelzer E, Glotzer DJ, Hartmann C, Thomas
D, Fukai N, Soker S and Olsen BR: Tissue specific regulation of
VEGF expression during bone development requires Cbfa1/Runx2. Mech
Dev. 106:97–106. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pratap J, Javed A, Languino LR, van Wijnen
AJ, Stein JL, Stein GS and Lian JB: The Runx2 osteogenic
transcription factor regulates matrix metalloproteinase 9 in bone
metastatic cancer cells and controls cell invasion. Mol Cell Biol.
25:8581–8591. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang X, Manner PA, Horner A, Shum L, Tuan
RS and Nuckolls GH: Regulation of MMP-13 expression by RUNX2 and
FGF2 in osteoarthritic cartilage. Osteoarthritis Cartilage.
12:963–973. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lim M, Zhong C, Yang S, Bell AM, Cohen MB
and Roy-Burman P: Runx2 regulates survivin expression in prostate
cancer cells. Lab Invest. 90:222–233. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
van der Deen M, Akech J, Lapointe D, Gupta
S, Young DW, Montecino MA, Galindo M, Lian JB, Stein JL, Stein GS
and van Wijnen AJ: Genomic promoter occupancy of runt-related
transcription factor RUNX2 in osteosarcoma cells identifies genes
involved in cell adhesion and motility. J Biol Chem. 287:4503–4517.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Elbashir SM, Harborth J, Weber K and
Tuschl T: Analysis of gene function in somatic mammalian cells
using small interfering RNAs. Methods. 26:199–213. 2002. View Article : Google Scholar : PubMed/NCBI
|