Introduction
Atrial fibrillation (AF) is an abnormal heart rhythm
characterized by rapid and irregular beating, which has a
prevalence of ~1% in the general population and >8% in patients
aged ≥80 years old (1,2). AF is not a life-threatening
arrhythmia; however, it impairs the quality of life of patients as
a result of its anatomic, hemodynamic, and hemocoagulative
consequences (3). AF markedly
influences the morbidity and mortality of patients (4), as it can increase the risk of heart
failure and stroke (5). In
addition, as its prevalence continuously increases, AF has become
an important cause of healthcare expenditure worldwide (6,7).
Therefore, the elucidation of the molecular mechanisms underlying
the pathophysiology of AF is of critical importance.
MicroRNAs (miRNAs) are a class of endogenous small
non-coding RNA molecules, with a length of 18–22 nucleotides
(8). miRNAs bind to the
3′-untranslated region of target mRNAs and thus regulate the
expression of numerous target genes (9); miRNAs have been reported to regulate
~60% of genes (10). Aberrant
miRNA expression has been implicated in various pathophysiological
processes, including the pathogenesis of cardiac diseases, and
their detection may have potential as a novel diagnostic tool
(11). Several miRNAs have been
reported to be abnormally expressed in AF, including miRNA
(miR)-328 (12), miR-1 (13) and miR-26 (14). The expression of miR-122 has
previously been associated with the development of cardiovascular
diseases, including acute coronary syndrome (15) and myocardial fibrosis (16). In addition, Liu et al
(9) demonstrated that serum
miR-122 levels were significantly decreased in rats with
experimental stroke produced by middle cerebral artery occlusion,
as well as in humans with ischemic stroke. Therefore, it is
hypothesized that miR-122 may be implicated in the development and
progression of AF.
In the present study, an AF mouse model was
generated by transesophageal rapid atrial stimulation.
Cardiomyocytes were isolated from AF mice, transfected with miR-122
inhibitors or negative controls and miR-122 expression was
assessed. In addition, cardiomyocyte proliferation and apoptosis
were evaluated. In order to investigate the molecular mechanisms
underlying the effect of miR-122 in AF, western blot analysis was
used to assess the expression of extracellular signal-regulated
kinase (ERK) and phosphorylated (p)-ERK, as well as of the
apoptosis-associated proteins caspase-3 and B-cell lymphoma 2-like
1 (Bcl-x). The present study aimed to provide evidence suggesting
the implication of miR-122 in the mechanisms underlying AF
pathophysiology.
Materials and methods
Animals
Male C57BL/6 mice were originally purchased from
Japan SLC, Inc. (Shizuoka, Japan), and were provided with standard
food and water. In the present study, 30 male mice (age, 12–14
weeks; weigh, 22–24 g) were used. The mice were housed in a
pathogen-free facility with free access to standard food and water.
The temperature was maintained at 25±2°C and the relative humidity
was 55±10% under a 12 h light/12 h dark. All experimental
procedures were approved by the Ethics Committee of Tianjin Medical
University General Hospital (Tianjin, China).
AF model
A total of 30 male C57BL/6 mice were divided into
the following three groups (n=10 mice/group): AF, sham-operation
and control group. Transesophageal rapid atrial stimulation was
used for the induction of AF, as previously described (17). Briefly, mice were anesthetized by
isoflurane inhalation (1.5–2% for maintenance). A 1.1 French
octapolar catheter with eight circular electrodes (0.5 mm) and an
interelectrode distance of 1 mm (cat no. EPR 800; Millar, Inc.,
Houston, TX, USA) was advanced through the esophagus. The catheter
was placed at the site with the lowest threshold for atrial
capture, as previously described (18). In order to ensure the correct
position of the pacing catheter, atrial capture with 1:1
atrioventricular conduction was measured prior to the burst pacing
period (19). Transesophageal
atrial burst pacing was then induced for 10 sec, at a stimulation
amplitude of 1.5 mA, using 10 msec cycle lengths and a pulse width
of 3 mA. Mice in the sham-operation group did not receive
transesophageal atrial burst pacing. The mice in the control group
received no intervention.
Cell isolation and culture
Two weeks following operation, the cardiomyocytes
were isolated from all the three groups as previously described
(20). Following 3–5 days of
culture, cardiomyocytes that exhibited regular spontaneous
contractions were used for experiments. Cardiomyocytes were
cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% (v/v) fetal calf serum (FCS; Invitrogen; Thermo Fisher
Scientific, Inc.), 100 IU/ml penicillin and 0.08 mg/ml gentamicin
and maintained in 95% air and 5% CO2 at 37°C.
Cell transfection
miR-122 inhibitors and inhibitor controls were
purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
miRNAs (100 nM) were transiently transfected into target cells from
AF mice (1×104 cells) using Lipofectamine
2000® (Invitrogen; Thermo Fisher Scientific, Inc.) as
the transfection reagent, according to the manufacturer's protocol.
Following 48 h of transfection, subsequent experiments were
performed. The sequences for miR-122 inhibitor was
5′-CAAACACCAUUGUCACACUCCA-3′ and control was
5′-GCCCAGGCAAAUGACACAGUUC-3′.
MTT assay
Cells were seeded in 96-well plates at a density of
103 cells/well. Cell viability was assessed following 72
h and 7 days of culture using an MTT colorimetric assay, according
to the manufacturer's protocol. Briefly, 10 µl (5 mg/ml) MTT
solution (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added
to each well. Following 4 h of incubation at 37°C, the samples were
centrifuged at 4°C and 600 × g for 5 min, and the formazan product
was dissolved with DMSO. The optical density of the supernatants
was measured at 570 nm using a UV-Vis microplate spectrophotometer
(BioTek China, Beijing, China). The experiment was repeated three
times.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick-end labeling
(TUNEL) assay
Cardiomyocyte apoptosis was assessed using the TUNEL
assay kit (Roche Diagnostics, Indianapolis, USA). Apoptotic nuclei
were fluorescently labeled with the 50 µl TUNEL reagent for 1 h at
37°C in the dark. Cell nuclei were counterstained with 10 µg/ml
Hoechst 33258 for 5 min at room temperature. The number of
apoptotic cells was determined under a confocal fluorescence
microscope (Olympus, Otsu, Japan). The apoptotic cells were
calculated as the number of TUNEL-positive and DAPI-stained cells
divided by the total number of cells in five random fields. This
assay was repeated at least three times.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. Total RNA was reverse transcribed into
cDNA using an RT Reagent kit (Qiagen, Inc., Valencia, CA, USA),
according to the manufacturer's protocol. qPCR was performed using
the SYBR-Green PCR Master Mix (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Thermocycling conditions
were as follows: Denaturation at 95°C for 30 sec, followed by 40
cycles of 95°C for 5 sec and 60°C for 34 sec. The primers for
miR-122 (cat no. 3416) and small nucleolar RNA D68 (Snord68; cat
no. 33712), used as the housekeeping gene, were synthesized by
Qiagen GmbH (Hilden, Germany). Relative miRNA expression was
quantified using the 2−ΔΔCq method (20). The primers were listed as follows:
miR-122 forward, 5′-GAGAGGCCTAAAGCCACAGA-3′ and reverse,
5′-CACTTACCCCCAGTCAGCTC-3′. Snord68 forward,
5′-ACGACTAGGGCTGTACTGACTTGATG-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGG-3′.
Western blot analysis
Cardiomyocytes were lysed using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) supplemented with a cocktail of
protease and phosphatase inhibitors on an ice bath. Following
ultrasonic fragmentation, lysates were centrifuged at 1,500 × g for
30 min at 4°C, and the supernatant was collected into an Eppendorf
tube. Protein concentration was determined using a bicinchoninic
acid protein assay kit (Beyotime Institute of Biotechnology). Equal
amounts (20 µg) of extracted protein samples were separated by
10–12% SDS-PAGE and transferred onto polyvinylidene difluoride
membranes (Thermo Fisher Scientific, Inc.). Membranes were blocked
with 5% nonfat dry milk at room temperature for 1 h, and then
probed with anti-Bcl-x (cat. no. 2764; dilution 1:1,000),
anti-caspase-3 (cat. no. 9662; dilution 1:1,000), anti-ERK (cat.
no. 4695; dilution 1:1,000) and anti-p-ERK (cat. no. 4370; dilution
1:1,000) primary antibodies (Cell Signaling Technology, Inc.,
Danvers, MA, USA) overnight at 4°C. Following three washes, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibody (cat. no. 7074; dilution 1:2,000) at room
temperature for 1 h (Cell Signaling Technology, Inc.) for 2 h.
β-actin (cat. no. 4970; dilution 1:1,000) was used as the internal
control (Cell Signaling Technology, Inc.). Protein bands were
visualized using an enhanced chemiluminescence system (Amersham
Biosciences; GE Healthcare, Chicago, IL, USA). Blots were
semi-quantified by densitometric analysis on a gel documentation
system using the Image Lab Software version 5.2.1 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Enumerated data were analyzed using a χ2
or Wilcoxon rank-sum test. The statistical significance of the
differences between groups was assessed by one-way analysis of
variance, followed by a post hoc Tukey test for multiple
comparisons. Data are expressed as the mean ± standard deviation
and the experiments were repeated at least three times. Statistical
analysis was performed using the SPSS software version 16.0 (SPSS
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-122 expression in
cardiomyocytes
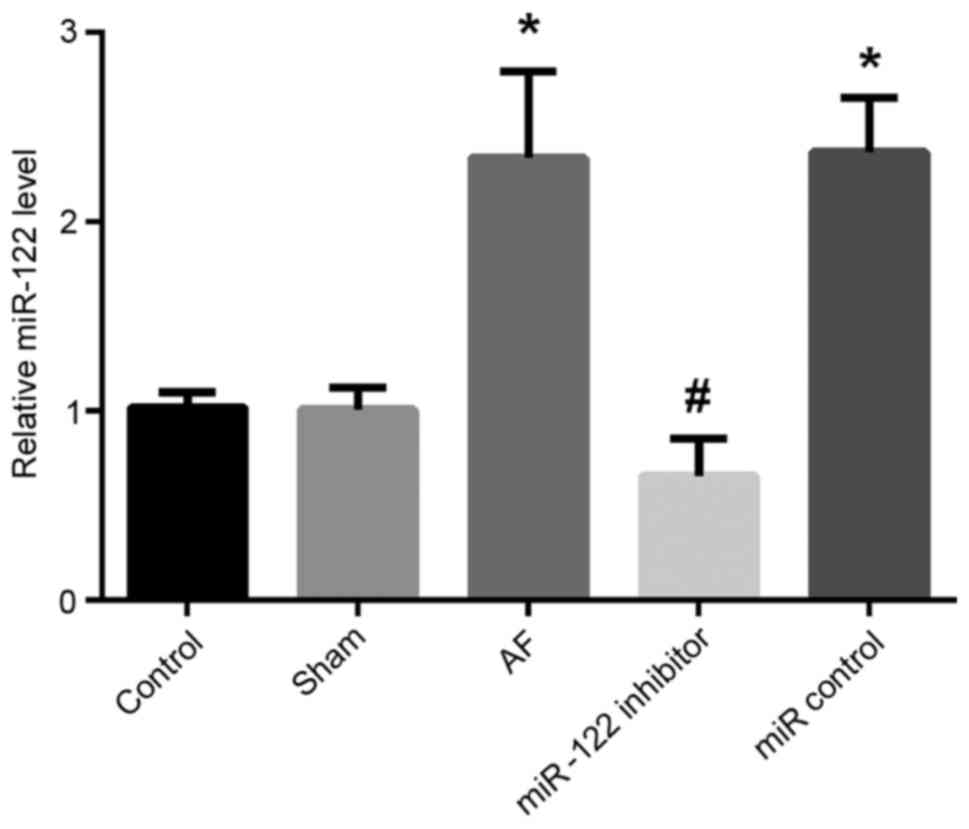
RT-qPCR was used to assess the expression of miR-122
in cardiomyocytes (Fig. 1). The
present results demonstrated that the expression of miR-122 in the
AF group was significantly increased compared with the
sham-operation and control groups (P<0.05). By contrast,
following transfection with a miR-122 inhibitor, the expression of
miR-122 in cardiomyocytes isolated from AF mice was significantly
decreased compared with AF cardiomyocytes transfected with a
miR-control (P<0.05).
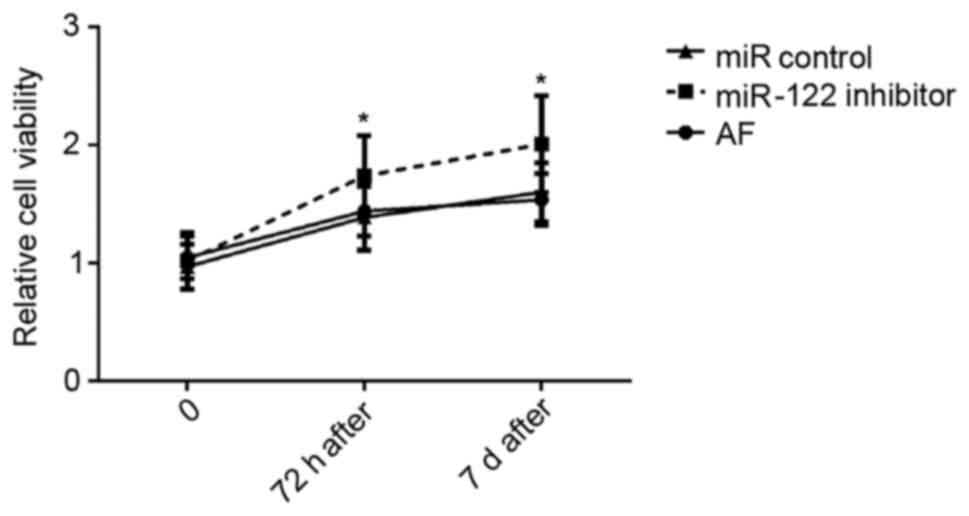
Cardiomyocyte viability
Cardiomyocytes isolated from AF mice were
transfected with a miR-122 inhibitor or an inhibitor control, and
cell viability was evaluated 72 h and 7 days post-transfection,
using an MTT assay (Fig. 2).
Following miR-122 inhibition, the viability of cardiomyocytes
appeared to be significantly enhanced 72 h and 7 days
post-transfection compared with cells transfected with the
miR-control (P<0.05).
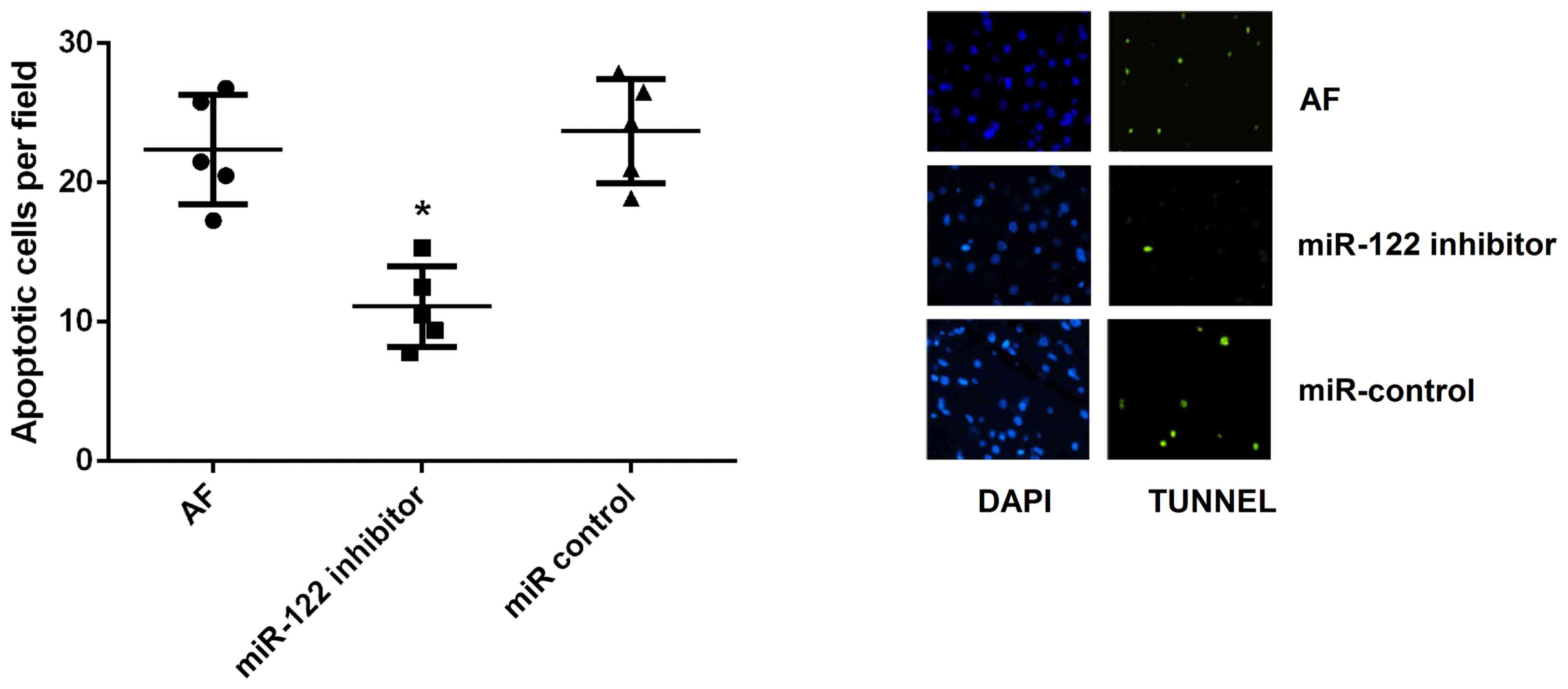
Cardiomyocyte apoptosis
Cardiomyocyte apoptosis is critical in the pathology
of persistent AF. In the present study, a TUNEL assay demonstrated
that the apoptotic rate of cardiomyocytes was significantly
decreased following miR-122 inhibition compared with cardiomyocytes
in the AF and miR-control groups (P<0.05; Fig. 3).
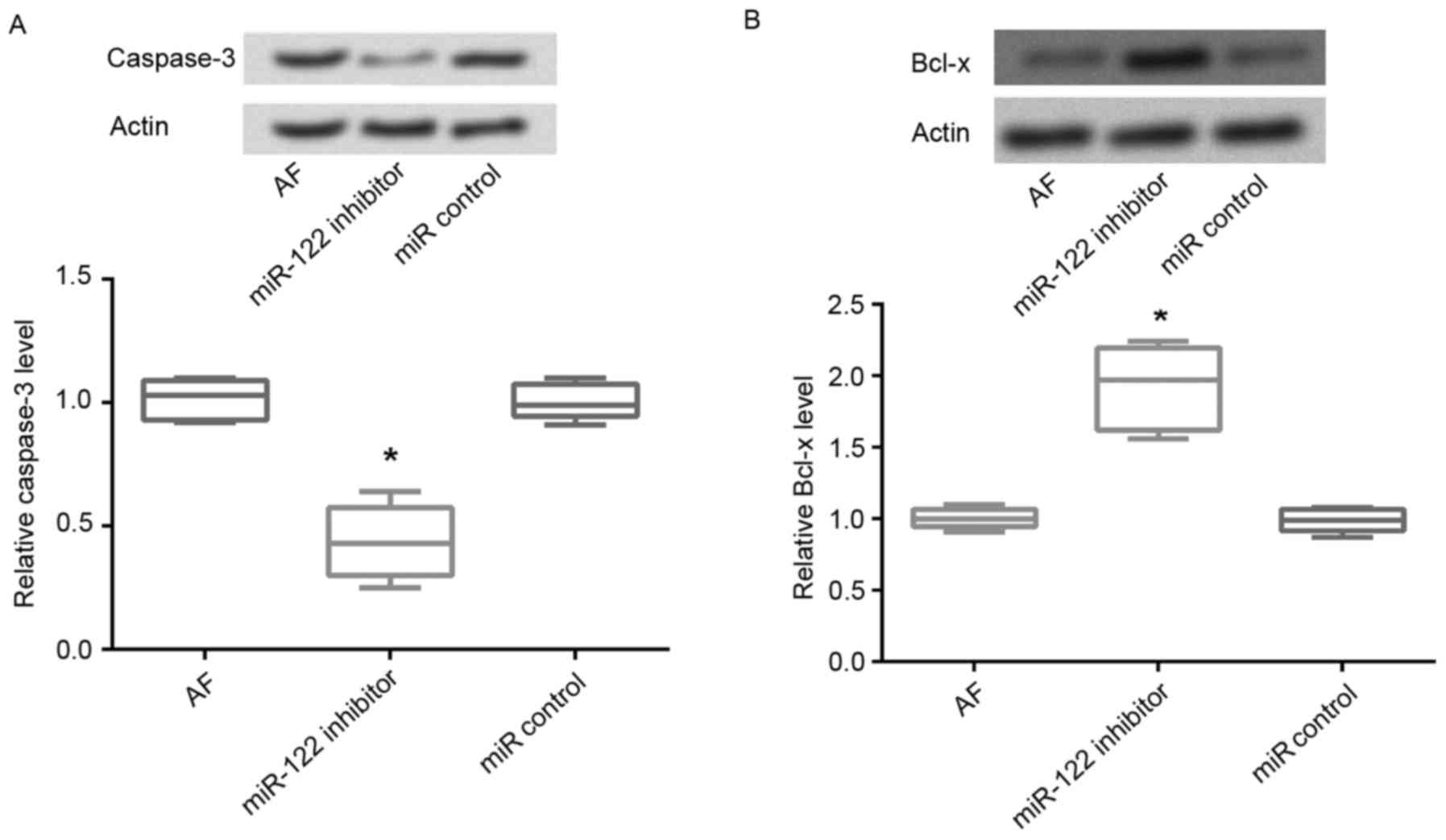
Expression of apoptosis-associated
proteins
The protein expression levels of the pro-apoptotic
factor caspase-3 and the anti-apoptotic factor Bcl-x were assessed
using western blot analysis. The present results revealed that
following miR-122 inhibition in AF cardiomyocytes, the protein
expression levels of caspase-3 were significantly decreased
compared with untransfected cells (P<0.05; Fig. 4A). Conversely, the protein
expression levels of Bcl-x were significantly upregulated following
miR-122 inhibition compared with untransfected AF cardiomyocytes
and cells transfected with the miR-control (P<0.05; Fig. 4B).
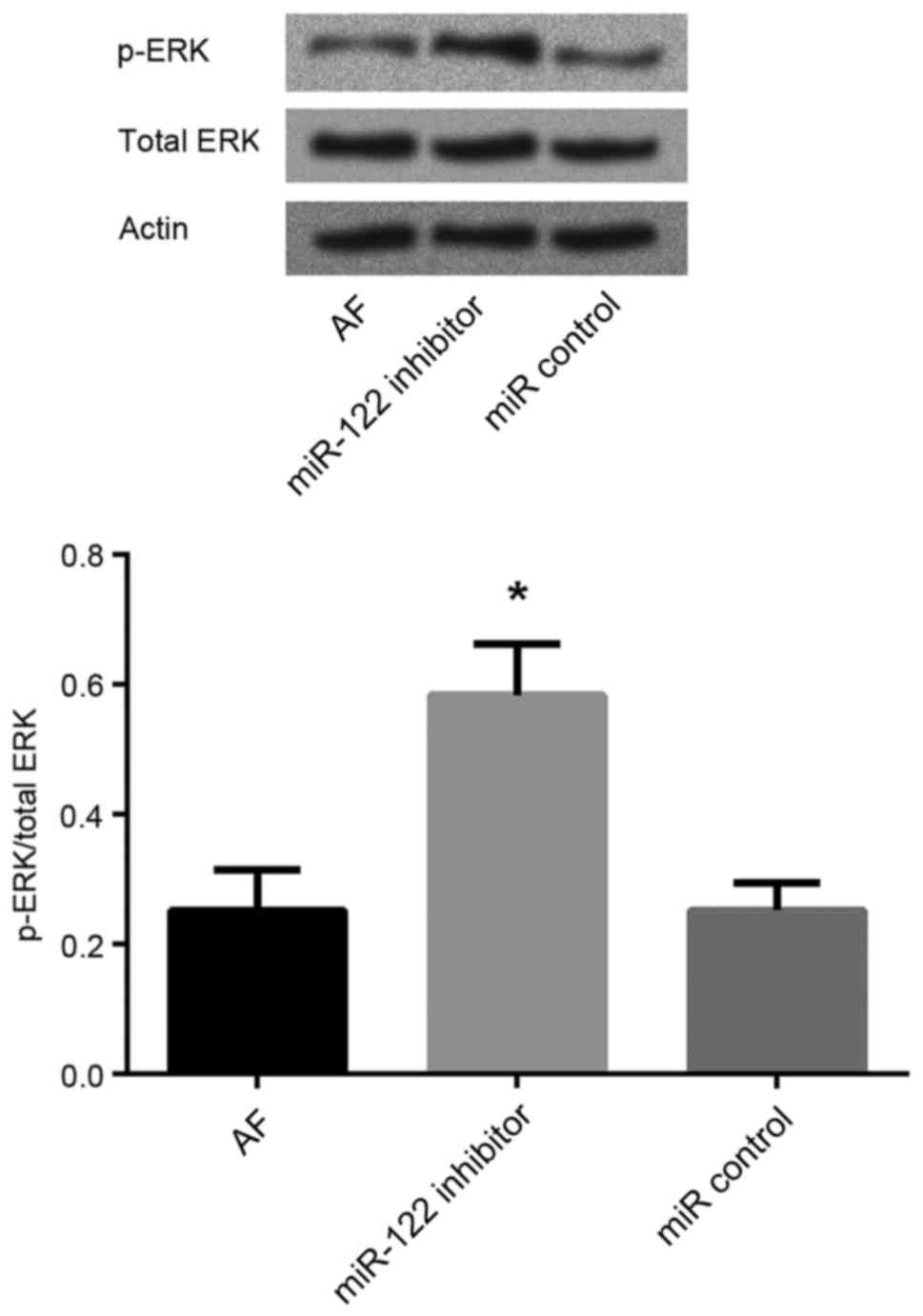
miR-122 inhibition enhances ERK
phosphorylation
To further investigate the molecular mechanisms
underlying the involvement of miR-122 in AF, the protein expression
levels of ERK and p-ERK in cardiomyocytes were assessed using
western blot analysis. Following miR-122 inhibition, the p-ERK/ERK
ratio was significantly increased in AF cardiomyocytes compared
with untransfected AF cardiomyocytes and cells transfected with the
miR-control (P<0.05; Fig.
5).
Discussion
AF is a major public health concern, as well as a
socioeconomic problem, due to its debilitating comorbidities, which
include cognitive disturbances, permanent disability and recurrent
hospitalization (21). Therefore,
the elucidation of the molecular mechanisms underlying AF
pathogenesis, and the development of novel therapeutic strategies
for the treatment of patients with AF is imperative. The present
study demonstrated that the expression of miR-122 was significantly
upregulated in cardiomyocytes isolated from mice with AF compared
with control mice. Following knockdown of miR-122 expression in AF
cardiomyocytes, using transfection with a miR-122 inhibitor, the
cell viability of cardiomyocytes was enhanced, whereas their
apoptosis was significantly suppressed. Furthermore, the protein
expression of the pro-apoptotic factor caspase-3 was significantly
decreased, whereas the expression of the anti-apoptotic Bcl-x was
significantly upregulated, following miR-122 silencing in AF
cardiomyocytes.
miR-122 has been revealed to be highly expressed in
liver tissue, where it may be implicated in the regulation of fatty
acid metabolism (22,23). Wei et al (24) suggested that miR-122 may be
associated with the presence and severity of coronary heart disease
in patients with hyperlipidemia. Under physiological conditions,
miR-122 expression levels are low in myocardial tissue (12); however, elevated miR-122 plasma
levels have been reported in patients with acute heart failure
(25). In accordance with the
aforementioned studies, the present results revealed that the
expression levels of miR-122 were significantly upregulated in
cardiomyocytes of mice following the induction of AF compared with
control mice. miR-122 has been suggested to be implicated in the
maintenance of the differentiated state of tissues, as it may
participate in the establishment of tissue-specific gene expression
patterns (26). Aberrant
expression of miRNAs and the subsequent dysregulation of the
expression of their targets have been associated with the
proliferation and apoptosis of tumor cells (27). Notably, miR-122 has been
demonstrated to regulate the expression of target molecules
involved in various biological processes, including cellular
proliferation, differentiation and apoptosis (28). The present results revealed that
the apoptotic rate of AF cardiomyocytes was significantly decreased
following miR-122 silencing compared with untransfected AF
cardiomyocytes; similarly, cardiomyocyte viability in the AF group
appeared to be lower compared with in the miR-122 inhibitor group.
These results suggested that miR-122 may exert pro-apoptotic and
anti-proliferative effects on myocardial cells.
Apoptosis is a complex biological process that
enables multicellular organisms to discard unwanted cells during
their development (29,30). In order to further investigate the
involvement of miR-122 in apoptotic pathways, the expression of
apoptosis-related proteins was evaluated in cardiomyocytes. The
caspase and Bcl-2 protein families have been identified as key
mediators involved in apoptotic pathways in various types of cells
(31,32). Caspase-3 is a member of the caspase
family and has been identified as a pro-apoptotic protein. Ghavami
et al (33) revealed that
caspase-3 was activated in apoptotic cells via intrinsic and
extrinsic pathways. Bcl-x has been identified as an anti-apoptotic
protein, and its overexpression has been revealed to inhibit cell
death (34). Constitutive
activation of the epidermal growth factor receptor has been
reported to increase Bcl-x expression, thus suppressing apoptosis
(35). In the present study, Bcl-x
was demonstrated to be downregulated in cardiomyocytes isolated
from mice with AF, thus suggesting the activation of apoptotic
pathways. Notably, following miR-122 inhibition, Bcl-x expression
was upregulated, whereas caspase-3 expression was downregulated in
AF cardiomyocytes. These results suggested that miR-122 may be
implicated in the regulation of cardiomyocyte apoptosis in AF,
through the modulation of caspase-3 and Bcl-x expression.
ERKs are ubiquitous intracellular signaling
molecules, which have been implicated in the regulation of several
cellular pathways, including meiosis, mitosis and post-mitotic
processes in differentiated cells (36). ERK phosphorylation leads to the
activation of kinase activity. Previous studies have suggested that
aberrant ERK activation may promote cell death (37,38).
Therefore, the protein expression levels of ERK and p-ERK were
assessed in cardiomyocytes in the current study, in order to
further investigate the molecular mechanisms underlying the effects
of miR-122 in myocardial apoptotic pathways. The present results
revealed that the p-ERK/ERK ratio in the AF cardiomyocytes was
significantly increased following miR-122 silencing, compared with
untransfected AF cardiomyocytes, thus suggesting that miR-122 may
promote cardiomyocyte apoptosis through the inhibition of ERK
activation. Notably, ERK activity has previously been associated
with the upregulation of the pro-apoptotic caspase-3 and the
downregulation of the anti-apoptotic Bcl-x (37), in accordance with the
aforementioned results.
In conclusion, the results of the present study
suggested that miR-122 may be associated with the regulation of
cardiomyocyte apoptosis in AF. Therefore, miR-122 may have
potential as a novel biomarker for the diagnosis of AF, as well as
a possible therapeutic target for the development of novel
strategies for the treatment of patients with AF.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XZ made substantial contributions to conception and
design and was involved in drafting the manuscript. XZ and WJ
performed the experiments. WJ revised the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Krogh-Madsen T, Abbott GW and Christini
DJ: Effects of electrical and structural remodeling on atrial
fibrillation maintenance: A simulation study. PLoS Comput Biol.
8:e10023902012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xiao J, Liang D, Zhao H, Liu Y, Zhang H,
Lu X, Liu Y, Li J, Peng L and Chen YH: 2-Aminoethoxydiphenyl
borate, a inositol 1,4,5-triphosphate receptor inhibitor, prevents
atrial fibrillation. Exp Biol Med (Maywood). 235:862–868. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zoni-Berisso M, Lercari F, Carazza T and
Domenicucci S: Epidemiology of atrial fibrillation: European
perspective. Clin Epidemiol. 6:213–220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gaita F, Corsinovi L, Anselmino M,
Raimondo C, Pianelli M, Toso E, Bergamasco L, Boffano C, Valentini
MC, Cesarani F and Scaglione M: Prevalence of silent cerebral
ischemia in paroxysmal and persistent atrial fibrillation and
correlation with cognitive function. J Am Coll Cardiol.
62:1990–1997. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wolf PA, Abbott RD and Kannel WB: Atrial
fibrillation as an independent risk factor for stroke: The
framingham study. Stroke. 22:983–988. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jones DG, Haldar SK, Hussain W, Sharma R,
Francis DP, Rahman-Haley SL, McDonagh TA, Underwood SR, Markides V
and Wong T: A randomized trial to assess catheter ablation versus
rate control in the management of persistent atrial fibrillation in
heart failure. J Am Coll Cardiol. 61:1894–1903. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Benjamin EJ, Wolf PA, D'Agostino RB,
Silbershatz H, Kannel WB and Levy D: Impact of atrial fibrillation
on the risk of death The framingham heart study. Circulation.
98:946–952. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu D, Jickling GC, Ander BP, Hull H, Zhan
X, Dykstra-Aiello C, Stamova B and Sharp FR: Abstract W P93:
MiR-122 improves stroke outcomes after middle cerebral artery
occlusion in rats. Stroke. 46:AWP932015.
|
|
10
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Latronico MV and Condorelli G: MicroRNAs
and cardiac pathology. Nat Rev Cardiol. 6:418–429. 2009. View Article : Google Scholar
|
|
12
|
Lu Y, Zhang Y, Wang N, Pan Z, Gao X, Zhang
F, Zhang Y, Shan H, Luo X, Bai Y, et al: MicroRNA-328 contributes
to adverse electrical remodeling in atrial fibrillation.
Circulation. 122:2378–2387. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Girmatsion Z, Biliczki P, Bonauer A,
Wimmer-Greinecker G, Scherer M, Moritz A, Bukowska A, Goette A,
Nattel S, Hohnloser SH and Ehrlich JR: Changes in microRNA-1
expression and IK1 up-regulation in human atrial fibrillation.
Heart Rhythm. 6:1802–1809. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo X, Pan Z, Shan H, Xiao J, Sun X, Wang
N, Lin H, Xiao L, Maguy A, Qi XY, et al: MicroRNA-26 governs
profibrillatory inward-rectifier potassium current changes in
atrial fibrillation. J Clin Invest. 123:1939–1951. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li X, Yang Y, Wang L, Qiao S, Lu X, Wu Y,
Xu B, Li H and Gu D: Plasma miR-122 and miR-3149 potentially novel
biomarkers for acute coronary syndrome. PLoS One. 10:e01254302015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Beaumont J, López B, Hermida N, Schroen B,
San José G, Heymans S, Valencia F, Gómez-Doblas JJ, De Teresa E,
Díez J and González A: microRNA-122 down-regulation may play a role
in severe myocardial fibrosis in human aortic stenosis through
TGF-β1 up-regulation. Clin Sci (Lond). 126:497–506. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schrickel JW, Bielik H, Yang A, Schimpf R,
Shlevkov N, Burkhardt D, Meyer R, Grohé C, Fink K, Tiemann K, et
al: Induction of atrial fibrillation in mice by rapid
transesophageal atrial pacing. Basic Res Cardiol. 97:452–460. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Verheule S, Sato T, Everett T IV, Engle
SK, Otten D, Rubart-von der Lohe M, Nakajima HO, Nakajima H, Field
LJ and Olgin JE: Increased vulnerability to atrial fibrillation in
transgenic mice with selective atrial fibrosis caused by
overexpression of TGF-beta1. Circ Res. 94:1458–1465. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Haugan K, Lam HR, Knudsen CB and Petersen
JS: Atrial fibrillation in rats induced by rapid transesophageal
atrial pacing during brief episodes of asphyxia: A new in vivo
model. J Cardiovasc Pharmacol. 44:125–135. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
European Heart Rhythm Association; Heart
Rhythm Society, . Fuster V, Rydén LE, Cannom DS, Crijns HJ, Curtis
AB, Ellenbogen KA, Halperin JL, Le Heuzey JY, et al: ACC/AHA/ESC
2006 guidelines for the management of patients with atrial
fibrillation-executive summary: A report of the American College of
Cardiology/American Heart Association Task Force on Practice
Guidelines and the European Society of Cardiology Committee for
Practice Guidelines (Writing Committee to Revise the 2001
Guidelines for the Management of Patients With Atrial
Fibrillation). J Am Coll Cardiol. 48:854–906. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chang J, Nicolas E, Marks D, Sander C,
Lerro A, Buendia MA, Xu C, Mason WS, Moloshok T, Bort R, et al:
miR-122, a mammalian liver-specific microRNA, is processed from hcr
mRNA and may downregulate the high affinity cationic amino acid
transporter CAT-1. RNA Biol. 1:106–113. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fernández-Hernando C, Suárez Y, Rayner KJ
and Moore KJ: MicroRNAs in lipid metabolism. Curr Opin Lipidol.
22:86–92. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei G, He HW, Wang ZM, Zhao H, Lian XQ,
Wang YS, Zhu J, Yan JJ, Zhang DG, Yang ZJ and Wang LS: Plasma
levels of lipometabolism-related miR-122 and miR-370 are increased
in patients with hyperlipidemia and associated with coronary artery
disease. Lipids Health Dis. 11–55. 2012.PubMed/NCBI
|
|
25
|
Corsten MF, Dennert R, Jochems S,
Kuznetsova T, Devaux Y, Hofstra L, Wagner DR, Staessen JA, Heymans
S and Schroen B: Circulating MicroRNA-208b and MicroRNA-499 reflect
myocardial damage in cardiovascular disease. Circ Cardiovasc Genet.
3:499–506. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma L, Liu J, Shen J, Liu L, Wu J, Li W,
Luo J, Chen Q and Qian C: Expression of miR-122 mediated by
adenoviral vector induces apoptosis and cell cycle arrest of cancer
cells. Cancer Biol Ther. 9:554–561. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Thompson CB: Apoptosis in the pathogenesis
and treatment of disease. Science. 267:1456–1462. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fuchs Y and Steller H: Programmed cell
death in animal development and disease. Cell. 147:742–758. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Danial NN and Korsmeyer SJ: Cell death:
Critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Galluzzi L, Vitale I, Abrams JM, Alnemri
ES, Baehrecke EH, Blagosklonny MV, Dawson TM, Dawson VL, El-Deiry
WS, Fulda S, et al: Molecular definitions of cell death
subroutines: Recommendations of the nomenclature committee on cell
death 2012. Cell Death Differ. 19:107–120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ghavami S, Hashemi M, Ande SR, Yeganeh B,
Xiao W, Eshraghi M, Bus CJ, Kadkhoda K, Wiechec E, Halayko AJ and
Los M: Apoptosis and cancer: Mutations within caspase genes. J Med
Genet. 46:497–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nagane M, Levitzki A, Gazit A, Cavenee WK
and Huang HJ: Drug resistance of human glioblastoma cells conferred
by a tumor-specific mutant epidermal growth factor receptor through
modulation of Bcl-XL and caspase-3-like proteases. Proc Natl Acad
Sci USA. 95:5724–5729. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cagnol S and Chambard JC: ERK and cell
death: Mechanisms of ERK-induced cell death-apoptosis, autophagy
and senescence. FEBS J. 277:2–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu H, Jiang C, Li G and Lü J: PKB/AKT and
ERK regulation of caspase-mediated apoptosis by methylseleninic
acid in LNCaP prostate cancer cells. Carcinogenesis. 26:1374–1381.
2005. View Article : Google Scholar : PubMed/NCBI
|