Introduction
The term ‘missed abortion (MA)’, a type of
miscarriage, refers to a pregnancy in which there is fetal demise
without outside intervention, and also no uterine activity that may
expel the product of conception (POC) prior to 20 weeks of
gestation (1). Multiple
epidemiological factors, including parental or embryonic
chromosomal abnormalities, infection, immunological factors,
hereditary thrombophilia, uterine abnormalities, endocrinological
disorders, and nutritional and environmental factors have been
associated with miscarriage (2,3).
Cytogenetic analysis of the retained POC is thought to be the most
effective test for identifying the cause of MA (4). Chromosomal abnormalities are the
primary cause of MA, with errors in chromosome number, copy number
variations and abnormalities resulting in structural defects
accounting for 60–80% of MAs (4–8).
However, the cause of 20–40% of MAs remains unknown, despite
current detection methods.
High-throughput sequencing technology is currently
widely used for identifying genetic alterations associated with MAs
of unknown causes. Single gene defects in the POC may additionally
be associated with MA; however, few studies have used
high-throughput sequencing methods to study genetic defects in the
POC.
In the present study, 19 unrelated MA POCs were
collected and whole-exome sequencing (WES) was performed on the
POC. Bioinformatics analysis was performed on sequence variants
from a list of 286 selected candidate genes that are associated
with early embryonic lethality and MA. A total of 36 sequence
variants in 32 genes potentially associated with MA were identified
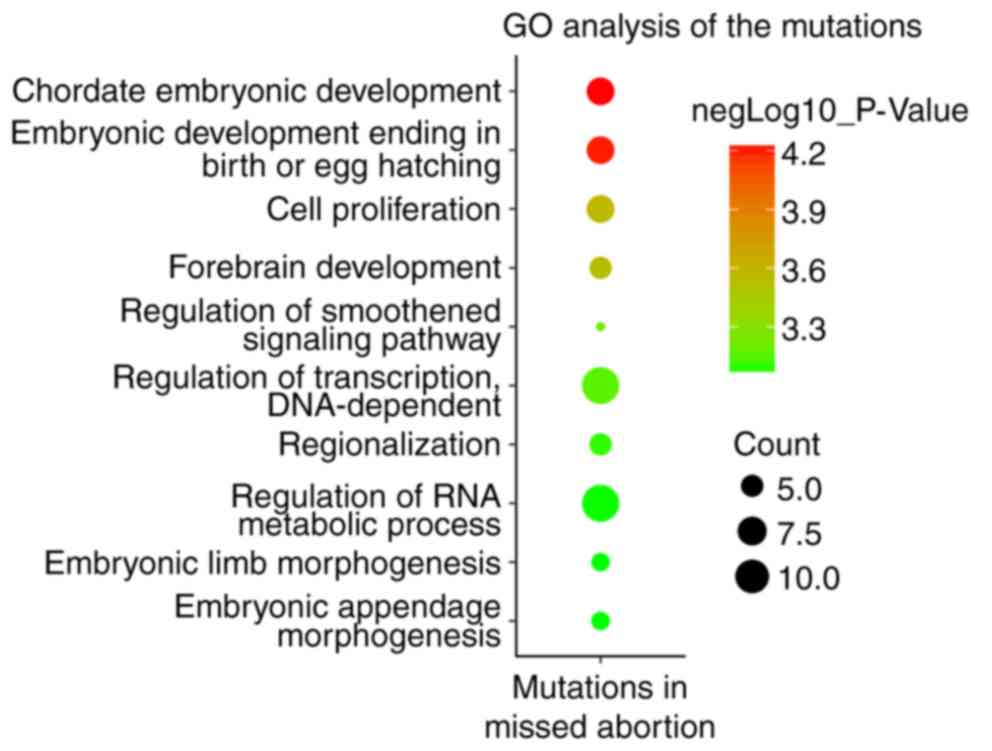
in 15 out of 19 POCs. Gene Ontology (GO) analysis suggested that
these genes were enriched in biological processes in early
embryonic development, including ‘chordate embryonic development’,
‘cell proliferation’ and ‘forebrain development’. The novel genes
and genetic alterations may increase knowledge of MA pathogenesis
and aid future genetic counseling for MA.
Materials and methods
Subjects
A total of 19 women (between 23 and 42 years of age)
participated in the present study, who experienced MA between 5–12
weeks of gestation and who were treated in the Department of
Obstetrics and Gynecology of the Chinese People's Liberation Army
(PLA) General Hospital (Beijing, China) between March 2017 and June
2017. Patient details are given in Table I. The inclusion criteria were as
follows: The participants with spontaneous abortions in early
pregnancy with unexplained etiology prior to the 12th week of
gestational age and lack of any successful pregnancy in the
previous history were included in the study. The exclusion criteria
were as follows: The patients with a history of risk factors,
including chronic infections, thrombosis, autoimmune diseases,
endocrinological disorders or genital malformation were not
included in the present study.
 | Table I.Clinical and sequencing features of
the patients and the embryos. |
Table I.
Clinical and sequencing features of
the patients and the embryos.
| Embryo ID | Patient age,
years | Gestational age,
weeks | Sequencing depth,
x | Karyotype analysis of
each embryo |
|---|
| QW001 | 30 | 6 | 170.95 | 46, XY |
| QW002 | 28 | 12 | 114.06 | 46, XX |
| QW003 | 28 | 7 |
96.61 | 46, XY |
| QW004 | 29 | 9 | 101.93 | 46, XX |
| QW005 | 30 | 7 |
92.99 | 46, XX |
| QW006 | 28 | 10 | 104.26 | 46, XY |
| QW007 | 30 | 8 | 106.04 | 46, XY |
| QW008 | 23 | 6 |
32.32 | 46, XX |
| QW009 | 42 | 14 | 131.72 | 46, XX |
| QW010 | 27 | 10 | 114.73 | 46, XX |
| QW011 | 28 | 8 | 124.83 | 46, XX |
| QW012 | 28 | 9 | 127.92 | 46, XY |
| QW013 | 28 | 8 | 116.69 | 46, XX |
| QW014 | 42 | 6 | 126.82 | 46, XY |
| QW015 | 32 | 5 |
90.60 | 46, XX |
| QW016 | 29 | 10 | 112.91 | 46, XY |
| QW017 | 31 | 7 |
87.46 | 46, XY |
| QW018 | 35 | 11 |
93.35 | 46, XY |
| QW019 | 24 | 8 |
93.06 | 46, XY |
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
Chinese PLA General Hospital's research committee and with the 1964
Helsinki declaration and its later amendments, or comparable
ethical standards. The present study was approved by the ethical
committee of Chinese People's Liberation Army (PLA) General
Hospital. Written informed consent was obtained from each
participant.
The present study was conducted on 19 preserved
chorionic villus samples with the normal chromosome number as
determined by next-generation sequencing (NGS). WES was performed
on chorionic villus genomic DNA from miscarriage samples of unknown
cause.
WES analysis
Each exome was captured using Roche Nimblegen SeqCap
EZ Exome v3.0 kit (Roche Applied Science, Madison, WI, USA)
according to the manufacturer's protocol. Subsequently, the
enriched exomes were sequenced using the Illumina HiSeq ×10
platform (Ilumina, Inc., San Diego, CA, USA). Reads were mapped
against the human reference genome hg38 (https://genome.ucsc.edu/index.html) using
Burrows-Wheeler Aligner (http://bio-bwa.sourceforge.net/). The single
nucleotide variants (SNV) were called by SAMTools (version 0.1.19,
http://samtools.sourceforge.net/) and
the Genome Analysis Toolkit (GATK, version 4.0,4.0, Broad
Institute, http://software.broadinstitute.org/gatk/), and ANNOVAR
(http://annovar.openbioinformatics.org/en/latest/)
was used for SNV annotation and filtering. Variants fulfilling the
following criteria were retained: i) Missense, nonsense,
frame-shift, or splice site variants; ii) absent in the dbSNP
(http://www.ncbi.nlm.nih.gov/snp/), 1000
Genomes (http://browser.1000genomes.org/index.html), ESP6500
(http://evs.gs.washington.edu/EVS/),
Exome Aggregation Consortium (ExAC; http://exac.broadinstitute.org/) and the Genome
Aggregation Database (gnomAD; http://gnomad.broadinstitute.org/) databases. Four
online functional prediction tools including Polyphen2 (http://genetics.bwh.harvard.edu/pph2/),
SIFT (http://sift.jcvi.org/), MutationTaster
(http://mutationtaster.org/) and
FATHMM-MKL (http://fathmm.biocompute.org.uk/fathmmMKL.htm), were
used to predict the variant effect on protein function. Constraint
Metrics Z score for missense variation (9), Loss Intolerance (pLI) (10) and Haploinsufficiency Score
(11) were used for evaluating the
haploinsufficiency effect of each gene. DECIPHER database
(https://decipher.sanger.ac.uk/) was used
for identifying the previous published copy number variations and
single nucleotide variants and the associated disease phenotypes.
DAVID Bioinformatics Resources 6.7 (https://david-d.ncifcrf.gov/) was used for conducting
the GO analysis.
Sanger sequencing validation
Sanger sequencing was used to validate the WES
results and verify whether the potentially disease-causing variants
identified were true variants or sequencing artifacts. Sanger
sequencing for the LDB1 variant in the POC QW013 was performed
using gene-specific primers as follows; the forward primer was
5′-AGGAGTGTCACAATGCTCAGATGAT-3′ and the reverse primer was
5′-GTAAACGGAGACTCAGATGGGAGAG-3′. Cycling parameters were an initial
denaturation at 94°C for 5 min followed by 35 cycles of
denaturation at 94°C for 20 sec, annealing at 60°C for 30 sec and
extension at 72°C for 1 min, followed by a final extension at 72°C
for 5 min. TransStart FastPfu DNA polymerase (TransGen Biotech Co.,
Ltd., Beijing, China) was used in the PCR reaction.
Results
WES analysis of MA embryos
A total of 19 patients (between 23 and 42 years of
age) with MA participated in the present study (Table I). A total of 19 MA POCs (chorionic
villus from 5–12 gestational weeks) were examined. The karyotypes
of all POC were normal (Table I).
WES was performed for each POC. The sequencing depth of each WES is
listed in Table I. Therefore,
WES-detected sequence variants identified in each embryo were
focused upon. The present study aimed to identify direct sequence
variants causing MA, which should not exist in live human beings.
Therefore, polymorphisms with a minor allele frequency absent in
the dbSNP, 1000 Genomes, ESP6500, ExAC and gnomAD databases were
retained. Subsequently, all variants were further filtered,
according to the list of 286 selected candidate genes that were
associated with early embryonic lethality and MA. A total of 36
sequence variants in 32 genes potentially associated with MA were
identified in 15 out of 19 patients (data not shown). All variants
were in the heterozygous state.
In silico analysis of the
variants
GO analysis suggested that these 32 genes were
enriched in biological processes in early embryonic development,
including ‘chordate embryonic development’, ‘cell proliferation’
and ‘forebrain development’ (Fig.
1). In silico analysis predicted that 12 of 36 variants were
considered to be pathogenic alleles by four online prediction
tools, including Polyphen-2, SIFT, Mutation Taster and FATHMM-MKL
(Table II). As all the variants
were heterozygous, the present study aimed to determine whether the
heterozygous state of the variants influenced disease tolerance.
The variation intolerance scores were analyzed using three scoring
systems, including the Constraint Metrics Z score for missense
variation (9), Loss Intolerance
(pLI) (10) and Haploinsufficiency
Score (11). Out of the 12 genes,
LIM domain binding 1 gene (LDB1) was the only gene that was
predicted as intolerant to variation by the three scoring systems
(Table II). Sequence variant
c.3064C>T; p.P1022S (Table II)
in another gene, death induced obliterator-1 (DIDO1), was
additionally a potential candidate gene causing MA. This variant
was predicted to be a pathogenic allele by Polyphen-2,
MutationTaster and FATHMM-MKL. The variant in DIDO1 was considered
loss-of-function-intolerant, as predicted by the Constraint Metrics
Z score for missense variation and pLI (Table II). Therefore, it was hypothesized
that the variant in LDB1 (c.662C>T; p.S221L) and DIDO1
(c.3064C>T; p.P1022S) was likely to be associated with embryo
lethality and MA.
| Table II.In silico analysis of sequence
variants demonstrated by whole-exome sequencing in the embryos from
cases of missed abortion. |
Table II.
In silico analysis of sequence
variants demonstrated by whole-exome sequencing in the embryos from
cases of missed abortion.
| Embryo ID | Gene | Reference mRNA
no. | Mutation type | Variants | Aa change | Polyphen-2 | SIFT | Mutation
Taster | FATHMM-MKL | M Z score | pLI | HI, % |
|---|
| QW001 | LIAS | NM_006859 | m | c.991T>C | p.W331R | D (0.997) | D (0) | D (1.0) | D (0.976) | 1.16 | 0.06 | 12.37 |
| QW002 | PADI6 | NM_207421 | m | c.122C>T | p.A41V | P (0.892) | N.A. | N.A. | N (0.422) | N.A. | N.A. | N.A. |
| QW004 | ATE1 | NM_007041 | m | c.929G>A | p.C310Y | P (0.855) | D (0.016) | D (1.0) | D (0.985) | −0.13 | 0.51 | 45.89 |
| QW005 | INTS1 | NM_001080453 | m | c.3934C>A | p.L1312M | B (0.183) | T (0.174) | D (0.82) | D (0.896) | 1.18 | 0.22 | 52.90 |
|
| PIKFYVE | NM_015040 | m | c.3683A>T | p.Q1228L | D (0.987) | T (0.245) | D (1.0) | D (0.989) | 1.58 | 0.98 | 20.31 |
|
| RAC1 | NM_018890 | m | c.230G>C | p.G77A | D (0.992) | T (0.137) | D (0.997) | D (0.962) | 3.42 | 0.57 | 0.72 |
|
| SCARB1 | NM_005505 | m | c.20C>A | p.A7E | D (0.997) | D (0.032) | N (1.0) | N (0.172) | 1.83 | 0.08 | 37.77 |
| QW007 | OTX2 | NM_172337 | m | c.475C>A | p.P159T | D (0.999) | D (0.002) | D (1.0) | D (0.985) | 1.02 | 0.74 | 0.71 |
| QW008 | BPTF | NM_182641 | m | c.2882T>A | p.I961K | B (0.017) | D (0.002) | D (0.999) | D (0.912) | 4.39 | 1.00 | 36.79 |
|
| CREBBP | NM_004380 | m | c.3107C>A | p.T1036K | B (0.008) | T (0.527) | D (0.981) | D (0.908) | 5.58 | 1.00 | 0.62 |
|
| HSF1 | NM_005526 | m | c.1462C>G | p.L488V | P (0.798) | D (0.007) | D (0.993) | D (0.938) | 0.54 | 0.59 | 48.62 |
|
| NF1 | NM_000267 | m | c.1648C>A | p.L550M | D (1.0) | T (0.092) | D (0.999) | D (0.899) | 6.22 | 1.00 | 0.87 |
|
| PIKFYVE | NM_015040 | m | c.3307A>G | p.K1103E | D (0.997) | T (0.222) | D (1.0) | D (0.998) | 1.58 | 0.98 | 20.31 |
|
| PTCH1 | NM_000264 | m | c.3470C>A | p.A1157E | D (0.973) | T (0.071) | D (1.0) | D (0.991) | 2.86 | 1.00 | 0.48 |
|
| RAPGEF2 | NM_014247 | m | c.3203T>A | p.V1068E | B (0.243) | T (0.319) | D (1.0) | D (0.993) | 3.22 | 1.00 | 21.18 |
| QW010 | RGS14 | NM_006480 | m | c.510C>G | p.S170R | D (1.0) | D (0) | D (1.0) | D (0.916) | 1.15 | 0.02 | 61.07 |
|
| TRIM28 | NM_005762 | m | c.361A>C | p.K121Q | D (0.997) | T (0.172) | D (1.0) | D (0.604) | 3.16 | 1.00 | 27.80 |
| QW011 | PTPRB | NM_001109754 | m | c.5561T>C | p.V1854A | B (0.002) | D (0.034) | N (0.97) | D (0.838) | 0.49 | 0.94 | 39.33 |
| QW012 | DIDO1 | NM_033081 | m | c.3064C>T | p.P1022S | P (0.759) | T (0.177) | D (0.929) | D (0.963) | 3.10 | 1.00 | 83.16 |
|
| KIF16B | NM_001199866 | m | c.3802G>T | p.V1268F | B (0.007) | D (0) | D (0.864) | N (0.096) | 0.69 | 0.00 | 44.93 |
|
| ZNF568 | NM_198539 | s | c.514A>T | p.R172X | N.A. | N.A. | D (1.0) | N (0.066) | 0.27 | 0.00 | 88.58 |
| QW013 | FAM208A | NM_015224 | m | c.1616A>G | p.H539R | D (0.991) | T (0.177) | D (1.0) | D (0.883) | 0.89 | 1.00 | 29.50 |
|
| KAT2A | NM_021078 | m | c.671C>A | p.P224H | D (1.0) | D (0) | D (1.0) | D (0.98) | 4.79 | 0.41 | 22.83 |
|
| KDM1A | NM_015013 | m | c.1759A>T | p.N587Y | D (1.0) | T (0.051) | D (1.0) | D (0.991) | 5.56 | 0.99 | 5.56 |
|
| LDB1 | NM_003893 | m | c.662C>T | p.S221L | D (0.985) | D (0.001) | D (1.0) | D (0.977) | 3.24 | 0.88 | 2.24 |
|
| NOTCH1 | NM_017617 | m | c.2953C>G | p.P985A | D (0.969) | T (0.352) | D (1.0) | D (0.966) | 4.48 | 1.00 | 0.15 |
|
| POGLUT1 | NM_152305 | m | c.832T>G | p.F278V | D (1.0) | D (0.001) | D (1.0) | D (0.937) | 1.08 | 0.00 | 20.35 |
| QW014 | CDH5 | NM_001795 | s | c.1138C>T | p.Q380X | N.A. | N.A. | A (1.0) | D (0.736) | 0.02 | 0.15 | 32.63 |
|
| GAS1 | NM_002048 | m | c.610C>T | p.R204C | D (1.0) | D (0.002) | D (1.0) | D (0.772) | 4.27 | 0.62 | 29.14 |
| QW015 | DLX3 | NM_005220 | m | c.314C>T | p.A105V | B (0.037) | T (0.119) | D (0.987) | D (0.918) | 1.87 | 0.01 | 40.57 |
|
| PTPRB | NM_001109754 | m | c.359T>C | p.V120A | B (0.39) | D (0.007) | N (1.0) | D (0.788) | 0.49 | 0.94 | 39.33 |
| QW016 | INTS1 | NM_001080453 | m | c.6475C>T | p.L2159F | D (0.999) | D (0.006) | D (1.0) | D (0.963) | 1.18 | 0.22 | 52.90 |
|
| SALL4 | NM_020436 | m | c.733C>A | p.H245N | B (0.067) | T (0.16) | D (1.0) | D (0.865) | 1.66 | 1.00 | 38.00 |
| QW018 | SRRT | NM_015908 | m | c.1148A>C | p.K383T | D (0.999) | T (0.249) | D (1.0) | D (0.987) | 4.61 | 0.98 | 30.36 |
|
| VPS26A | NM_004896 | m | c.758C>T | p.A253V | B (0.388) | D (0.019) | D (1.0) | D (0.996) | 0.94 | 0.66 | 11.04 |
| QW019 | CREBBP | NM_004380 | m | c.2917C>A | p.P973T | P (0.952) | T (0.055) | D (1.0) | D (0.992) | 5.58 | 1.00 | 0.62 |
Analysis of the variant in LDB1
LDB1 serves important roles in the regulation of a
variety of processes in early embryonic development, including
heart formation (12), head and
brain development (12–17), limb patterning (18), and eye development (16). In previous studies, the development
of LDB1 null mutant mice was arrested at embryonic day 8.5 (E8.5)
and mice succumbed at E9-E10 (12,19).
Therefore, LDB1 may be a good candidate gene for human early
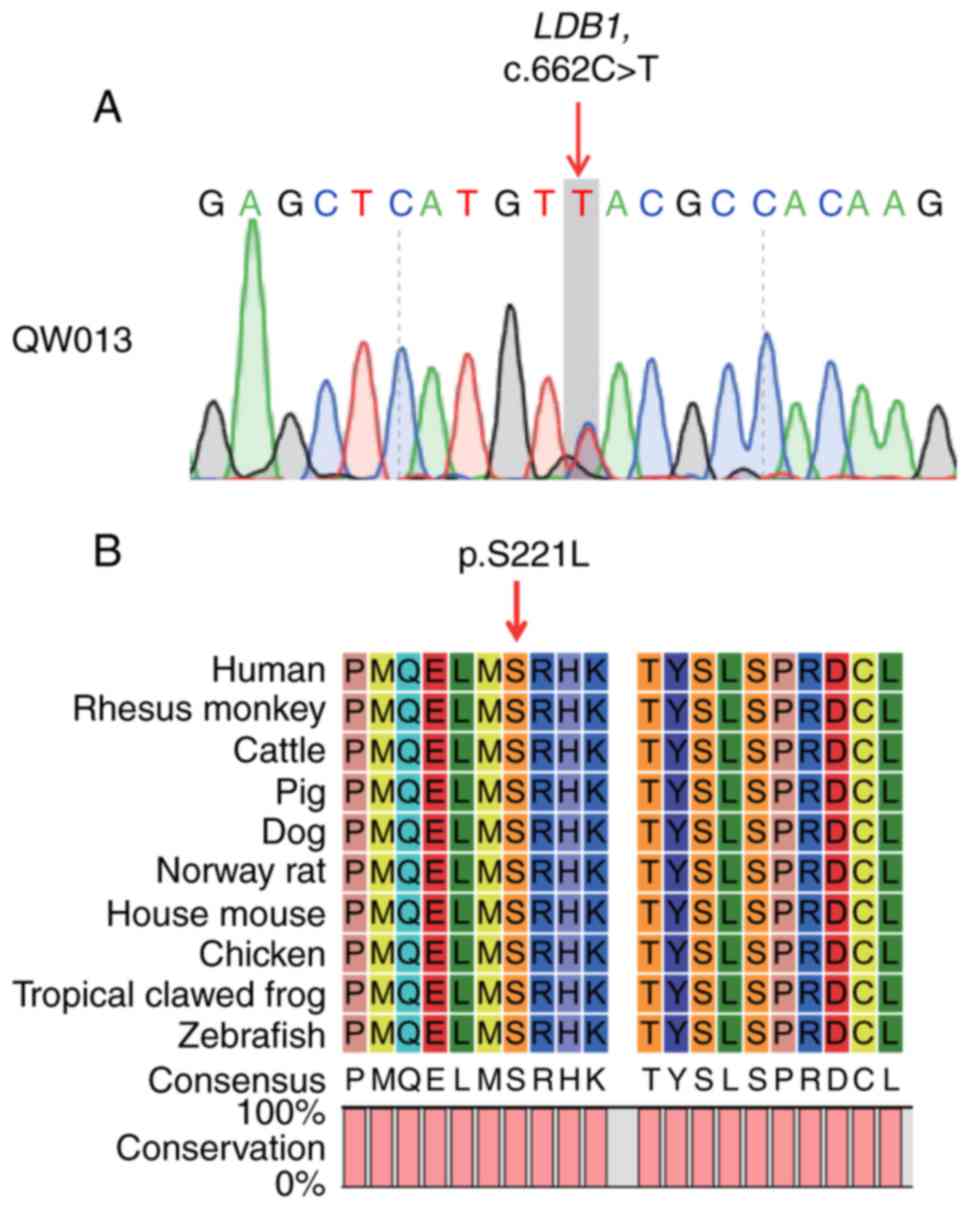
embryonic lethality and MA. In the present study, the heterozygous
c.662C>T variant in LDB1 was also validated by Sanger sequencing
(Fig. 2A). DNA samples from the
patient and her husband were unavailable, therefore it was not
possible to analyze whether this variant was inherited or generated
de novo. This variant and the flanking region were additionally
highly conserved in humans and other animals including zebrafish
(Fig. 2B), which reflects the
highly conserved role of LDB1 in early embryonic development across
different species.
Discussion
In the present study, WES was performed on 19 POC
and using a strict filtering strategy, 36 rare sequence variants
associated with MA were identified. GO analysis suggested that
these 32 genes were enriched in biological processes in early
embryonic development, including ‘chordate embryonic development’,
‘cell proliferation’ and ‘forebrain development’. Further strict
in silico bioinformatics analysis predicted the LDB1
(c.662C>T; p.S221L) variant to be a highly pathogenic
variant.
Previous studies suggest that chromosomal
abnormalities account for 60–80% of MA cases (4–8).
However, to the best of the authors' knowledge, no studies have
been performed on POC using NGS technology to elucidate single gene
defects. Therefore, the present study attempted to dissect the
genetic causes of the remaining 20–40% of MA cases. A total of 36
rare sequence variants in 32 genes potentially associated with MA
were identified in 15 out of 19 patients. Of the 32 genes, seven
genes [lycine acetyltransferase 2A, lycine demethylase 1A
(KDM1A), spalt like transcription factor 4, heat shock
transcription factor 1, integrator subunit complex 1, patched 1 and
growth arrest specific 1] were significantly enriched in the
biological process of ‘chordate embryonic development’, and five
genes (KDM1A, notch 1, neurofibromin 1, orthodenticle
homeobox 2 and Rac family small GTPase 1) were enriched in
‘forebrain development’. The embryos succumbed or arrested if each
of the above genes were knocked out.
LDB1 is a critical gene involved in embryonic
morphogenesis (12–17,19).
LDB1 gene deficiency leads to early embryonic arrest and
embryo loss between E9-E10 (12,19).
Therefore, LDB1 is a good candidate gene for embryonic
lethality. Additionally, LDB1 was searched for in Online
Mendelian Inheritance in Man (OMIM) and PubMed; however, no study
demonstrating LDB1 gene mutations associated with
developmental diseases was identified. Therefore, it was
hypothesized that human embryos harboring pathogenic mutations in
LDB1 may lead to embryonic lethality, meaning that no child
carrying the LDB1 pathogenic mutation would be born; this
may explain why no LDB1 mutation was observed in OMIM.
Furthermore, in the present study, the c.662C>T; p.S221L variant
in LDB1 was predicted as a pathogenic allele by a number of
prediction tools, which, along with the extreme rarity and
conservation of the variant and flanking regions, suggested that
the LDB1 mutation in the POC was associated with MA.
Ablation of DIDO1 in mice causes embryonic
lethality during the gastrulation stage (20). DIDO heterozygous deletion
mice demonstrate abnormalities in their spleen, bone marrow and
peripheral blood (21). By
checking the DECIPHER database (https://decipher.sanger.ac.uk/), two heterozygous
missense variants in DIDO1 were identified, one of which may
be associated with abnormalities in the head, cardiovascular
system, ear, integument, nervous system and skeletal system.
Therefore, the variant in the present study may also be associated
with embryonic developmental abnormalities.
In conclusion, the present study identified 36 rare
sequence variants in 19 POCs associated with MA. Further
bioinformatics analysis predicted that the LDB1
(c.662C>T; p.S221 L) variant is a highly pathogenic variant and
may be associated with embryonic lethality. Taken together, the
results of the present study provide researchers and clinicians
with a better understanding of the etiology and molecular mechanism
of human embryonic lethality and MA.
Acknowledgements
The authors would like to thank the participants in
the present study for their consent and support to publish this
article.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81571411) and the
Military Medical Innovation Project (grant no. 16JS011).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MF and SM conducted the experiments; MF, CW, SJ and
LL performed the WES data analysis and in silico analysis of
sequence variants; MF, SJ, YM, SM and CW collected the POC samples;
and YM and HP designed the experiments and wrote the manuscript.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
Chinese PLA General Hospital's research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. This research was approved by the ethical committee of
Chinese People's Liberation Army (PLA) General Hospital. Written
informed consent was obtained from each participant.
Patient consent for publication
Written informed consent was obtained from each
participant.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Griebel CP, Halvorsen J, Golemon TB and
Day AA: Management of spontaneous abortion. Am Fam Physician.
72:1243–1250. 2005.PubMed/NCBI
|
|
2
|
Clifford K, Rai R, Watson H and Regan L:
An informative protocol for the investigation of recurrent
miscarriage: Preliminary experience of 500 consecutive cases. Hum
Reprod. 9:1328–1332. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hatasaka HH: Recurrent miscarriage:
Epidemiologic factors, definitions, and incidence. Clin Obstet
Gynecol. 37:625–634. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Segawa T, Kuroda T, Kato K, Kuroda M, Omi
K, Miyauchi O, Watanabe Y, Okubo T, Osada H and Teramoto S:
Cytogenetic analysis of the retained products of conception after
missed abortion following blastocyst transfer: A retrospective,
large-scale, single-centre study. Reprod Biomed Online. 34:203–210.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alberman ED and Creasy MR: Frequency of
chromosomal abnormalities in miscarriages and perinatal deaths. J
Med Genet. 14:313–315. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang HK, Luo FW, Geng Q, Li J, Liu QZ,
Chen WB, Li F and Xie JS: Analysis of fetal chromosomal karyotype
and etiology in 252 cases of early spontaneous abortion. Zhonghua
Yi Xue Yi Chuan Xue Za Zhi. 28:575–578. 2011.(In Chinese).
PubMed/NCBI
|
|
7
|
Muñoz M, Arigita M, Bennasar M, Soler A,
Sanchez A and Borrell A: Chromosomal anomaly spectrum in early
pregnancy loss in relation to presence or absence of an embryonic
pole. Fertil Steril. 94:2564–2568. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Philipp T, Philipp K, Reiner A, Beer F and
Kalousek DK: Embryoscopic and cytogenetic analysis of 233 missed
abortions: Factors involved in the pathogenesis of developmental
defects of early failed pregnancies. Hum Reprod. 18:1724–1732.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Samocha KE, Robinson EB, Sanders SJ,
Stevens C, Sabo A, McGrath LM, Kosmicki JA, Rehnström K, Mallick S,
Kirby A, et al: A framework for the interpretation of de novo
mutation in human disease. Nat Genet. 46:944–950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lek M, Karczewski KJ, Minikel EV, Samocha
KE, Banks E, Fennell T, O'Donnell-Luria AH, Ware JS, Hill AJ,
Cummings BB, et al: Analysis of protein-coding genetic variation in
60,706 humans. Nature. 536:285–291. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang N, Lee I, Marcotte EM and Hurles ME:
Characterising and predicting haploinsufficiency in the human
genome. PLoS Genet. 6:e10011542010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mukhopadhyay M, Teufel A, Yamashita T,
Agulnick AD, Chen L, Downs KM, Schindler A, Grinberg A, Huang SP,
Dorward D and Westphal H: Functional ablation of the mouse Ldb1
gene results in severe patterning defects during gastrulation.
Development. 130:495–505. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim S, Zhao Y, Lee JM, Kim WR, Gorivodsky
M, Westphal H and Geum D: Ldb1 is essential for the development of
isthmic organizer and midbrain dopaminergic neurons. Stem Cells
Dev. 25:986–994. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao Y, Flandin P, Vogt D, Blood A,
Hermesz E, Westphal H and Rubenstein JL: Ldb1 is essential for
development of Nkx2.1 lineage derived GABAergic and cholinergic
neurons in the telencephalon. Dev Biol. 385:94–106. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao Y, Kwan KM, Mailloux CM, Lee WK,
Grinberg A, Wurst W, Behringer RR and Westphal H: LIM-homeodomain
proteins Lhx1 and Lhx5, and their cofactor Ldb1, control Purkinje
cell differentiation in the developing cerebellum. Proc Natl Acad
Sci USA. 104:13182–13186. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Plautz CZ, Zirkle BE, Deshotel MJ and
Grainger RM: Early stages of induction of anterior head ectodermal
properties in Xenopus embryos are mediated by transcriptional
cofactor ldb1. Dev Dyn. 243:1606–1618. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Costello I, Nowotschin S, Sun X, Mould AW,
Hadjantonakis AK, Bikoff EK and Robertson EJ: Lhx1 functions
together with Otx2, Foxa2, and Ldb1 to govern anterior mesendoderm,
node, and midline development. Genes Dev. 29:2108–2122. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tzchori I, Day TF, Carolan PJ, Zhao Y,
Wassif CA, Li L, Lewandoski M, Gorivodsky M, Love PE, Porter FD, et
al: LIM homeobox transcription factors integrate signaling events
that control three-dimensional limb patterning and growth.
Development. 136:1375–1385. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L, Lee JY, Gross J, Song SH, Dean A and
Love PE: A requirement for Lim domain binding protein 1 in
erythropoiesis. J Exp Med. 207:2543–2550. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Futterer A, Raya A, Llorente M,
Izpisúa-Belmonte JC, de la Pompa JL, Klatt P and Martínez-A C:
Ablation of Dido3 compromises lineage commitment of stem cells in
vitro and during early embryonic development. Cell Death Differ.
19:132–143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fütterer A, Campanero MR, Leonardo E,
Criado LM, Flores JM, Hernández JM, Miguel San JF and Martínez-A C:
Dido gene expression alterations are implicated in the induction of
hematological myeloid neoplasms. J Clin Invest. 115:2351–2362.
2005. View
Article : Google Scholar : PubMed/NCBI
|