Introduction
Intervertebral disc degeneration (IDD) is one of the
major causes of low back pain (LBP), a condition associated with
high morbidity and disability. As a result, LBP has become a
significant financial burden worldwide (1). An intervertebral disc consists of
three parts: The nucleus pulposus (NP); the annulus fibrosus and
the endplate cartilage. The NP is located in the center of the disc
and consists of a mixture of nucleus pulposus cells (NPCs) and
extracellular matrix, while the outer annulus fibrosus is mainly
composed of collagen fibers (2).
The NP is the main tissue responsible for axial compression of the
spine in the intervertebral disc. As the number of NPCs within a
disc decreases and functional capability is lost, the extracellular
matrix, which features proteoglycan and type II collagen, becomes
degraded. This leads to an increase in the synthesis of type I
collagen which ultimately causes discs to degenerate (3). Previous studies have shown that IDD
is associated with aging and that spinal disease is an age-related
disease. Furthermore, accumulating evidence demonstrates that the
senescence of NPCs, as induced by different forms of stress, plays
a crucial role in the progression of IDD (4). As NPCs become senescent, their
ability to proliferate is impeded and their functional capability
is lost (5).
Over recent years, an increasing number of studies
have reported that IDD is associated with osteoporosis, although
the association remains controversy (6). For example, Luo et al
(6) identified a close
relationship between IDD and osteoporosis in ovariectomized rats,
and speculated that the mechanism involved was related to the
integrity and function of adjacent structures of intervertebral
discs in the spine. Adjacent segment disc degeneration is very
common in women with osteoporosis, thus indicating that
osteoporosis might represent one of the causative agents of IDD
(7). Research has shown that the
endplate of patients with osteoporosis is under high pressure and
that this can cause endplate calcification. This structural anomaly
exerts influence upon the nutritional infiltration of the end
plate, resulting in malfunction of the nutritional supply to
intervertebral discs and, ultimately, to IDD (8).
Parathyroid hormone (PTH) is widely used in the
treatment of osteoporosis because it is known to promote the
synthesis of bone (9). PTH is also
known to regulate the synthesis and metabolism of intervertebral
disc cells and inhibit potential calcification molecules via the
mitogen-activated protein kinase (MAPK) and protein kinase A (PKA)
signaling pathways (9). It has
also been reported that PTH cannot only promote anabolic
metabolism, but can also inhibit degradation of the extracellular
matrix within intervertebral discs (7). Indeed, animal experiments have
established that PTH can relieve disc degeneration in rat models of
osteoporosis (7). Therefore, PTH
has potential clinical significance for the treatment of disc
degeneration in patients with osteoporosis, although the exact
mechanism involved remains unclear. Previous studies have shown
that PTH can promote autophagy in osteoblasts and chondrocytes
(10) and can also alleviate
osteoarthritis by activating autophagy in articular chondrocytes
(11). However, the effect of PTH
upon autophagy in NPCs has yet to be evaluated. Dexamethasone
(DXM), a type of glucocorticoid, is commonly used in clinical
scenarios and is known to cause osteoporosis (12). In the present study, DXM and PTH
were used to treat NPCs. We then investigated the levels of
senescence and autophagy in order to determine the effect of PTH on
NPCs treated with DXM, and also to analyze the potential
involvement of autophagy and the mTOR pathway.
Materials and methods
Ethical approval
The experimental protocol of this study was approved
by the Animal Ethics Committee of the Affiliated Hospital of Hebei
University of Engineering (Handan, China).
Reagents and antibodies
Alcian dye, rat-derived recombinant PTH and type II
collagenase were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). Primary antibodies against LC3b, Beclin-1,
P62, ATG5, type II collagen, SOX-9 and β-actin were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA). β-galactosidase
staining kits, immunofluorescence reagents, western blotting
reagents, DAPI, and cell protein extraction kits were purchased
from Beyotime Institute of Biotechnology (Shanghai, China).
Finally, cell culture reagents, including Dulbecco's modified
Eagle's medium (DMEM), fetal bovine serum (FBS) and 0.25% trypsin
were purchased from Gibco (Thermo Fisher Scientific, Inc., Waltham,
MA, USA).
Culturing rat primary NPCs
Thirty male Sprague-Dawley rats, aged 3 months and
weighing approximately 250–300 g were obtained from SLAC Laboratory
Animal Company (Shanghai, China) and treated as described
previously (10). In brief, after
comforting, the dorsal anatomy was dissected under aseptic
conditions and the entire lumbar spine (L1-L6) was dissected and
transferred to a clean bench. Each segment of NP (which had a
jelly-like appearance) was carefully removed under a microscope and
transferred to a centrifuge tube. Collagenase type II was then
treated with 0.1% collagenase for 4 h at 37°C and filtered through
a 200 mesh filter. Cells were then cultured in DMEM medium
containing 10% FBS and 1% penicillin in an incubator at 37°C and 5%
CO2. In order to prevent the dedifferentiation of NPCs,
all cells in the follow-up experiments were all second
generation.
Experimental design
NPCs were treated with 1,25 and 50 µg/ml of DXM for
48 h. The effect of PTH on autophagy and the mTOR signaling pathway
in NPCs was analyzed by dividing cells into four treatment groups:
A Control group; DXM group; PTH group; and a DXM + PTH group. All
treatments were administered for 48 h. The DXM + PTH group were
first treated with 25 µg/ml DXM for 1 h; 10 nM PTH was then added
for the remainder of the48 h study period. The effect of autophagy
on PTH-induced NPC-induced NPC senescence was also analyzed by
dividing cells into five treatment groups: A control group; DXM
group; DXM + PTH group; DXM + PTH + Control siRNA group; and a DXM
+ PTH + ATG5 siRNA group. All treatments were administered for 48
h.
Alcian staining
NPCs were plated on 24-well plates at a cell density
of 70–80% and experiments were performed overnight. Cells were
fixed with 4% paraformaldehyde for 10 min and washed three times
with PBS. Then, cells were added with 1 ml of Alcian dye (prepared
with acetic acid) and incubated at room temperature for 30 mins.
Finally, cells were observed under an inverted microscope
(magnification, ×10).
Cellular immunofluorescence
Following overnight culture in a 24-well plate,
cells were fixed with 4% paraformaldehyde for 10 min and then
treated with 0.2% Triton X-100 for 15 min. After blocking with 5%
goat serum for 30 min, cells were incubated overnight at 4°C in a
refrigerator with primary antibodies against type II collagen and
SOX-9 primary (1:100). The following morning, cells were incubated
with Alexa Fluor-555 labeled secondary antibody (Beyotime Institute
of Biotechnology) for 2 h at room temperature, stained for 5 min at
room temperature with DAPI (4′,6-diamidino-2-phenylindole)
(Beyotime Institute of Biotechnology) and finally observed under a
fluorescence microscope (Olympus Corporation, Tokyo, Japan).
Measurement of cell surface area
NPCs were treated with different concentrations of
DXM for 48 h and then observed under a microscope and photographed
with a 20X objective lens. Then, at least six cells were selected
from each image with Image-Pro Plus software (Carlsbad, CA, USA)
and measured using the ‘Area Measure’ function. At least six images
from six fields were selected for each treatment group, with at
least six cells selected for each image.
β-galactosidase staining
NPCs were treated with DXM at 25 and 50 µg/ml for 48
h and then treated with β-galactosidase (senescence-associated
β-galactosidase, SA-β-gal) at room temperature for 15 min, in
accordance with the manufacturer's instructions. After washing
three times with PBS, the cells was stained with SA-β-gal for 12 h
at room temperature. After washing with PBS, the cells were finally
observed under an inverted microscope at ×200, and the proportion
(%) of positive cells present in the total cell population was
determined.
Western blotting
Radio-Immunoprecipitation Assay (RIPA, Pikungen,
Shanghai), containing 1% phenylmethanesulfonyl fluoride (PMSF) was
used to extract total cellular protein and the bicinchoninic acid
(BCA) method used to determine protein concentration. Extracted
total proteins were then separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred onto polyvinylidene difluoride (PVDF) membranes
(Beyotime Institute of Biotechnology) using a wet-spinning method.
After blocking with 5% non-fat milk for 2 h at room temperature,
membranes were washed with TBST for 3 times and then incubated with
LC3b, Beclin-1, P62 and ATG5 primary antibodies (1:1,000; Cell
Signaling Technology, Inc.) overnight at 4°C. After 3 washes, the
membranes were incubated with horseradish peroxidase-labeled
secondary antibody for 2 h at room temperature. Finally, membranes
were treated with ECL luminescence (Thermo Fisher Scientific, Inc.)
and positive immune-staining was detected on an imaging system
(PerkinElmer, Inc., Waltham, MA, USA). Semi-quantitative analysis
of protein bands was performed using AlphaEaseFC 4.0 software
(Alpha Innotech Corp., San Leandro, CA, USA).
ATG5 siNRA transcription
The specific design of ATG5 siRNA is shown in
Table I (Shanghai GenePharma Co.,
Ltd., Shanghai, China). 1 ml NPCs with a density of
2×105/ml were added to 6-well plates and cultured
overnight. Cells were transfected using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.), in accordance
with the manufacturer's protocol. At 48 h after transfection, cells
were treated with DXM and PTH as required by the experimental
design. The control RNA was a non-specific, non-targeting siRNA.
Following transfection, total cellular protein was extracted and
analyzed by western blotting.
 | Table I.Sequences of ATG5 siRNA. |
Table I.
Sequences of ATG5 siRNA.
| Primer | Direction | Sequence 5′-3 |
|---|
| ATG5 | Sense |
GGCCUUUCAUUCAGAAGCUTT |
|
| Antisense |
AGCUUCUGAAUGAAAGGCCTT |
| Negative | Sense |
UUCUCCGAACGUGUCACGUTT |
| control | Antisense |
ACGUGACACGUUCGGAGAATT |
GFP-LC3 transfection
The GFP-LC3 adenovirus with 1010/ml titer
was obtained from Han Heng Biology (Shanghai, China). According to
our previous study (13), rat NPCs
were cultured on a glass bottom dish overnight and transfected with
adenovirus with 100 multiplicity of infection (MOI). According to
the 100 MOI and 1–2×106 cells in each well, cells were
treated with 20 µl and 2×108 virus, which was diluted
into 1 ml serum-free medium. NPCs were cultured with virus for 2 h
in a 37°C incubator and then treated with 2 ml normal medium
overnight. Then, the transfection efficiency was detected under
fluorescence microscopy. Autophagosomes were observed using a laser
confocal microscopy (Leica TCS, SP8; Leica Microsystems GmbH,
Wetzlar, Germany) after treatments as the experimental design
described.
Statistical analysis
All experiments were replicated in triplicate and
statistically analyzed by SPSS 15 software package (SPSS, Inc.,
Chicago, IL, USA). Differences between groups were analyzed by
one-way analysis of variance (ANOVA). If ANOVA proved to be
statistically significant, then we also used the least significant
difference (LSD) method for pairwise analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of rat primary NPCs in
culture
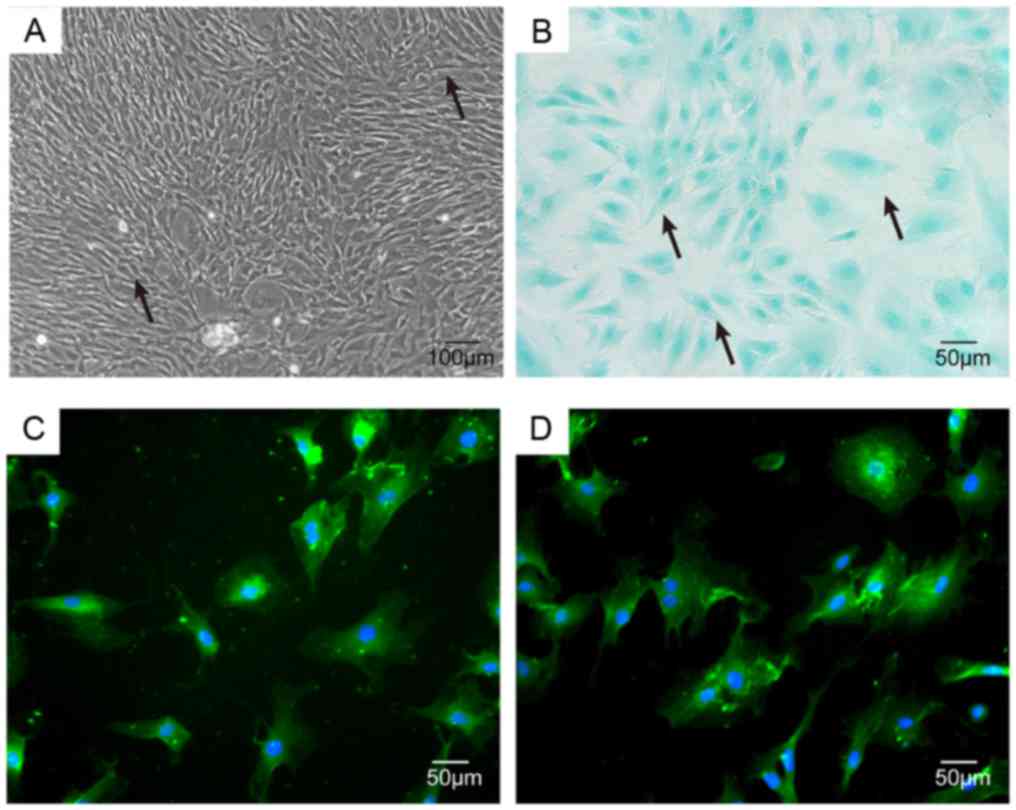
NP cells were identified as having apolygonal or
star-shaped appearance under inverted phase contrast microscopy,
and abundant particles were observed in the cytoplasm of NPCs
(Fig. 1A). Notochord cells
containing large number of vacuoles also could be observed. NPCs
treated by Alcian staining were light blue in coloration (Fig. 1B) due to the high cellular content
of proteoglycan. Further immunofluorescence studies showed that the
cytoplasm of NPCs contained large amounts of type II collagen and
SOX-9, thus confirming that NPCs exhibit cartilage-like
characteristics (Fig. 1C and
D).
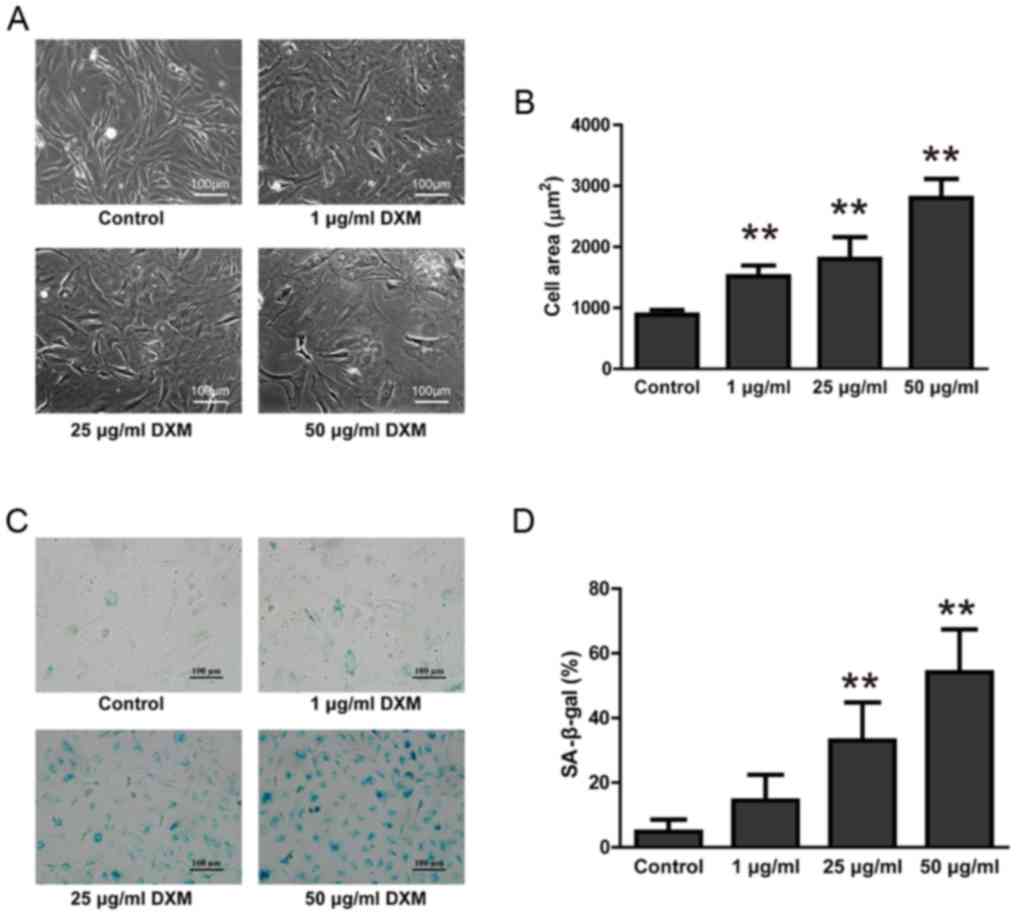
DXM promoted NPC senescence
After treatment with 1, 25 and 50 µg/ml of DXM for
48 h, the cell size of NPCs was significantly increased,
particularly in the 50 µg/ml group (P<0.05). NPC nuclei were
enlarged and the cytoplasm was stretched flat, creating a ‘fried
egg-like’ appearance (Fig. 2A).
Calculations of cell surface area also demonstrated that
glucocorticoid treatment significantly increased the surface area
of NPCs (P<0.05; Fig. 2B). The
senescence of NPCs was further confirmed by SA-β-gal staining. The
cytoplasm of NPCs treated with 25 and 50 µg/ml DXM were more
intensively stained than controls. Semi-quantitative analysis
further found that the proportion of NPCs in these two groups that
were positively stained with SA-β-gal was significantly increased
(P<0.05; Fig. 2C and D),
indicating that NPCs treated with 25 and 50 µg/ml DXM were
senescent.
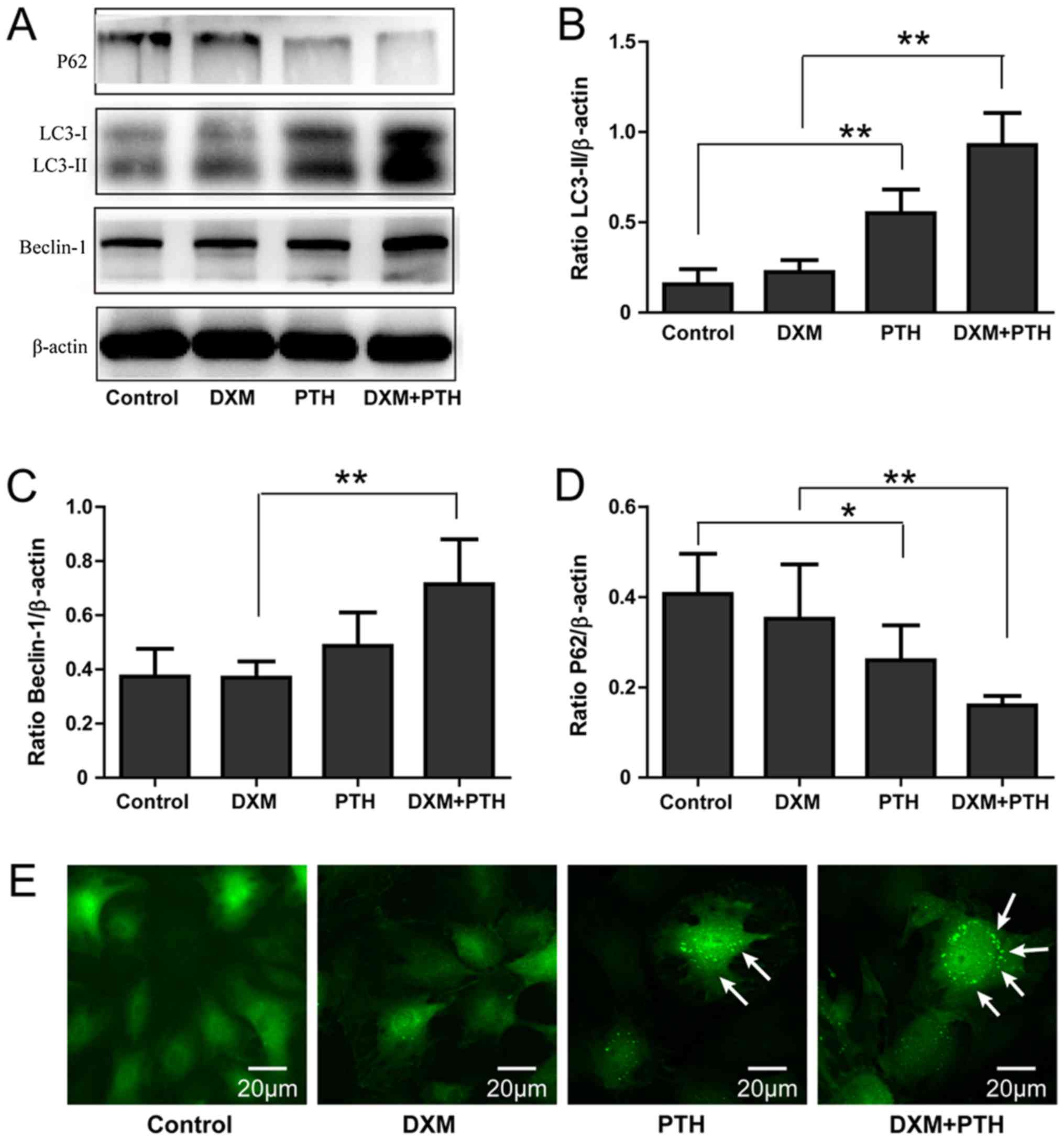
The effect of PTH on autophagy levels
in NPCs
To analyze the effect of PTH on autophagy in NPCs,
we used western blotting and GFP-LC3 assay to analyze the protein
expression levels of LC3-II, Beclin-1 and P62, as well as
autophagosomes. Compared with the control group, the administration
of 10 nM PTH led to a significant increase in the levels of LC3-II
protein in NPCs 48 h later (P<0.05). The levels of LC3-II in
NPCs following DXM + PTH treatment was significantly higher than
that of DXM alone (P<0.05), indicating that PTH cannot only
induce NPCs to produceLC3-II protein but can also lead to increased
expression of LC3-II in cells treated with DXM (P<0.05; Fig. 3A and B). Changes in Beclin-1 levels
were similar to those observed with LC3-II (P<0.05; Fig. 3A and C). Since P62 is a
‘transporter’ of autophagic degradation-specific proteins, P62
levels gradually decline with increasing PTH concentrations, thus
confirming the autophagy of NPCs under PTH-activated DXM
(P<0.05; Fig. 3A and D).
Compared with control and DXM group, GFP-LC3 analysis showed that
PTH significantly enhanced the number of LC3-positive cells and the
number of LC3-positive green dots in one cell (Fig. 3E).
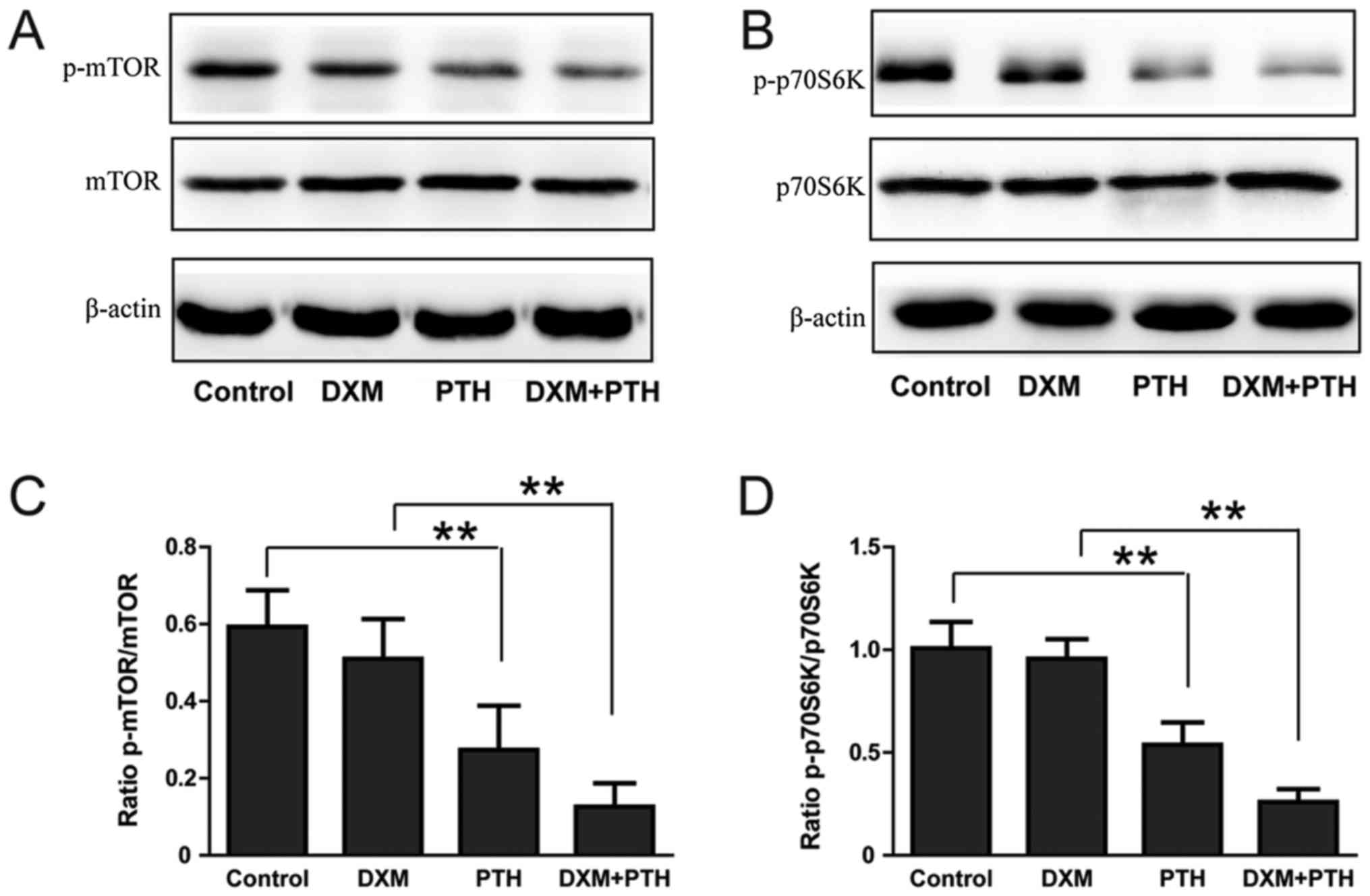
The effect of PTH upon the mTOR
signaling pathway in NPCs
mTOR-dependent and mTOR-independent signaling is the
main activation mechanism of autophagy, and p70S6K is a signaling
molecule downstream of mTOR. The expression of p-mTOR and p-p70S6K
in NPCs was significantly reduced after treatment with 10-nM PTH
for 48 h (P<0.05); a combination of DXM + PTH further reduced
the expression of p-mTOR and p-p70S6K compared with 25 µg/ml of DXM
alone (P<0.05; Fig. 4A-D),
indicating that PTH can inhibit the mTOR signaling pathway in NPCs
exposed to DXM.
Inhibition of autophagy reversed the
protective role of PTH on NPC senescence
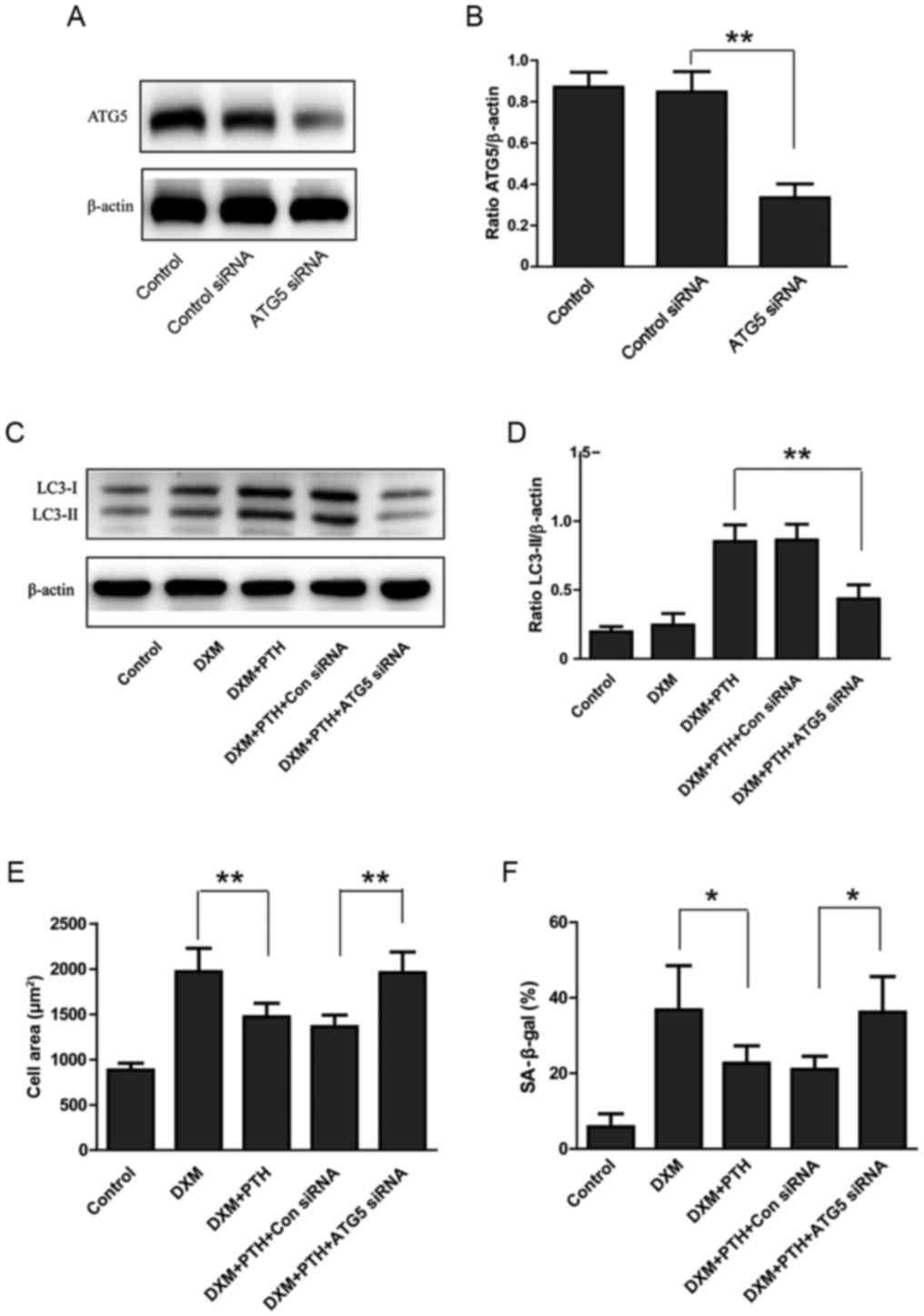
Next, we transfected NPCs with ATG5 siRNA to
suppress autophagy in an effort to analyze the potential
involvement of autophagy during the PTH-induced inhibition of
senescence in NPCs. Just 48 h after transfection, Western blotting
data showed a significant reduction of ATG5 levels (P<0.05;
Fig. 5A and B). Furthermore, ATG5
siRNA significantly inhibited LC3-II levels in NPCs treated with
PTH + DXM, demonstrating that ATG5 siRNA resulted in reduced levels
of autophagy (P<0.05; Fig. 5C and
D). Compared with DXM alone, a combination of DXM + PTH
significantly reduced the surface area of NPCs and reduced the
proportion of SA-β-gal-positive cells, indicating that PTH can
inhibit the senescence of NPCs induced by DXM (P<0.05; Fig. 5E and F). However, the transfection
of ATG5 siRNA reversed the inhibitory effect of PTH on the surface
area of NPCs and proportion of SA-β-gal-positive cells (P<0.05;
Fig. 5E and F). Therefore,
autophagy may be involved in the senescent effect of PTH on
NPCs.
Discussion
The treatment of disc degeneration with
glucocorticoids is still controversial. One previous study found
that injections of glucocorticoid could relieve short-term pain in
patients with acute disc pain, but was not effective over the
long-term (11). Although
glucocorticoids have the potential to treat IDD as inflammatory
reactions, they are also associated with a larger number of
complications. For example, Quan et al (14) found
that DXM can inhibit the nuclear translocation of the NF-κB
signaling pathway induced by TNF-α and then inhibit the
inflammatory response of NPCs in a short time period. However, the
prolonged use of DXM can stimulate the auto-phosphorylation of
NF-κB. The biological characteristics of chondrocyte is similar to
that of NPCs. The long-term use of DXM can promote apoptosis and
senescence in cartilage cells, prevent the growth of chondrocytes
and reduce the functional activity of chondrocytes (15–17).
However, the effect of DXM on senescence in NPCs has yet to be
reported.
In this study, we found that the cell size of NPCs
increased with increasing DXM concentration, and that 25 and 50
µg/ml DMX could significantly increase the proportion of cells that
were SA-β-gal-positive, thus indicating that DXM can promote
senescence in NPCs. The mechanism of senescence predominantly
involves to pathways: Replicative senescence and precocious
senescence. The mechanism underlying replicative senescence is
predominantly the p19Arf/p53/p21Cip1/Waf1 signaling pathway, while
the mechanism involved in premature senescence is mainly
p16INK4a/Rb (16,17); however, the mechanism by which DXM
causes senescence in NPCs still needs further study.
Autophagy maintains intracellular homeostasis by
clearing damaged organelles and aberrant proteins in cells by
fusion with lysosomes (18).
Autophagy is activated in NPCs when stimulated extracellularly by
acid, hypoxia, hypertonicity and starvation. This process leads to
the creation of an autophagosome which forms a bilayer membrane
structure in the cell. Proteins and organelles, together with
lysosomes, also form autolysosomes that degrade damaged organelles
and aberrant proteins and thus resist changes in the
microenvironment; however, the inhibition of autophagy can
accelerate apoptosis in NPCs (19,20).
In vivo studies of animals have found that the activation of
autophagy can inhibit senescence and apoptosis in NPCs and that IDD
can be partially alleviated (21).
In the present study, Western blot analysis of the expression of
LC3-II, Beclin-1 and P62, established markers of autophagy, and
GFP-LC3 assay showed that PTH could activate DXM-induced autophagy
in NPCs. Simialrly, Zhu et al (10) reported that 10 nM PTH could promote
bone cell autophagy and the inhibition of apoptosis in osteoblast.
In another study, Chen et al (11) reported that 0.1 ml/kg of 10 nM PTH
could relieve the progression of osteoarthritis in the knee joint
of rats by activating autophagy in chondrocytes. Although DXM has
been reported to increase the level of autophagy in chondrocytes
and osteocytes (15), the present
study indicated that this effect was not likely to be related to
the type of cell or the duration of action.
The key molecule underlying the activation of
autophagy is the mTOR protein and the mechanisms of involved is
usually divided into mTOR-dependent and mTOR independent pathways
(22). The activation of
mTOR-dependent autophagy involves the inhibition of mTOR expression
and a series of processes that cause autophagy in cells after the
inhibition of mTOR expression via the downstream mTOR signals
p70S6K and 4EBP1 (22). The
present study found that PTH inhibits the expression of p-mTOR and
p-p70S6K proteins, demonstrating that the mechanism by which PTH
activates autophagy may involve an mTOR-dependent signaling
pathway. In a previous study, Jiang et al (23) found that high osmotic pressure
activated autophagy by inhibiting the mTOR signaling pathway, thus
indicating that the mTOR signaling pathway in NPCs might represent
the main signaling pathway for the activation of autophagy.
The senescence of NPCs represents one of the
mechanisms that causes pathological changes in IDD. Cell senescence
blocks the cell cycle, reduces cell viability, increases the
expression of catabolic factors and extracellular matrix-degrading
enzymes, ultimately leading to changes involving the degradation of
extracellular matrix and disc structure (21). Autophagy is closely related to
senescence. In human bronchial epithelial cells, Sirt6 activates
autophagy via the IGF-Akt-mTOR signaling pathway to suppress cell
senescence (24). In NPCs,
silencing of mTORC1 by autophagy after mTORC1 activation protects
NPCs from inflammation-induced apoptosis, senescence and
degradation of the extracellular matrix (25). The present study found that PTH
could inhibit DXM-induced senescence in NPCs. However, in cells
transfected with ATG5 siRNA, this protective effect was reversed,
indicating that autophagy may mediate the inhibitory effect of PTH
on senescence in NPCs. Since ATG5 is an important protein in the
formation of autophagosomes, ATG5 siRNA is able toinhibit
acutophagic activation. The causes of senescence in NPCs often
include oxidative stress and inflammatory reactions. However,
previous studies have reported that autophagy can inhibit the
degree of oxidative stress and inflammation, so this may represent
the mechanism underlying the autophagic inhibition of senescence,
although this needs to be confirmed by further study.
Collectively our current data showed that DXM at 25
and 50 µg/ml promoted senescence in NPCs. PTH at a concentration of
10 mM increased the level of autophagy in cells treated by DXM and
inhibited the mTOR signaling pathway. PTH could therefore alleviate
senescence in NPCs under the action of DXM, while silencing ATG5 to
inhibit the level of autophagy could reverse the protective effect
of PTH. These results suggest that PTH may induce autophagy via the
mTOR signaling pathway to inhibit DXM-induced senescence in NPCs
and to protect NPCs.
Acknowledgements
Not applicable.
Funding
This work was supported by the project of Handan
Municipal Science and Technology Bureau (grant no.
1723208067-2).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XYW and LYJ designed and performed the experiments.
XYW, LYJ, JLH, ZAF and RJG analyzed the data. XYW, LYJ and JLH
prepared all the figures. XYW, JLH and RJG wrote the paper.
Ethics approval and consent to
participate
The present study was conducted in accordance with
the ethical guidelines of the 1975 Declaration of Helsinki and was
approved by the Committee of the Affiliated Hospital of Hebei
University of Engineering. Animal experiments were performed in
compliance with the National Institutes of Health Guide for the
Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rampersaud YR, Bidos A, Fanti C and
Perruccio AV: The need for multidimensional stratification of
chronic low back pain (LBP). Spine (Phila Pa 1976). 42:E1318–E1325.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hoy D, Bain C, Williams G, March L, Brooks
P, Blyth F, Woolf A, Vos T and Buchbinder R: A systematic review of
the global prevalence of low back pain. Arthritis Rheum.
64:2028–2037. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Neidlinger-Wilke C, Galbusera F, Pratsinis
H, Mavrogonatou E, Mietsch A, Kletsas D and Wilke HJ: Mechanical
loading of the intervertebral disc: From the macroscopic to the
cellular level. Eur Spine J. 23 Suppl 3:S333–S343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang L, Zhang X, Zheng X, Ru A, Ni X, Wu
Y, Tian N, Huang Y, Xue E, Wang X and Xu H: Apoptosis, senescence
and autophagy in rat nucleus pulposus cells: Implications for
diabetic intervertebral disc degeneration. J Orthop Res.
31:692–702. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gao C, Ning B, Sang C and Zhang Y:
Rapamycin prevents the intervertebral disc degeneration via
inhibiting differentiation and senescence of annulus fibrosus
cells. Aging (Albany NY). 10:131–143. 2018.PubMed/NCBI
|
|
6
|
Luo Y, Zhang L, Wang WY, Hu QF, Song HP,
Su YL and Zhang YZ: Alendronate retards the progression of lumbar
intervertebral disc degeneration in ovariectomized rats. Bone.
55:439–448. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jia H, Ma J, Lv J, Ma X, Xu W, Yang Y,
Tian A, Wang Y, Sun L, Xu L, et al: Oestrogen and parathyroid
hormone alleviate lumbar intervertebral disc degeneration in
ovariectomized rats and enhance Wnt/β-catenin pathway activity. Sci
Rep. 6:275212016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mizrahi J, Silva M, Keaveny T, Edwards W
and Hayes W: Finite-element stress analysis of the normal and
osteoporotic lumbar vertebral body. Spine (Phila Pa 1976).
18:2088–2096. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Madiraju P, Gawri R, Wang H, Antoniou J
and Mwale F: Mechanism of parathyroid hormone-mediated suppression
of calcification markers in human intervertebral disc cells. Eur
Cell Mater. 25:268–283. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu L, Chen J, Zhang J, Guo C, Fan W, Wang
YM and Yan Z: Parathyroid Hormone (PTH) induces autophagy to
protect osteocyte cell survival from dexamethasone damage. Med Sci
Monit. 23:4034–4040. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen CH, Ho ML, Chang LH, Kang L, Lin YS,
Lin SY, Wu SC and Chang JK: Parathyroid hormone (1–34) ameliorated
knee osteoarthritis in rats via autophagy. J appl physiol (1985).
124:1177–1185. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yin H, Wang S, Zhang Y, Wu M, Wang J and
Ma Y: Zuogui Pill improves the dexamethasone-induced osteoporosis
progression in zebrafish larvae. Biomed Pharmacother. 97:995–999.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He JL, Dong XH, Li ZH, Wang XY, Fu ZA and
Shen N: Pterostilbene inhibits reactive oxygen species production
and apoptosis in primary spinal cord neurons by activating
autophagy via the mechanistic target of rapamycin signaling
pathway. Mol Med Rep. 17:4406–4414. 2018.PubMed/NCBI
|
|
14
|
Quan M, Park SE, Lin Z, Hong MW, Park SY
and Kim YY: Steroid treatment can inhibit nuclear localization of
members of the NF-kB pathway in human disc cells stimulated with
TNF-α. Eur J Orthop Surg Traumatol. 25 Suppl 1:S43–S51. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zaman F, Chrysis D, Huntjens K, Chagin A,
Takigawa M, Fadeel B and Sävendahl L: Dexamethasone differentially
regulates Bcl-2 family proteins in human proliferative
chondrocytes: Role of pro-apoptotic Bid. Toxicol Lett. 224:196–200.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hong D, Chen HX, Yu HQ, Wang C, Deng HT,
Lian QQ and Ge RS: Quantitative proteomic analysis of
dexamethasone-induced effects on osteoblast differentiation,
proliferation and apoptosis in MC3T3-E1 cells using SILAC.
Osteoporos Int. 22:2175–2186. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xue E, Zhang Y, Song B, Xiao J and Shi Z:
Effect of autophagy induced by dexamethasone on senescence in
chondrocytes. Mol Med Rep. 14:3037–3044. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen K, Lv X, Li W, Yu F, Lin J, Ma J and
Xiao D: Autophagy is a protective response to the oxidative damage
to endplate chondrocytes in intervertebral disc: Implications for
the treatment of degenerative lumbar disc. Oxid Med Cell Longev.
2017:40417682017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang L, Jin Y, Wang H, Jiang Y and Dong
J: Glucosamine protects nucleus pulposus cells and induces
autophagy via the mTOR-dependent pathway. J Orthop Res.
32:1532–1542. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen J, Xie JJ, Jin MY, Gu YT, Wu CC, Guo
WJ, Yan YZ, Zhang ZJ, Wang JL, Zhang XL, et al: Sirt6
overexpression suppresses senescence and apoptosis of nucleus
pulposus cells by inducing autophagy in a model of intervertebral
disc degeneration. Cell Death Dis. 9:562018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alayev A and Holz MK: mTOR signaling for
biological control and cancer. J Cell Physiol. 228:1658–1664. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang LB, Cao L, Yin XF, Yasen M, Yishake
M, Dong J and Li XL: Activation of autophagy via Ca-dependent
AMPK/mTOR pathway in rat notochordal cells is a cellular adaptation
under hyperosmotic stress. Cell Cycle. 14:867–879. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
He J, Zhang G, Pang Q, Yu C, Xiong J, Zhu
J and Chen F: SIRT6 reduces macrophage foam cell formation by
inducing autophagy and cholesterol efflux under ox-LDL condition.
FEBS J. 284:1324–1337. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ito M, Yurube T, Kakutani K, Maeno K,
Takada T, Terashima Y, Kakiuchi Y, Takeoka Y, Miyazaki S, Kuroda R
and Nishida K: Selective interference of mTORC1/RAPTOR protects
against human disc cellular apoptosis, senescence and extracellular
matrix catabolism with Akt and autophagy induction. Osteoarthritis
Cartilage. 25:2134–2146. 2017. View Article : Google Scholar : PubMed/NCBI
|