Introduction
Obesity has become a worldwide public health
problem. It has previously been demonstrated that excessive energy
intake and obesity are considered to be risk factors for
cardiovascular diseases (1,2).
Calorie restriction (CR), an effective non-pharmacological
intervention of reduced energy intake, increases the health-span
and life-span of worms and yeast to rats and fish (3). Several studies have reported that CR
delays or decreases the development of various age-associated
diseases, including cardiovascular disease, diabetes, and various
forms of cancer (4–6). CR is important in regulating
cardiovascular diseases via activation of silent information
regulator 2 (SIR2) family members.
SIR2 is an NAD+_dependent deacetlyase and
is associated with life-span extension in CR. There are seven SIR2
orthologs in mammals, sirtuin (SIRT) 1–7. SIRT1 is the closest and
most well-characterized homolog of SIR2. SIRT1 mediates cellular
metabolism and exerts corresponding effects on gene expression via
deacetylating proteins, including the forkhead transcription
factors (FOXOs), myogenic differentiation 1, the p53 tumor
suppressor, and peroxisome proliferator-activated receptor-γ
co-activator 1α (7,8). Previous studies have demonstrated
that the SIR2 family exhibits an important regulatory role in the
pathogenesis of cardiovascular disease (9–12).
In the mouse 12.5 day embryo, SIRT1 is expressed only in myocardial
cells, however in adult rat myocardium, SIRT1 is expressed in the
nucleus and the cytoplasm (13).
SIRT1 deletion in rats leads to heart abnormalities, including
cardiac atrial septal defects, ventricular septal defects and
valvular deficiencies (14).
In addition to SIRT1, SIRT3, a stress-responsive
deacetylase, which is localized in mitochondria, exhibits a
positive effect in cardiac tissues. SIRT3 protects cardiomyocytes
in stress situations via deacetylating Ku70 (15). Studies reveal SIRT3 knockout rats
result in accelerated signs of aging in the heart including cardiac
hypertrophy (16).
It has been demonstrated that CR leads to
upregulated expression of SIRT in numerous organs, including rodent
brain, heart, liver and in white adipose tissues (17,18).
However, the extent to which CR affects cardiac function and SIRT
expression is largely unknown.
The present study investigated the impact of
moderate and severe calorie restriction compared with a high-fat
dietary regimen on morphology and function of the cardiovascular
system of aged rats. Furthermore, the effects of 25 and 45% CR
diets on myocardial SIRT1 and SIRT3 expression levels were
characterized.
Materials and methods
Ethics statement
The experiments were approved by the Institutional
Animal Care and Use Committee of Shantou University Medical College
(Shantou, China), and were conducted according to the Guide for the
Care and Use of Laboratory Animals (The Regulation of Experimental
Animals in Guangdong Province).
Materials
Primary antibodies against SIRT1, SIRT3 and β-actin
were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). SIRT1 rabbit polyclonal IgG, cat. no. sc-15404; SIRT3, rabbit
polyclonal IgG, cat. no. sc-49743; β-actin mouse monoclonal IgG,
cat. no. sc-47778. An enhanced chemiluminescence (ECL) western blot
detection system was obtained from GE Healthcare (Chicago, IL,
USA).
Animals and treatment
A total of 60 8-week-old female Sprague-Dawley rats,
weighing ~178.1 g, were purchased from the Animal Center, Shantou
University Medical College. The rats were housed separately in a
temperature- and humidity-controlled animal house at 22–25°C with a
12 h-12 h light-dark cycle. Following one week of acclimatization
feeding, rats were randomly divided into four groups (15 rats
each): a normal control group (NC) fed with standard rodent chow
containing 4% fat, 4.5% fiber, and 24% protein, ad libitum;
a 25% calorie restriction group (25% CR), fed with 75% of the total
caloric intake from the NC group; a 45% calorie restriction group
(45% CR), fed with 55% of the total caloric intake from the NC
group; a high-fat group (HF) fed with a high-fat diet containing
10% lard, 3% yolk, 1% cholesterol, 0.5% sodium cholate, and 85.5%
standard rodent chow. Sufficient vitamins and minerals were present
in each of the diets, and the rats had unlimited access to water
for 8 weeks. Rats were monitored and weighed daily.
Cardiac haemodynamic measurements
A total of eight weeks following treatment, left
ventricular (LV) performance was measured in rats anaesthetized
with intraperitoneal injections of 10% chloral hydrate (250 mg/kg).
After observation for 5 min, if the anaesthetic effect was not
satisfactory, another dose of 100 mg/kg was added. The rats were
placed on controlled heating pads, and the core temperature,
measured via a rectal probe, was maintained at 36–38°C. According
to a previously described method (19), a small cannula filled with heparin
saline (500 U/ml) was inserted into the left ventricle through the
apex with the chest open and mechanically ventilated, and
positioned along the cardiac longitudinal axis. Following
stabilization for 2 min, the pressure signal was continuously
recorded using a MacLab A/D converter (AD Instruments, Mountain
View, CA, USA). The left ventricular systolic and end-diastolic
pressures were measured, and the maximal slope of systolic pressure
increment (+dP/dt) and diastolic pressure decrement
(-dP/dt) were calculated. Following obtainment of the
haemodynamic measurements, the rats were sacrificed.
Sample collection and tissue
processing
Blood samples were collected from tail veins every
two weeks following fasting for 16 h. Sera were immediately
isolated from blood samples by centrifugation at 1,200 × g for 5
min at 4°C and stored at −20°C until analyzed. Hearts were
harvested and flushed with 0.9% normal saline solution, prior to
being sectioned into three parts. The central sections of each rat
heart, which included dual atriums and dual ventricles, were used
for western blot analysis and were immediately stored at −80°C.
Western blot analysis
Protein was extracted from heart tissues using
radioimmunoprecipitation assay buffer (1% Triton-X-100, 150 mmol/l
NaCl, 5 mmol/l EDTA and 10 mmol/l Tris-HCl, pH 7.0) containing a
protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). The tissue lysates were subjected to centrifugation at
13,200 g for 10 min at 4°C. Protein concentration was determined
using the Bradford method. A total of 50 µg protein per lane was
separated by 12% SDS-PAGE and transferred onto a polyvinylidene
fluoride membrane (EMD Millipore, Billerica, MA, USA). Following
three washes with PBS, the membranes were soaked in 5% non-fat dry
milk for 2 h at room temperature and incubated with primary
antibodies against β-actin (dilution, 1:1,000), SIRT1 (dilution,
1:200) and SIRT3 (dilution, 1:200) overnight at 4°C. Membranes were
then incubated with goat anti-rabbit (cat. no. sc-2004) and goat
anti-mouse (cat. no. sc-2005) horseradish peroxidase-conjugated IgG
antibodies (dilution, 1:2,000; Santa Cruz Biotechnology Inc.,
Dallas, TX, USA) for 1 h at room temperature. Immune complexes were
subsequently visualized using ECL and the band intensities were
measured and quantified using Quantity One® software
(Bio-Rad Laboratories Inc., Hercules, CA, USA).
Statistical analysis
All data are presented as the mean ± standard
deviation of at least three independent experiments. Statistical
analyses were performed using one-way analysis of variance followed
by Student-Newman-Keuls post hoc test for the comparison of
multiple groups. All data were analyzed using SPSS v. 19.0 (IBM
Corp., Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of CR on the body weight (BW),
heart weight (HW) and left ventricle mass index (LVMI) in rats
The 25% CR rats maintained their initial weight
until the end of the study, and their body weights were decreased
compared with NC and HF groups (P<0.05; Fig. 1A), however their HW and LVMI were
similar to the NC and HF groups (Fig.
1B and C). The 45% CR rats exhibited the lowest BW following
the experiment (P<0.01; Fig.
1A), and HW and LVMI were also the lowest (P<0.05; Fig. 1B and C). There was no significant
difference between HF and NC groups in BW and HW (Fig. 1A and B), however the LVMI of the HF
group was increased significantly compared with the other three
groups (P<0.05; Fig. 1C).
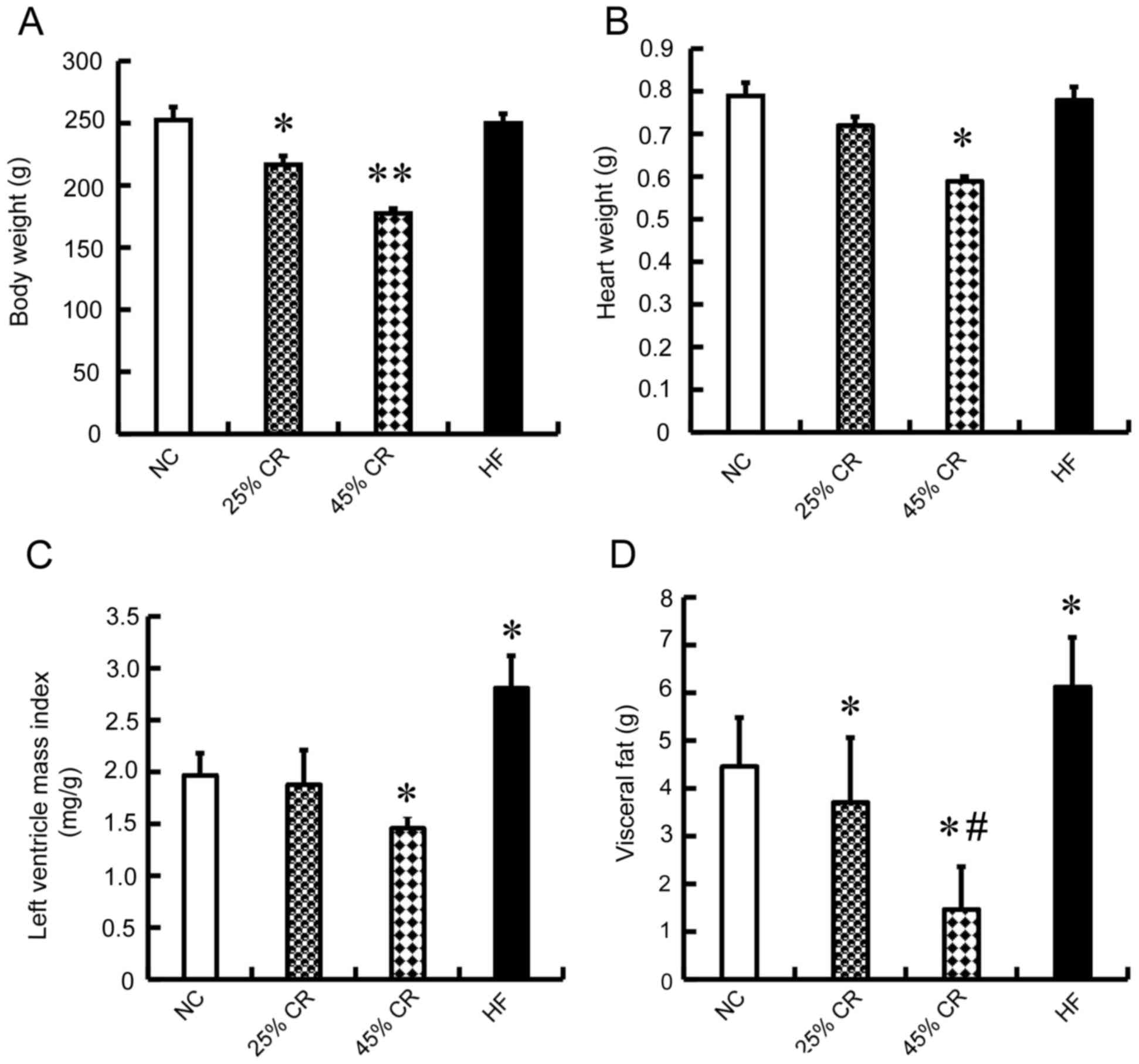 | Figure 1.Effect of CR on the BW, HW, LVMI and
VF in rats. The rats were randomized into four groups (15 mice
each) with similar BW, a NC group, 25, 45% CR and a HF group. (A)
BW, (B) HW, (C) LVMI and (D) VF were significantly decreased with
45% CR. There was no significant difference between HF and NC
groups in BW and HW, however the LVMI and VF weight of the HF group
was increased significantly compared with the other three groups.
Data are presented as the mean ± standard deviation. *P<0.05 vs.
the NC group; **P<0.01 vs. the NC group; #P<0.05
vs. the 25% CR group. BW, body weight; HW, heart weight; LVMI, left
ventricle mass index; VF, visceral fat; 25% CR, a 25% calorie
restriction group; 45% CR, 45% calorie restriction group; HF,
high-fat; NC, negative control. |
Effect of CR on the visceral fat (VF),
triglyceride (TG) and low density lipoprotein levels (LDL) in
rats
As presented in Fig.
1D, compared with the NC group, the two CR treated rats had
decreased VF weight (P<0.05), whereas the VF weight was
significantly increased in the HF group (P<0.05). The 45% CR
group had the lowest, and there was a significant difference
between the VF levels of the 25 and 45% CR groups (P<0.05). TG
levels in the HF-treated and NC-treated groups increased rapidly in
the first two weeks, decreased in the following weeks, however were
increased at all time points compared with the CR groups
(P<0.05). However, there was no significant difference between
HF and NC groups. The TG levels of the 25 and 45% CR rats decreased
(P<0.05), however there was no significant difference between
the CR groups (Table I). The LDL
levels the in HF-treated rats was significantly increased at all
time points (P<0.05). The LDL levels in other three groups
demonstrated similar alterations, slightly increased, however there
was no significant difference among the three groups (Table I).
 | Table I.Effect of CR on the TG and LDL levels
in rats. |
Table I.
Effect of CR on the TG and LDL levels
in rats.
| Variable | NC | 25% CR | 45% CR | HF |
|---|
| TG (mmol/l) |
|
|
|
|
| Week
2 | 0.78±0.11 | 0.82±0.08 | 0.88±0.05 | 0.89±0.09 |
| Week
4 | 1.09±0.15 |
0.89±0.21a,b |
0.84±0.13a,b |
1.32±0.06a |
| Week
6 | 0.81±0.23 |
0.57±0.18a,b |
0.57±0.31a,b |
0.96±0.19a |
| Week
8 | 0.86±0.17 |
0.67±0.13a,b |
0.55±0.13a,b |
1.05±0.19a |
| LDL (mmol/l) |
|
|
|
|
| Week
2 | 0.34±0.10 | 0.31±0.06 | 0.33±0.12 | 0.33±0.08 |
| Week
4 | 0.38±0.05 |
0.31±0.11b |
0.32±0.12b | 0.41±0.15 |
| Week
6 |
0.40±0.13b |
0.37±0.09b |
0.43±0.20b | 0.53±0.14 |
| Week
8 |
0.46±0.05b |
0.44±0.05b |
0.43±0.12b | 0.61±0.19 |
Effect of CR on the LV function in
rats
The heart rate is a good indicator of the cardiac
function. The present study measured the heart rate and it was
revealed that the 45% CR group rats' heart rate decreased, whereas
the 25% CR group increased (P<0.05; Fig. 2A). Haemodynamic measurements were
also recorded to determine the effect of CR on LV function in rats.
Compared with the NC group, the HF group rats exhibited a
significant deterioration in haemodynamic indices in LVSP
(96.4±6.46 vs. 112.8±7.08 mmHg, P<0.05; Fig. 2B), LVEDP (7.6±0.81 vs. 6.8±0.53
mmHg, P<0.05; Fig. 2C),
+dP/dt (5,212±453 vs. 6,030±486 mmHg/s, P<0.05;
Fig. 2D), and -dP/dt
(4,302±368 vs. 5,120±467 mmHg/s, P<0.05; Fig. 2E). However, rats treated with 25%
CR exhibited significant improvement in these haemodynamic indices
(P<0.05, Fig. 2B-E). Notably,
the 45% CR group had a significant deterioration in haemodynamic
indices compared with the 25% CR group in LVSP (81.7±9.56 vs.
131.5±7.14 mmHg, P<0.05; Fig.
2B), LVEDP (7.9±0.55 vs. 6.0±1.02 mmHg, P<0.05; Fig. 2C), +dP/dt (5,004±512
vs. 6,860±566 mmHg/s, P<0.05; Fig.
2D), and -dP/dt (4,198±499 vs. 5,978±412 mmHg/s,
P<0.05; Fig. 2E).
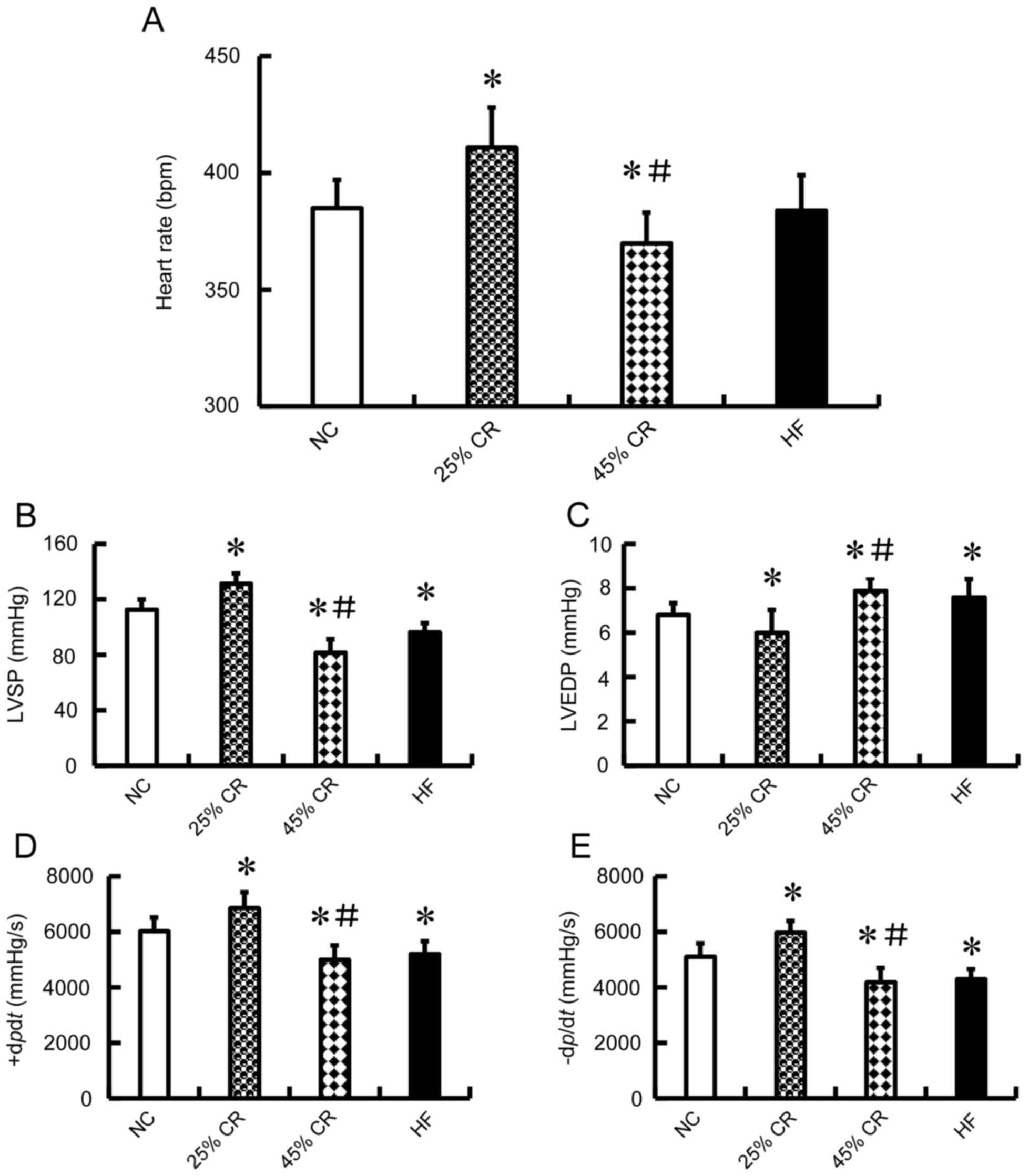 | Figure 2.Effect of CR on the LV function in
rats. The rats were randomized into four groups (15 mice each) with
similar BW, a NC group, 25, 45% CR and a HF group. A total of eight
weeks following treatment, (A) HR was measured and it was
demonstrated that the 45% CR group HR decreased, whereas the 25% CR
group HR increased. Furthermore, LV performance was measured in
rats anaesthetized by intraperitoneal injection of chloral hydrate.
The haemodynamic variables between different treatment groups are
presented. Alterations in (B) LVSP and (C) LVEDP. (D) The maximal
slope of systolic pressure increment (+dP/dt). (E)
The maximal slope of diastolic pressure decrement
(-dP/dt). Data are presented as the mean ± standard
deviation. *P<0.05 vs. the NC group; #P<0.05 vs.
the 25% CR group; n=8. HR, heart rate; LVSP, left ventricular
systolic pressure; LVEDP, left ventricular end-diastolic pressure;
25% CR, a 25% calorie restriction group; 45% CR, 45% calorie
restriction group; HF, high-fat; NC, negative control; LV, left
ventricular. |
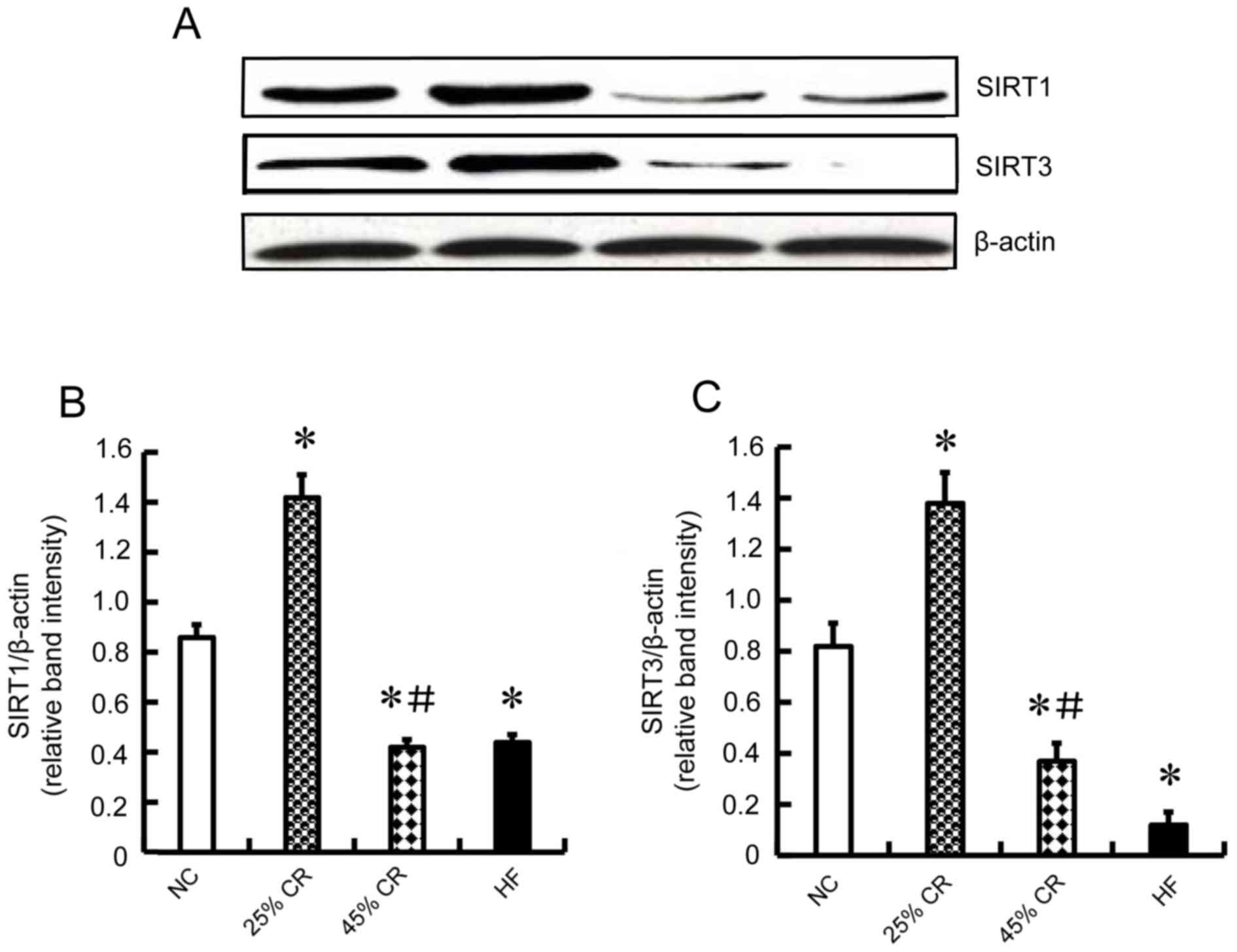
Western blot analysis of SIRT1 and
SIRT3 protein expression
To explore the molecular mechanism of CR on cardiac
function, immunoblotting was used to detect the protein expression
of SIRT1 and SIRT3. The expression of SIRT1 and SIRT3 in the 25% CR
group was increased compared with the other three groups
(P<0.05, Fig. 3A-C). The 45% CR
and HF groups had decreased levels (P<0.05, Fig. 3A-C) compared with NC group, and
there was no difference between 45% CR and HF groups.
Discussion
Obesity raises the risk of morbidity from
hypertension, dyslipidemia, type 2 diabetes mellitus (diabetes),
coronary heart disease, stroke, gallbladder disease,
osteoarthritis, sleep apnea, respiratory problems and various
cancers (20). Obesity is also
associated with increased risk in all-cause and cardiovascular
disease mortality (20). Although
CR as a nonpharmacological intervention of reduced energy intake
exhibits various benefits, including extending lifespan and
delaying age-associated diseases (3–5),
excessive CR may lead to a converse result.
In the present study, rats treated with a HF diet
had the heavier BW, increased LDL level, greater TG level, and more
abdominal fat. Conversely, rats treated with a 25% CR diet and a
45% CR diet were much healthier with decreased BW, lower LDL
levels, lower TG levels, and less abdominal fat. The 45% CR group
demonstrated the greatest decrease. However, the rats of the 45% CR
group exhibited various pathological features; their fur was
sparser compared with the other three groups. Their daily
activities were also not as energetic. The heart rate is a good
indicator of cardiac function. In the appropriate range, the faster
the heart rate, the better the cardiac function. At the end of the
study, the heart rates were measured and it was demonstrated that
the 45% CR group heart rates decreased, whereas the 25% CR group
heart rates increased. This indicated that excessive CR may be
harmful to health.
It is well known that excessive energy intake may
induce insulin resistance, central obesity, dyslipidemia and
hypertension, which are all important risk factors of
cardiovascular disease (1,2,21).
Dyslipidemia has been proven to have an important role in the
development of cardiovascular diseases (22,23).
SIRT1 and SIRT3 may regulate lipid metabolism. SIRT1 is a positive
regulator of liver X receptor proteins, nuclear receptors that
function as cholesterol sensors and regulate whole body cholesterol
and lipid homeostasis. SIRT1 knockout rats with a high-fat diet
lead to high TG and high LDL levels (24). SIRT3 may activate mitochondrial
fatty acid oxidation enzymes (12), suggesting SIRT3 may modulate lipid
metabolism. All these studies may explain why HF rats accumulated
fat mass and high blood fat whereas CR rats did not, although their
body weights have no significant difference.
SIRTs demonstrate diverse cellular localizations and
numerous cellular functions. The authors previously demonstrated
that SIRT1 was identified in the nucleus and cytoplasm, whereas
SIRT3 was predominantly observed in the cytoplasm (25). Moderate CR may upregulate SIRT
expression in various organs, including cardiac tissues. It has
previously been demonstrated that the mRNA and protein expression
of SIRT1-4 and −7 are significantly increased in the cardiac
tissues of rats in CR groups (25). However, it is still unclear whether
excessive CR has any effect on SIRT expression. The results of the
present study suggested that excessive CR resulted in a decline in
SIRT expression. Expression levels of SIRT1 and SIRT3 in 45% CR
were decreased compared with the 25% CR group, and were also
decreased compared with NC.
Previous studies demonstrate that high expression of
SIRT1 in transgenic rat hearts decreases cell apoptosis, cardiac
dysfunction and cardiac hypertrophy (26,27).
Increasing expression of SIRT3 protects cardiac tissues from
oxidative stress-mediated cell death (28). SIRT3-deficient mice at 8-weeks of
age have a basal level of cardiac hypertrophy, and a concomitant
decline in the protective effect of cardiac tissues (16). In the present study, the rats HW of
the HF group was not significantly different compared with the NC
and 25% CR groups; however, the LVMI of the HF rats was
significantly different compared with the other groups. The rats HW
and LVMI of the 45% CR group were the lowest. These results
demonstrated that the HF group rats exhibited left ventricular
hypertrophy, due to a decline of SIRT1 and SIRT3 expression. It was
additionally demonstrated that the haemodynamic indices LVSP,
+dP/dt and -dP/dt in the 45% CR group
decreased, and LVEDP increased via monitoring heart function. The
HF group demonstrated similar alterations, whereas the 25% CR group
demonstrated an adverse alteration. It may be hypothesized and
investigated in the future, that the decrease of SIRT1 and SIRT3
expression levels leads to a concomitant decline in the protective
effect of cardiac tissues.
Previous studies have demonstrated that lifelong
calorie restriction of 40% increases the cardiac expression of
autophagic markers, which suggests that it may have a
cardioprotective effect by decreasing oxidative damage brought on
by aging and cardiovascular diseases (29,30).
In response to glucose deprivation, cardiomyocytes initiate the
nuclear translocation of FoxO1 and FoxO3 to the nucleus where the
transcription of genes responsible for autophagy are activated
(31,32). Under a starvation state, SIRT1 is
upregulated (18,33). SIRT1 mediates the deacetylation of
FoxO1 and upregulation of Rab7, which functions as the center for
mediating increased autophagic flux in response to starvation,
which in turn maintains left ventricular function during these
events (31). Overall, these
findings unanimously support that calorie restriction may mediate
its beneficial effects by stimulating autophagy in the heart,
indicating the potential for cardioprotective therapy.
It is important to note that the present study had
several limitations. It is well known that age-associated
alterations primarily include changes in body weight, body fat,
blood glucose, insulin, triglyceride and cholesterol. The present
study only focused on indicators associated with cardiac function,
including body weight, heart weight, left ventricle mass index,
visceral fat, triglyceride and low density lipoprotein, however did
not include blood glucose and insulin. The authors aim to further
observe the alterations in blood glucose, insulin and other
indicators in future studies. In addition, the present study only
observed the effects of CR on SIRT1 and SIRT3 protein expression,
but not mRNA expression. In future studies, the authors will use
reverse transcription-fluorescence quantitative polymerase chain
reaction to observe the mRNA expression of SIRT1 and SIRT3.
In conclusion, the data demonstrated that moderate
calorie restriction (25% CR), rather than severe calorie
restriction (45% CR), attenuated age-associated alterations and
improved myocardial function in rats. The molecular mechanisms
responsible for the protective effect of moderate calorie
restriction may involve the expression of SIRT1 and SIRT3. Moderate
calorie restriction may upregulate the expression levels of SIRT1
and SIRT3, however severe calorie restriction may lead to a
decline. Therefore, the findings of the present study indicated
that moderate calorie restriction may be an effective therapeutic
approach for obesity-associated diseases.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81270382/H0215).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WY and WW conceived and designed the study. WY and
JJQ performed the experiments. WY, JJQ, CJC, YCF and WW collected
and analyzed the data. WY and WW wrote the manuscript. All authors
reviewed and revised the manuscript.
Ethics approval and consent to
participate
The experiments were approved by the Institutional
Animal Care and Use Committee of Shantou University Medical College
(Shantou, China), and were conducted according to the Guide for the
Care and Use of Laboratory Animals (The Regulation of Experimental
Animals in Guangdong Province).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ginsberg HN and MacCallum PR: The obesity,
metabolic syndrome, and type 2 diabetes mellitus pandemic: Part I.
Increased cardiovascular disease risk and the importance of
atherogenic dyslipidemia in persons with the metabolic syndrome and
type 2 diabetes mellitus. J Cardiometab Syndr. 4:113–119. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lavie CJ, Milani RV and Ventura HO:
Obesity and cardiovascular disease: Risk factor, paradox, and
impact of weight loss. J Am Coll Cardiol. 53:1925–1932. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Civitarese AE, Carling S, Heilbronn LK,
Hulver MH, Ukropcova B, Deutsch WA, Smith SR and Ravussin E;
CALERIE Pennington Team, : Calorie restriction increases muscle
mitochondrial biogenesis in healthy humans. PLoS Med. 4:e762007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Varady KA and Hellerstein MK: Do calorie
restriction or alternate-day fasting regimens modulate adipose
tissue physiology in a way that reduces chronic disease risk? Nutr
Rev. 66:1–342. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lefevre M, Redman LM, Heilbronn LK, Smith
JV, Martin CK, Rood JC, Greenway FL, Williamson DA, Smith SR and
Ravussin E; Pennington CALERIE team, : Caloric restriction alone
and with exercise improves CVD risk in healthy non-obese
individuals. Atherosclerosis. 203:206–213. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Holloszy JO and Fontana L: Caloric
restriction in humans. Exp Gerontol. 42:709–712. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taylor DM, Maxwell MM, Luthi-Carter R and
Kazantsev AG: Biological and potential therapeutic roles of sirtuin
deacetylases. Cell Mol Life Sci. 65:4000–4018. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Al-Regaiey KA, Masternak MM, Bonkowski M,
Sun L and Bartke A: Long-lived growth hormone receptor knockout
mice: Interaction of reduced insulin-like growth factor i/insulin
signaling and caloric restriction. Endocrinology. 146:851–860.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fontana L, Klein S, Holloszy JO and
Premachandra BN: Effect of long-term calorie restriction with
adequate protein and micronutrients on thyroid hormones. J Clin
Endocrinol Metab. 91:3232–3235. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Potente M, Ghaeni L, Baldessari D,
Mostoslavsky R, Rossig L, Dequiedt F, Haendeler J, Mione M, Dejana
E, Alt FW, et al: SIRT1 controls endothelial angiogenic functions
during vascular growth. Genes Dev. 21:2644–2658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hsu CP, Odewale I, Alcendor RR and
Sadoshima J: Sirt1 protects the heart from aging and stress. Biol
Chem. 389:221–231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sundaresan NR, Gupta M, Kim G, Rajamohan
SB, Isbatan A and Gupta MP: Sirt3 blocks the cardiac hypertrophic
response by augmenting Foxo3a-dependent antioxidant defense
mechanisms in mice. J Clin Invest. 119:2758–2771. 2009.PubMed/NCBI
|
|
13
|
Tanno M, Sakamoto J, Miura T, Shimamoto K
and Horio Y: Nucleocytoplasmic shuttling of the NAD+-dependent
histone deacetylase SIRT1. J Biol Chem. 282:6823–6832. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sakamoto J, Miura T, Shimamoto K and Horio
Y: Predominant expression of Sir2alpha, an NAD-dependent histone
deacetylase, in the embryonic mouse heart and brain. FEBS Lett.
556:281–286. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sundaresan NR, Samant SA, Pillai VB,
Rajamohan SB and Gupta MP: SIRT3 is a stress-responsive deacetylase
in cardiomyocytes that protects cells from stress-mediated cell
death by deacetylation of Ku70. Mol Cell Biol. 28:6384–6401. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hafner AV, Dai J, Gomes AP, Xiao CY,
Palmeira CM, Rosenzweig A and Sinclair DA: Regulation of the mPTP
by SIRT3-mediated deacetylation of CypD at lysine 166 suppresses
age-related cardiac hypertrophy. Aging (Albany NY). 2:914–923.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wolf G: Calorie restriction increases life
span: A molecular mechanism. Nutr Rev. 64:89–92. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cohen HY, Miller C, Bitterman KJ, Wall NR,
Hekking B, Kessler B, Howitz KT, Gorospe M, de Cabo R and Sinclair
DA: Calorie restriction promotes mammalian cell survival by
inducing the SIRT1 deacetylase. Science. 305:390–392. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu L, Zhang X, Qian B, Min X, Gao X, Li
C, Cheng Y and Huang J: Over-expression of heat shock protein 27
attenuates doxorubicin-induced cardiac dysfunction in mice. Eur J
Heart Fail. 9:762–769. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jensen MD, Ryan DH, Apovian CM, Ard JD,
Comuzzie AG, Donato KA, Hu FB, Hubbard VS, Jakicic JM, Kushner RF,
et al: 2013 AHA/ACC/TOS guideline for the management of overweight
and obesity in adults: A report of the American College of
Cardiology/American Heart Association Task Force on Practice
Guidelines and The Obesity Society. Circulation. 129 25 Suppl
2:S102–S138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee M and Aronne LJ: Weight management for
type 2 diabetes mellitus: Global cardiovascular risk reduction. Am
J Cardiol. 99:68B–79B. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bernardi R, Cosentino ER and Borghi C:
Metabolic syndrome and hypertension: Prevention and treatment.
Minerva Med. 97:123–141. 2006.(In Italian). PubMed/NCBI
|
|
23
|
Ambroskina VV, Kriachok TA, Larionov OP,
Talaieva TV and Bratus' VV: Hyperlipidemia and decrease of lipid
tolerance as factors of atherogenesis. Fiziol Zh. 53:19–28.
2007.PubMed/NCBI
|
|
24
|
Li X, Zhang S, Blander G, Tse JG, Krieger
M and Guarente L: SIRT1 deacetylates and positively regulates the
nuclear receptor LXR. Mol Cell. 28:91–106. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu W, Zhou HF, Lin RB, Fu YC and Wang W:
Short-term calorie restriction activates SIRT1-4 and −7 in
cardiomyocytes in vivo and in vitro. Mol Med Rep. 9:1218–1224.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Alcendor RR, Gao S, Zhai P, Zablocki D,
Holle E, Yu X, Tian B, Wagner T, Vatner SF and Sadoshima: Sirt1
regulates aging and resistance to oxidative stress in the heart.
Circ Res. 100:1512–1521. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu W, Fu YC, Zhou XH, Chen CJ, Wang X, Lin
RB and Wang W: Effects of resveratrol on H(2)O(2)-induced apoptosis
and expression of SIRTs in H9c2 cells. J Cell Biochem. 107:741–747.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen CJ, Fu YC, Yu W and Wang W: SIRT3
protects cardiomyocytes from oxidative stress-mediated cell death
by activating NF-κB. Biochem Biophys Res Commun. 11:798–803. 2013.
View Article : Google Scholar
|
|
29
|
Ahn J and Kim J: Nutritional status and
cardiac autophagy. Diabetes Metab J. 37:30–35. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wohlgemuth SE, Julian D, Akin DE, Fried J,
Toscano K, Leeuwenburgh C and Dunn WA Jr: Autophagy in the heart
and liver during normal aging and calorie restriction. Rejuvenation
Res. 10:281–292. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hariharan N, Maejima Y, Nakae J, Paik J,
Depinho RA and Sadoshima J: Deacetylation of FoxO by Sirt1 plays an
essential role in mediating starvation-induced autophagy in cardiac
myocytes. Circ Res. 107:1470–1482. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sengupta A, Molkentin JD and Yutzey KE:
FoxO transcription factors promote autophagy in cardiomyocytes. J
Biol Chem. 284:28319–28331. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nemoto S, Fergusson MM and Finkel T:
Nutrient availability regulates SIRT1 through a forkhead-dependent
pathway. Science. 306:2105–2108. 2004. View Article : Google Scholar : PubMed/NCBI
|