Introduction
Microglia, a type of glial cell, is an important
immune line of defence in the central nervous system. The effects
of microglia in brain tissue and spinal cord are similar to those
of macrophages (1). Of all the
glial cells found in brain tissue, 20% are microglia (2). Microglia hyperproliferative
activation is found in Alzheimer's disease, Parkinson's disease,
and other diseases, and the overactivation of microglia has been
shown to lead to neurotoxicity (3).
Chemokine CC ligand 2 (CCL2) is a member of the
chemokine family, which is secreted by human vascular endothelial
cells, astrocytes, microglia, smooth muscle cells, and other types
of cell. CCL2 is able to regulate macrophage chemotaxis, the
activation of basophils, as well as being involved in the processes
of cell proliferation and apoptosis (4,5).
α-synuclein is a type of nerve protein that is widely distributed
in brain tissue, where it serves an important role in maintaining
synaptic function (6). The
over-aggregation of α-synuclein leads to Lewy body dysfunction,
triggering learning, memory and other functional deficiencies
(7). A previous study has
demonstrated that CCL2 may promote the proliferation of microglia,
and also promote the secretion of inflammatory factors, thus
leading to the toxicity produced by excessive aggregation of
α-synuclein (8). In the present
study, microglia were used as the cell type of choice to
investigate the role of CCL2 in α-synuclein-mediated microglia
proliferation and neuronal apoptosis, in order to provide a
theoretical basis underpinning the development of therapies for
treatment of neurological diseases.
Materials and methods
Materials
A total of 10 SPF C57/BL6 mice (5 female/5 male),
aged 1–3 days and 200±20 g in weight, were purchased from the
Experimental Animal Center of Xinxiang Medical University
(Xinxiang, China). All animals were housed in sterile conditions at
19–21°C, with a relative humidity of 50–60%, ad libitum
intake of water and food and under 12-h of light/dark cycle. The
present study was performed in strict accordance with the
recommendations given in the Guide for the Care and Use of
Laboratory Animals of the National Institutes of Health. The animal
use protocol was reviewed and approved by the Institutional Animal
Care and Use Committee (IACUC) of Xinxiang Medical University.
Main instruments and reagents
DMEM/F12 medium and trypsin were purchased from
Sigma-Aldrich; Merck KGaA, (Darmstadt, Germany). Fetal bovine serum
(FBS; Gibco™) was purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Penicillin-streptomycin
solution was purchased from Beijing Dingguo Biotechnology Co., Ltd.
(Beijing, China). A HERAcell 150i CO2 incubator and an
MK3 microplate reader were purchased from Thermo Fischer
Scientific, Inc. Monoclonal antibodies raised against α-synuclein,
cleaved caspase-3, Akt and phosphorylated (p)-Akt were purchased
from R&D Systems, Inc. (Minneapolis, MN, USA). A bicinchoninic
acid (BCA) protein quantification kit was purchased from Shanghai
Yi Sheng Biotechnology Co., Ltd. (Shanghai, China). Tumor necrosis
factor-α (TNF-α) and interleukin-1β (IL-1β) detection kits were
purchased from Shanghai Bai Rui Biotechnology Co., Ltd. (Shanghai,
China). A nitric oxide (NO) content detection kit was purchased
from Applygen Technologies, Inc. (Beijing, China).
Primary isolation and culture of
microglia and primary neurons
The mice, within 24 h of birth, were sacrificed
prior to subsequent investigations, and the numbers of mice that
were sacrificed were dependent on the requirements for each
individual set of experiments. The cerebral cortex vessels were
removed and placed in DMEM/F12 medium, after having been treated
with 75% alcohol. Following excision of the brain tissues, 10 ml
0.125% trypsin was added, and the samples were placed in a water
bath at37°C to perform a rapid digestion reaction for 5 min. FBS
was then added to terminate the digestion reaction, and the samples
were placed at room temperature for 3 min; tissue precipitation was
observed in the lower layer, and the supernatant was transferred to
EP tubes by filtering through a 70 µm filter. The filtered solution
was centrifuged at 168 × g for 10 min at 25°C, and the supernatant
was then discarded. DMEM/F12 medium containing 15% FBS and 100 U/ml
penicillin-streptomycin was added to the pellet and the cells
suspension was transferred into a cell culture flask coated with
poly-d-lysine for incubation in a 5% CO2 incubator at
37°C for 24 h following by replacement of fresh cell culture. After
the cells in the flask were cultured for 10 days and incubated with
agitation for 60 min, the medium was removed and the cells were
washed with PBS. The cells were then collected and seeded into a
6-well cell culture plate. A total of 3×105 cells were
added to each well. Following incubation for 30 min, the adherent
cells were microglia cells; following 5 days of culture, it was
possible to collect the primary microglia.
Following sieving, the filtrate was centrifuged at
300 × g at 4°C for 10 min, and the precipitated cells were
suspended in DMEM/F12 cell culture medium containing 10% FBS, 2%
B27 and 100 U/ml penicillin-streptomycin solution. Cells were
subsequently inoculated into a lysine-coated cell flask, and
following 24 h, the medium was replaced by the same medium lacking
FBS. Primary neurons were obtained three days after isolation and
culture.
Experimental grouping
The cells were divided into a control group, a CCL2
group, an α-synuclein group and a CCL2 plus α-synuclein group. In
the control group, the cells were treated with an identical amount
of PBS, whereas the cells in the CCL2 group were cultured in medium
containing CCL2 (1.632 mg/ml) at a concentration of 0.05 ng/µl;
cells in the α-synuclein group were treated with medium containing
α-synuclein (1.120 mg/ml) at a concentration of 0.2 ng/µl; and
cells in the CCL2 plus α-synuclein group were cultured in medium
containing 0.05 ng/µl CCL2 and 0.2 ng/µl α-synuclein.
MTT detection of cell
proliferation
Washed cells were seeded into 96-well cell culture
plates, and 100 µl cell suspension was added to each well. The
cells were incubated at 37°C in a 5% CO2 incubator for
24 h. The medium was replaced as described in the Primary isolation
and culture of microglia and neurons section above, 20 µl MTT
solution (5 mg/ml) was added to each well, and the cells were
further incubated in the CO2 incubator for 4 h, after
which time 150 µl dimethyl sulfoxide solution was added. After
allowing the reaction to proceed for 10 min, the absorbance values
per well were measured at 450 nm using a microplate reader.
Detection of NO by the Griess
method
NO gives rise to nitrite (NO2−) anions in
an aqueous environment, although in an acidic environment,
NO2− may react with sulfonamide compounds to form
diazonium salts, as well as coupling with naphthylethylenediamine
(Griess reagent). The more that these compounds are generated, the
higher is the absorbance measured at 540 nm, reflecting the higher
NO content. Cells in each group were collected, and the cells were
cultured for 24 h. The NO content in the cell culture medium was
detected by following the protocol provided with the NO content
detection kit.
Determination of the TNF-α and IL-1β
content by the ELISA method
Cells in each group were collected and cultured in
the incubator at 37°C for 24 h. The levels of inflammatory factors
in the cell culture medium were measured using an ELISA kit of
TNF-α (cat. no. T04140; Groundwork Biotechnology Diagnosticate,
Ltd., San Diego, USA) or IL-1β (cat. no. V006-10; Groundwork
Biotechnology Diagnosticate, Ltd., San Diego, CA, USA). The
absorbance of each well at 450 nm was measured using a microplate
reader.
α-Synuclein levels in cells detected
by western blot analysis
Cells in each group were collected and cultured
after 24 h. The medium was discarded. The cells were incubated with
lysate for 10 min, and the lysate was then transferred into
centrifuge tubes and centrifuged at 168 × g for 10 min at 4°C. The
supernatant was transferred into the EP tube, and the extracted
protein concentration was detected using a BCA protein
quantification kit. The protein sample was subsequently mixed with
loading buffer and boiled at 100°C for 5 min. The denatured protein
samples were added to the sample wells of an SDS-PAGE gel (10%),
and the initial voltage for electrophoresis was set at 90 V. When
the bromophenol blue was observed to have run into the edge of the
stacking and removing gels, the voltage was adjusted to 120 V until
the end of the electrophoresis was reached. The proteins (20
µg/well) loaded in the gel were electro-transferred onto a
polyvinylidene difluoride (PVDF) membrane for 90 min at 4°C. The
PVDF membrane was sealed in 5% skimmed milk powder at 37°C for 90
min, followed by reaction with the primary
antibody-anti-α-synuclein antibody (cat. no. ab138501; 1:500; 4°C
incubation overnight; Abcam, Cambridge, UK), followed by the
secondary antibody-goat anti-rabbit IgG H&L (HRP) (37°C
incubation for 90 min; cat. no. ab6721; 1:1,000; Abcam). After the
reaction was allowed to develop with the colour substrate using
Visualizer™ Western Blot Detection kit (EMD Millipore,
Billerica, MA, USA), the protein expression levels were analyzed,
comparing against GAPDH as the internal reference protein.
Detection of apoptosis by flow
cytometry
Cells in each group were cultured for 24 h according
to the identical procedure as that described for culture of the
microglial cells. Cell culture medium was added to the neurons for
24 h, following which time the culture medium was discarded, the
cells were treated with 0.125% trypsin, and following digestion,
the mixture was centrifuged at 300 × g at 4°C for 5 min. The
supernatant was then discarded, the cells were washed once in PBS,
binding buffer was added, and the cells were suspended and mixed,
adjusting the concentration of the cells to 3×106
cells/ml. A total of 5 µl of Annexin V and propidium iodide (PI)
were added to each 100 µl cell suspension, and the reactions were
allowed to proceed at room temperature for 10 min. Apoptosis was
detected using flow cytometry FACSARIA III (BD) with FlowJo 10
(LLC, Ashland, OR, USA) software using a Dead Cell Apoptosis kit
(cat. no. V13242; Invitrogen; Thermo Fisher Scientific, Inc.).
Detection of the levels of cleaved
caspase-3, Akt and p-Akt in neurons by western blotting
Again, cells in each group were cultured for 24 h
according to the identical procedure as that described for culture
of the microglial cells. Cell culture medium was added to the
neurons for 24 h, after which time the culture medium was discarded
and the proteins in the neurons were collected. The levels of
cleaved caspase-3, Akt and p-Akt in the neurons was detected by
western blotting, as described above.
Statistical analysis
Experimental data were analysed using IBM SPSS
statistics software, version 22 (IBM Corps, Armonk, NY, USA). Data
were expressed as the mean ± standard deviation (x±s).
Comparisons between two of the groups were made using the t-test.
One-way analysis of variance was used to compare multiple groups.
Multiple comparisons between the groups were performed using
Tukey's post-hoc test. P<0.05 and P<0.01 were considered to
indicate a statistically significant difference.
Results
Cell proliferation
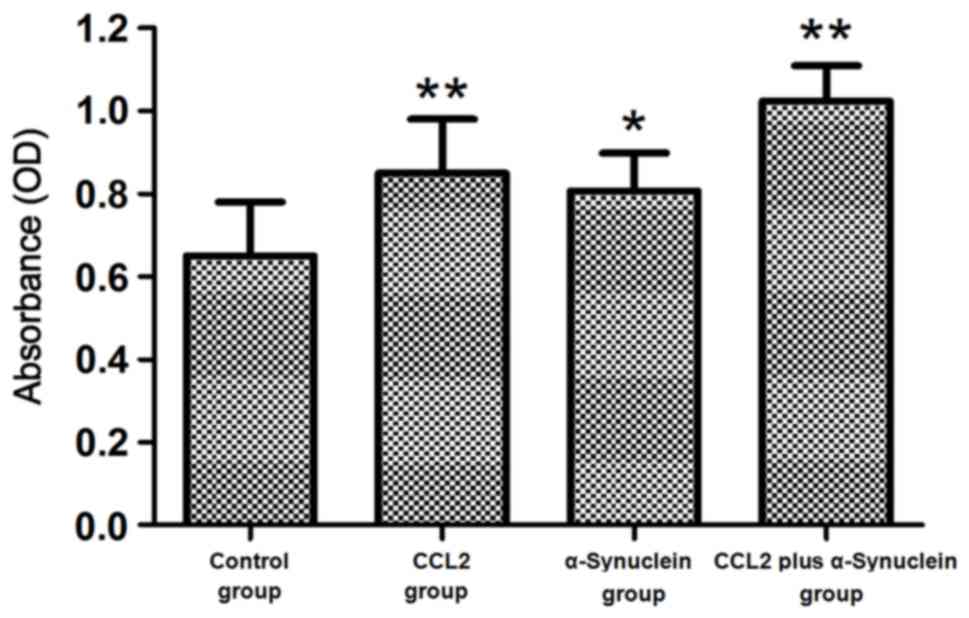
The proliferation of cultured microglia cells was
detected by MTT assay after the cells were treated with 0.05 ng/µl
CCL2, 0.2 ng/µl α-synuclein and 0.05 ng/µl CCL2 plus 0.2 ng/µl
α-synuclein. The results demonstrated that proliferation of the
α-synuclein cells was significantly higher compared with that of
the control group (P<0.05). Furthermore, proliferation of the
cells in the CCL2 and CCL2 plus α-synuclein groups was
significantly higher compared with that in the control group
(P<0.01; Fig. 1).
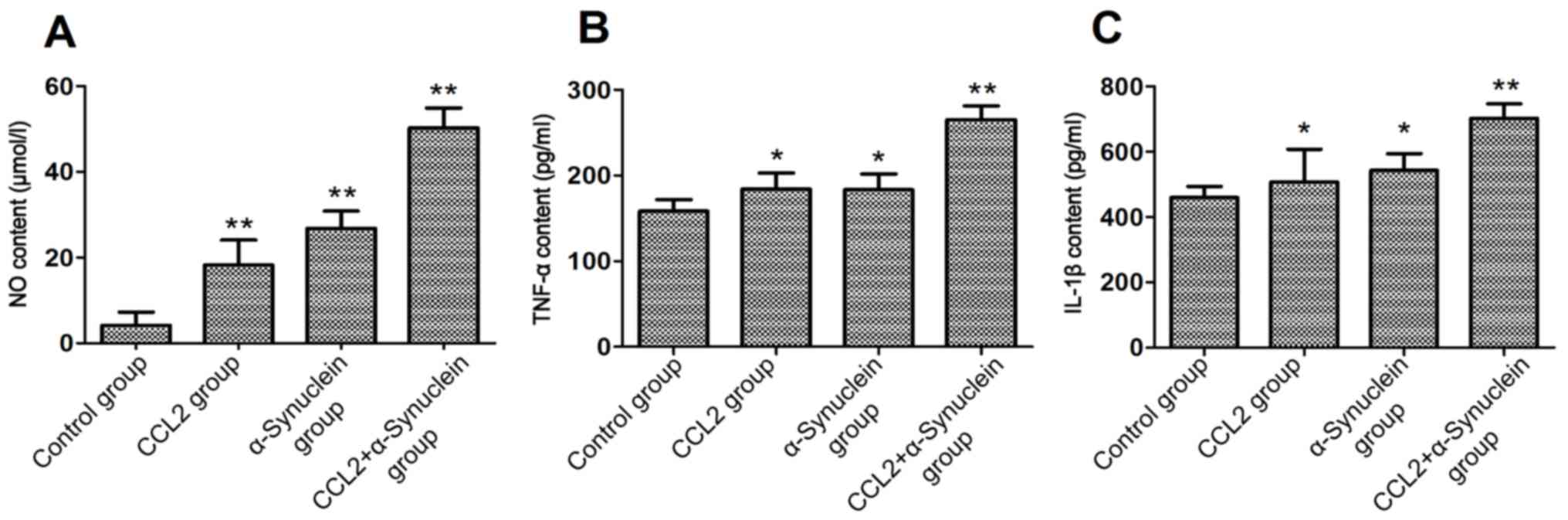
Detection of the levels of NO, TNF-α
and IL-1β
After 24 h culture, the medium for each experimental
group was collected, and the levels of NO, TNF-α and IL-1β in the
supernatant were subsequently measured. The results showed that the
levels of NO in the CCL2, α-synuclein and CCL2 plus α-synuclein
groups were significantly higher compared with that in the control
group (P<0.01). In addition, the levels of TNF-α and IL-1β in
the CCL2 and α-synuclein groups were significantly higher compared
with that in the control group (P<0.05). The levels of TNF-α and
IL-1β in the CCL2 plus α-synuclein group were significantly higher
compared with that in the control group (P<0.01; Fig. 2).
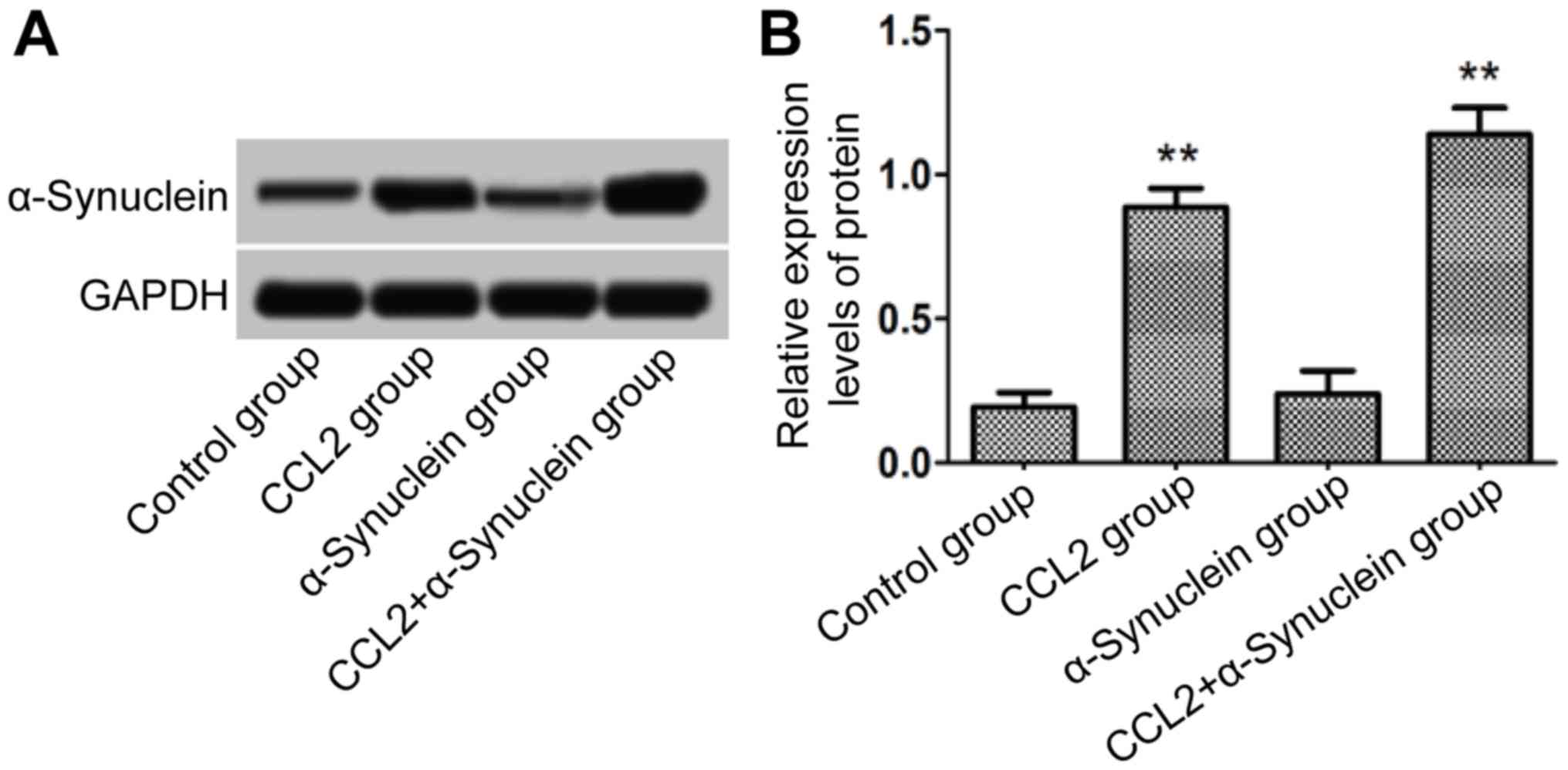
α-Synuclein level in cells
The cells in each group following 24 h culture were
collected, and total protein was then extracted. The levels of
α-synuclein were detected using western blot analysis. The results
demonstrated that, compared with the control group, there was no
obvious increase in the expression level of α-synuclein for the
α-synuclein group. However, the levels of α-synuclein in the CCL2
and CCL2 plus α-synuclein groups were significantly higher compared
with that in the control group (P<0.01; Fig. 3).
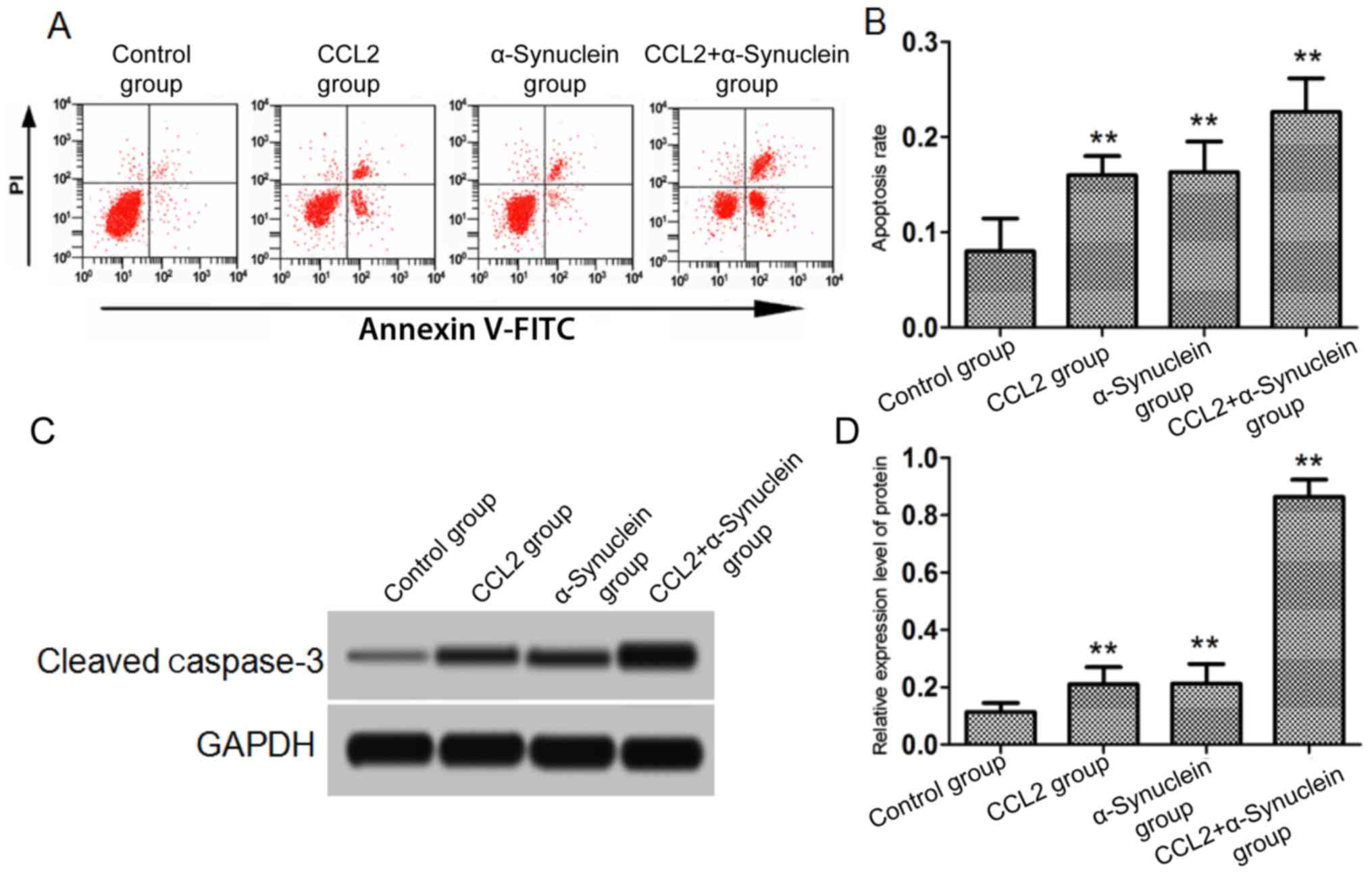
Neuronal apoptosis
After the cells in each group were cultured for 24
h, microglia cells were added to the neurons. Cells undergoing
apoptosis were subsequently detected using flow cytometric
analysis, and the level of cleaved caspase-3 in the neurons after
24 h was detected by western blotting. The results showed that the
apoptotic rate of the CCL2, α-synuclein and CCL2 plus α-synuclein
groups was significantly higher compared with that of the control
group (P<0.01). Similarly, the levels of cleaved caspase-3
protein in the CCL2, α-synuclein and CCL2 plus α-synuclein groups
were significantly higher compared with that in control group
(P<0.01; Fig. 4).
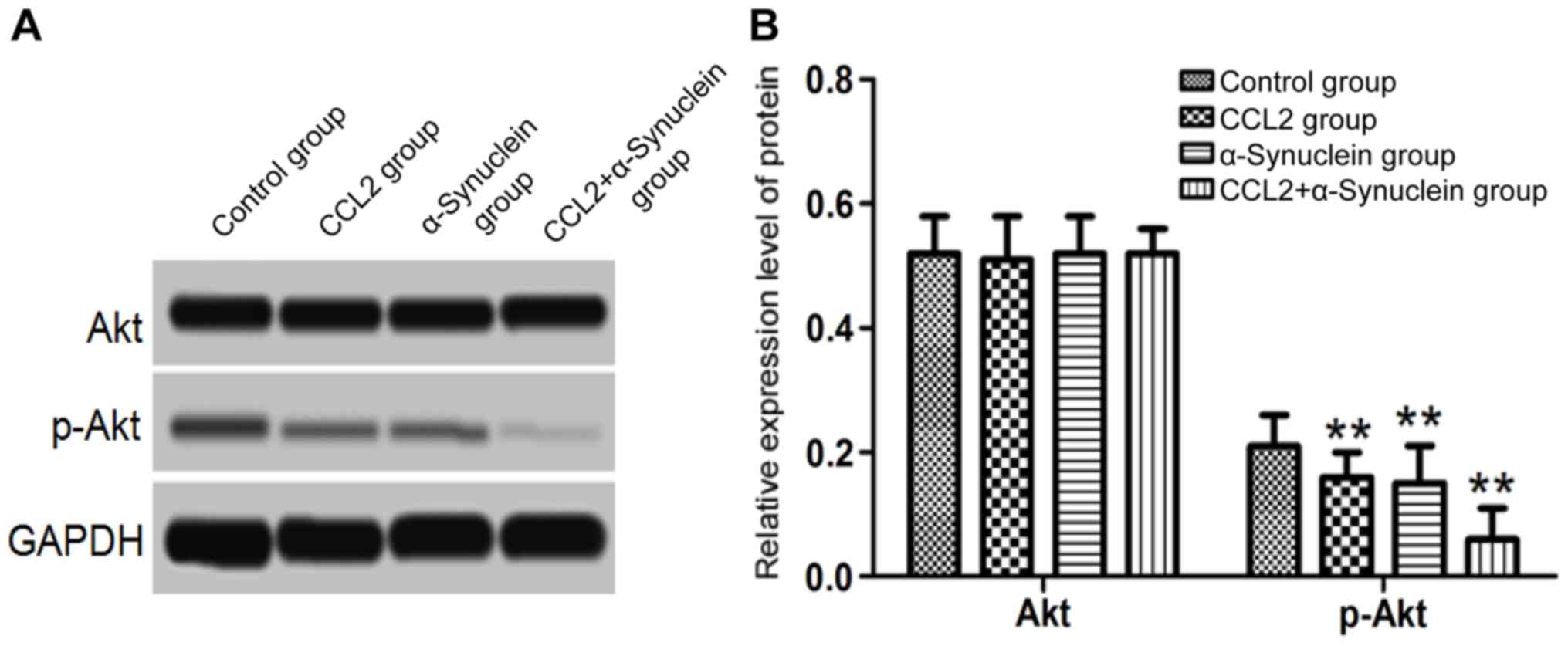
Levels of Akt and p-Akt in the cells
determined by western blot analysis
Following culture for 24 h, the microglia cell
culture medium was added to the neurons. The levels of Akt and
p-Akt were then detected by western blotting. The results
demonstrated that no significant differences were identified in the
levels of Akt protein in the CCK2, α-synuclein and CCL2 plus
α-synuclein groups compared with the control group (P>0.05).
However, the levels of p-Akt protein in the CCL2, α-synuclein and
CCL2 plus α-synuclein groups were significantly lower compared with
those in the control group (P<0.01; Fig. 5).
Discussion
Microglia originate in the neuroectoderm. When the
central nervous system becomes damaged, microglia are able to
engulf harmful substances in the nervous system (9). In mature brain tissue, microglia are
in a quiescent state, and are able to sense abnormal pathological
changes in the surrounding environment. Microglia are also able to
promote neuronal proliferation, and to protect the brain's tissue
structure. Microglia activation may lead to the secretion of a
variety of immune factors and NO (10). On the one hand, NO has a role in
the killing of pathogens; on the other hand, neurons themselves may
exert a certain toxic effect. A previous study has demonstrated
that neurodegenerative diseases are associated with abnormal
activation of the microglia, leading to the production of
inflammatory factors, which, in turn, have a certain toxicity for
the nervous system (11).
The gene for α-synuclein is located on chromosome 4
and contains 7 exons (12).
α-Synuclein protein is widely expressed in various structures of
the brain, including the cerebral cortex, neuronal synapse,
hippocampus, and so forth, and these are involved in the processes
of normal synaptic signal transmission. Under normal circumstances,
microglia may engulf and degrade α-synuclein protein (13). However, when the microglia are
abnormally activated, their ability to engulf and degrade
α-synuclein protein decreases, thus leading to a large accumulation
of α-synuclein proteins that produce neurotoxicity (14). A previous study has shown that the
overexpression of α-synuclein in the bodies of animals leads to
abnormal activation of the microglia, thereby leading to an
increased rate of neuronal death (15). Upon stimulation of cultured
microglia in vitro with α-synuclein protein, microglia
activation was demonstrated to be increased, leading to the
production of a large number of inflammatory factors and NO
(16). In the present study,
α-synuclein was used to stimulate the microglia cells, and the
results demonstrated that the microglia proliferation capacity was
increased; also, the secretion of NO, TNF-α and IL-1β in the cells
was increased, results which were consistent with those reported
previously.
CCL2 is a monocyte chemotactic factor. Recent
studies have shown that CCL2 expression is increased in the
peripheral blood and cerebrospinal fluid of patients with
neurodegenerative diseases, whereas the activity of microglia upon
inhibition of CCL2 expression was found to decrease as the
cognitive function of the biological organism was being restored
(17,18). CCL2 was also shown to be involved
in the inflammatory response of the central nervous system, and was
associated with the body's NO content and oxygen free radical
levels (19). Previous studies
have demonstrated that 0.05 ng/µl CCL2 may significantly promote
the proliferation of microglia (20). In the present study, microglia were
treated with 0.05 ng/µl CCL2 protein, and it was demonstrated that
CCL2 could promote the proliferation of microglia and the secretion
of NO, TNF-α and IL-1β.
In order to study further the effect of CCL2 on
neurons, the primary neurons were cultured in vitro and
treated with microglia culture medium containing CCL2 and
α-synuclein. The results demonstrated that the
CCL2/α-synuclein-treated microglia culture mixture was able to
promote neuronal apoptosis. Furthermore, the rate of neuronal
apoptosis was found to increase following treatment with a
combination of CCL2 and α-synuclein. Additionally, the expression
of cleaved caspase-3 and other components of the Akt signaling
pathway in neurons were detected: The level of cleaved caspase-3
protein was increased whereas that of p-Akt decreased, suggesting
that CCL2 promoted the proliferation of α-synuclein-induced
microglia, thus causing neuronal apoptosis which may be associated
with the Akt signaling pathway.
In conclusion, the present study has shown that CCL2
is able to promote the proliferation of α-synuclein-induced
microglia, via which the secretion of inflammatory factors and NO
by microglia is also promoted, thereby inducing neuronal apoptosis.
The pro-apoptotic mechanisms may be associated with the Akt
signaling pathway. The results thus obtained have laid a foundation
for further study of the pathogenesis of neurodegenerative
diseases.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Key Science
and Technology Project of Henan Province, China (grant no.
172102310685).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BY made substantial contributions to the design of
the present study. LZ, PM, QG, LS and LW performed the experiments
of cell culture, MTT, detection of NO and interpreted the data. LZ,
QG, LS and LM performed the experiments of ELISA and western
blotting. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was performed in strict accordance
with the recommendations given in the Guide for the Care and Use of
Laboratory Animals of the National Institutes of Health. The animal
use protocol was reviewed and approved by the Institutional Animal
Care and Use Committee (IACUC) of Xinxiang Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Glebov K, Löchner M, Jabs R, Lau T, Merkel
O, Schloss P, Steinhäuser C and Walter J: Serotonin stimulates
secretion of exosomes from microglia cells. Glia. 63:626–634. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang Y, Gao H, Zhang W, Zhang W and Fang
L: Thymoquinone inhibits lipopolysaccharide-induced inflammatory
mediators in BV2 microglial cells. Int Immunopharmacol. 26:169–173.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim M, Choi SY, Lee P and Hur J:
Neochlorogenic acid inhibits lipopolysaccharide-induced activation
and pro-inflammatory responses in BV2 microglial cells. Neurochem
Res. 40:1792–1798. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chun E, Lavoie S, Michaud M, Gallini CA,
Kim J, Soucy G, Odze R, Glickman JN and Garrett WS: CCL2 promotes
colorectal carcinogenesis by enhancing polymorphonuclear
myeloid-derived suppressor cell population and function. Cell Rep.
12:244–257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tsaur I, Rutz J, Makarević J, Juengel E,
Gust KM, Borgmann H, Schilling D, Nelson K, Haferkamp A, Bartsch G
and Blaheta RA: CCL2 promotes integrin-mediated adhesion of
prostate cancer cells in vitro. World J Urol. 33:1051–1056. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mbefo MK, Fares MB, Paleologou K, Oueslati
A, Yin G, Tenreiro S, Pinto M, Outeiro T, Zweckstetter M, Masliah E
and Lashuel H: Parkinson disease mutant E46 K enhances α-synuclein
phosphorylation in mammalian cell lines, in yeast and in vivo. J
Biol Chem. 290:9412–9427. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moree B, Yin G, Lázaro DF, Munari F,
Strohäker T, Giller K, Becker S, Outeiro TF, Zweckstetter M and
Salafsky J: Small molecules detected by second-harmonic generation
modulate the conformation of monomeric α-synuclein and reduce its
aggregation in cells. J Biol Chem. 290:27582–27593. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kempuraj D, Thangavel R, Yang E, Pattani
S, Zaheer S, Santillan DA, Santillan MK and Zaheer A: Dopaminergic
Toxin 1-Methyl-4-Phenylpyridinium, proteins α-synuclein and glia
maturation factor activate mast cells and release inflammatory
mediators. PLoS One. 10:e01357762015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shinozaki Y, Shibata K, Yoshida K,
Shigetomi E, Gachet C, Ikenaka K, Tanaka KF and Koizumi S:
Transformation of Astrocytes to a neuroprotective phenotype by
microglia via P2Y1 receptor downregulation. Cell Rep. 19:1151–1164.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mangino G, Famiglietti M, Capone C, Veroni
C, Percario ZA, Leone S, Fiorucci G, Lülf S, Romeo G, Agresti C, et
al: HIV-1 myristoylated Nef treatment of murine microglial cells
activates inducible nitric oxide synthase, NO2 production and
neurotoxic activity. PLoS One. 10:e01301892015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cox DJ, Field RH, Williams DG, Baran M,
Bowie AG, Cunningham C and Dunne A: DNA sensors are expressed in
astrocytes and microglia in vitro and are upregulated during
gliosis in neurodegenerative disease. Glia. 63:812–825. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Van der Perren A, Macchi F, Toelen J,
Carlon MS, Maris M, de Loor H, Kuypers DR, Gijsbers R, van den
Haute C, Debyser Z and Baekelandt V: FK506 reduces
neuroinflammation and dopaminergic neurodegeneration in an
α-synuclein-based rat model for Parkinson's disease. Neurobiol
Aging. 36:1559–1568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang Y and Le W: Differential roles of M1
and M2 microglia in neurodegenerative diseases. Mol Neurobiol.
53:1181–1194. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Harms AS, Cao S, Rowse AL, Thome AD, Li X,
Mangieri LR, Cron RQ, Shacka JJ, Raman C and Standaert DG: MHCII is
required for α-synuclein-induced activation of microglia, CD4 T
cell proliferation and dopaminergic neurodegeneration. J Neurosci.
33:9592–9600. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Daher JP, Volpicelli-Daley LA, Blackburn
JP, Moehle MS and West AB: Abrogation of α-synuclein-mediated
dopaminergic neurodegeneration in LRRK2-deficient rats. Proc Natl
Acad Sci USA. 111:9289–9294. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Codolo G, Plotegher N, Pozzobon T, Brucale
M, Tessari I, Bubacco L and de Bernard M: Triggering of
inflammasome by aggregated α-synuclein, an inflammatory response in
synucleinopathies. PLoS One. 8:e553752013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim RY, Hoffman AS, Itoh N, Ao Y, Spence
R, Sofroniew MV and Voskuhl RR: Astrocyte CCL2 sustains immune cell
infiltration in chronic experimental autoimmune encephalomyelitis.
J Neuroimmunol. 274:53–61. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kempuraj D, Thangavel R, Fattal R, Pattani
S, Yang E, Zaheer S, Santillan DA, Santillan MK and Zaheer A: Mast
cells release chemokine CCL2 in response to parkinsonian toxin
1-Methyl-4-Phenyl-Pyridinium (MPP+). Neurochem Res. 41:1042–1049.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bose S and Cho J: Role of chemokine CCL2
and its receptor CCR2 in neurodegenerative diseases. Arch Pharm
Res. 36:1039–1050. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gómez-Nicola D, Schetters ST and Perry VH:
Differential role of CCR2 in the dynamics of microglia and
perivascular macrophages during prion disease. Glia. 62:1041–1052.
2014. View Article : Google Scholar : PubMed/NCBI
|