Introduction
Postoperative cognitive dysfunction (POCD) refers to
an impactful decline in cognitive function that is commonly
presented in older patients after surgery (1). The characteristics of POCD usually
manifest as disorientation and impaired memory, concentration and
thinking, but the etiology of POCD remains unclear (2,3).
Moreover, there are various risk factors for POCD, including age,
cognitive function before surgery, duration of the surgery, and
respiratory complications and infection after surgery (4). Recently, POCD has been considered a
serious problem affecting people's health and has an influence on
both the outcome and elevated costs of patients, but until now,
only a few useful treatments have been discovered (5). POCD can last for several days or even
several years; consequently, the quality of life of the patients is
extremely disrupted, there is an increase in hospitalization and in
extra hospital nursing expenses, and POCD is also blamed for
increasing morbidity and mortality during surgery (2). According to previous, efficient
studies, it is suggested that the regulation of certain signaling
pathways are implicated in POCD (3,6–8).
Calcium regulation [such as phosphatidylinositol
3-kinase (PI3K) pathway] is closely related to neurodegenerative
diseases such as POCD (9).
According to Hu et al (10), the inhibition of PI3K is partially
involved in different stages of POCD, but the exact mechanism is
not clear. There are several articles about the negative regulation
between BACE1 and PI3K activation (11,12),
but the specific molecular mechanism is not clarified. The protein
kinase B (Akt) signaling pathway is considered one of the potential
mechanisms in the progression of POCD, and the expression of the
β-site amyloid precursor protein cleavage enzyme 1 (BACE1) gene is
downregulated by activating the PI3K/Akt pathway (6,12).
It has been demonstrated that deregulation of the PI3K pathway is
associated with prostate cancer and that myocardial viability is
partially improved by activation of the PI3K/Akt pathway (13,14).
Amyloid-β (Aβ) protein, a major factor causing neuritic plaques in
the brains of Alzheimer's disease (AD) patients, is generated by
BACE1 (15). Genetic studies have
found an important relationship between Aβ and neurodegenerative
diseases, and Aβ involves several pathways, such as PI3K and C-Jun,
involved in the development of the disease (16,17).
Besides, Aβ promotes the production of reactive oxygen species
(ROS) and induces neuronal apoptosis, and activation of iNOS
pathway induces neurotoxicity; Aβ upregulates intracellular calcium
concentration to regulate synaptic plasticity of dendritic cells
(16,17). However, the different size of Aβ
peptide region and the different biological process of the involved
pathway regulation lead to the unknown mechanism of Aβ in disease
(18,19). It has been proven that an increased
BACE1 level elevates Aβ production and plays a role in promoting AD
(20). Moreover, BACE1 is a main
target to modify the treatment for AD, and by inhibiting BACE1
expression, AD can be relieved and even prevented through
inhibiting the expression of BACE1 (21,22).
These evidences suggest that BACE1 might participate in the POCD
process by mediating the production of Aβ and regulating the PI3K
pathway. Therefore, the present paper aims to analyze the effects
of BACE1 gene silencing in rats with POCD through the PI3K/Akt
pathway, which will potentially provide an extra option for POCD
treatment.
Materials and methods
Model establishment
A total of 120 7-day-old male Sprague-Dawley (SD)
rats with a weight of 15 to 20 g were purchased from the
Experimental Animal Center of Lanzhou University Second Hospital.
Twenty rats without any treatment were enrolled in the control
group, and the remaining rats were placed in an organic glass box
(20×40×15 cm) and included in the model group. The inlet and
exhaust ports were at opposite sides of the box. A Vapor 2000
Anesthetic Vaporizer (Draeger Medical Equipment Co., Ltd.,
Shanghai, China) was attached to the inlet port to infuse oxygen
and isoflurane, and the 5250 Respiratory Gas Monitor (Ohmeda,
Louisville, CO, USA) was connected with the exhaust port to detect
the concentration of oxygen, isoflurane and carbon dioxide. An
electric heating blanket was placed on the bottom of the box to
keep the rats' body temperature at 38 to 39°C. The rats in the
control group inhaled pure oxygen for 4 h at a flow rate of 4
l/min. The rats in the model group inhaled isoflurane for 4 h, and
the concentration of isoflurane was 3.4 to 3.6% in 1 h, 2.1 to 2.3%
after 1 h and 1.7 to 1.8% after 2 to 4 h. The loss of a righting
reflex in the rats was considered the onset of anesthesia. All the
animal experiments in the study were in accordance with the Guide
for the Care and Use of Laboratory Animals (23) issued by the National Institutes of
Health.
BACE1 siRNA vector construction and
grouping
The small interfering RNA (siRNA) of BACE1 was
synthesized using the in vitro transcription method. The
complete sequence of BACE1 mRNA in rats was obtained from GenBank
(GenBank accession no. NM012104). The target sequence of siRNA was
5′-GACGCUCAACAUCCUGGUG-3′. According to the design principles for
short hairpin RNA (shRNA), a loop region, reverse complementary
sequence and transcription terminator were added to the sequence
above for the transcription of shRNA. Restriction enzyme sites Sal
I (GTCGAC), Hind III (AAGCTT) and Xho I (CTCGAG) and protective
bases were also added to the cloned fragments to obtain a more
efficient insertion into the vector. In addition, the
single-stranded DNA of BACE1 siRNA was obtained. The synthesized
single-stranded DNA was diluted to 20 pmol/l with sterile water,
and double-stranded DNA was obtained after annealing. The product
after annealing was connected to the linearized vector.
Escherichia coli DH5α was transformed, applied on flat
plates, and cultured overnight at 37°C. Two colonies were selected
with sterile toothpicks from each plate corresponding to each
sequence, lysed for 10 min at 95°C, and added into the reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
system with the primers of multiple cloning sites in the vectors. A
total of 5 µl of PCR product was obtained and analyzed by 2%
agarose gel electrophoresis to select the positive colonies. In
addition, the plasmid DNA was then extracted, and the sequences
were analyzed. The endotoxin-free DNA was extracted from the
transformed strain corresponding to the recombinant expression
vector with the identified correct sequence in strict accordance
with the instructions of the Plasmid Maxi Preparation kit (Shanghai
Yanhui Biotech Co., Ltd., Shanghai, China). After the successful
construction of the recombinant vector, the BACE1 siRNA plasmid and
blank plasmid were extracted and diluted to 0.1 to 3 µg/µl with
interventions to the rats on the first day after successful
modeling. In addition, 80 rats from the model group were equally
divided into 4 groups, as follows: (1) The si-BACE1 group: 10 µl of BACE1
siRNA plasmid DNA solution was injected through the caudal vein.
(2) The wortmannin group: PI3K
inhibitor wortmannin (S2758; Selleck Chemicals, Houston, TX, USA)
at a concentration of 2 mg/kg was injected through the caudal vein
30 min before anesthesia, and the model was established with the
same method described above. (3)
The negative control (NC) group: A total of 10 µl blank plasmid
without siRNA was transfected. (4)
The si-BACE1 + wortmannin group: PI3K inhibitor wortmannin (S2758;
Selleck Chemicals) at a concentration of 2 mg/kg was injected
through the caudal vein 30 min before anesthesia; the model was
established with the same method described above, and 10 µl BACE1
siRNA plasmid DNA solution was injected through the caudal vein
after modeling. At the same time, the rats in the control group and
model group were all injected with the same dosage of saline once a
day for 7 days.
Blood gas analysis
Four h after transfection, 200 µl of blood was
extracted from the left ventricle of the rats in each group, and an
I-STAT portable blood gas analyzer (Boyu Medical Equipment Co.,
Ltd., Shanghai, China) was applied to measure the blood glucose
level, pH, PaO2 and PaCO2.
Spontaneous locomotor activity
test
A ZZ-6 autonomic activity tester purchased from the
Beijing Institute of Respiratory Medicine (Beijing, China) was
applied to test the locomotor activity. The box was made of 4 black
opaque sealed cylinders with a diameter of 50 cm and a height of 40
cm containing 4 phototubes on the inner surface to record the
locomotion of rats. During the experiment, the rats were gently
placed in the box, and the lid was closed. The duration of each
test was 10 min, and the test was performed daily for 5 days. The
equipment was cleaned with water to prevent odors from interfering
with the judgment of the rats after each experiment. The room was
kept quiet before and after the experiment, and the temperature was
kept constant at 24°C.
Fear conditioning test
Fear conditioning was measured to evaluate the
cognitive function of rats using the fear conditioning analysis
system (XR.XC404; Shanghai Xinruan Information Technology Co.,
Ltd., Shanghai, China). On the first day, sound stimulation (2000
Hz, 90 dB, 30 sec) was conducted after 3 min of an adaptive phase,
and electric current stimulation (1 mA, 2 sec) started at the last
2 sec of sound stimulation and ended at the same time as the sound
stimulation, with an interval of 1 min. On the second day, cued and
contextual fear conditioning experiments were conducted. For the
contextual conditioning, the rats were placed in the same scene on
the first day without sound or electric current stimulation and
recorded for 8 min. For the cued conditioning, the rats were placed
in a different scene from the first day, and after 3 min of the
adaptive phase, sound stimulation (2000 Hz, 90 dB, 30 sec) was
released 3 times with an interval of 1 min without an electric
current and recorded for 8 min. Image analysis software (Shenzhen
RWD Life Science Co., Ltd., Shenzhen, China) was used to calculate
the percentage of stagnation time of the rats in cued and
contextual fear conditioning.
Morris water maze test
A water maze (Chinese Academy of Medical Sciences,
Beijing, China) was adopted with a diameter of 214 cm, a height of
50 cm, a depth of 30 cm and a black bottom. At the beginning of the
experiment, Chinese ink was dropped into the water to darken the
water. Four white spots were marked in equidistance
counter-clockwise on the wall of the pool. The four quadrants of
the pool were west, south, east and north, and a circular platform
(10 cm in diameter and 28 cm in height) was placed in the north
quadrant. A camera was installed right above the pool with a clear
vision of the entire water maze. The experiment was conducted in a
quiet room with artificial lighting. During the experiment, the
water temperature was 20 to 23°C, and the water was changed every
day. The rats were fasted for 4 h before the experiment, and their
fur was dried after the experiment. A week after the drug
injection, the rats were pretrained 3 times a day (with an interval
of 2 h each time) for 2 days. During the training, the rats were
randomly put in different quadrants of the pool with their heads
against the wall. The rats were guided to swim in a straight line
toward the platform and to stay on the platform for 30 sec.
For the orientation navigation experiment, the rats
were put into the water from different quadrants, each time with
their heads against the wall. When the rats entered the water, the
escape latency and swim distance were recorded. A Morris water maze
was performed in 15 rats of each group. The time period between the
rats getting into the water and finding the platform was defined as
the escape latency. The rats that could not find the platform in 60
sec were placed on the platform for 10 sec, and the escape latency
was recorded as 60 sec. The training was performed 4 times a day
for 5 days, and the locations of water entry were randomly chosen
in different quadrants every time. The average value of the entire
experiments was regarded as the ability of learning and memory, and
the escape latency represented the spatial learning ability of
rats. For the space exploration experiment, the platform was
removed the next day after the orientation navigation experiment.
The rats were put into the water from the contralateral quadrant of
the previous quadrant of the original platform. The swimming trace
for 120 sec, the time when the rats first reached the platform and
the number of times that the rats passed the target area within 120
sec was recorded to reflect the spatial learning ability of the
rats. After comparison and analysis, the memory ability of rats to
find the original platform was investigated. If the rats did not
pass the target area in 120 sec, then the time was recorded as 120
sec, and the number of passing times was 0. The data collection and
processing were accomplished by the Morris software.
RT-qPCR
After the behavioral experiments, the rats in each
group were sacrificed, the skulls were opened, and the brain
tissues were taken out, frozen immediately by liquid nitrogen and
stored at −80°C in a refrigerator. The total RNA was extracted from
brain tissues according to the instructions of the kit (Shanghai
Yanhui Biotech Co., Ltd.). The RNA was reversely transcribed into
cDNA. The total volume of the PCR reaction was 10 µl, including 1
µl of cDNA, 5 µl of 2X SYBR® Premix Ex Taq XmlI, 0.4 µl
of forward primer, 0.4 µl of reverse primer, and 3.2 µl of double
distilled water. The conditions were 2 min of predenaturation at
94°C, 30 cycles of 45 sec at 94°C, 45 sec at 55°C and 1 min at
72°C, and a final extension for 10 min at 72°C. GAPDH served as an
internal reference. The primer sequences are shown in Table I. PCR products were detected by 2%
agarose gel electrophoresis. The integrated optical density (OD)
was calculated as the gray value multiplying the area. The relative
gene expression was represented by the 2−ΔΔCq method and
the ratio of integrated OD of the target gene to the reference gene
(24). In addition, ∆Cq
represented the difference between the Cq value of the target gene
and the Cq value of the reference gene (25). RT-qPCR was also used to detect the
mRNA expression of caspase-3 and PI3K/Akt signaling pathway-related
genes.
 | Table I.Primer sequences for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Forward primer
sequence (5′-3′) | Reverse primer
sequence (5′-3′) |
|---|
| BACE1
(NM012104) |
TGGTGGACACGGGCAGTAGTAA |
TCGGAGGTCTCGGTATGTACTGG |
| Caspase-3
(NM012922) |
TACCCTGAAATGGGCTTGTGT |
GTTAACACGAGTGAGGATGTG |
| PI3K
(NM053481) |
TGGACGGCGAAGTAAAGCATT |
AGTGTGACATTGAGGGAGTCG |
| Akt (NM033230) |
CTCATTCCAGACCCACGAC |
ACAGCCCGAAGTCCGTTA |
| GAPDH
(NM017008) |
GACAACTTTGGCATCGTGGA |
ATGCAGGGATGATGTTCTGG |
Western blot analysis
Brain tissues were added to a protein extraction
reagent (Beijing Biolab Technology Co., Ltd., Beijing, China)
according to a 1: 10 (g/l) ratio. The components of the reagent
were 20 mM Tris (pH, 7.5), 150 mM NaCl, 1% Triton X-100, Na2HPO4,
β-glycerophosphate, ethylenediamine tetraacetic acid (EDTA),
Na3VO4, PVP40 and leupeptin. The extracted protein was centrifuged
at 12,500 rpm for 15 min to obtain the supernatant, and the protein
was quantitatively determined by the bicinchoninic acid (BCA) kit
(Beyotime Institute of Biotechnology, Jiangsu, China). A sample
with the same amount of the protein was taken and diluted to 20 µl
with 4× loading buffer and normal saline. After denaturation at
100°C for 5 min, sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) was prepared according to the relative
molecular weight of the target protein. The proteins were
transferred onto the membrane according to the concentration and
loading quantity of the protein. Five percent skim milk was used to
block the membrane for 1 h. After gently shaking at 37°C for 2 h,
GAPDH (ab9485, 1:5,000) and the primary antibodies BACE1 (ab108394,
1:1,000), PI3K (ab86714, 1:1,000), phosphorylated (p)-PI3K
(ab182651, 1:1,000), Akt (ab8805, 1:500), p-Akt (ab8933, 1:1,000),
caspase-3 (ab13847, 1:500) and cleaved-caspase-3 (ab49822, 1:500;
all purchased from Abcam, Cambridge, MA, USA) were added for
incubation at 4°C overnight. The membrane was washed 3 times, each
time for 5 min. POD-conjugated goat anti-rabbit antibody (A5795,
1:5,000; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added
to incubate for 1.5 h at 37°C, and the membrane was washed 3 times
for 10 min each time. Luminescent reagent was added, and the film
was exposed by X-ray, developed and fixed in the darkroom. Quantity
One software was employed for the semiquantitative analysis for
BACE1 expression. The ratio of the gray value of the internal
reference to that of BACE1 on film represented the expression level
of BACE1. This method was also applied to detect the protein
expression of caspase-3 and PI3K/Akt signaling pathway-related
genes.
Hematoxylin and eosin (HE)
staining
The brain tissues were removed, and the hippocampus
of one side of the cerebral hemisphere was separated on the ice,
immediately frozen by liquid nitrogen and stored at −80°C in the
refrigerator. The other side of the cerebral hemisphere was fixed
in a 500 ml mixture of 2% paraformaldehyde and 2% glutaraldehyde at
4°C for 2 h, and the brain was then removed and soaked in a freshly
mixed fixative solution at 4°C for 12 h. The hippocampus was
separated the next day. The tissues in the hippocampal CA1 region
were paraffin-embedded, and serial coronal sections (4 mm) were cut
behind the optic chiasm. The sections were baked at 60°C overnight
and dewaxed in xylene I and xylene II for 20 min. The sections were
placed into 100, 95, 80 and 70% ethanol for 5 min respectively,
stained by hematoxylin for 10 min, and rinsed by running water for
15 min. Eosin was used to stain the sections for 30 sec, and double
distilled water was used to wash the sections. After the sections
were dehydrated by ethanol, cleared in xylene and sealed with
neutral balsam, the ultrastructure and number of pyramidal cells in
the hippocampal CA1 area were observed and photographed using an
electron microscope.
TUNEL staining
Using the same method of HE staining, hippocampal
CA1 tissues of rats were treated by TUNEL staining. Then, they were
cut into sections, and later, these tissues were dewaxed in xylene
I and II for 20 min and then were placed in 100, 100, 95, 80 and
70% ethanol for 5 min separately. After washing with phosphate
buffered saline (PBS) twice (5 min each time), the tissues were
fixed in 4% paraformaldehyde for 15 min. After again washing twice
with PBS (5 min each time), then 100 µl proteinase K (20 µg/ml) was
added to each section. After being treated at room temperature for
15 min, the tissues were twice washed in PBS (for 5 min each).
Then, the TUNEL kit of Promega Corporation (Madison, WI, USA; cat.
no. G3250) was used for staining. The specific steps were in line
with the instructions. After adding 100 µl equilibrium liquid, the
tissues were put in a wet box for balance for 10 min. The TUNEL
reaction mixture was made as follows: the treatment group was mixed
with 1 µl rTdT+1 µl biotin-labeled dUTP + 98 µl equilibrium liquid,
and the negative control group was mixed with threefold-distilled
water rather than rTdT. In the positive control group, the tissues
were incubated after adding 100 µl DNase 1 buffer for 5 min. After
removing the liquid, the tissues were enzymatically digested by
adding 100 µl DNase 1 (10 U/ml) for 10 min. After washing with
deionized water 4 times and with PBS for 5 min, 100 µl TUNEL
reaction mixture was added to the sample. Then, the sample was
placed in a dark wet box after being covered by a cover glass or
sealing film for reaction at 37°C × 1 h. The TUNEL reaction was
stopped after the sample was immersed in 2× SSC for 15 min. After
being washed in PBS 3 times (5 min for each), the sample was closed
in peroxidase (POD), immersed in 0.3% H2O2 for 15 min, and then
again immersed for washing 3 times with PBS (for 5 min each). The
sample was mixed with 100 µl streptavidin-labeled HRP (diluted
based on 1:500 of PBS) for a 30 min reaction and immersed by again
washing 3 times with PBS (for 5 min each). Subsequently, 100 µl DAB
mixture (50 µl DAB + 50 µl DAB substrate buffer + 50 µl H2O220 × +
950 µl threefold-distilled water) was supplemented for a 10 min
reaction. When a light brown background appeared under the
microscope, the sample was rinsed with deionized water and redyed
with hematoxylin for 3 sec. Then, the sample was immediately washed
with running water. After being dehydrated by gradient alcohol (50,
70, 85, 95, 100, 100%, 1 min for each) and cleared by xylene II,
the sample was mounted by neutral balata. Finally, the positive
apoptotic cells were observed under a microscope, with 10 pictures
taken in each group to determine the apoptosis rate of positive
cells in the hippocampal CA1 region.
Enzyme-linked immunosorbent assay
(ELISA)
The other side of the frozen hippocampus
(approximately 100 mg) was homogenized and centrifuged, and then
the supernatant was extracted according to the instructions of the
ELISA kits for TNF-α, IL-1, IL-6, Aβ and amyloid precursor protein
(APP) purchased from Shanghai Westang Biotech Co., Ltd., (Shanghai,
China). The kits were placed at room temperature for 20 min, and
the washing solution was prepared. A total of 10 standard wells
were arranged on the ELISA plate, including 2 blank wells without
samples or ELISA reagents. The standard solution was subjected to
gradient dilution, and the standard curve was created. The sample
was diluted 10-fold with normal saline and then added to the wells
of the ELISA plate. The plate was gently shaken and incubated at
37°C for 30 min after being sealed. The liquid in the wells was
removed, and the washing solution was added and then removed after
30 sec, and the step was repeated 5 times before drying the plate.
In addition, 50 µl of ELISA reagent was added and incubated at 37°C
for 30 min, and the liquid in the wells was removed. The washing
solution was added and then removed after 30 sec, and the step was
repeated 5 times before drying the plate. Next, 50 µl of
chromogenic reagent A and 50 µl of chromogenic reagent B was added
to each well and incubated at 37°C for 15 min in the dark after
shaking, followed by the addition of 50 µl stop solution. The
microplate reader (Bio-Rad, Hercules, CA, USA) was applied to
measure the OD value (450 nm) of each well at 10 min. The
concentration standard curve was drawn with the OD value as the
vertical coordinate and the standard solution as the horizontal
coordinate. According to the OD value, the concentration of the
sample was determined and recorded.
Statistical analysis
SPSS v.22.0 software (IBM Corp., Armonk, NY, USA)
was used for statistical analysis. All tests were repeated three
times, and the means and standard deviations were calculated.
Measurement data are expressed as the mean ± standard deviation.
The comparison between groups was conducted by one-way analysis of
variance, which was combined with Tukey's Honest Significant
Difference post hoc test, and repeated measures analysis of
variance combined with the Bonferroni post hoc test was used to
analyze the differences in a group at different time points.
P<0.05 was considered to indicate a statistically significant
difference.
Results
No hypoxia or carbon dioxide
accumulation were found in each group
Measurement of the arterial blood gas index and
blood glucose was carried out. The results showed that among the
rats of 6 groups, all the pH values were in the normal range
without hypoxia or carbon dioxide accumulation in each group. The
pH, PaO2, PaCO2 and blood
glucose level had no significant difference, as shown in Table II (all P>0.05).
| Table II.Blood gas indices and blood glucose
levels in the rat artery among the six groups. |
Table II.
Blood gas indices and blood glucose
levels in the rat artery among the six groups.
| Group | pH |
PaCO2 (mmHg) |
PaO2 (mmHg) | Blood glucose
(mmol/l) |
|---|
| Control | 7.88±0.12 | 37.81±5.11 | 105.21±6.40 | 5.14±0.82 |
| Model | 7.73±0.17 | 39.47±4.30 | 100.24±8.30 | 4.84±0.75 |
| NC | 7.84±0.23 | 38.62±4.62 | 102.63±5.64 | 4.64±0.60 |
| Si-BACE1 | 7.78±0.16 | 39.33±5.20 | 101.53±7.30 | 4.05±0.70 |
| Si-BACE1 +
wortmannin | 7.90±0.25 | 38.93±4.80 | 103.15±6.50 | 4.93±0.55 |
| Wortmannin | 7.80±0.27 | 38.12±4.04 | 102.85±6.70 | 4.75±0.96 |
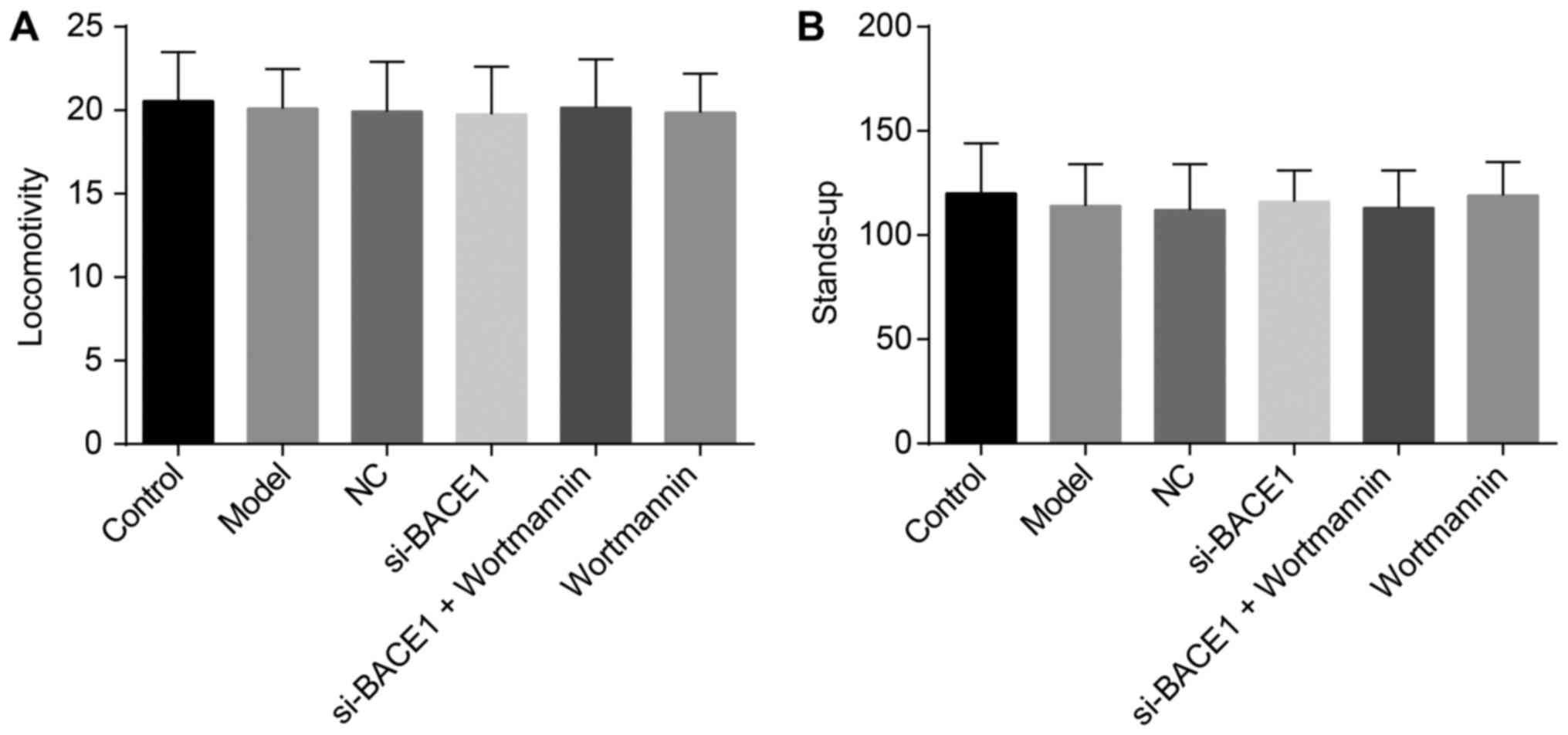
The spontaneous locomotor activity and
standing up of rats exhibited no significant differences among the
six groups
According to the results of experiments on the
spontaneous locomotor and standing activities of rats (Fig. 1), no significant differences were
found in the standing and locomotor activities of rats in each
group (all P>0.05).
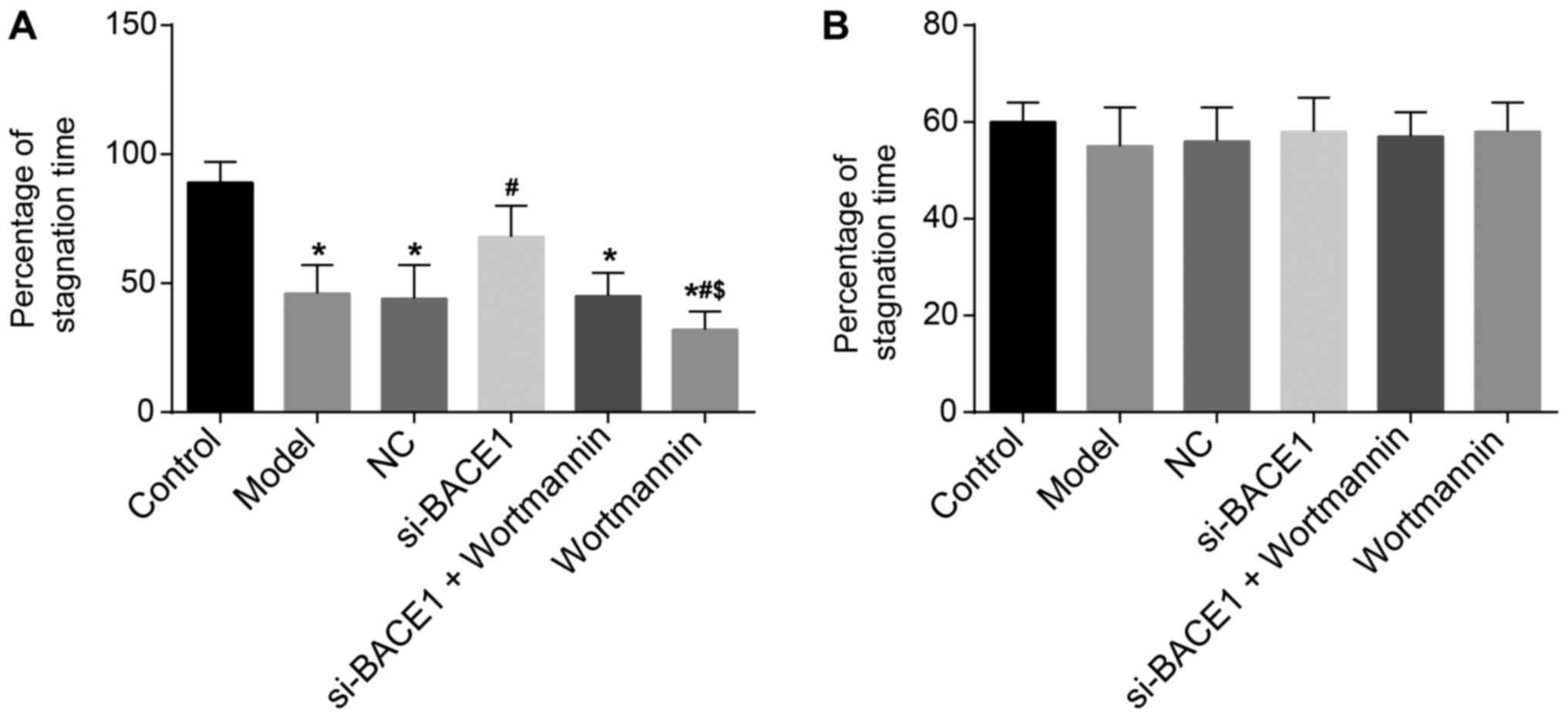
BACE1 gene silencing increased the
percentage of stagnation time in the fear conditioning test of
rats
The contextual fear conditioning test was performed
for observing the percentage of stagnation time of rats. Its
results indicated that compared with the control group, the model,
NC, si-BACE1 + wortmannin and wortmannin groups had a lower
percentage of stagnation time (all P<0.05). The percentage of
stagnation time in the si-BACE1 group was significantly higher than
that in the model group (P<0.05), while the percentage of
stagnation time in the wortmannin group was remarkably lower than
that in the model group and si-BACE1 + wortmannin group (all
P<0.05). There were no significant differences in the percentage
of stagnation time among the si-BACE1 + wortmannin, model and NC
groups (all P>0.05). In the cued fear conditioning test, the
percentage of stagnation time was not significantly different in
the 6 groups (all P>0.05, Fig.
2). Consequently, the results of the study confirmed that the
percentage of stagnation time of rats can be increased by BACE1
gene silencing.
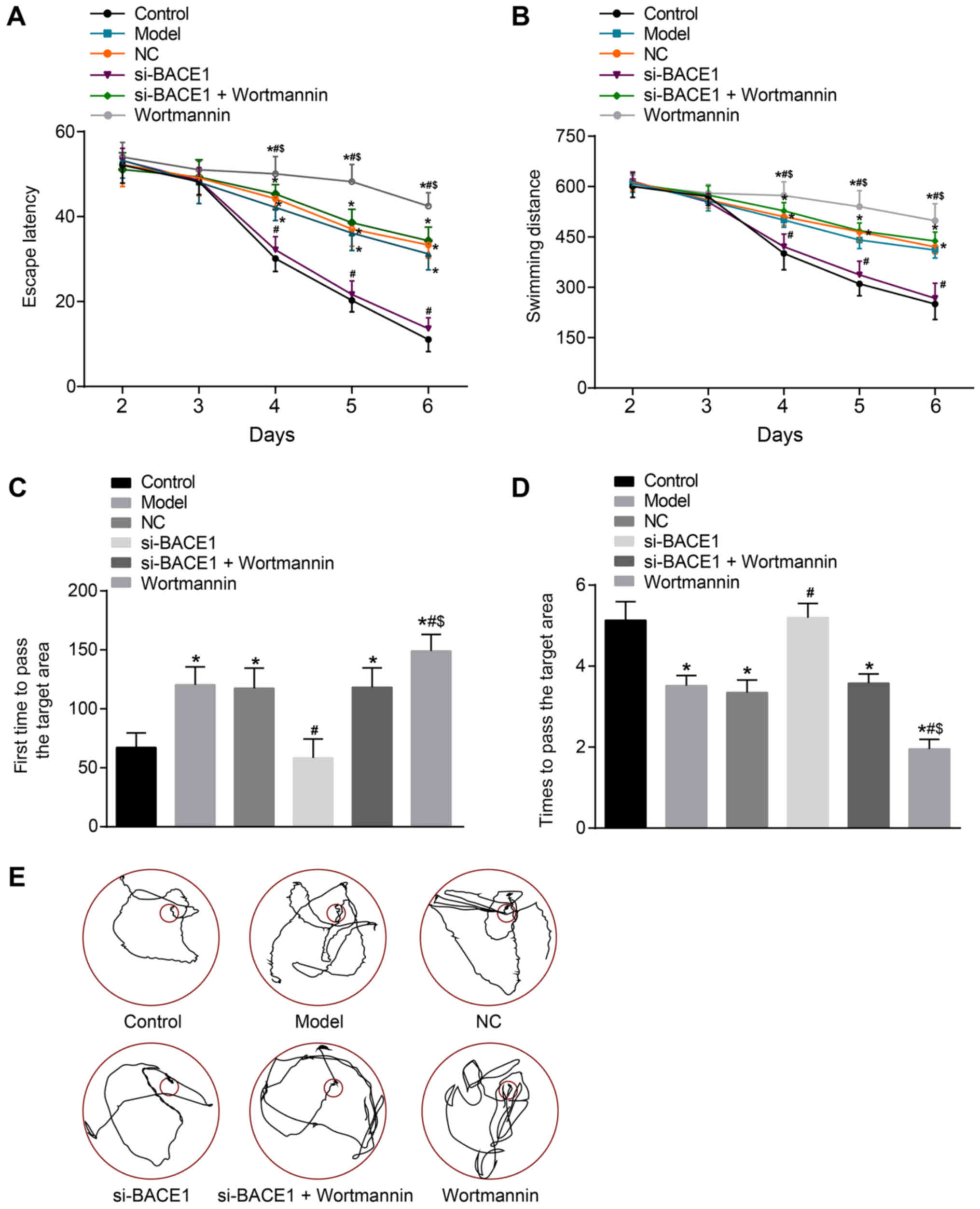
BACE1 silencing enhanced the spatial
memory ability of rats
Several experiments were carried out to measure the
spatial memory ability of rats. The escape latency and swimming
distance of rats in the control group and si-BACE1 group decreased
significantly during the 5-day training. However, the escape
latency and the swimming distance were longer and decreased slowly
in the model, NC, si-BACE1 + wortmannin and wortmannin groups
compared to the control group during the fourth to sixth day
(P<0.05). On the seventh day of the space exploration
experiment, in comparison to the control group, the model, NC and
si-BACE1 + wortmannin groups spent more time to first pass the
target area and passed the target area remarkably fewer times (all
P<0.05). Compared to the model group, the escape latency and
swimming distance was obviously reduced from the fourth to the
sixth day, the time that the rats spent in first passing the target
area shortened, and the times of passing the target area rose in
the si-BACE1 group. However, when compared to the model and
si-BACE1 + wortmannin group, the wortmannin group had a
significantly longer escape latency and swimming distance from the
fourth to sixth day, and the time the rats spent in first passing
the target area extended and the times of passing the target area
reduced (all P<0.05). In addition, there was no significant
difference in the escape latency, swimming distance and exploration
time among the model group, NC group and si-BACE1 + wortmannin
group (all P>0.05, Fig. 3). In
brief, the findings verified that BACE1 gene silencing enhances the
spatial memory ability of rats.
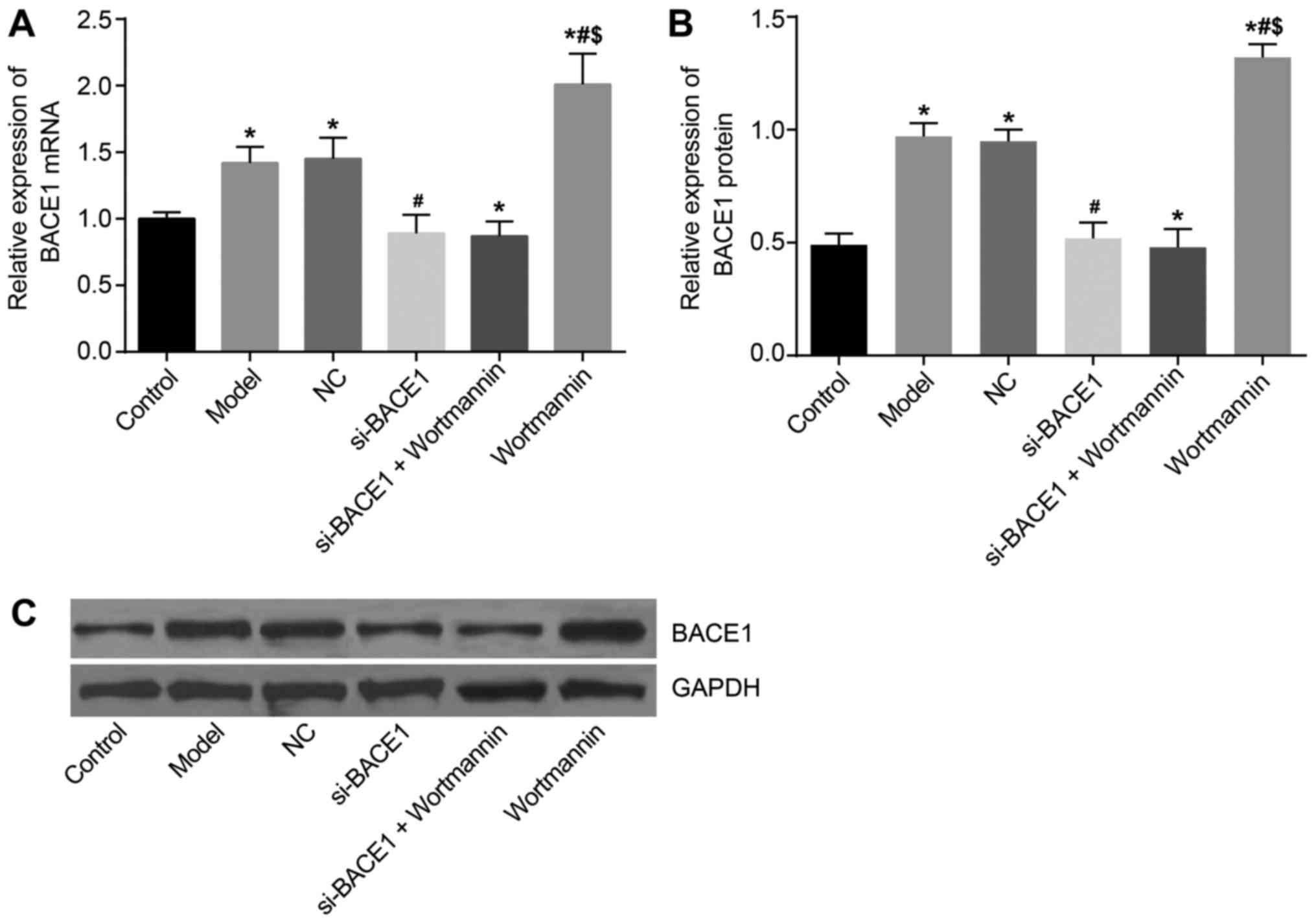
mRNA and protein expression of BACE1
was the highest in the wortmannin group
RT-qPCR results showed that compared with the
control group, the expression of BACE1 in rats in the model, NC and
wortmannin groups was upregulated. In addition, the expression in
the si-BACE1 group and si-BACE1 + wortmannin group was lower than
that in the model group. The expression of BACE1 in the wortmannin
group was higher than that in the model group (P<0.05). There
was no significant difference between the NC and model groups
(P>0.05) (Fig. 4A), and the
results of western blot analysis detection of protein were
consistent with those of RT-qPCR (Fig.
4B and C).
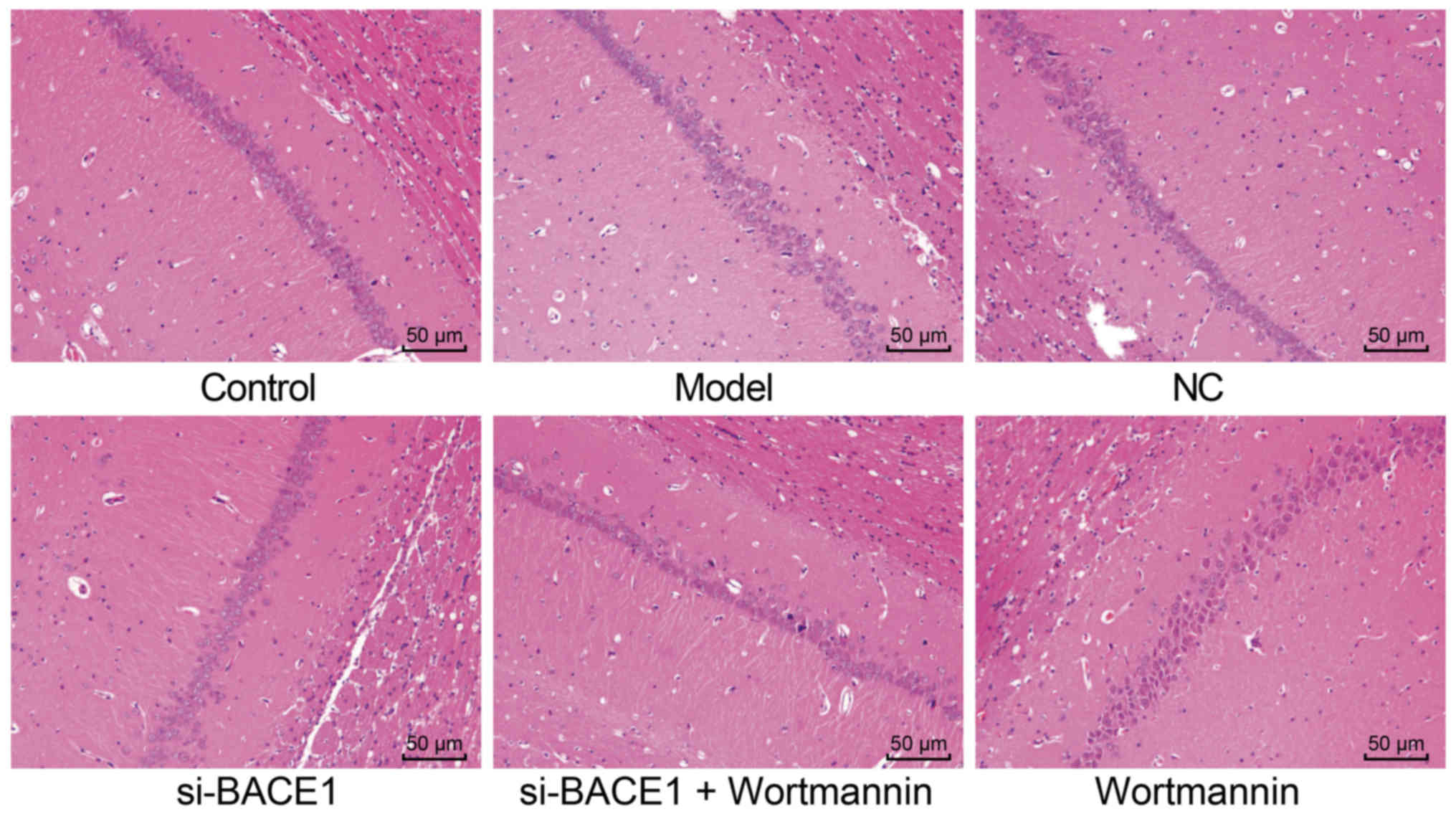
Silencing BACE1 improved the
pathological state of immature rats induced by isoflurane
anesthesia
Isoflurane anesthesia was performed, and its results
showed that the intensively stained hippocampal pyramidal cells
were annular. The hippocampal CA1 neurons in the control group were
regularly arranged, and the cells were morphologically normal and
orderly arranged. The neurons were of similar size with clear
structures of mitochondria and synapse. The synapses were evenly
distributed, the presynaptic and postsynaptic membranes were close,
and the synaptic vesicles were abundant and concentrated. The
hippocampal CA1 neurons in the model, NC and si-BACE1 + wortmannin
groups were irregularly arranged, without a clear structure of
mitochondria. The synaptic vesicles were reduced and scattered, and
the presynaptic and postsynaptic membranes were not clear. The
synaptic cleft widened, and the synaptic vesicles were sparsely
distributed. The nuclei were intensively stained with karyopyknosis
and dissolution, and there was vacuolization in some neurons. In
the si-BACE1 group, the hippocampal CA1 neurons were rather
regularly arranged, with a clear structure of mitochondria. The
synapses were evenly distributed, and the presynaptic and
postsynaptic membranes were rather close, with relatively close
synaptic vesicles. There was less pathological damage in the
si-BACE1 group compared to the model group, but the pathological
damage in the wortmannin group worsened compared to that in the
model and si-BACE1 + wortmannin groups (Fig. 5). Taken together, silencing BACE1
improves the pathological state of immature rats induced by
isoflurane anesthesia.
BACE1 gene silencing decreased TNF-α,
IL-1β, IL-6, APP and Aβ levels in the rat hippocampus
Next, TNF-α, IL-1β, IL-6, APP and Aβ levels in the
rat hippocampi among the six groups were detected. The TNF-α level
in the hippocampus was similar among the 6 groups (all P>0.05).
Compared to the control group, the hippocampal IL-1β, IL-6, APP and
Aβ levels in the model, NC, si-BACE1 + wortmannin and wortmannin
groups were remarkably elevated (all P<0.05). The hippocampal
IL-1β, IL-6, APP and Aβ levels in the si-BACE1 group were notably
lower than those in the model group (all P<0.05). The levels of
IL-1β, IL-6, APP and Aβ in the wortmannin group were significantly
higher than those in the si-BACE1 + wortmannin and model groups
(all P<0.05). No differences were noted among the model, NC and
si-BACE1 + wortmannin groups or between the si-BACE1 and control
groups (all P>0.05, Table
III). These results suggested that silencing BACE1 attenuates
the inflammatory response and the levels of APP and Aβ in
hippocampal tissues.
| Table III.TNF-α, IL-1β, IL-6, APP and Aβ levels
in the rat hippocampus of the six groups. |
Table III.
TNF-α, IL-1β, IL-6, APP and Aβ levels
in the rat hippocampus of the six groups.
| Group | TNF-α (pg/g) | IL-1β (pg/g) | IL-6 (pg/g) | APP | Aβ |
|---|
| Control | 130.23±18.07 | 108.03±16.12 | 47.24±7.06 | 10.04±3.45 | 205.24±19.45 |
| Model | 134.12±16.06 |
169.45±19.04a |
84.24±9.12a |
25.67±4.02a |
390.74±15.06a |
| NC | 132.14±14.73 |
166.54±20.34a |
82.32±6.67a |
26.78±3.46a |
387.19±16.09a |
| si-BACE1 | 129.35±15.09 |
120.78±18.02b |
52.35±5.35b |
9.76±2.67b |
210.36±10.92b |
| si-BACE1 +
wortmannin | 136.78±17.07 |
168.65±21.02a |
80.76±7.14a | 24.9 0±
2.26a |
396.54±13.67a |
| Wortmannin | 133.67±12.08 |
192.08±22.12a–c |
104.21±9.45a–c |
44.32±5.34a–c |
450.45±20.09a–c |
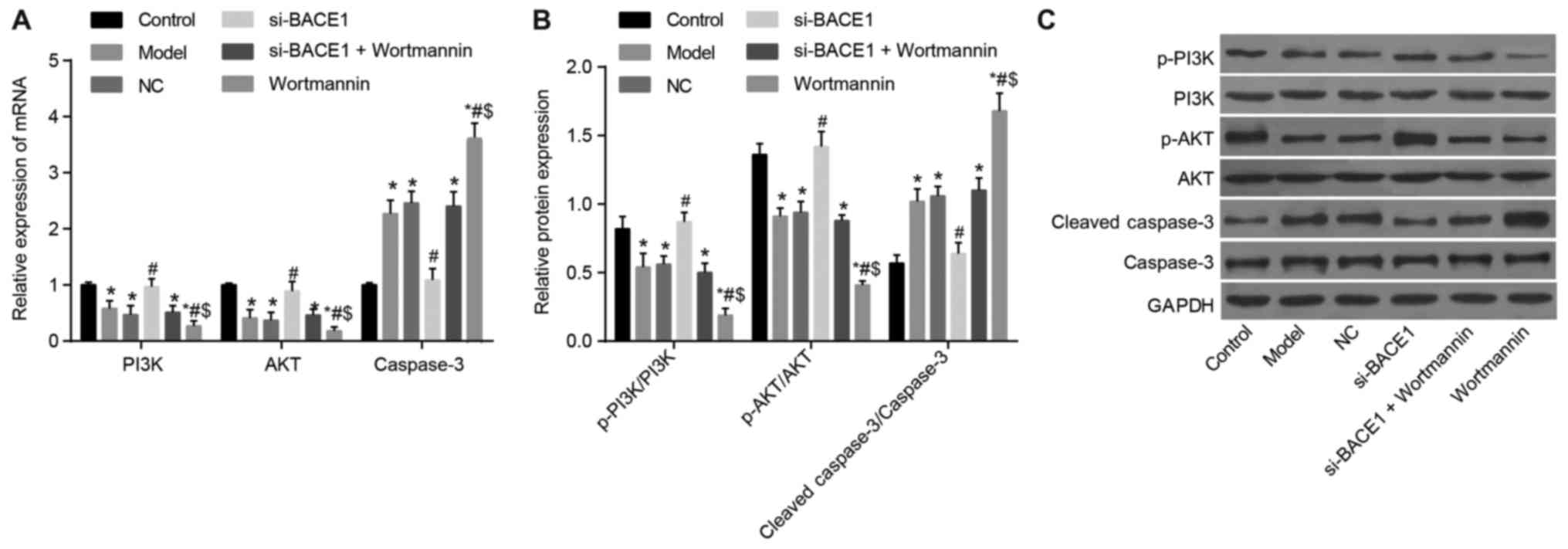
BACE1 gene silencing activated the
PI3K/Akt signaling pathway
RT-qPCR was used to detect the mRNA expression of
related genes. The results revealed that the PI3K and Akt mRNA
expression levels in the hippocampus among the model, NC, si-BACE1
+ wortmannin and wortmannin groups exhibited a notable decrease,
while the mRNA expression of caspase-3 remarkably increased in
comparison to the control group (all P<0.05). Compared to the
model and NC groups, the relative mRNA expression levels of PI3K
and Akt in the si-BACE1 group were significantly elevated, but the
mRNA expression of caspase-3 was reduced (all P<0.05); the
relative mRNA expression levels of PI3K and Akt were significantly
lower but the mRNA expression level of caspase-3 was remarkably
higher in the wortmannin group than in the si-BACE1 + wortmannin
group (all P<0.05). The mRNA expression levels of PI3K and Akt
in the wortmannin group were lower and the caspase-3 protein
expression level was significantly higher than those in the
si-BACE1 + wortmannin group (all P<0.05). The results of western
blot analysis demonstrated that the protein expression levels of
PI3K and Akt and the extent of PI3K and Akt phosphorylation in the
model, NC, si-BACE1 + wortmannin and wortmannin groups declined,
while the protein expression level of caspase-3 obviously increased
compared to that in the control group (all P<0.05). However, the
si-BACE1 group and control group exhibited no significant
difference (P>0.05). The protein expression levels of
p-PI3K/PI3K and p-AKT/AKT in the si-BACE1 group were higher, and
the cleaved-caspase-3/caspase-3 expression level decreased notably
compared with the levels in the model and NC groups (all
P<0.05), and the wortmannin group showed the opposite tendency.
The protein expression levels of PI3K and Akt and the extent of
PI3K and Akt phosphorylation in the wortmannin group were lower
than those in the si-BACE1 + wortmannin group, and the caspase-3
protein expression level was significantly higher (all P<0.05).
The PI3K, Akt and caspase-3 expression levels in the model, NC and
si-BACE1 + wortmannin groups exhibited no significant difference
(all P>0.05, Fig. 6). All these
results indicated that BACE1 gene silencing activates the PI3K/AKT
signaling pathway.
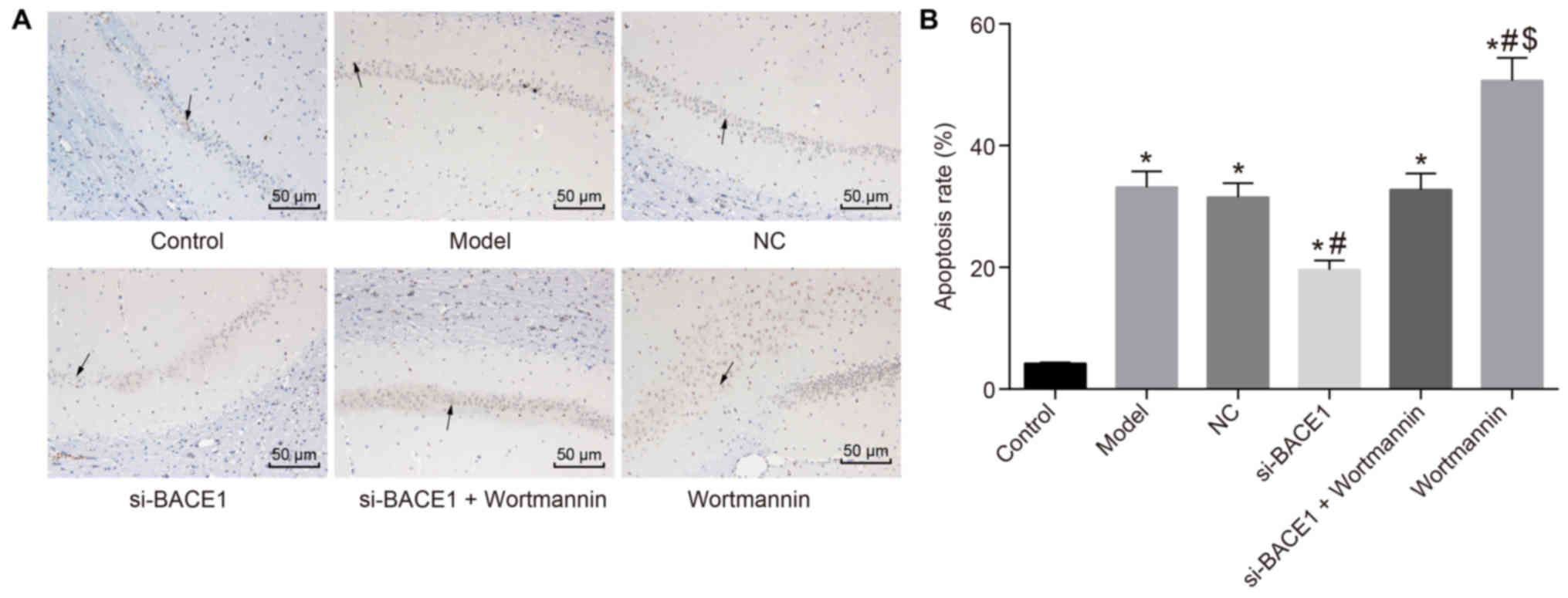
Silencing BACE1 promoted the
activation of the PI3K/Akt signaling pathway, thereby inhibiting
the apoptosis of the hippocampal CA1 region
To further prove that increased expression of
caspase3 induced cell apoptosis in the hippocampal CA1 region of
rats, we performed TUNEL staining. The results showed that in the
model, NC, si-BACE1 + wortmannin and wortmannin groups, the
positive rate of TUNEL staining in rat hippocampus CA1 was
significantly higher than that in the control group. Compared with
the model and NC groups, the si-BACE1 group had a lower positive
rate in the hippocampal CA1 area, and the positive rate in the
wortmannin group was significantly higher. In contrast to the
si-BACE1 + wortmannin group, the wortmannin group had an increased
positive rate of TUNEL staining in the hippocampal CA1 region
(P<0.05). There was no significant difference in the model, NC
and si-BACE1 + wortmannin groups (P>0.05; Fig. 7). Consequently, the findings
revealed that silencing BACE1 can activate the PI3K/Akt signaling
pathway, thus inhibiting the apoptosis of hippocampal CA1
region.
Discussion
It has been suggested that at present, few
satisfactory treatments for POCD are achieved and that use of
anesthetics is correlated with POCD (5). However, interestingly, according to
previous studies, various examples have been given to demonstrate
the potential role of signaling pathways in POCD (3,6–8),
indicating that studies of POCD targeting the regulation of
signaling pathways might be of great importance to the development
of treatments for POCD. In this study, the effects of BACE1 gene
silencing on POCD after isoflurane anesthesia via the PI3K/Akt
signaling pathway were studied in immature rats, and the results
might provide some evidence for the improvement of POCD treatment
with BACE1 gene silencing through activation of the PI3K/Akt
signaling pathway and inhibition of the generation of Aβ by
APP.
Initially, in this study, BACE1 gene silencing was
found to activate the PI3K/Akt signaling pathway. The cleavage of
the APP by enzymes, which are generally denoted as β- and
γ-secretase, is a crucial process in the pathological mechanism of
AD (26). BACE1, which is highly
expressed in the pancreas, is an aspartyl protease that is bound to
the membrane and is closely related to AD, leading to the
generation of Aβ from APP (27).
The inhibition of BACE1 along with acetylcholinesterase and
butyrylcholinesterase plays a key role in preventing and treating
AD (28). PI3K has been discovered
to have an important role in regulating many cellular processes
such as the survival, proliferation and differentiation of cells,
and Akt is expressed as AKT1, AKT2 and AKT3 encoded separately by
the PKBα, PKBβ, and PKBγ genes (29). There are multiple studies
indicating that activation of the PI3K/Akt signaling pathway
inhibits the growth and survival of cancer cells (29,30).
Additionally, wortmannin, as a specific inhibitor of PI3K, could
decrease the expression of PI3K and Akt. All PI3Ks are suppressed
by the drugs wortmannin and LY294002, of which wortmannin is a
commonly used reagent in cell biology and has been applied to
inhibit DNA repair, receptor-mediated endocytosis and cell
proliferation (31). A previous
study suggested that wortmannin, a PI3K/Akt inhibitor, decreases
the inflammatory cytokines in severe acute pancreatitis (SAP) rats,
demonstrating that its regulatory mechanisms may occur via the
suppression of NF-κB and p38 MAPK activity (32).
Furthermore, the levels of IL-1β, IL-6, APP and Aβ
in the rat hippocampus were remarkably reduced in the si-BACE1
group compared to the model group. A previous study discovered that
partial inhibition of the BACE1 gene had an influence on the APP's
β-cleavage to decrease the Aβ levels in the brain to improve the
cognitive dysfunctions associated with AD, including implicit and
explicit memory components (33).
The soluble oligomers of Aβ, the main component of senile plaques
in the brain, are considered to lead to the dysfunction of synapses
and cognition during the early phases of AD (34,35).
IL-1β, a proinflammatory cytokine, leads to neurogenic inflammation
in many central nervous system diseases as well as inflammatory
reaction development in the brain, and it has been revealed that
POCD has a certain correlation with the concentrations of
peripheral markers of inflammation, especially IL-6 and S-100β
(2). In addition, IL-1β
overexpression is associated with metaplasia, chronic gastritis,
severe stunted growth and even an elevated risk of solid
malignancy, suggesting that chronic inflammation triggers the
initiation and development of cancers (36). IL-6 is a cytokine involved in the
pathological mechanism of rheumatoid arthritis by its various
effects on immunological and inflammatory reactions, which also
plays a regulatory part in the process of metabolism, regeneration
and neurons (37,38).
In addition, isoflurane caused POCD in immature rats
via the regulation of IL-1β and caspase-3. It has been reported
that caspase-3 is an indicator of neuronal apoptosis caused by
increased expression of caspase-3 in the development of POCD
(39). IL-1β is an indicator of
the inflammatory response in the nervous system (40). These are two independent indicators
with no direct correlation, both of which are the factors related
to POCD. POCD is a type of severe neural sequela occurring after
anesthesia and surgery and is associated with anesthesia duration
and advanced age (41). At
present, a study has reported that isoflurane can result in a
significantly higher incidence of POCD (42). Chen et al (43), have reported that caspase-3, a
regulator of neuroapoptosis induced by isoflurane, plays a negative
regulatory role in the apoptosis of hippocampal neural precursor
cells (NPCs) and that NPCs are related to cognition impairment. In
addition, Lin et al (44),
have confirmed that isoflurane exposure may result in POCD in rats,
which has a correlation with cell damage and overexpression of
inflammatory mediators in the hippocampus, induced by isoflurane.
Isoflurane anesthesia has been found to increase the IL-1 level in
streptozotocin-induced diabetic rat models and aggravate
streptozotocin-induced cognitive impairment (45).
In conclusion, the present study reveals that BACE1
gene silencing activates the PI3K/Akt signaling pathway and
inhibits the generation of Aβ by APP, thus improving POCD induced
by isoflurane anesthesia in rats. The present study provides some
ideas for the development of new BACE1-targeting inhibitors that
can relieve the symptoms of POCD.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the corresponding author upon
reasonable request.
Authors' contributions
YBW, JQX, WL and RZZ made substantial contributions
to the design of the present study. YBW, JQX, WL collated the data,
and designed and developed the data in the manuscript. RZZ, SHH and
YHX performed data analyses and produced the initial draft of the
manuscript. All authors have read and approved the final submitted
manuscript.
Ethics approval and consent to
participate
All animal experimentation was approved by the
Animal Ethics Committee of Lanzhou University Second Hospital
(Lanzhou, China) and abided by relevant protocols.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jungwirth B, Zieglgansberger W, Kochs E
and Rammes G: Anesthesia and postoperative cognitive dysfunction
(POCD). Mini Rev Med Chem. 9:1568–1579. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Peng L, Xu L and Ouyang W: Role of
peripheral inflammatory markers in postoperative cognitive
dysfunction (POCD): A meta-analysis. PLoS One. 8:e796242013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ma Y, Cheng Q, Wang E, Li L and Zhang X:
Inhibiting tumor necrosis factor-α signaling attenuates
postoperative cognitive dysfunction in aged rats. Mol Med Rep.
12:3095–3100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ballard C, Jones E, Gauge N, Aarsland D,
Nilsen OB, Saxby BK, Lowery D, Corbett A, Wesnes K, Katsaiti E, et
al: Optimised anaesthesia to reduce post operative cognitive
decline (POCD) in older patients undergoing elective surgery, a
randomised controlled trial. PLoS One. 7:e374102012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liebert AD, Chow RT, Bicknell BT and
Varigos E: Neuroprotective effects against POCD by
Photobiomodulation: Evidence from assembly/disassembly of the
cytoskeleton. J Exp Neurosci. 10:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang C, Li C, Xu Z, Zhao S, Li P, Cao J
and Mi W: The effect of surgical and psychological stress on
learning and memory function in aged C57BL/6 mice. Neuroscience.
320:210–220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, He H, Li D, Zhu W, Duan K, Le Y,
Liao Y and Ou Y: The role of the TLR4 signaling pathway in
cognitive deficits following surgery in aged rats. Mol Med Rep.
7:1137–1142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu N, Wang C, Zheng Y, Ao J, Zhang C, Xie
K, Li Y, Wang H, Yu Y and Wang G: The role of the
Wnt/β-catenin-Annexin A1 pathway in the process of
sevoflurane-induced cognitive dysfunction. J Neurochem.
137:240–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Small DH, Mok SS and Bornstein JC:
Alzheimer's disease and Abeta toxicity: From top to bottom. Nat Rev
Neurosci. 2:595–598. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu N, Wang M, Xie K, Wang H, Wang C, Wang
C, Wang C, Li Y, Yu Y and Wang G: Internalization of GluA2 and the
underlying mechanisms of cognitive decline in aged rats following
surgery and prolonged exposure to sevoflurane. Neurotoxicology.
49:94–103. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hamilton DL, Findlay JA, Montagut G,
Meakin PJ, Bestow D, Jalicy SM and Ashford ML: Altered amyloid
precursor protein processing regulates glucose uptake and oxidation
in cultured rodent myotubes. Diabetologia. 57:1684–1692. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
He XL, Yan N, Chen XS, Qi YW, Yan Y and
Cai Z: Hydrogen sulfide down-regulates BACE1 and PS1 via activating
PI3K/Akt pathway in the brain of APP/PS1 transgenic mouse.
Pharmacol Rep. 68:975–982. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sarker D, Reid AH, Yap TA and de Bono JS:
Targeting the PI3K/AKT pathway for the treatment of prostate
cancer. Clin Cancer Res. 15:4799–4805. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arslan F, Lai RC, Smeets MB, Akeroyd L,
Choo A, Aguor EN, Timmers L, van Rijen HV, Doevendans PA,
Pasterkamp G, et al: Mesenchymal stem cell-derived exosomes
increase ATP levels, decrease oxidative stress and activate
PI3K/Akt pathway to enhance myocardial viability and prevent
adverse remodeling after myocardial ischemia/reperfusion injury.
Stem Cell Res. 10:301–312. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Deng Y, Wang Z, Wang R, Zhang X, Zhang S,
Wu Y, Staufenbiel M, Cai F and Song W: Amyloid-β protein (Aβ) Glu11
is the major β-secretase site of beta-site amyloid-β precursor
protein-cleaving enzyme 1 (BACE1) and shifting the cleavage site to
Aβ Asp1 contributes to Alzheimer pathogenesis. Eur J Neurosci.
37:1962–1969. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi X, Cai X, Di W, Li J, Xu X, Zhang A,
Qi W, Zhou Z and Fang Y: MFG-E8 selectively inhibited Aβ-induced
microglial M1 polarization via NF-κB and PI3K-Akt pathways. Mol
Neurobiol. 54:7777–7788. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chiang HC, Wang L, Xie Z, Yau A and Zhong
Y: PI3 kinase signaling is involved in Abeta-induced memory loss in
Drosophila. Proc Natl Acad Sci USA. 107:7060–7065. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qiang W, Yau WM, Lu JX, Collinge J and
Tycko R: Structural variation in amyloid-β fibrils from Alzheimer's
disease clinical subtypes. Nature. 541:217–221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Szaruga M, Munteanu B, Lismont S, Veugelen
S, Horrè K, Mercken M, Saido TC, Ryan NS, De Vos T, Savvides SN, et
al: Alzheimer's-causing mutations shift Aβ length by destabilizing
γ-secretase-Aβn interactions. Cell. 170:443–456 e414. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao J, Fu Y, Yasvoina M, Shao P, Hitt B,
O'Connor T, Logan S, Maus E, Citron M, Berry R, et al: Beta-site
amyloid precursor protein cleaving enzyme 1 levels become elevated
in neurons around amyloid plaques: Implications for Alzheimer's
disease pathogenesis. J Neurosci. 27:3639–3649. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Portelius E, Dean RA, Andreasson U,
Mattsson N, Westerlund A, Olsson M, Demattos RB, Racke MM,
Zetterberg H, May PC and Blennow K: β-site amyloid precursor
protein-cleaving enzyme 1(BACE1) inhibitor treatment induces Aβ5-X
peptides through alternative amyloid precursor protein cleavage.
Alzheimers Res Ther. 6:752014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hunt KW, Cook AW, Watts RJ, Clark CT,
Vigers G, Smith D, Metcalf AT, Gunawardana IW, Burkard M, Cox AA,
et al: Spirocyclic β-site amyloid precursor protein cleaving enzyme
1 (BACE1) inhibitors: From hit to lowering of cerebrospinal fluid
(CSF) amyloid β in a higher species. J Med Chem. 56:3379–3403.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mason TJ and Matthews M: Aquatic
environment, housing and management in the eighth edition of the
guide for the care and use of laboratory animals: Additional
considerations and recommendations. J Am Assoc Lab Anim Sci.
51:329–332. 2012.PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative pcr and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tuo YL, Li XM and Luo J: Long noncoding
RNA UCA1 modulates breast cancer cell growth and apoptosis through
decreasing tumor suppressive miR-143. Eur Rev Med Pharmacol Sci.
19:3403–3411. 2015.PubMed/NCBI
|
|
26
|
Holsinger RM, Goense N, Bohorquez J and
Strappe P: Splice variants of the Alzheimer's disease
beta-secretase, BACE1. Neurogenetics. 14:1–9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Johnson JL, Chambers E and Jayasundera K:
Application of a bioinformatics-based approach to identify novel
putative in vivo BACE1 substrates. Biomed Eng Comput Biol. 5:1–15.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kuk EB, Jo AR, Oh SI, Sohn HS, Seong SH,
Roy A, Choi JS and Jung HA: Anti-Alzheimer's disease activity of
compounds from the root bark of Morus alba L. Arch Pharm Res.
40:338–349. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Engelman JA: Targeting PI3K signalling in
cancer: Opportunities, challenges and limitations. Nat Rev Cancer.
9:550–562. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Abliz A, Deng W, Sun R, Guo W, Zhao L and
Wang W: Wortmannin, PI3K/Akt signaling pathway inhibitor,
attenuates thyroid injury associated with severe acute pancreatitis
in rats. Int J Clin Exp Pathol. 8:13821–13833. 2015.PubMed/NCBI
|
|
32
|
Xu P, Wang J, Yang ZW, Lou XL and Chen C:
Regulatory roles of the PI3K/Akt signaling pathway in rats with
severe acute pancreatitis. PLoS One. 8:e817672013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Devi L and Ohno M: Genetic reductions of
beta-site amyloid precursor protein-cleaving enzyme 1 and
amyloid-beta ameliorate impairment of conditioned taste aversion
memory in 5XFAD Alzheimer's disease model mice. Eur J Neurosci.
31:110–118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nishitsuji K, Tomiyama T, Ishibashi K, Ito
K, Teraoka R, Lambert MP, Klein WL and Mori H: The E693Delta
mutation in amyloid precursor protein increases intracellular
accumulation of amyloid beta oligomers and causes endoplasmic
reticulum stress-induced apoptosis in cultured cells. Am J Pathol.
174:957–969. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thinakaran G and Koo EH: Amyloid precursor
protein trafficking, processing, and function. J Biol Chem.
283:29615–29619. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tu S, Bhagat G, Cui G, Takaishi S,
Kurt-Jones EA, Rickman B, Betz KS, Penz-Oesterreicher M, Bjorkdahl
O, Fox JG and Wang TC: Overexpression of interleukin-1beta induces
gastric inflammation and cancer and mobilizes myeloid-derived
suppressor cells in mice. Cancer Cell. 14:408–419. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Smolen JS, Beaulieu A, Rubbert-Roth A,
Ramos-Remus C, Rovensky J, Alecock E, Woodworth T and Alten R;
OPTION Investigators, : Effect of interleukin-6 receptor inhibition
with tocilizumab in patients with rheumatoid arthritis (OPTION
study): A double-blind, placebo-controlled, randomised trial.
Lancet. 371:987–997. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Scheller J, Chalaris A, Schmidt-Arras D
and Rose-John S: The pro- and anti-inflammatory properties of the
cytokine interleukin-6. Biochim Biophys Acta. 1813:878–888. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Miao HH, Zhen Y, Ding GN, Hong FX, Xie ZC
and Tian M: Ginsenoside Rg1 attenuates isoflurane-induced caspase-3
activation via inhibiting mitochondrial dysfunction. Biomed Environ
Sci. 28:116–126. 2015.PubMed/NCBI
|
|
40
|
Cibelli M, Fidalgo AR, Terrando N, Ma D,
Monaco C, Feldmann M, Takata M, Lever IJ, Nanchahal J, Fanselow MS
and Maze M: Role of interleukin-1beta in postoperative cognitive
dysfunction. Ann Neurol. 68:360–368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Luo X, Yang L, Chen X and Li S: Tau
hyperphosphorylation: A downstream effector of isoflurane-induced
neuroinflammation in aged rodents. Med Hypotheses. 82:94–96. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Berger M, Nadler JW, Browndyke J, Terrando
N, Ponnusamy V, Cohen HJ, Whitson HE and Mathew JP: Postoperative
cognitive dysfunction: Minding the gaps in our knowledge of a
common postoperative complication in the elderly. Anesthesiol Clin.
33:517–550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen X, Wang W, Zhang J, Li S, Zhao Y, Tan
L and Luo A: Involvement of caspase-3/PTEN signaling pathway in
isoflurane-induced decrease of self-renewal capacity of hippocampal
neural precursor cells. Brain Res. 1625:275–286. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lin D and Zuo Z: Isoflurane induces
hippocampal cell injury and cognitive impairments in adult rats.
Neuropharmacology. 61:1354–1359. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yang C, Zhu B, Ding J and Wang ZG:
Isoflurane anesthesia aggravates cognitive impairment in
streptozotocin-induced diabetic rats. Int J Clin Exp Med.
7:903–910. 2014.PubMed/NCBI
|