Introduction
The dried root of Astragalus membranaceus
(AM), alternatively termed Astragali Radix or
Huangqi, is a globally popular medicinal herb primarily
grown in Northern China, Mongolia and Korea (1). AM has been used in traditional
Chinese medicine (TCM) for over two millennia, where it is
traditionally believed to ease urination, reduce purulent
discharge, and promote soft tissue repair (1,2). To
this day, the water-soluble extract of AM is still applied for the
alternative treatment of various conditions, including fatigue,
anorexia, anemia, fever, allergies, gastric ulcers and diarrhea
(1). There are >100
biologically-active compounds that have been identified in AM,
including flavonoids, saponins, polysaccharides, amino acids and
trace elements (1,3). It is hypothesized that this
combination of bioactive compounds are responsible for AM's
observed immunomodulatory, anti-hyperglycemic, anti-inflammatory,
antioxidant and antiviral properties (1).
In particular, AM has been indicated to demonstrate
antioxidant effects in diabetic nephropathy, heart disease and
liver disease (1). Several
previous studies have reported that AM root extract demonstrates
antioxidant activity in vitro (4–6).
Although these previous findings reveal AM's beneficial antioxidant
properties, its cytoprotective effects at the cellular and
mitochondrial levels remain largely unknown.
To address this question, in vitro
administration of the bacterial endotoxin lipopolysaccharide (LPS)
is a well-established experimental model for generating
intracellular oxidative stress (7,8). In
the present study, the authors applied LPS to a human pulmonary
type II-like epithelial lung adenocarcinoma cell line, a human
umbilical vein endothelial cell line and a human bladder carcinoma
cell line in order to construct in vitro models of
intracellular oxidative stress. They then assayed the cellular and
mitochondrial cytoprotective effects of varying doses of AM root
extract upon these three LPS-treated cell lines. These findings
should better the understanding of AM's cytoprotective effects upon
human cells.
Materials and methods
Cell culture and experimental group
construction
The root extract of Astragalus membranaceus
(AM) and lipopolysaccharide (LPS; 1 µg/ml) were purchased from
Sigma-Aldrich (Merck KGaA; Darmstadt, Germany). The human pulmonary
type II-like epithelial lung adenocarcinoma cell line A549, the
human umbilical vein endothelial cell (HUVEC) line CRL-1730, and
the ECV304 cell line were purchased from American Type Culture
Collection (Manassas, VA, USA). Notably, the ECV304 cell line was
originally thought to be derived from HUVECs from a healthy donor,
however is now known to be cross-contaminated with the T24 bladder
carcinoma cell line (9). Trypsin,
propidium iodide (PI), and RNase A were purchased from
Sigma-Aldrich. Fetal calf serum (FCS) was purchased from Hyclone;
GE Healthcare Life Sciences (Logan, UT, USA), and Dulbecco's
modified Eagles medium (DMEM) was purchased from Gibco; Thermo
Fisher Scientific, (Waltham, MA, USA).
The three logarithmic-phase cell lines were seeded
onto six-well plates at a density of 1×106 cells/ml and
cultured in DMEM supplemented with 5% FCS at 37°C to the point of
adherence. Then, adherent cells were divided into six experimental
groups: LPS group treated with LPS (1 µg/ml) alone, four LPS+AM
groups treated with various concentrations of AM (25, 50, 100, and
200 µg/ml) in addition to LPS (1 µg/ml), and a control group
receiving neither LPS nor AM. The three cell lines were cultured
under these conditions at 37°C for 24 h prior to performance of the
following assays.
Cell apoptosis assay
As previously described with minor modifications
(10), flow cytometric analysis
was used to differentiate early and late apoptotic cells using an
Annexin V-FITC/PI apoptosis detection kit (Nanjing KGI Biological
Technology Development Co., Ltd., Nanjing, China) according to the
manufacturer's instructions. Briefly, cells (1×106
cells/ml) were trypsinized, and the cell suspension was transferred
into a centrifuge tube for centrifugation (200 × g, 30 min, 4°C).
The supernatant was aspirated out, and the cells were washed three
times with phosphate-buffered saline (PBS). A total of 100,000
cells were resuspended in 100 µl binding buffer containing Annexin
V-FITC and PI. Samples were incubated for 5 min at room temperature
in the dark. Quantification of Annexin V-FITC and PI binding was
performed using a BD-FACS Canto™ II flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA). BD CellQuest™ Pro version 5.2.1 (BD
Biosciences) was then used to perform the quadrant analysis.
Experiments were repeated three times.
Cell cycle progression assay
Cell cycle progression was determined by flow
cytometry following PI staining, as previously described with minor
modifications (11). Cells were
trypsinized, and the cell suspension was transferred into a
centrifuge tube for centrifugation (1,000 × g, 5 min, 37°C). The
supernatant was aspirated out, the cells were washed three times
with PBS. Cells were fixed in 70% ethanol at 4°C for 24 h. The
cells were then stained with a PI/RNase staining buffer for 1 h at
37°C. Stained cells were analyzed on a BD-FACS Canto™ II flow
cytometer (BD Biosciences) to calculate the proportion of cells in
the various phases of the cell cycle using Mod Fit LT version 3.0
(Verity Software House Inc., Topsham, ME, USA). Experiments were
repeated three times.
Reactive oxygen species (ROS)
detection assay
The 2′,7′-dichlorofluorescein-diacetate (DCFH-DA)
kit was used to quantitate ROS generation as an index for
cytotoxicity, and was purchased from Cell Biolabs, Inc. (San Diego,
CA, USA). ROS generation was analyzed with a DCFH-DA assay as
previously described, with minor modifications (12). The culture medium was aspirated
out, cells were washed three times in PBS, and then incubated in
ROS staining solution (DCFH-DA) at 37°C for 20 min. Following
washing three times with PBS, DCFH fluorescence intensity was
detected with a BD-FACS Canto™ II flow cytometer (BD Biosciences)
at a 488 nm excitation wavelength and a 525 nm emission wavelength
(channel FL1). Results were calculated relative to the mean
fluorescence intensity of channel FL1 in the untreated control
group. Experiments were repeated three times.
Mitochondrial membrane potential
(Δψmit) assay
Rhodamine 123 is a mitochondrial-specific
fluorescent probe that stains in a membrane potential-dependent
fashion. This was used in the present study and was. The
mitochondrial membrane potential (Δψmit) was analyzed
with a rhodamine 123 assay purchased from Thermo Fisher Scientific,
Inc, as previously described with minor modifications (13). Cells were trypsinized, and the cell
suspension was transferred into a centrifuge tube for
centrifugation (400 × g, 15 min, 37°C). The supernatant was
aspirated, washed three times with PBS, and incubated with 5 µM
rhodamine 123 at 37°C for 30 min in the dark. Rhodamine 123
fluorescence intensity was then detected with a BD-FACS Canto™ II
flow cytometer (Becton-Dickinson) and CellQuest™ Pro version 5.2.1
(BD Biosciences). Experiments were repeated three times.
Western blotting
Western blotting was performed as previously
described, with minor modifications (14). Cells were lysed for 30 min in
MBST/OG buffer (25 mM MES; 150 mM NaCl; 60 mM octylglucopyranoside;
1% Triton X-100; pH 6.4) on ice. The lysate was boiled for 10 min
to denature the proteins and then cooled. The protein concentration
was determined using the Bradford assay, and equal amounts of
lysate protein (50 µg/lane) were then resolved by 10% SDS-PAGE.
Following electrophoresis, the proteins were transferred to a
polyvinylidene difluoride membrane, which was blocked for 1 h in 5%
skim milk solution in Tris-buffered saline containing 0.1% Tween-20
at room temperature. The membrane was incubated overnight at 4°C
with primary antibodies (Abcam, Cambridge, UK) against human
cleaved caspase-3 (1:100; cat. no. ab2302), human p53 (1:500; cat.
no. ab26), human B cell lymphoma (Bcl)-2 (1:1,000; cat. no.
ab32124), and human β-actin (1:500; cat. no. ab8226). The membrane
was subsequently incubated for 1 h at room temperature with
horseradish peroxidase-conjugated secondary antibodies (1:5,000;
anti-rabbit, cat. no. ab7090; anti-mouse, cat. no. ab97040; Abcam).
The immunoreactive bands were visualized using an enhanced
chemiluminescence detection kit (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Band intensity was quantified using ChemiDoc
XRS (Bio-Rad, Laboratories, Inc.) and normalized to β-actin.
Experiments were repeated three times.
Statistical analysis
Statistical analysis was performed using SPSS
software, version 17.0 (SPSS, Inc., Chicago, IL, USA). Data are
presented as the mean ± standard deviation from at least three
independent experiments. Statistically significant differences
between experimental groups were assessed by one-way analysis of
variance followed by Bonferroni's multiple comparison test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
AM therapy reduces apoptosis rate
The present study first assayed apoptosis rates
under varying doses of AM root extract in the three LPS-treated
human cell lines. Across all three cell lines, LPS administration
significantly increased apoptosis rates compared with controls
(P<0.05; Fig. 1). The addition
of either 100 µg/ml or 200 µg/ml AM therapy significantly reduced
apoptosis rates in LPS-treated cells (P<0.05; Fig. 1). As a general trend, increasing
concentrations of AM therapy produced progressively greater
reductions in the apoptosis rate.
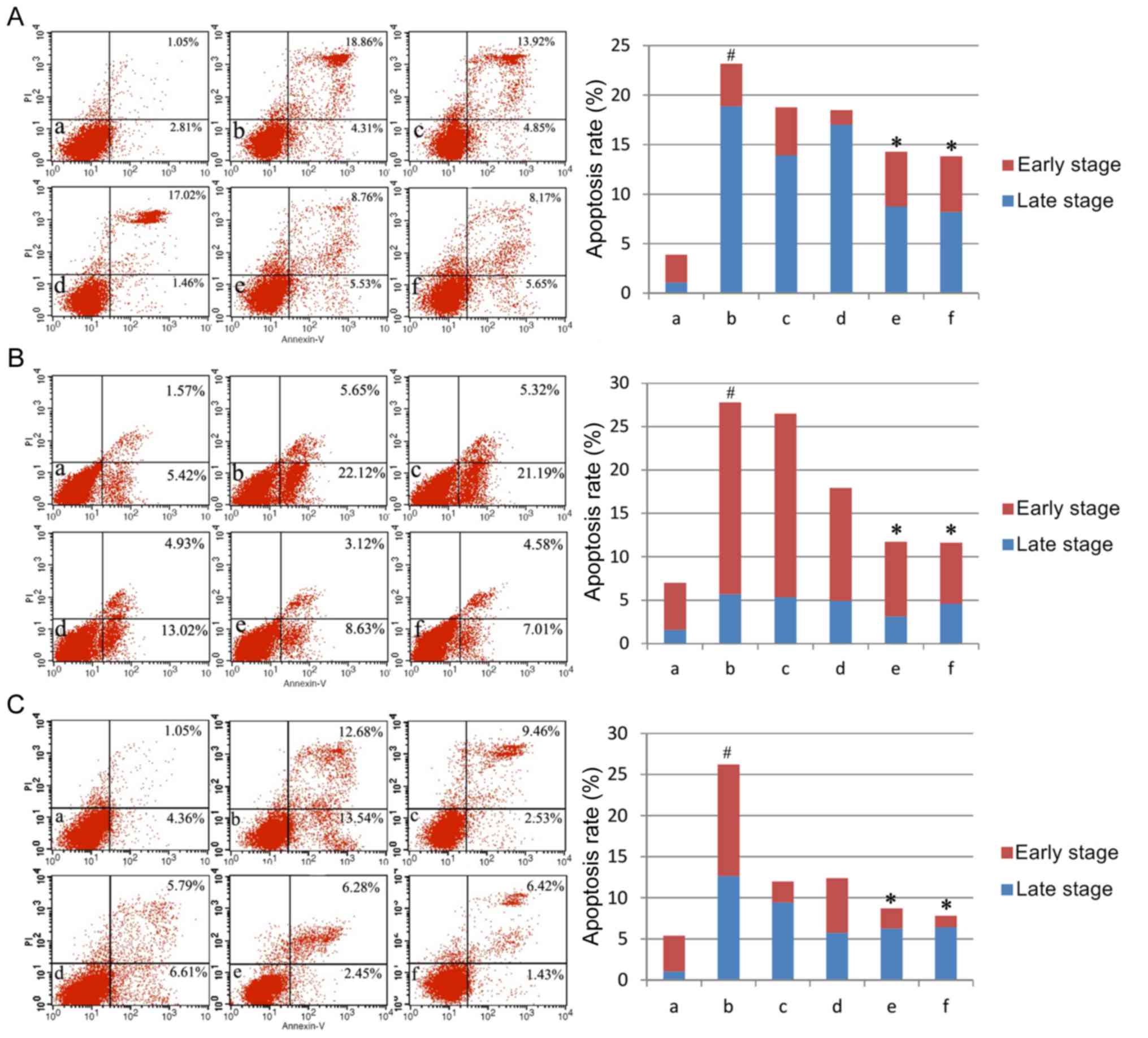 | Figure 1.Apoptosis rates in the three cell
lines. Apoptosis rates of (A) the human pulmonary type II-like
epithelial lung adenocarcinoma cell line A549, (B) the human
umbilical vein endothelial cell line CRL-1730, and (C) the human
bladder carcinoma cell line ECV304. A, Untreated control group (no
LPS, no AM); b, LPS group (1 µg/ml), and the four LPS+AM groups
treated with LPS (1 µg/ml); plus c, 25; d, 50; e, 100; and f, 200
µg/ml of AM. Data are presented as the mean ± standard deviations
from three independent experiments. #P<0.05 vs. control group a,
and *P<0.05 vs. LPS group b, with one-way analysis of variance
followed by Bonferroni post-hoc test. AM, Astragalus membranaceus;
LPS, lipopolysaccharide; PI, propidium iodide. |
AM therapy affects cell cycle
progression
Having established that AM therapy significantly
reduces apoptosis rate in LPS-treated cells, the present study
assayed cell cycle phase distributions under varying doses of AM
root extract in the three LPS-treated human cell lines. Across all
three cell lines, LPS administration produced a significant shift
to S/G2 phase relative to controls (P<0.05; Fig. 2). In A549 cells, the addition of
either 25, 100 or 200 µg/ml AM therapy significantly reduced the
S/G2 phase % in LPS-treated cells (P<0.05; Fig. 2A). In CRL-1730 cells, the addition
of all AM therapy doses significantly reduced the S/G2 phase % in
LPS-treated cells (P<0.05; Fig.
2B). In ECV304 cells, the addition of 200 µg/ml AM therapy
significantly reduced the S/G2 phase % in LPS-treated cells
(P<0.05; Fig. 2C). As a general
trend, increasing concentrations of AM therapy produced
progressively greater reductions in S/G2 phase %.
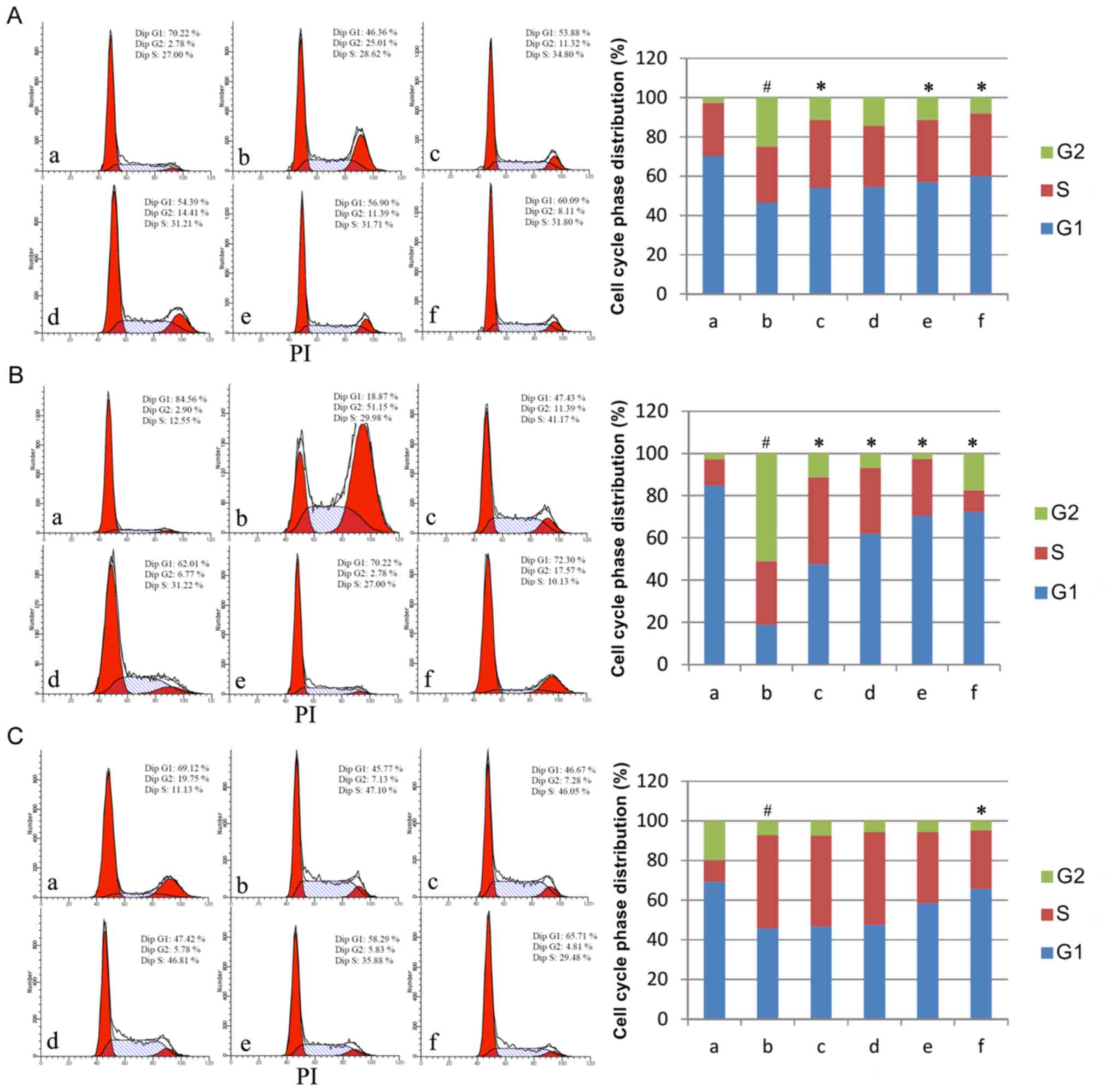 | Figure 2.Cell cycle phase distributions in the
three cell lines. Cell cycle phase distributions of (A) the human
pulmonary type II-like epithelial lung adenocarcinoma cell line
A549, (B) the human umbilical vein endothelial cell line CRL-1730,
and (C) the human bladder carcinoma cell line ECV304. A, Untreated
control group (no LPS, no AM); b, LPS group (1 µg/ml), and the four
LPS+AM groups treated with LPS (1 µg/ml); plus c, 25; d, 50; e,
100; and f, 200 µg/ml of AM. Data are presented as the mean ±
standard deviations from three independent experiments. #P<0.05
vs. control group a, and *P<0.05 vs. LPS group b, with one-way
analysis of variance followed by Bonferroni post-hoc test. AM,
Astragalus membranaceus; LPS, lipopolysaccharide. |
AM therapy reduces ROS production
The present study next assayed ROS production under
varying doses of AM root extract in the three LPS-treated human
cell lines. Across all three cell lines, LPS administration
significantly increased ROS production relative to controls
(P<0.05; Fig. 3A). Across all
three cell lines, the addition of either 100 µg/ml or 200 µg/ml AM
therapy significantly reduced ROS production in LPS-treated cells
(P<0.05; Fig. 3A). As a general
trend, increasing concentrations of AM therapy produced
progressively greater reductions in ROS production.
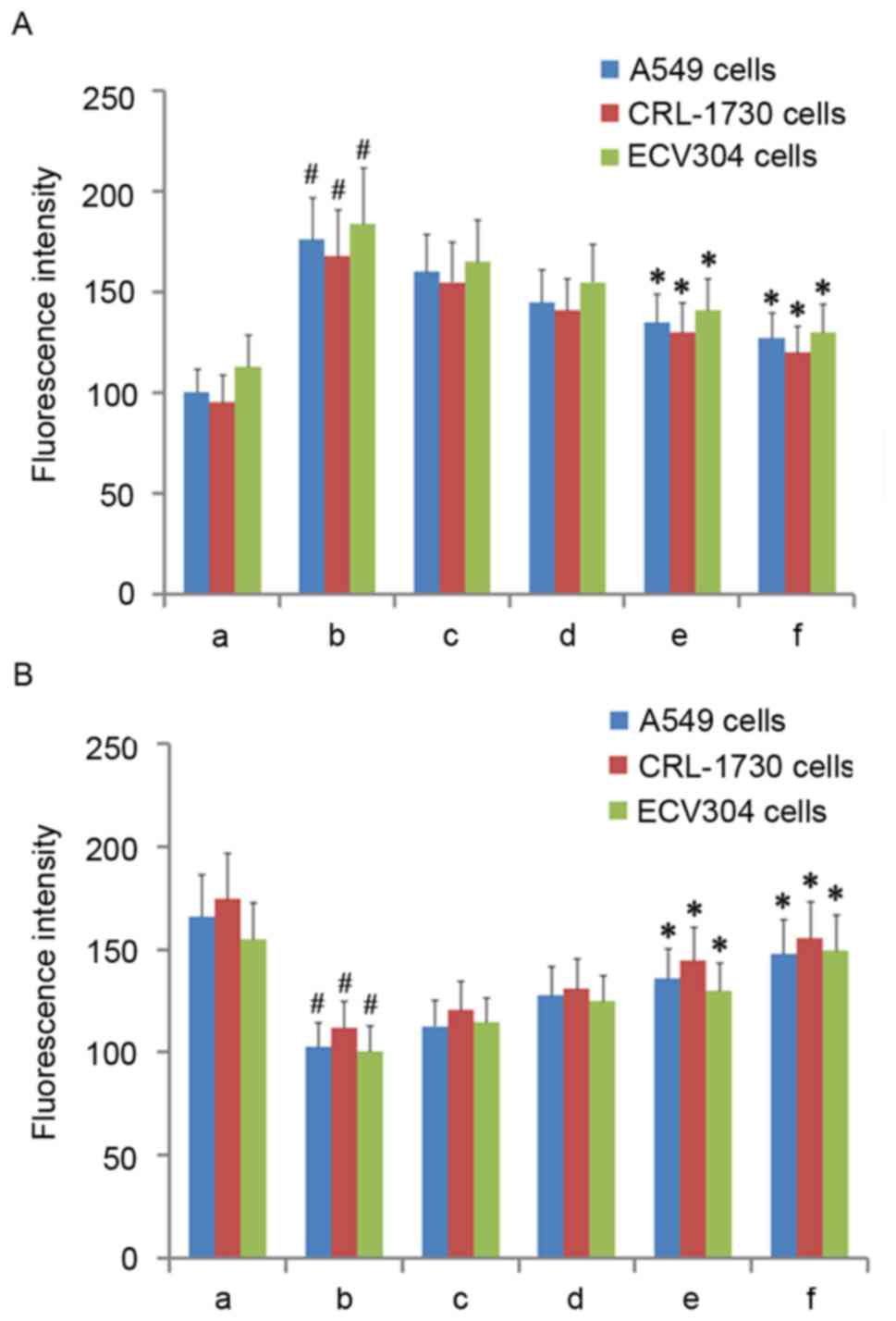 | Figure 3.ROS and mitochondrial membrane
potential in the three cell lines. (A) Fluorescence intensity of
ROS in the three cell lines. (B) Fluorescence intensity of
rhodamine 123 in the three cell lines. A, Untreated control group
(no LPS, no AM); b, LPS group (1 µg/ml), and the four LPS+AM groups
treated with LPS (1 µg/ml); plus c, 25; d, 50; e, 100; and f, 200
µg/ml of AM. Data are presented as the mean ± standard deviations
from three independent experiments. #P<0.05 vs. control group a,
and *P<0.05 vs. LPS group b, with one-way analysis of variance
followed by Bonferroni post-hoc test. AM, Astragalus membranaceus;
LPS, lipopolysaccharide; ROS, reactive oxygen species. |
AM therapy increases Δψmit
levels
Having established that AM therapy significantly
reduces ROS production in LPS-treated cells, Δψmit
levels were then assayed under varying doses of AM root extract in
the three LPS-treated human cell lines. As diffusion of the
cationic dye rhodamine 123 is directly proportional to the degree
of membrane polarization, Δψmit levels were examined by
measuring the relative differences in rhodamine 123 fluorescence
intensity between the experimental groups (15). Across all three cell lines, LPS
administration significantly reduced Δψmit relative to
controls (P<0.05; Fig. 3B).
Across all three cells lines, the addition of either 100 µg/ml or
200 µg/ml AM therapy significantly increased Δψmit in
LPS-treated cells (P<0.05; Fig.
3B). As a general trend, increasing concentrations of AM
therapy produced progressively greater increases in
Δψmit.
AM therapy affects expression of
apoptosis-associated proteins
Having established that AM therapy significantly
increases Δψmit levels in LPS-treated cells, the
expression of several apoptosis-associated proteins (cleaved
caspase-3, p53, and Bcl-2) were assayed under varying doses of AM
root extract in the three LPS-treated human cell lines. In all
three cell lines, LPS administration significantly increased
cleaved caspase-3 and p53 expression and significantly decreased
Bcl-2 expression, relative to controls (P<0.05; Fig. 4). In A549 cells, the addition of
all AM therapy doses significantly reduced cleaved caspase-3 and
p53 expression (P<0.05; Fig.
4A); however, only 200 µg/ml AM therapy significantly increased
Bcl-2 expression in LPS-treated A549 cells (P<0.05; Fig. 4A). In CRL-1730 cells, the addition
of all AM therapy doses significantly reduced cleaved caspase-3 and
p53 expression (P<0.05; Fig.
4B); however, only the addition of either 100 µg/ml or 200
µg/ml AM therapy significantly increased Bcl-2 expression in
LPS-treated CRL-1730 cells (P<0.05; Fig. 4B). In ECV304 cells, the addition of
either 100 µg/ml or 200 µg/ml AM therapy significantly reduced
cleaved caspase-3 and p53 expression (P<0.05; Fig. 4C); however, only the addition of
200 µg/ml AM therapy significantly increased Bcl-2 expression in
LPS-treated ECV304 cells (P<0.05; Fig. 4C). Interestingly, the addition of
25 µg/ml AM therapy increased p53 expression, and 50 µg/ml AM
therapy increased cleaved caspase-3 and p53 expression; however,
these alterations were not significant (P>0.05; Fig. 4C). As a general trend, increasing
concentrations of AM therapy produced progressively reduced cleaved
caspase-3 and p53 expression, and increased Bcl-2 expression
levels.
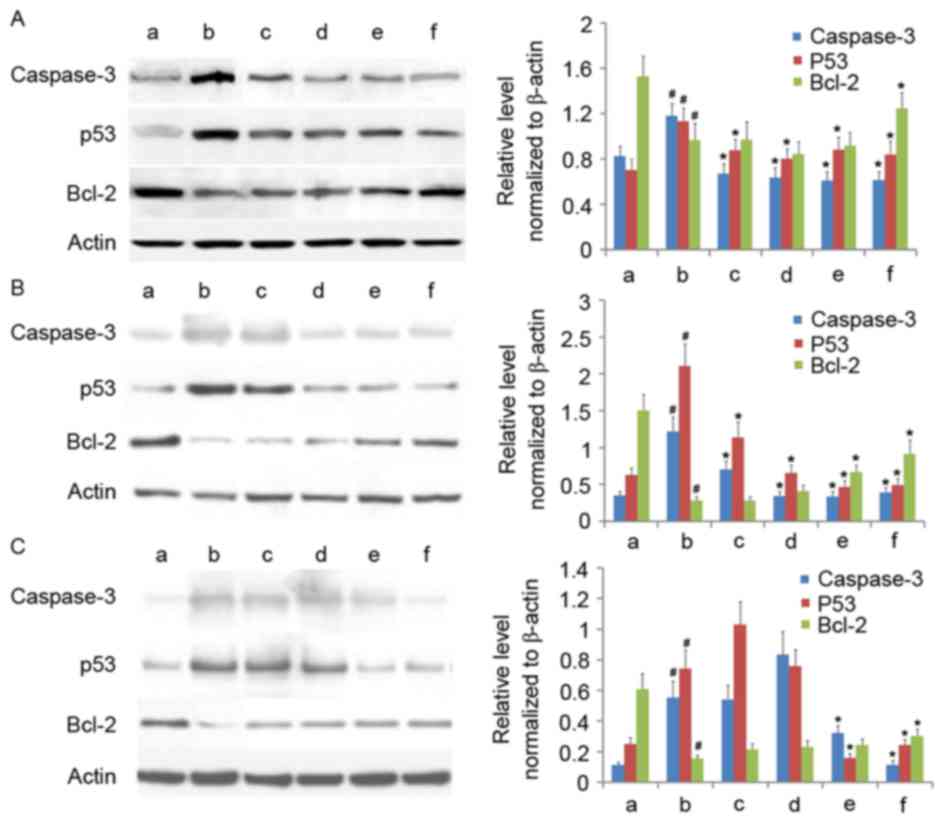 | Figure 4.Western blotting of cleaved caspase-3,
p53 and Bcl-2 in the three cell lines. Western blotting of (A) the
human pulmonary type II-like epithelial lung adenocarcinoma cell
line A549, (B) the human umbilical vein endothelial cell line
CRL-1730, and (C) the human bladder carcinoma cell line ECV304. A,
Untreated control group (no LPS, no AM); b, LPS group (1 µg/ml),
and the four LPS+AM groups treated with LPS (1 µg/ml); plus c, 25;
d, 50; e, 100 and f, 200 µg/ml of AM. β-actin served as a loading
control. Data are presented as the mean ± standard deviations from
three independent experiments. #P<0.05 vs. control group a and
*P<0.05 vs. LPS group b with one-way analysis of variance
followed by Bonferroni post-hoc test. AM, Astragalus membranaceus;
LPS, lipopolysaccharide; Bcl-2, B cell lymphoma-2. |
Discussion
Oxidative stress is defined as the excess production
of ROS (and/or other oxidant species) that overwhelms the cell's
antioxidant capacity, with mitochondrial electron transport
recognized as an important source of ROS in the majority of
eukaryotic cell types (16,17).
Oxidative stress induces macromolecular damage (oxidative damage of
DNA bases, polyunsaturated fatty acids, and amino acid residues)
and is involved in the pathogenesis of several disorders, including
inflammatory diseases, atherosclerosis, diabetes, and cancer
(16–18). Although several synthetic compounds
have been developed and marketed to address oxidative stress
(butylated hydroxytoluene, butylated hydroxyanisole, and
nordihydroguaiaretic acid), these synthetic compounds may produce
adverse side effects (18,19). As a result, there is increasing
interest in the discovery and investigation of non-synthetic,
naturally-occurring therapies to address oxidative stress.
The TCM herb Astragalus membranaceus (AM,
Astragali Radix, or Huangqi) has been demonstrated to
exhibit antioxidant effects in vitro (4–6).
Previous research on eight TCM herb pairs demonstrated that
antioxidant activity is largely driven by alterations in total
flavonoid content (20). In
accordance with this, antioxidant assays on five AM extracts (ABTS,
DPPH, reducing power, and •OH assays) demonstrated that AM's
phenolic compounds (including flavonoids) are primarily responsible
for its antioxidant effects (4).
However, there is little available information on the cellular and
mitochondrial cytoprotective effects of AM root extract in human
cells.
Therefore, the present study examined the cellular
and mitochondrial cytoprotective effects of varying doses of AM
root extract applied to three LPS-treated human cell lines. Across
all three cell lines, it was demonstrated that AM treatment reduced
the negative cellular and mitochondrial effects induced by LPS in a
dose-dependent manner, through reduced apoptosis rates, reduced
S/G2 phase arrest, reduced ROS production, increased
Δψmit, reduced cleaved caspase-3, reduced p53 expression
and increased Bcl-2 expression. As increased ROS production and
loss of Δψmit have been associated with the activation
of caspase-induced apoptotic cascades (15,21),
these results suggested that AM therapy reduced apoptosis through
dampening ROS production and dampening Δψmit loss, which
downregulated the p53-induced caspase-mediated pro-apoptotic
pathway and upregulated the nuclear factor-κB-induced
Bcl-2-mediated anti-apoptotic pathway. Accordingly, this precise
pattern of anti-apoptotic findings has been previously observed
with other naturally-occurring therapies in vitro, including
isofraxidin, quercetin, cyanidin, arjunolic acid, and taurine
(21–25).
There are several limitations to the present study.
Firstly, the present study only analyzed three human cell lines,
therefore results are restricted to the three cell lines and AM
therapy may have different effects in other cell types. Secondly,
the in vitro LPS model used here may not precisely match the
oxidative stress conditions for the three cell types in
vivo. Finally, an AM root extract solution that contains
numerous bioactive compounds was applied; therefore, the present
study did not establish which compound(s) in the AM root extract
were responsible for the observed effects. Future studies on AM
root extract should apply high performance liquid chromatography
and mass spectrometry analytical techniques in order to identify
the constituent bioactive compounds responsible for AM's beneficial
effects (26). In addition,
although the results suggest it, the present study did not
conclusively establish that AM therapy reduces apoptosis through
downregulating the p53-induced caspase-mediated pro-apoptotic
pathway and/or upregulating the NF-κB-induced Bcl-2-mediated
anti-apoptotic pathway. Therefore, further experiments involving
gene silencing and overexpression are required to establish the
precise molecular mechanism(s) underlying AM's beneficial
effects.
In conclusion, AM treatment protects human pulmonary
and bladder epithelial cells as well as human endothelial cells
from LPS-induced apoptosis in a dose-dependent manner. The present
evidence suggests that AM therapy reduces ROS production and
mitochondrial membrane depolarization, thereby preventing the
activation of caspase-induced apoptotic cascades. Further research
on the molecular effects of AM's constituent compounds are needed
to better understand the molecular mechanism(s) underlying AM's
beneficial effects.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Heilongjiang
Province Research Program (grant no. D201179). The funders had no
role in study design, data collection and analysis, decision to
publish, or preparation of the manuscript.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Author's contributions
XW and YL conceived of and designed the study. XW,
WZ, QW and PC performed the experiments. XW, WZ, QW and PC analyzed
and interpreted the data. XW wrote the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have competing
interests.
References
|
1
|
Fu J, Wang Z, Huang L, Zheng S, Wang D,
Chen S, Zhang H and Yang S: Review of the botanical
characteristics, phytochemistry, and pharmacology of Astragalus
membranaceus (Huangqi). Phytother Res. 28:1275–1283. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang HW, Lin ZX, Xu C, Leung C and Chan
LS: Astragalus (a traditional Chinese medicine) for treating
chronic kidney disease. Cochrane Database Syst Rev.
22:CD0083692014.
|
|
3
|
Ma XQ, Shi Q, Duan JA, Dong TT and Tsim
KW: Chemical analysis of Radix Astragali (Huangqi) in China: A
comparison with its adulterants and seasonal variations. J Agric
Food Chem. 50:4861–4866. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li X, Chen D, Mai Y, Wen B and Wang X:
Concordance between antioxidant activities in vitro and chemical
components of Radix Astragali (Huangqi). Nat Prod Res.
26:1050–1053. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu HW, Fang J, Tang LY, Lu P, Xu HY, Zhao
Y, Li DF, Fu MH and Yang HJ: Quality evaluation of astragali radix
based on DPPH radical scavenging activity and chemical analysis.
Chin Herbal Med. 6:282–289. 2014. View Article : Google Scholar
|
|
6
|
Jiao J, Gai QY, Fu YJ, Ma W, Peng X, Tan
SN and Efferth T: Efficient production of isoflavonoids by
Astragalus membranaceus hairy root cultures and evaluation of
antioxidant activities of extracts. J Agric Food Chem.
62:12649–12658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang Y, Wang GZ, Rabinovitch PS and Tabas
I: Macrophage mitochondrial oxidative stress promotes
atherosclerosis and nuclear factor-κB-mediated inflammation in
macrophages. Circ Res. 114:421–433. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Müller JM, Ziegler-Heitbrock HL and
Baeuerle PA: Nuclear factor kappa B, a mediator of
lipopolysaccharide effects. Immunobiology. 187:233–256. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
MacLeod RA, Dirks WG, Matsuo Y, Kaufmann
M, Milch H and Drexler HG: Widespread intraspecies
cross-contamination of human tumor cell lines arising at source.
Int J Cancer. 83:555–563. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Scharstuhl A, Mutsaers HA, Pennings SW,
Russel FG and Wagener FA: Involvement of VDAC, Bax and ceramides in
the efflux of AIF from mitochondria during curcumin-induced
apoptosis. PLoS One. 4:e66882009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Arora S, Bhardwaj A, Srivastava SK, Singh
S, McClellan S, Wang B and Singh AP: Honokiol arrests cell cycle,
induces apoptosis, and potentiates the cytotoxic effect of
gemcitabine in human pancreatic cancer cells. PLoS One.
6:e215732011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang DY, Wang HJ and Tan YZ:
Wnt/β-catenin signaling induces the aging of mesenchymal stem cells
through the DNA damage response and the p53/p21 pathway. PLoS One.
6:e213972011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang S, Zhang Y, Zhuang Y, Wang J, Ye J,
Zhang S, Wu J, Yu K and Han Y: Matrine induces apoptosis in human
acute myeloid leukemia cells via the mitochondrial pathway and Akt
inactivation. PLoS One. 7:e468532012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Scotland RL, Allen L, Hennings LJ, Post GR
and Post SR: The ral exchange factor rgl2 promotes cardiomyocyte
survival and inhibits cardiac fibrosis. PLoS One. 8:e735992013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cao J, Liu Y, Jia L, Zhou HM, Kong Y, Yang
G, Jiang LP, Li QJ and Zhong LF: Curcumin induces apoptosis through
mitochondrial hyperpolarization and mtDNA damage in human hepatoma
G2 cells. Free Radic Biol Med. 43:968–975. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ray PD, Huang BW and Tsuji Y: Reactive
oxygen species (ROS) homeostasis and redox regulation in cellular
signaling. Cell Signal. 24:981–990. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hekimi S, Lapointe J and Wen Y: Taking a
‘good’ look at free radicals in the aging process. Trends Cell
Biol. 21:569–576. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jin M, Zhao K, Huang Q and Shang P:
Structural features and biological activities of the
polysaccharides from Astragalus membranaceus. Int J Biol Macromol.
64:257–266. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carocho M and Ferreira IC: A review on
antioxidants, prooxidants and related controversy: Natural and
synthetic compounds, screening and analysis methodologies and
future perspectives. Food Chem Toxicol. 51:15–25. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang WJ, Li DP, Li JK, Li MH, Chen YL and
Zhang PZ: Synergistic antioxidant activities of eight traditional
Chinese herb pairs. Biol Pharm Bull. 32:1021–1026. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li P, Zhao QL, Wu LH, Jawaid P, Jiao YF,
Kadowaki M and Kondo T: Isofraxidin, a potent reactive oxygen
species (ROS) scavenger, protects human leukemia cells from
radiation-induced apoptosis via ROS/mitochondria pathway in
p53-independent manner. Apoptosis. 19:1043–1053. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sharma DR, Sunkaria A, Verma D and Gill
KD: Quercetin attenuates aluminum-induced apoptosis in rat
hippocampus, by preventing cytochrome c translocation, Bcl-2
decrease, bax elevation, caspase-3 and p53 activation. Free Radical
Biol Med. 53:S44–S45. 2012. View Article : Google Scholar
|
|
23
|
Gao S, Chen T, Choi MY, Liang Y, Xue J and
Wong YS: Cyanidin reverses cisplatin-induced apoptosis in HK-2
proximal tubular cells through inhibition of ROS-mediated DNA
damage and modulation of the ERK and AKT pathways. Cancer Lett.
333:36–46. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ghosh J, Das J, Manna P and Sil PC: The
protective role of arjunolic acid against doxorubicin induced
intracellular ROS dependent JNK-p38 and p53-mediated cardiac
apoptosis. Biomaterials. 32:4857–4866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chang CY, Shen CY, Kang CK, Sher YP, Sheu
WH, Chang CC and Lee TH: Taurine protects HK-2 cells from oxidized
LDL-induced cytotoxicity via the ROS-mediated mitochondrial and
p53-related apoptotic pathways. Toxicol Appl Pharmacol.
279:351–363. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu X, Li F, Zhang X, Li P, Zhang X, Wu Z
and Li D: In vitro synergistic antioxidant activity and
identification of antioxidant components from Astragalus
membranaceus and Paeonia lactiflora. PLoS One. 9:e967802014.
View Article : Google Scholar : PubMed/NCBI
|














