Introduction
Lung cancer is one of the most common and aggressive
types of cancer, and the leading cause of cancer-associate
mortality worldwide (1). Among all
patients with lung cancer, non-small-cell lung cancer (NSCLC)
accounts for 75–80% of all cases (2). Surgery and radiotherapy remain the
predominant therapeutic approaches for treating NSCLC (3); however, the majority of patients with
NSCLC are diagnosed at advanced stages accompanied with metastasis,
which leads to highly unsatisfactory outcomes of therapy (4). Thus, to develop novel strategies for
patients with NSCLC, it is necessary to investigate the mechanism
underlying the development and progression of NSCLC.
LIM domain containing 2 (LIMD2) belongs to the LIMD
protein family (5). In the human
genome, 135 identifiable LIM-encoding sequences have been located
within 58 genes (5). LIMD2 has
been demonstrated to serve as a biomarker for papillary thyroid
carcinoma lymph node metastasis (6); however, knowledge regarding role of
LIMD2 in other types of cancer and its functional mechanism remain
very limited. Thus, investigation is required to determine whether
LIMD2 regulates the aggressive behaviors associated with NSCLC,
including proliferation and invasion.
MicroRNAs (miRNAs) are a class of noncoding RNAs
with a length of ~22 nucleotides (7). MiRNAs can regulate gene expression by
binding to the complementary sequence of 3′-untranslated region
(UTR) of target mRNAs (8).
Accumulating evidence has indicated that miRNAs exert notably
important functions in various biological processes, including cell
survival, proliferation, migration and invasion (9,10).
For instance, Xie et al (11) reported that Let-7c inhibits the
growth of cholangiocarcinoma. Wang et al (12) revealed that miR-598 inhibits the
cell proliferation and invasion of glioblastoma by directly
targeting MACC1 (12). At present,
the importance of miRNAs in a variety of cancers is widely
acknowledged (11,12).
In the present study, the function of LIMD2 and its
expression mechanism in NSCLC was investigated. The expression of
LIMD2 was significantly upregulated in NSCLC tissues and cell
lines; knockdown of LIMD2 suppressed the proliferation and invasion
of NSCLC cells as reported by the present study. Additionally,
LIMD2 expression was regulated by miR-34a. Collectively, the
results of the present study demonstrated the important roles of
LIMD2 in the progression of NSCLC.
Materials and methods
Patient samples
NSCLC tissues and adjacent non-cancerous tissues
were collected from 42 patients at The Second Affiliated Hospital
of Harbin Medical University (Harbin, China) between May 2014 and
September 2016. Tissue samples were frozen and stored at −80°C. The
samples were divided into LIMD2 low expression group and LIMD2 high
expression group (the median value of LIMD2 expression was used as
the cutoff value). The associations between the expression of LIMD2
and the clinicopathological features of patients with
non-small-cell lung cancer is given in Table I. The present study was approved by
the Research Ethics Committee of the Second Affiliated Hospital of
Harbin Medical University. Informed consent was obtained from
patients for the collection of samples in accordance with the
Declaration of Helsinki. All patients had not undergone radical
prostatectomy and/or any other treatment prior to the present
study.
 | Table I.Association between the expression of
LIMD2 with the clinicopathological features of patients with
non-small-cell lung cancer. |
Table I.
Association between the expression of
LIMD2 with the clinicopathological features of patients with
non-small-cell lung cancer.
| Features | Low LIMD2 expression
(n=19) | High LIMD2 expression
(n=23) | P-value |
|---|
| Age (years) |
|
| 0.366 |
| ≤50 | 12 | 11 |
|
|
>50 | 7 | 12 |
|
| Sex |
|
| 0.750 |
| Male | 13 | 14 |
|
|
Female | 6 | 9 |
|
| TNM stage |
|
| 0.016a |
| I–II | 14 | 8 |
|
|
III–IV | 5 | 15 |
|
| Tumor size (cm) |
|
| 0.026a |
| ≤4 | 11 | 5 |
|
|
>4 | 8 | 18 |
|
| Lymph node
metastasis |
|
| 0.049a |
|
Negative | 9 | 4 |
|
|
Positive | 10 | 19 |
|
Cell culture and transfection
Human NSCLC cell lines H1299, A549, H358 and H460
were purchased from the American Type Culture Collection (Manassas,
VA, USA). Human bronchial epithelial cells (HBEC) were obtained
from the Chinese Academy of Sciences (Shanghai, China). Cells were
cultured in RPMI-1640 medium (Invitrogen; Thermo Fischer
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Invitrogen; Thermo Fischer Scientific, Inc.) at 37°C
in a humidified 5% CO2 incubator.
Small interfering (si)RNA against LIMD2
(5′-GACCCACCAACTTACCATA-3′) or negative control (si-NC;
5′-AATTCTCCGAACGTGTCAC-3′) and miR-34a
(5′-UGGCAGUGUCUUAGCUGGUUGU-3′) mimic or negative control (NC mimic;
5′-ACAUCUGCGUAAGAUUCGAGUCUA-3′) were synthetized by Invitrogen
(Thermo Fischer Scientific, Inc.) and 50 nM per plasmid was
transfected into H1299 and A549 cells, in which LIMD2 expression
levels were higher compared with the other cell lines, using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The LIMD2 coding sequence was cloned into the
pcDNA3.1 vector (Invitrogen) between Not I and EcoR I, and 1 mg
plasmid was transfected into H1299 and A549 cells as well as 50 nM
miR-34a or NC mimics using Lipofectamine® 3000. Then, 48
h after transfection, the transfection efficiency was determined
using reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) as described below.
Cell proliferation
Each well of a 96-well plate was seeded with ~5,000
transfected cells. Cell proliferation was assessed using a Cell
Counting Kit-8 assay (CCK-8, Shanghai U-Sea biotech, Shanghai,
China). Following incubation for 72 h at 37°C, 10 µl CCK-8 was
added to each well and incubated for 2 h in an incubator at 37°C.
The absorbance was determined using a multimode microplate reader
(Berthold Technologies GmbH & Co. KG, Bad Wildbad, Germany) at
450 nm.
Transwell assay
Cell invasion assays were performed using Transwell
chambers (Corning Incorporated, Corning, NY, USA). Transfected
cells (2×104) were seeded in the upper chamber
(pre-coated with Matrigel) and incubated in serum-free RPMI-1640
media, and the lower chamber was coated with complete RPMI-1640
medium supplemented with 10% fetal bovine serum (FBS). After 24 h
at 37°C, migrated cells on the bottom surface of the filters were
fixed with 4% paraformaldehyde for 1 h at 25°C, and stained with
0.1% crystal violet solution (MedChem Express, Shanghai, China) for
30 min at 25°C, and counted under an inverted microscope at ×100
magnifications; 3 fields per view were analyzed. The experiments
were performed in triplicate.
RT-qPCR
Total RNA was extracted from tumor tissues or
cultured cells with TRIzol (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocols. Then, cDNA was
synthesized with the M-MLV reverse transcriptase (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
protocol. Subsequently, transcripts were analyzed via qPCR using a
TaqMan MicroRNA Assay kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) on an ABI 7300 qPCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). Each experiment was repeated 3
times. The thermocycling conditions were: Denaturation at 95°C for
10 min, followed by 40 cycles of denaturation at 95°C for 15 sec
and elongation at 60°C for 1 min. The relative expression levels or
LIMD2 and miR-34a were calculated and normalized to endogenous that
of GAPDH and U6, respectively. The primer sequences were: miR-34a
forward, 5′-AACGAGACGACGACAGAC-3′ and reverse,
5′-UGGCAGUGUCUUAGCUGGUUGU-3′; U6 forward, 5′-AACGAGACGACGACAGAC-3′
and reverse, 5′-GCAAATTCGTGAAGCGTTCCATA-3′; LIMD2 forward,
5′-GCCCATCCTGTTGTGACCAA-3′ and reverse, 5′-ATGGTGAGTCTGCACCCTTC-3′;
GAPDH forward, 5′-ATGTTGCAACCGGGAAGGAA-3′ and reverse,
5′-AGGAAAAGCATCACCCGGAG-3′. Expression fold was calculated based on
the 2−ΔΔCq method (13).
Western blotting
Following transfection, total protein was extracted
from tumor tissues or cultured cells by radioimmunoprecipitation
assay buffer (Thermo Fisher Scientific, Inc.). All the protein
lysates were separated using 10% SDS-PAGE and transferred onto a
polyvinylidene fluoride membrane (Thermo Fisher Scientific, Inc.).
The membrane was blocked using 5% non-fat milk in PBS (Thermo
Fisher Scientific, Inc.) containing 0.1% Tween-20 (Sigma-Aldrich;
Merck KGaA) at room temperature for 3 h. Then, the membrane was
incubated for 2 h at 25°C with specific primary anti-human
antibodies: GAPDH (1:5,000; cat. no. SAB2701826; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) and LIMD2 (1:2,000; cat. no.
SAB1307182; Sigma-Aldrich; Merck KGaA), followed by incubation for
1 h at 25°C with a goat horseradish peroxidase-conjugated secondary
antibody (1:2,000; sc-2005; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA). Membranes were then washed with PBS for 10 min, and the
protein bands were visualized using an Enhanced Chemiluminescence
Western Blotting kit (Pierce; Thermo Fisher Scientific, Inc.), in
accordance with the manufacturer's protocol. Protein densitometry
was performed using ImageJ software (version 1.41; National
Institutes of Health, Bethesda, MD, USA). GAPDH was used as a
control. The experiment was repeated 3 times.
Luciferase reporter assay
The target gene LIMD2 and potential binding site
were predicted using TargetScan7 (http://www.targetscan.org/vert_71/). A luciferase
reporter assay was performed to determine the direct binding of
miR-34a to the target gene LIMD2. The wild type (WT) and mutant
(Mut) 3′-UTR sequence of LIMD2 was directly synthesized by Shanghai
GenePharma Co., Ltd. (Shanghai, China) and then inserted into pGL3
plasmid (Ambion; Thermo Fisher Scientific, Inc.) between Mlu I and
Bgl II. Cells (2×104) were cultured in 24-well plates,
and each well was transfected with 0.2 µg firefly luciferase
reporter plasmid, and equal amounts of miR-34a (50 nM) and NC
mimics (50 nM) using Lipofectamine® 2000. After 24 h,
Firefly and Renilla luciferase activities were measured
using the Luc-Pair miRNA Luciferase Assay kit (GeneCopoeia, Inc.,
Rockville, MD, USA) according to the manufacturer's protocols.
Luciferase activities were normalized to that of Renilla
luciferase.
Statistical analysis
All experiments were performed 3 times and results
expressed as mean ± standard deviation. All statistical analyses
were performed using SPSS software version 20.0 (IBM Corp., Armonk,
NY, USA). For comparisons between multiple groups, one-way analysis
of variance followed by a Tukey's post-hoc test was applied and a
two-tailed Student's t-test was performed for comparisons between
two groups. Pearson's correlation coefficient analysis was used to
determine the correlation between LIMD2 and miR-34a expression. The
association between the expression of LIMD2 with the
clinicopathological features of patients with non-small-cell lung
cancer was analysis using a χ2 test. P<0.05 was
considered to indicate a statistically significant difference.
Results
LIMD2 is upregulated in NSCLC
tissues
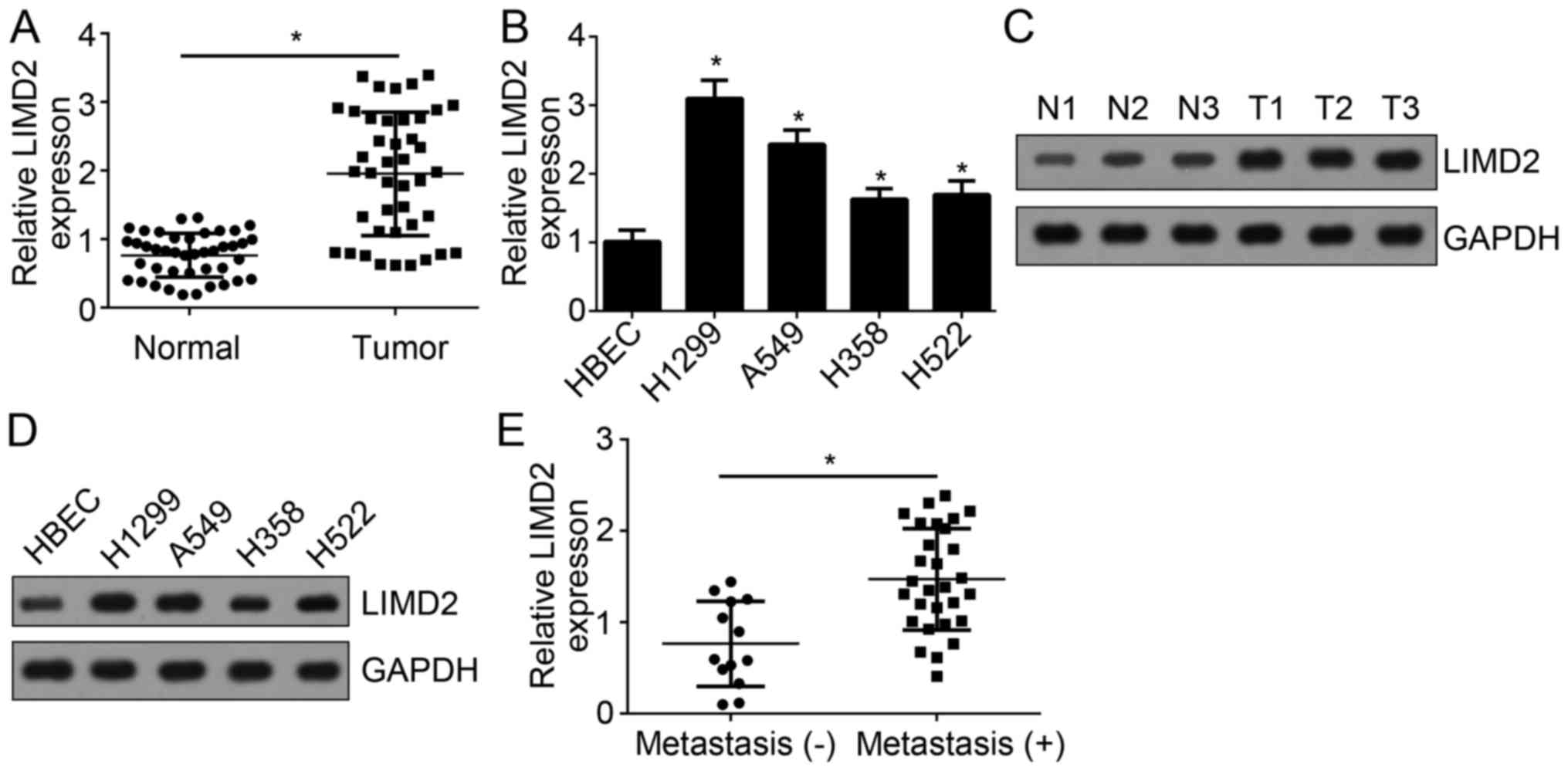
To investigate the function of LIMD2 in NSCLC, the
expression levels of LIMD2 were determined by RT-qPCR. The present
study reported that LIMD2 expression levels were significantly
upregulated in NSCLC tissues (n=41) compared with in adjacent
normal tissues (n=41) (Fig. 1A).
Additionally, the expression levels of LIMD2 in NSCLC cell lines
were determined. The results indicated that the expression levels
of LIMD2 were significantly upregulated in NSCLC cell lines,
including H1299, A549, H358 and H522 cells, compared with human
bronchial epithelial cells (HBECs) (Fig. 1B). Consistently, the protein
expression levels of LIMD2 in NSCLC tissues and cell lines were
notably upregulated compared with in normal tissue and HBECs,
respectively (Fig. 1C and D). In
addition, the present study reported that LIMD2 expression levels
were significantly higher in NSCLC tissues with metastasis than
tissues without (Fig. 1E). These
data suggest LIMD2 may be involved in NSCLC progression.
LIMD2 knockdown suppresses the
proliferation and invasion of NSCLC cells
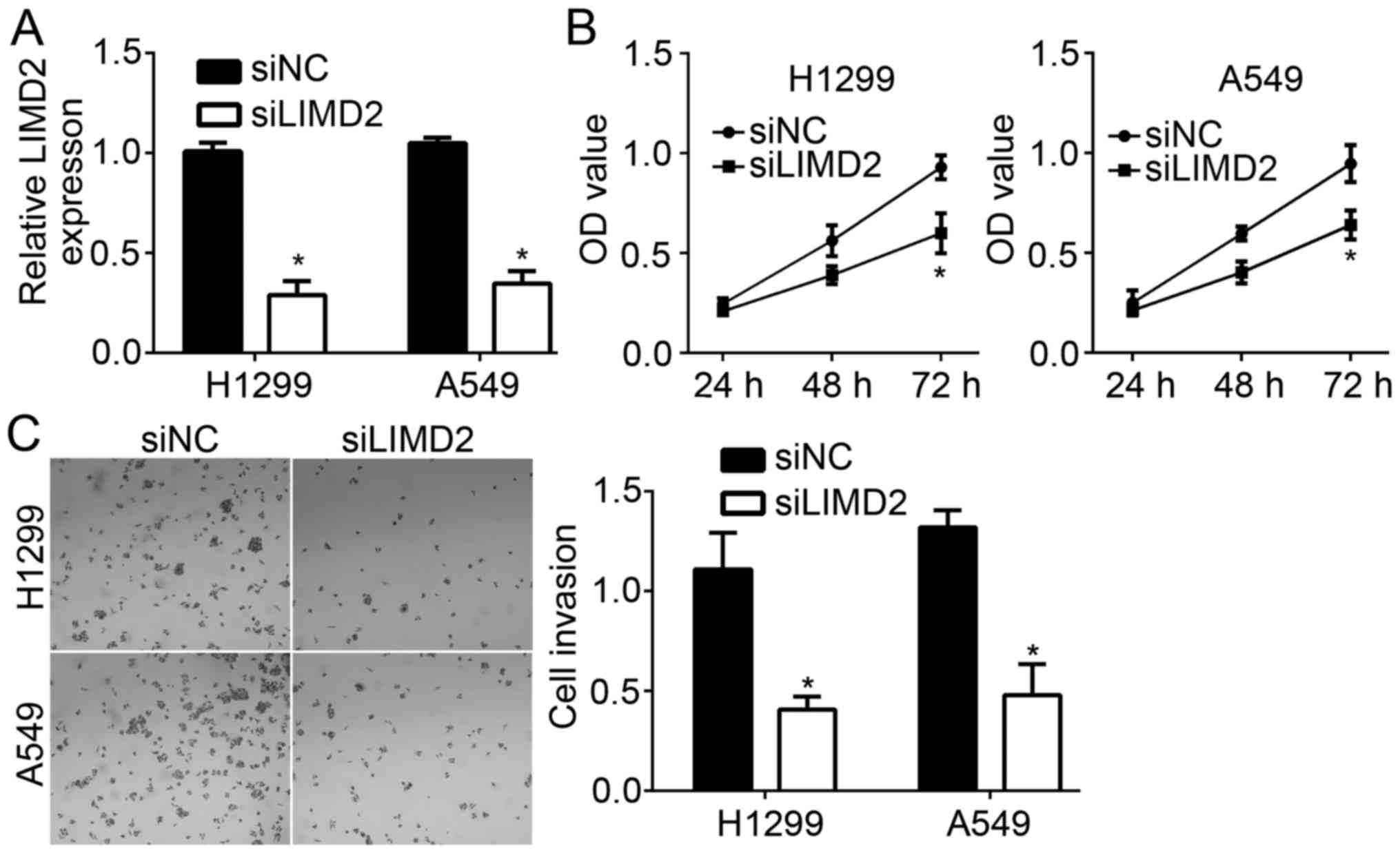
To determine the effects of LIMD2 on NSCLC cells,
LIMD2 expression was downregulated in H1299 and A549 cells via
transfection with specific siRNAs. RT-qPCR analysis revealed that
the expression levels of LIMD2 were significantly downregulated in
H1299 and A549 cells transfected with siLIMD2 compared with in the
control group (Fig. 2A). The
present study also performed CCK-8 and Transwell assays to
determine the effects of LIMD2 on cell proliferation and invasion,
respectively. The CCK-8 assay indicated that knockdown of LIMD2
significantly suppressed the proliferation of NSCLC cells at 72 h
compared with in the control group at 72 h (Fig. 2B). Furthermore, the present study
reported that LIMD2 silencing significantly reduced the number of
migrated cells compared with in the control group (Fig. 2C). The above data indicated that
LIMD2 contributes to NSCLC progression.
LIMD2 is downregulated by miR-34a
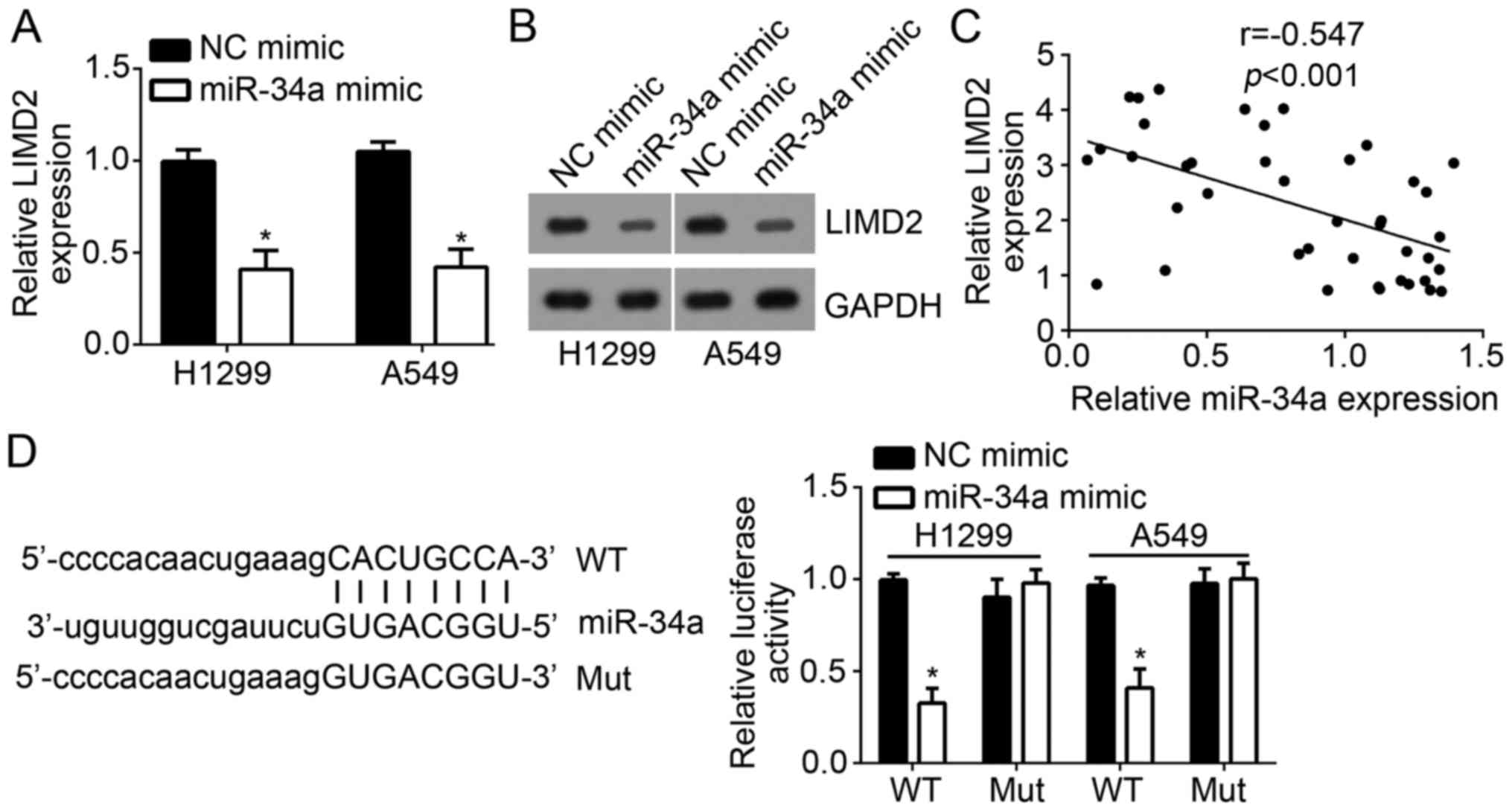
The present study investigated the mechanism
underlying the regulation of LIMD2 expression in NSCLC cells. The
results demonstrated that overexpression of miR-34a significantly
suppressed the mRNA expression levels of LIMD2 in H1299 and A549
cells compared with in the control (Fig. 3A). Additionally, western blot
analysis revealed that ectopic expression of miR-34a markedly
decreased the protein expression levels of LIMD2 in H1299 and A549
cells compared with in the control (Fig. 3B). Furthermore, an inverse
correlation between the expression of LIMD2 and miR-34a was
observed in NSCLC tissues (Fig.
3C).
miRNAs have been demonstrated to regulate the
degradation of target mRNAs (12).
To investigate whether LIMD2 is a direct target of miR-34a,
bioinformatics analysis was conducted using TargetScan. The results
indicated that LIMD2 is a potential target of miR-34a. To verify
this prediction, luciferase reporter assays with the WT and Mut
3′-UTR sequence of LIMD2 mRNA were performed. The results
demonstrated that overexpression of miR-34a significantly
suppressed the luciferase activity in H1299 and A549 cells
transfected with WT-LIMD2 3′-UTR reporter plasmid compared with in
the control group; no notable differences were observed within
cells overexpressing miR-34a containing the Mut-3′UTR compared with
in the controls (Fig. 3D). These
results demonstrated that miR-34a directly targets LIMD2.
MiR-34a is downregulated in NSCLC
tissues
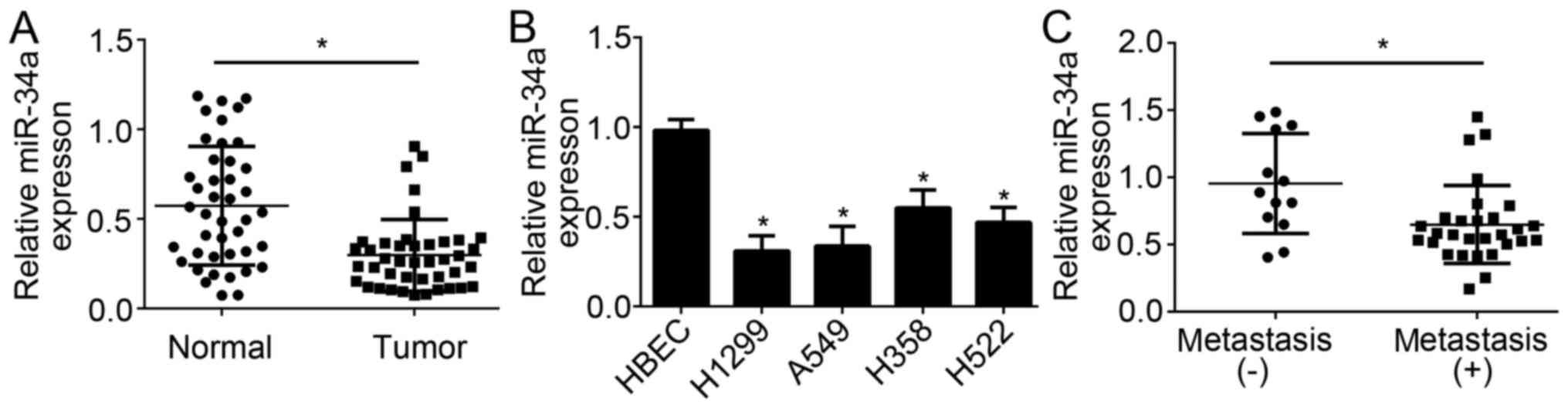
LIMD2 was proposed to act as an oncogene in NSCLC in
the present study; however, miR-34a inhibited the expression of
LIMD2 in NSCLC cells. To determine the role of miR-34a in NSCLC,
the expression levels of miR-34a were determined by RT-qPCR. The
results demonstrated that miR-34a expression levels were
significantly downregulated in NSCLC tissues compared with in
adjacent normal tissues (Fig. 4A).
Additionally, lower expression levels of miR-34a were observed in
NSCLC cell lines compared with in HBEC cells (Fig. 4B). Furthermore, the expression
levels of miR-34a were significantly lower in NSCLC tissues with
metastasis compared with in those without metastasis (Fig. 4C). These data implied that miR-34a
may be a tumor suppressor in NSCLC.
MiR-34a suppresses NSCLC cell
proliferation and invasion
The present study investigated whether the effects
of miR-34a on NSCLC cells are associated with LIMD2. RT-qPCR
analysis indicated that the overexpression of LIMD2 significantly
increased the mRNA expression levels of LIMD2 in H1299 and A549
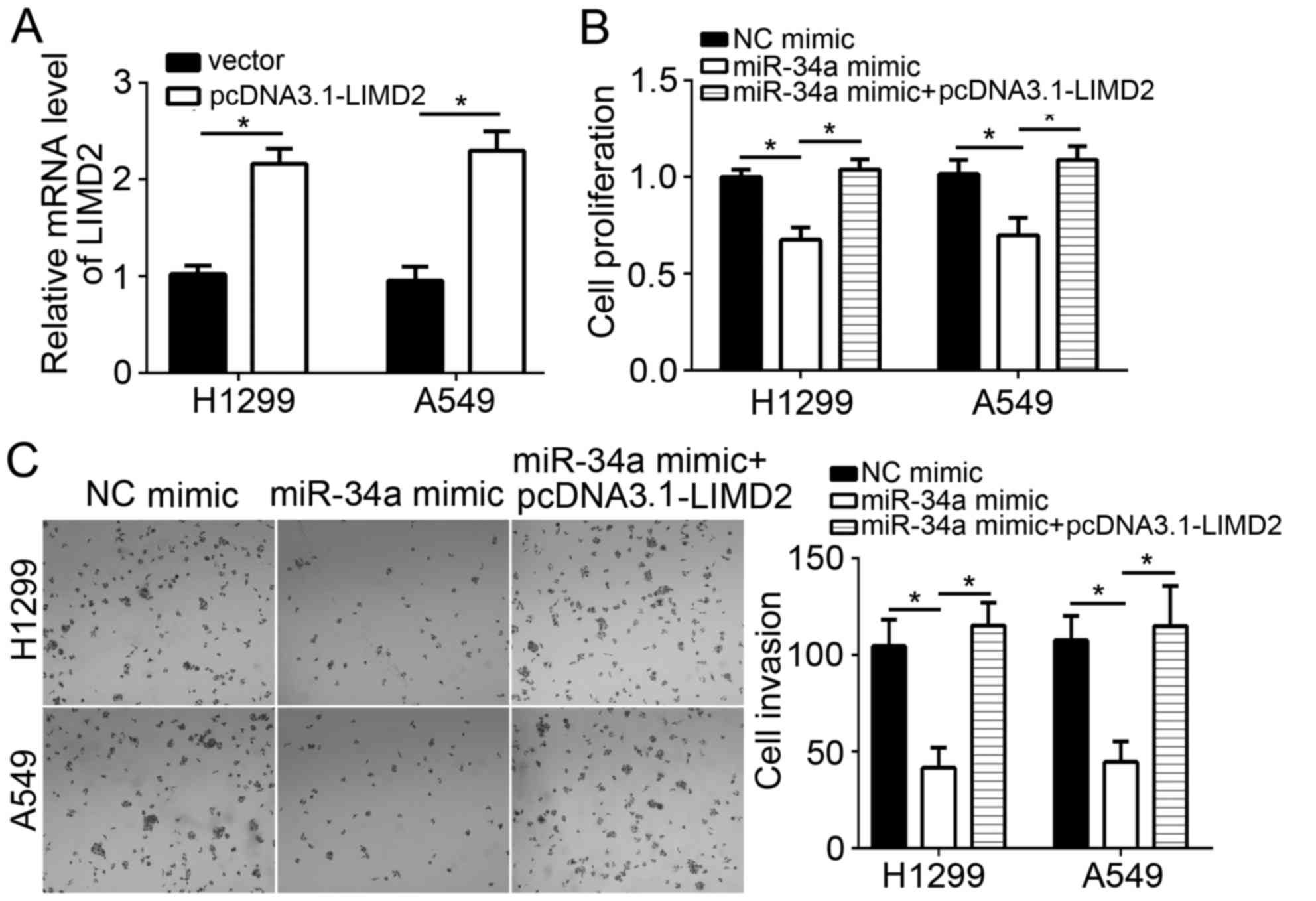
cells compared with in the vector-transfected group (Fig. 5A). CCK-8 and Transwell assays
revealed that the overexpression of miR-34a significantly
suppressed the proliferation and invasion, of NSCLC cells
respectively, compared with in the control group (Fig. 5B). Of note, restoration of LIMD2
expression significantly reversed the inhibitory effects of miR-34a
on H1299 and A549 cell proliferation and invasion compared with in
the miR-34a overexpression group (Fig.
5B and C). Taken together, the data from the present study
demonstrated that miR-34a suppressed NSLCL progression through
targeting LIMD2.
Discussion
As one of the most prevalent and malignant types of
tumors worldwide, lung cancer accounts for numerous
cancer-associated mortalities (1).
NSCLC contributes to 80% of all lung cancer cases (12). The study of lung cancer has
received increasing attention. The mechanism underlying the
development and progression of lung cancer remains largely unknown;
effective therapeutic strategies for the treatment of lung cancer
are urgently required. In the present study, it was proposed that
LIMD2 promoted the proliferation and invasion of NSCLC cells. In
addition, the regulatory mechanism of LIMD2 expression was
investigated in the present study, in which miR-34a was
demonstrated to directly target LIMD2 in NSCLC cells.
Previous studies have indicated that LIMD2 is a
novel metastasis-associated cytoplasmic protein, whose endogenous
expression levels are correlated with malignant behaviors in breast
cancer, bladder and thyroid cancers, and melanoma (5,14).
Knockdown of LIMD2 has been revealed to inhibit cell invasive
ability, motility and cell morphology, which was consistent with
tumor grade in human thyroid cancer (14); however, whether LIMD2 serves a
similar role in other cancers remains unknown. To determine the
correlation between LIMD2 and NSCLC, the expression levels of LID2
in NSCLC tissues were analyzed by RT-qPCR in the present study;
LIMD2 expression levels were higher in NSCLC cell lines than in
normal cells. Additionally, the present study reported that LIMD2
expression was positively correlated with tumor metastasis, and
tumor, node and metastasis stages in clinical samples (Table I). Furthermore, the expression
levels of LIMD2 were higher in the metastatic cell line H1299 than
within A549 cells. This further indicated that LIMD2 is associated
with tumor metastasis. These results suggested that LIMD2 is
involved in the progression of NSCLC. Functional experiments
demonstrated that LIMD2 knockdown significantly inhibited the
proliferation and invasion potentials of NSCLC cells in the present
study.
MiRNAs are a group of short noncoding RNAs of ~22
nucleotides in length (12).
MiRNAs have been widely demonstrated to regulate gene expression
via binding to the mRNA of target genes (15–17);
miRNAs are often aberrantly expressed in cancer cells (18). As miRNAs have been observed to
regulate the proliferation, migration, invasion and other
biological processes, a close association between miRNA expression
and tumor progression has been reported (19). For instance, Qiao et al
(20) reported that miR-154
inhibited the growth and metastasis of gastric cancer cells by
directly targeting metadherin (20). Cao et al (17) revealed that miR-552 promoted tumor
cell proliferation and migration by directly targeting dachshund
family transcription factor 1 via the Wnt/β-catenin signaling
pathway in colorectal cancer (17). The mechanism underlying the
regulation of LIMD2 expression by miRNAs is not well studied. As of
the important role of LIMD2 in NSCLC, it is necessary to determine
the regulatory mechanism of LIMD2 expression. In the present study,
miR-34a was reported to directly target LIMD2 mRNA; overexpression
of miR-34a notably inhibited the mRNA and protein expression levels
of LIMD2 in NSCLC cells. A recent report reported that miR-34a
serves as a tumor suppressor in lung cancer (21). The present study also indicated
that miR-34a inhibited the proliferation and invasion of NSCLC
cells.
In conclusion, the present study demonstrated the
essential role of LIMD2 in NSCLC cells; LIMD2 suppressed the
proliferation and invasion of NSCLC cells, which suggested that
LIMD2 may be a promising therapeutic target for the treatment of
NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FW and ZL performed the majority of the experiments
and analyzed the data. LX, YiL, YL, XZ and YW collected sample
tissues and performed some experiments. DL made substantial
contributions to the concept and design of the present study and
wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
For the use of human samples, the present study was
approved by the Institutional Ethics Committee of the Second
Affiliated Hospital of Harbin Medical University (Harbin, China)
and all enrolled patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ramalingam SS, Owonikoko TK and Khuri FR:
Lung cancer: New biological insights and recent therapeutic
advances. CA Cancer J Clin. 61:91–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paz-Ares L: Beyond first-line NSCLC
therapy: Chemotherapy or erlotinib? Lancet Oncol. 13:225–227. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Geng GJ, Yang YT, Jiang J, Yu XY and Fa
XE: MicroRNA-30a suppresses non-small-cell lung cancer by targeting
Myb-related protein B. Exp Ther Med. 15:1633–1639. 2018.PubMed/NCBI
|
|
5
|
Peng H, Talebzadeh-Farrooji M, Osborne MJ,
Prokop JW, McDonald PC, Karar J, Hou Z, He M, Kebebew E, Orntoft T,
et al: LIMD2 is a small LIM-only protein overexpressed in
metastatic lesions that regulates cell motility and tumor
progression by directly binding to and activating the
integrin-linked kinase. Cancer Res. 74:1390–1403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cerutti JM, Oler G, Michaluart P Jr,
Delcelo R, Beaty RM, Shoemaker J and Riggins GJ: Molecular
profiling of matched samples identifies biomarkers of papillary
thyroid carcinoma lymph node metastasis. Cancer Res. 67:7885–7892.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang X, Tang S, Le SY, Lu R, Rader JS,
Meyers C and Zheng ZM: Aberrant expression of oncogenic and
tumor-suppressive microRNAs in cervical cancer is required for
cancer cell growth. PLoS One. 3:e25572008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu H, Lei C, He Q, Pan Z, Xiao D and Tao
Y: Nuclear functions of mammalian MicroRNAs in gene regulation,
immunity and cancer. Mol Cancer. 17:642018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie F, Huang Q, Liu CH, Lin XS, Liu Z, Liu
LL, Huang DW and Zhou HC: MiR-1271 negatively regulates AKT/MTOR
signaling and promotes apoptosis via targeting PDK1 in pancreatic
cancer. Eur Rev Med Pharmacol Sci. 22:678–686. 2018.PubMed/NCBI
|
|
10
|
Xia D, Tian S, Chen Z, Qin W and Liu Q:
miR302a inhibits the proliferation of esophageal cancer cells
through the MAPK and PI3K/Akt signaling pathways. Oncol Lett.
15:3937–3943. 2018.PubMed/NCBI
|
|
11
|
Xie Y, Zhang H, Guo XJ, Feng YC, He RZ, Li
X, Yu S, Zhao Y, Shen M, Zhu F, et al: Let-7c inhibits
cholangiocarcinoma growth but promotes tumor cell invasion and
growth at extrahepatic sites. Cell Death Dis. 9:2492018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang N, Zhang Y and Liang H: microRNA-598
inhibits cell proliferation and invasion of glioblastoma by
directly targeting metastasis associated in colon cancer-1. Oncol
Res. Feb 14–2018.(Epub ahead of print). View Article : Google Scholar
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(T)(-Delta Delta C) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pinheiro Dos, Santos MJC, Bastos AU, da
Costa VR, Delcelo R, Lindsey SC, Colozza-Gama GA, Peng H, Rauscher
FJ III, Oler G and Cerutti JM: LIMD2 is overexpressed in BRAF
V600E-positive papillary thyroid carcinomas and matched lymph node
metastases. Endocr Pathol. 29:222–230. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tong X, Wang X, Wang C and Li L: Elevated
levels of serum MiR-152 and miR-24 in uterine sarcoma: Potential
for inducing autophagy via SIRT1 and deacetylated LC3. Br J Biomed
Sci. 75:7–12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
You Y, Tan J, Gong Y, Dai H, Chen H, Xu X,
Yang A, Zhang Y and Bie P: MicroRNA-216b-5p Functions as a
Tumor-suppressive RNA by Targeting TPT1 in pancreatic cancer cells.
J Cancer. 8:2854–2865. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao J, Yan XR, Liu T, Han XB, Yu JJ, Liu
SH and Wang LB: MicroRNA-552 promotes tumor cell proliferation and
migration by directly targeting DACH1 via the Wnt/β-catenin
signaling pathway in colorectal cancer. Oncol Lett. 14:3795–3802.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang M, Huang S and Long D: MiR-381
inhibits migration and invasion in human gastric carcinoma through
downregulatedting SOX4. Oncol Lett. 14:3760–3766. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao S, Gao X, Zang S, Li Y, Feng X and
Yuan X: MicroRNA-383-5p acts as a prognostic marker and inhibitor
of cell proliferation in lung adenocarcinoma by cancerous inhibitor
of protein phosphatase 2A. Oncol Lett. 14:3573–3579. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qiao W, Cao N and Yang L: MicroRNA-154
inhibits the growth and metastasis of gastric cancer cells by
directly targeting MTDH. Oncol Lett. 14:3268–3274. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song C, Lu P, Sun G, Yang L and Wang Z and
Wang Z: miR-34a sensitizes lung cancer cells to cisplatin via
p53/miR-34a/MYCN axis. Biochem Biophys Res Commun. 482:22–27. 2017.
View Article : Google Scholar : PubMed/NCBI
|