Introduction
Fatty liver disease, particularly non-alcoholic
fatty liver disease (NAFLD), is a major subtype of liver disease
(1). Fatty liver disease is
characterized by lipid accumulation in the liver and is closely
associated with mitochondrial abnormality (2). Generally, abnormal mitochondrial
function and increased mitochondrial reactive oxygen species (ROS)
affect fatty liver disease via impaired insulin sensitivity
(3). Elevated mitochondrial ROS
production can also cause steatohepatitis, due to fatty acid
oxidation disorder, which is an increased inflammatory response in
the liver (4). Therefore,
mitochondrial-targeted antioxidants are considered potential,
effective therapeutic compounds for the treatment of fatty liver
disease (5).
Millions of people worldwide suffer from chronic
hepatitis B virus (HBV) infection (6). Chronic HBV infection affects numerous
organ systems and is associated with several human diseases
(7). The pathogenesis of liver
disease is the most studied aspect of HBV infection, since the
liver is the most affected organ in HBV carriers (8); however, the underlying molecular
mechanisms by which HBV affects the liver remain poorly understood.
Recently, a case-control study involving different populations
indicated that patients with HBV infection suffer from fewer fatty
liver diseases, including NAFLD (9); however, some studies hold the
opposite view, that HBV infection increases the prevalence of NAFLD
(10,11).
The HBV genome is a partially double-stranded,
circular DNA molecule that encodes four main HBV proteins: Proteins
C, X, P and S. Of these, hepatitis B protein X (HBx) is a
multifunctional regulator involved in cellular signal transduction
pathways and gene transcription (12). HBV infection and HBx expression in
cells affect cellular processes, including cell growth, cell cycle
progression and apoptosis, thus indicating a pivotal role for HBx
in HBV infection (13). Therefore,
it is not surprising that HBV infection is associated with NAFLD
(14). HBx was originally detected
in the cytoplasm and nucleus (12), and due to its transcriptional
function, HBx can alter the gene expression of the host cell
(15). In mitochondria, HBx has
been reported serve as an enhancer of mitochondrial function by
direct interaction with the mitochondrial respiratory chain complex
subunit (15). However, HBx may
also contribute to mitochondrial function via increased fission and
mitophagy (16). These conflicting
findings increase the difficulty of analyzing the mitochondrial
role in HBV/HBx-associated fatty liver disease, specifically NAFLD.
While there is no consensus in the literature regarding the
association between HBV and protection against NAFLD, to understand
the effects of HBV infection on NAFLD, the present study conducted
a large-scale case-control study on Han Chinese individuals. In
addition, a functional study using HBx-expressing hepatocytes was
performed to validate the effect of HBx-mediated HBV infection on
NAFLD.
Patients and methods
Subjects
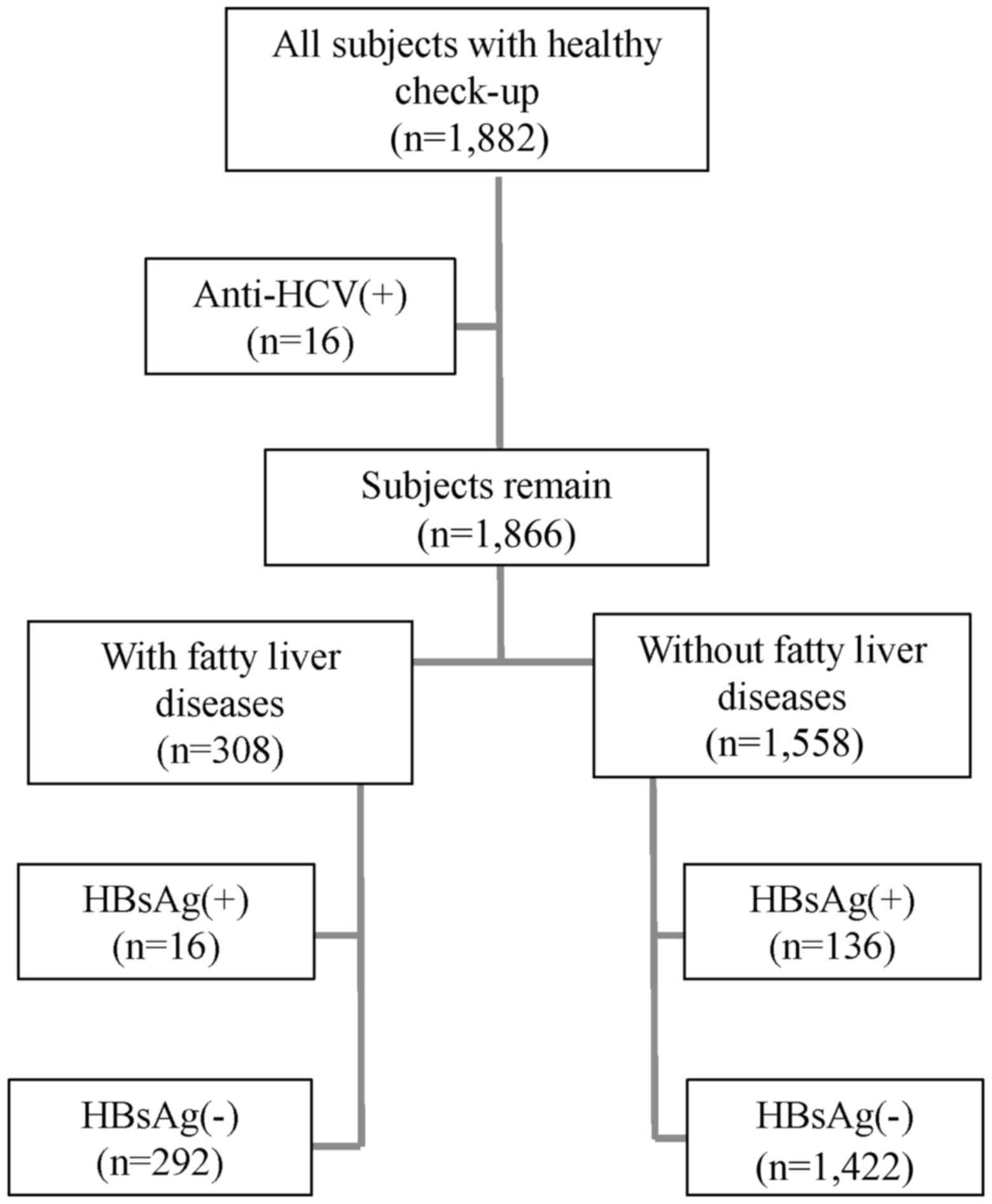
A total of 1,882 female subjects that underwent
comprehensive health examination were recruited at the First
Affiliated Hospital of Wenzhou Medical University (Wenzhou, China)
between October 2011 and March 2014 (mean age ± standard deviation,
46.42±10.291 years; median age, 45 years; age range 19–83 years).
Since subjects with hepatitis C virus (HCV) have alterations in
lipid metabolism (17), HCV
infection was tested by Roche Cobas7 Amplicor HCV Monitor Test,
version 2.0, (Roche Molecular Diagnostics, Pleasanton, CA, USA).
Subsequently, 16 subjects with chronic HCV infection were excluded
from the present study; the response rate for the enrolled subjects
was thus 99.1% (1,866/1,882; Fig.
1). By ultrasonography, using the Hitachi VISION Preirus
(Hitachi, Ltd., Tokyo, Japan), according to four ultrasound
criteria (liver brightness, deep attenuation, hepatorenal echo
contrast and vascular blurring), 308 and 1,558 subjects were
diagnosed with fatty liver and non-fatty liver disease,
respectively. The presence of hepatitis B virus antigen (HBsAg) was
determined in all enrolled subjects using the Roche Cobas 6800/8800
Systems (Roche Diagnostics). As demonstrated in Fig. 1, 16 subjects from the fatty liver
group and 136 subjects from the non-fatty liver group were
considered to have HBV, due to the presence of HBsAg (9). Informed consent was obtained from all
subjects and this study was approved by the Ethical Committee of
Wenzhou Medical University.
Cells and culture conditions
The hepatocyte cell lines Huh7 and MIHA were
purchased from the Cell Resource Center, Chinese Academy of Medical
Sciences (Beijing, China); the two cell types are widely used for
the study of HBV infection (18,19).
All cells were cultured in high-glucose Dulbecco's modified Eagle's
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and were maintained at 37°C in a humidified
atmosphere containing 5% CO2.
Plasmid infection
The coding sequence of HBx with a c-Myc tag at the C
terminus (Gene ID: 944566) was synthesized and cloned into a pCDH
plasmid (donated by Dr. Zhenfen Chi, Beijing Institute of
Genomics). 293T cells were from Cell Resource Center, Chinese
Academy of Medical Sciences (Shanghai, China). Briefly, 293T cells
(~1×106) were co-transfected with 1 µg pCDH with or
without HBx, 1 µg psPAX2, and 0.5 µg PMD2G (donated by Dr. Zhenfen
Chi; 2:2:1) for 72 h. Culture medium with packaged lentivirus was
used to infect Huh7 and MIHA cells for 24 h. Cells expressing HBx
were selected using 4 µg/ml puromycin (Sigma-Aldrich; Merck KGaA).
An empty plasmid was used as a control.
Antibodies and immunoblotting
Protein was extracted from whole cell lysates using
radioimmunoprecipitation assay lysis buffer (Cell Signaling
Technology, Inc., Danvers, MA, USA). Protein concentration was
quantified using Pierce BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.). Samples containing 30 µg proteins were
electrophoresed through 12% Sodium dodecyl sulfate polyacrylamide
gel electrophoresis (SDS-PAGE), and transferred onto PVDF membranes
with 0.2 µm pore size (Thermo Fisher Scientific, Inc.), the
membrane was blocked with tris buffered saline with 1% Tween 20
(TBST) buffer containing 10% non-fat milk at room temperature for 1
h. Proteins were probed with anti-HBx antibody (ab2741; 1:1,000;
Abcam, Cambridge, UK) or anti-β-actin at 4°C for 16 h (3700;
1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA).
Horseradish peroxidase conjugated anti mouse IgG (4410; 1:2,000;
Cell Signaling Technology, Inc.) was used as secondary antibodies
and incubated for 1 h at room temperature. Signals were detected
with Super Signal West Pico chemiluminescent substrate (Thermo
Fisher Scientific, Inc.). Integrated optical density was
semi-quantified using a Gel-Pro Analyzer 4.0 (Media Cybernetics,
Inc., Rockville, MD, USA).
Immunofluorescence staining
Cells (3×105) were treated with 100 nM
MitoTracker Red (Invitrogen; Thermo Fisher Scientific, Inc.) for 10
min at 37°C and were fixed with 4% paraformaldehyde for 30 min in
room temperature. After treatment with 0.2% Triton X-100 for 3 min,
cells were incubated overnight with anti-c-Myc antibody (2276s;
1:1,000; Cell Signaling Technology, Inc.). Subsequently, cells were
washed with PBS and incubated with an Alexa Fluor®
488-labeled immunoglobulin secondary antibody (9854; 1:250; Cell
Signaling Technology, Inc.) for 45 min at room temperature. A final
incubation was performed to stain the cells with DAPI (Beyotime
Institute of Biotechnology, Haimen, China) for 5 min at room
temperature. Images were captured using a confocal laser microscope
at a magnification ×400 (Nikon Corporation, Tokyo, Japan).
Measurement of endogenous oxygen
consumption
The endogenous oxygen consumption of intact cells
was determined using a Clark-type oxygen electrode (Oroboros
Instruments Corp., Innsbruck, Austria), as described previously
(20). After recording basal
respiration, oligomycin (an ATP synthase inhibitor; 0.2 µg/ml;
Sigma-Aldrich; Merck KGaA) was added to measure uncoupling
respiration, and carbonyl cyanide m-chlorophenyl hydrazone (CCCP; a
mitochondrial oxidative phosphorylation uncoupling agent; 0.1 µM;
Sigma-Aldrich; Merck KGaA) was added to measure maximum
respiration.
ATP, mitochondrial membrane potential
(MMP) and ROS measurements
ATP was measured using an ATP determination kit
(A22066; Molecular Probes; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Briefly, cells were grown
in 6-well plates to ~80% confluence. Cells (~1×106) were
then washed with cold PBS and boiled in 100 µl boiling buffer (100
mM Tris, 4 mM EDTA, adjusted to pH 7.75 with acetic acid) for 90
sec. Supernatants were retrieved by centrifugation at 10,000 × g
for 1 min. ATP content was determined by measuring the luminescence
of supernatants mixed with luciferase assay buffer and luminescence
measured at ~560 nm using a Varioskan™ Flash Multimode Reader (both
Thermo Fisher Scientific, Inc.).
Mitochondrial ROS levels were measured as described
previously (21). Briefly,
1×106 cells were washed in Hank's buffered salt solution
(HBSS; Sigma-Aldrich; Merck KGaA) resuspended in HBSS containing 5
µM Mito SOX (Molecular Probes; Thermo Fisher Scientific, Inc.), and
incubated at 37°C for 15 min. Cells were then washed twice with
HBSS, and fluorescence with excitation/emission at 510/580 nm was
recorded using a Varioskan™ Flash Multimode Reader. To measure the
ROS level in cells with NAC, Huh7 and MIHA cells were cultured with
10 mM NAC for 48 h at 37°C in a cell culture incubator
(Sigma-Aldrich; Merck KGaA) for 48 h. MMP was determined using the
cationic fluorescent redistribution dye TMRM (Thermo Fisher
Scientific, Inc.) as described previously (22). ATP, MMP and ROS signals were
normalized by the protein concentration measured with the Pierce
BCA Protein Assay kit.
Oil Red O (ORO) staining
Cells (5×105) were fixed with 4%
formaldehyde at room temperature for 25 min, and were stained with
freshly diluted ORO reagent (Sangon Biotech Co., Ltd., Shanghai,
China; diluted distilled water:Oil red O 4:6) for 20 min at room
temperature. Subsequently, cells were stained with 0.5% hematoxylin
(Sangon Biotech Co., Ltd.) for 5 min at room temperature and washed
with PBS twice. Lipid accumulation in the cells was observed using
an inverted phase contrast optical microscope at magnification,
×200 (ECLIPSE Ti-S; Nikon Corporation). To measure lipid droplet
accumulation in cell with NAC treatment, Huh7 and MIHA cells were
cultured with 10 nm NAC for 48 h at 37°C in a cell culture
incubator before ORO staining.
Statistical analysis
Clinical data are presented as means ± standard
deviation. Categorical variables were compared using the
χ2 test. The association between fatty liver and HBV
infection was adjusted for age using the logistic regression. Data
from cell models are presented as the means ± standard error of the
mean. To evaluate cell model data, an independent Student's t-test
was used. P<0.05 was considered to indicate a statistically
significant difference. All statistical analyses were performed
using SPSS 21.0 (IBM Corp., Armonk, NY, USA).
Results
HBV infection is not associated with
fatty liver disease occurrence
Only 30% of women in China consume alcohol, and few
of them have at least one drink per day (23). Correspondingly, in women with fatty
liver disease, few are diagnosed with alcoholic fatty liver disease
(24). Therefore, to assess the
risk of HBV infection in NAFLD, women undergoing health check-ups
were recruited to the present study. The clinical characteristics
in subjects with or without HBV infection were initially analyzed.
As demonstrated in Table I,
HBV-infected subjects had significantly higher levels of alanine
aminotransferase, aspartate aminotransferase and γ-glutamyl
transferase than HBV-free subjects, which indicated that hepatocyte
injury was present in HBV carriers. Additionally, although the
levels of high-density lipoprotein, a protective factor against
fatty liver disease, were significantly lower in HBV-infected
patients compared with in HBV-free subjects, other fatty liver
disease-associated factors, such as total cholesterol, low-density
lipoprotein and triglycerides, were not significantly affected by
HBV (Table I). Furthermore, the
incidence of fatty liver in HBV-infected subjects (16/152, 10.5%)
was not significantly different from that in HBV-free subjects
(292/1,714, 17%) when adjusted for age (Table II). These findings indicated that
HBV was not associated with a decreased incidence of NAFLD.
 | Table I.Factors associated with HBV and
non-HBV subjects. |
Table I.
Factors associated with HBV and
non-HBV subjects.
| Characteristic |
HBsAg+ |
HBsAg− |
P-valuea |
|---|
| Number | 152 | 1,714 | – |
| Age (years) | 45.1±8.7 | 46.5±10.4 | 0.052 |
| BMI
(kg/m2) | 23.94±3.16 | 22.85±3.07 | <0.001 |
| SBP (mmHg) | 118.6±14.9 | 115.9±14.9 | 0.010 |
| DBP (mmHg) | 78.7±9.7 | 75.3±9.3 | <0.001 |
| ALT (U/I) | 37.1±41.0 | 19.8±17.6 | <0.001 |
| AST (U/I) | 30.9±29.8 | 20.5±10.2 | <0.001 |
| GGT (U/I) | 36.2±64.2 | 21.2±19.6 | <0.001 |
| Glucose
(mg/dl) | 5.6±1.1 | 5.5±1.0 | 0.363 |
| Total cholesterol
(mg/dl) | 4.7±0.8 | 4.9±0.9 | 0.261 |
| LDL-C (mg/dl) | 2.8±0.7 | 2.7±0.8 | 0.094 |
| HDL-C (mg/dl) | 1.4±0.4 | 1.5±0.3 | P<0.001 |
| Triglycerides
(mg/dl) | 1.4±0.8 | 1.2±1.0 | 0.061 |
| Table II.Unadjusted prevalence and
age-adjusted odds ratios for fatty liver by HBV status. |
Table II.
Unadjusted prevalence and
age-adjusted odds ratios for fatty liver by HBV status.
|
|
| Fatty liver |
|---|
|
|
|
|
|---|
| Group | Total number | Total cases
(prevalence %) | OR | 95% CI |
P-valuea |
|---|
|
HBsAg− | 1,714 | 292 (17.0) | 1 |
|
|
|
HBsAg+ |
152 | 16 (10.5) | 0.656 | 0.379–1.134 | 0.131 |
Expression of HBx protein in Huh7 and
MIHA cells increases mitochondrial function
HBx is the major HBV component that contributes to
steatohepatitis (25). To examine
if and how HBV affects the occurrence of NAFLD, the HBx protein was
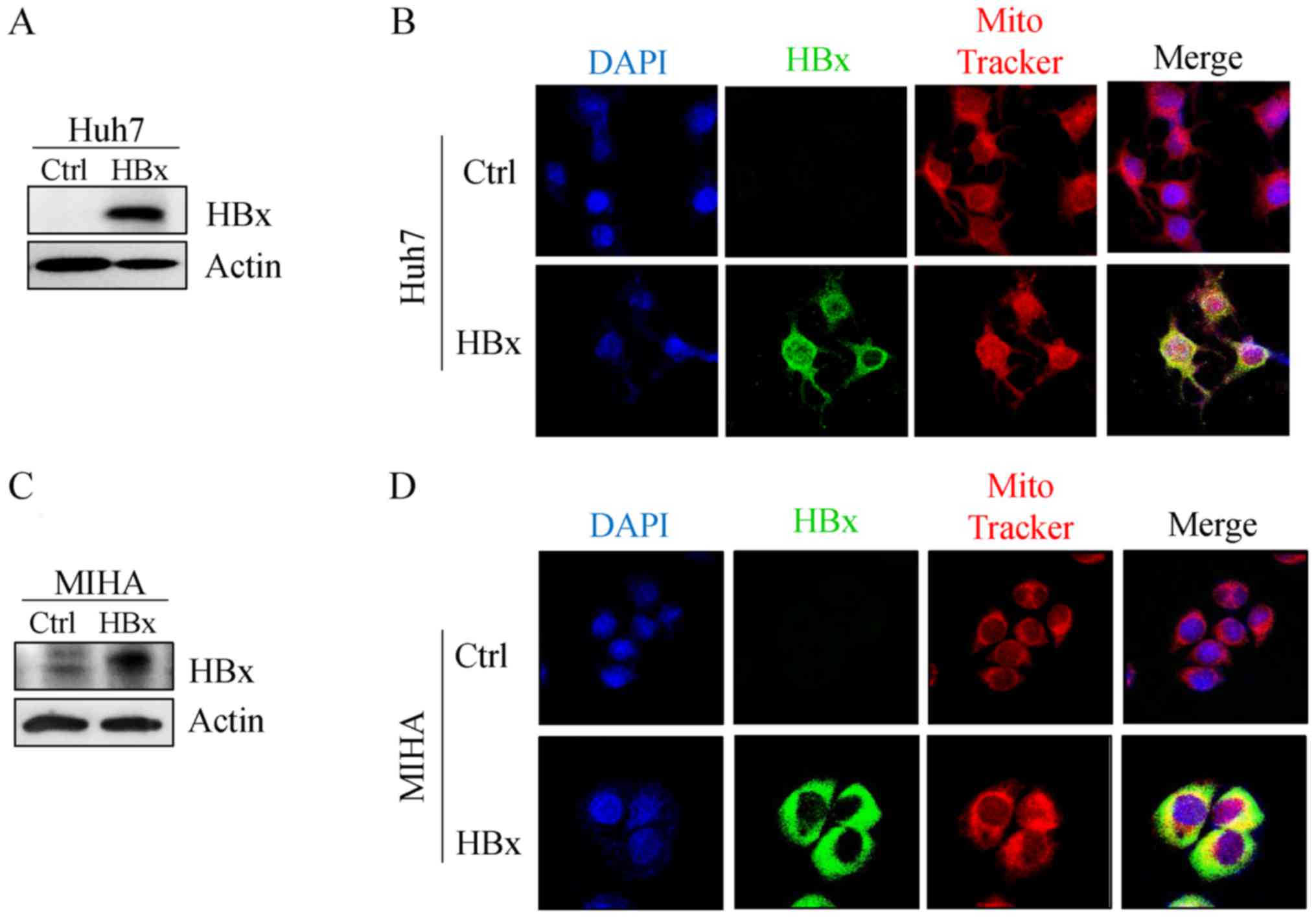
expressed in Huh7 and MIHA cell lines. Mitochondrial localization
of HBx was observed in Huh7 (Fig. 2A
and B) and MIHA cells (Fig. 2C and
D), which confirmed that HBx targeted the mitochondria in the
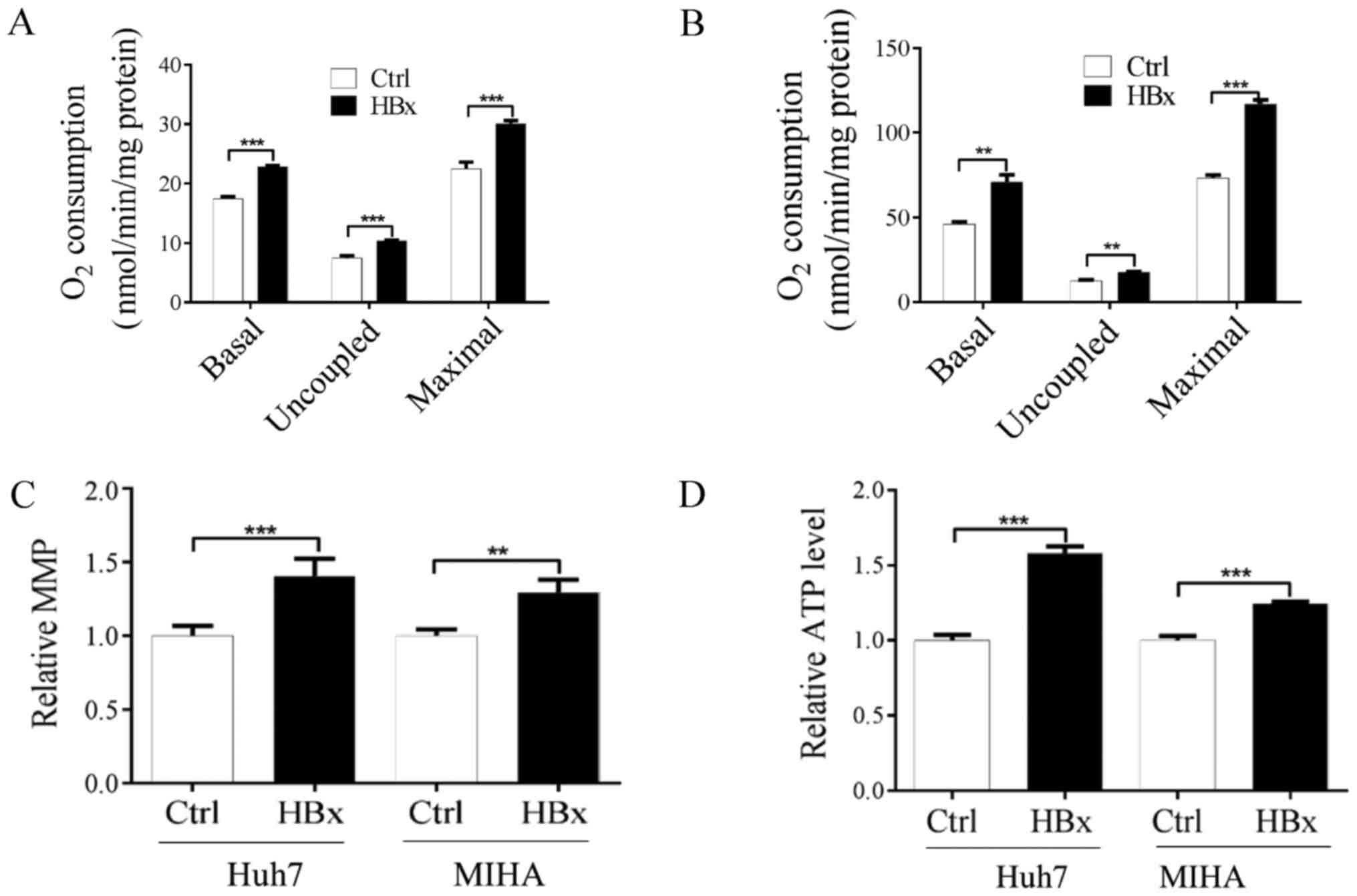
cytoplasm. Subsequently, whether HBx affects mitochondrial function
was investigated by measuring mitochondrial respiration, MMP and
ATP content. As shown in Fig. 3A and
B, endogenous basal mitochondrial respiration in cells
expressing HBx was significantly increased compared with in the
control cells. In addition, lowest mitochondrial respiration
(uncoupled respiration) in the presence of oligomycin (0.2 µg/ml)
and maximal mitochondrial respiration in the presence of CCCP (0.1
µM) were significantly increased in the two HBx-expressing cells
compared with the control cells (Fig.
3A and B). To further clarify the alterations of mitochondrial
function in cells expressing HBx, the MMP and ATP content were
investigated. As shown in Fig. 3C and
D, MMP and ATP content were significantly increased in
HBx-expressing Huh7 and MIHA cells compared with in control cells.
Although the underlying mechanism is not currently known, the
present results indicated that HBx may be associated with increased
mitochondrial function.
HBx mediates lipid droplet
accumulation in Huh7 and MIHA cells by altering mitochondrial ROS
levels
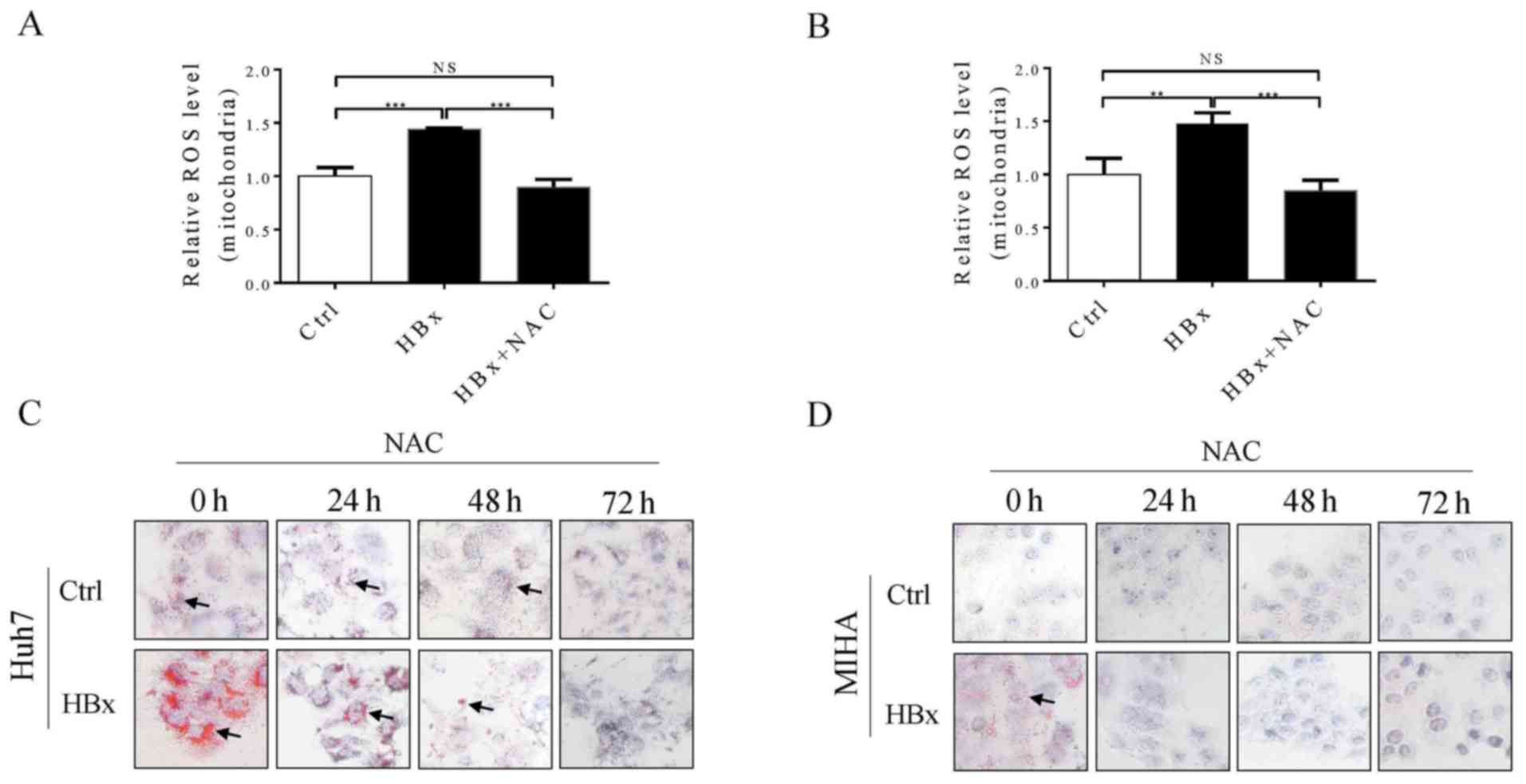
Excessive mitochondrial ROS production serves an
important role in the development of NAFLD (26). Therefore, mitochondrial ROS levels
in Huh7 and MIHA cells with or without HBx were investigated. As
shown in Fig. 4A and B,
mitochondrial ROS levels were significantly increased in Huh7 and
MIHA cells expressing HBx compared with in control cells without
HBx, and this effect was reversed by treating cells with
N-acetylcysteine for up to 48 h. Furthermore, it was demonstrated
that Huh7 and MIHA cells expressing HBx exhibit increased fatty
droplet formation compared with in the control cells (Fig. 4C and D), which further confirmed
that HBV infection may increase the pathogenesis of fatty liver
disease. Alternatively, increased ROS production in hepatocytes
expressing HBx may promote lipid droplet accumulation, and this
effect was reversed by treating the cells with a ROS scavenger in a
time-dependent manner (Fig. 4C and
D). Therefore, the present findings indicated that, HBV, or at
least the HBx protein, may not be a protective factor against
NAFLD; however, this requires further validation.
Discussion
Viral infections, such as chronic HBV or HCV, can
cause a multitude of liver diseases, including hepatic fibrosis,
cirrhosis and liver cancer (27,28).
Unlike HCV, conflicting conclusions regarding the association
between HBV infection and fatty liver disease have been reported in
different populations (29–31).
The prevalence of NAFLD is 28.3% in Hong Kong (age range, 18–70)
(31) and 43.9% in Taiwan (mean
age, 51.9 years), whereas in patients with HBV, the prevalence of
NAFLD decreases to 13.5 and 38.9% in Hong Kong and Taiwan,
respectively (31,32). However, in Hong Kong, a significant
difference in the prevalence of NAFLD between HBV-infected
individuals and controls was detected only in the population aged
between 40 and 59 years. Conversely, whereas HBV infection has been
reported to be negatively associated with NAFLD in patients >40
years, the same association is not true in younger patients
(32). Despite this contradiction,
a recent large cohort study in Korea supported the hypothesis that
HBV is a protective factor against NAFLD (9). In the present study, female patients
with fatty liver disease were recruited, in order to study the
association between HBV infection and fatty liver disease. The
majority of women in China do not consume alcohol, and few have a
history of daily consumption, due to traditional Chinese culture;
therefore, few female patients with fatty liver disease are
diagnosed with alcoholic fatty liver disease. The prevalence of
female fatty liver disease in the present study (24.8% in 50–59
year old age group) is similar to that reported for Chinese female
patients with NAFLD in the same age range (22.8%) (33). Studying the association between
female patients with fatty liver disease and HBV infection may
provide important information regarding the effects of HBV
infection on NAFLD. Since age has been demonstrated to be a major
contributor to HBV-associated NAFLD (31,32),
logistic regression analysis was applied to adjust for the risks
associated with age and HBV infection in fatty liver disease.
Although the prevalence of fatty liver disease was lower in
patients with HBV (10.5%) compared with in HBV-free patients (17%),
this finding was not significant. These findings suggested that HBV
may not be a protective factor against fatty liver disease or NAFLD
in mainland China.
The present epidemiological data is preliminary and
limitations exist. Factors such as anti-viral usage, the stage of
HBV infection and infection history in HBV-infected patients were
not considered. Furthermore, information regarding alcohol
consumption and drug administration in the enrolled subjects were
not included. However, Joo et al (9) reported similar limitations, but the
large sample size used in their study helped to overcome them. In
the present study, the effects of HBx on lipid levels in the two
cell lines were investigated to further evaluate the effects of
HBV, specifically HBx, on fatty liver disease. The present results
demonstrated that HBx increased the formation of lipid droplets
in vitro, a key indicator of NAFLD, suggesting that HBx may
serve as an inducer of NAFLD. HBx is believed to act as a promoter
in hepatic lipid accumulation through HBx/fatty acid-binding
protein 1/hepatocyte nuclear factor 3-β (HNF3β),
CCAAT-enhancer-binding protein α and peroxisome
proliferator-activated receptor α axis in HBV-induced NAFLD
(34). In addition, HBx can
directly interact with liver X receptor (LXR)α to upregulate the
levels of sterol regulatory element-binding protein 1 (SREBP1c) to
support lipogenesis, thereby causing fatty liver disease (35). In the present study, it was
demonstrated that HBx may promote lipid droplet formation via
increased mitochondrial ROS levels, whereas the addition of a ROS
scavenger inhibited lipid droplet formation. The underlying
mechanism of ROS in lipid droplet formation and NAFLD has been
widely studied (36). In addition
to the HBx/LXRα/SREBP1 axis, an HBx/mitochondrial ROS/c-Jun
N-terminal kinase axis can also promote SREBP1c to support the
formation of lipid droplet formation (37). Although increased generation of ROS
production has been implicated in damaged mitochondria, increased
mitochondrial ROS are also detected in cells with elevated aerobic
metabolism (38). Consistently,
our finding of increased mitochondrial function and ROS production
in HBx transfected cells imply a possibility of increased electron
leakage from over-activated electron transfer.
In conclusion, unlike previous studies, the present
study demonstrated that HBV infection may not act as a protective
factor against NAFLD in the Chinese population. Conversely, it was
revealed that HBV may serve a role in the development of NAFLD
in vitro via increased production of mitochondrial ROS;
however, this requires further validation in future studies.
Acknowledgements
The authors thank Nianqi Hu (Wenzhou Medical
University) for her assistance on data analysis of this
manuscript.
Funding
The present study was supported by grants from the
Chinese National Science Foundation (grant no. 31370795).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
HZ conceived the study, performed research and wrote
the manuscript; BW and WL performed research, analyzed data and
wrote the manuscript; BW conducted the statistical analysis; BW, HZ
and HF analyzed data and participated in the critical
discussion.
Ethics approval and consent to
participate
Informed consent was obtained from all subjects, and
this study was approved by the Ethical Committee of Wenzhou Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Benedict M and Zhang X: Non-alcoholic
fatty liver disease: An expanded review. World J Hepatol.
9:715–732. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gusdon AM, Song KX and Qu S: Nonalcoholic
Fatty liver disease: Pathogenesis and therapeutics from a
mitochondria-centric perspective. Oxid Med Cell Longev.
2014:6370272014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Besse-Patin A and Estall JL: An intimate
relationship between ROS and insulin signalling: Implications for
antioxidant treatment of fatty liver disease. Int J Cell Biol.
2014:5191532014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mello T, Materozzi M and Galli A: PPARs
and mitochondrial metabolism: From NAFLD to HCC. PPAR Res.
2016:74032302016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bravo E, Palleschi S, Rossi B, Napolitano
M, Tiano L, D'Amore E and Botham KM: Coenzyme Q metabolism is
disturbed in high fat diet-induced non-alcoholic fatty liver
disease in rats. Int J Mol Sci. 13:1644–1657. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schweitzer A, Horn J, Mikolajczyk RT,
Krause G and Ott JJ: Estimations of worldwide prevalence of chronic
hepatitis B virus infection: A systematic review of data published
between 1965 and 2013. Lancet. 386:1546–1555. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hsu A, Lai CL and Yuen MF: Update on the
risk of hepatocellular carcinoma in chronic hepatitis B virus
infection. Curr Hepat Rep. 10:106–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tu T, Budzinska MA, Shackel NA and Urban
S: HBV DNA integration: Molecular Mechanisms and Clinical
Implications. Viruses. 9(pii): E752017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Joo EJ, Chang Y, Yeom JS and Ryu S:
Hepatitis B virus infection and decreased risk of nonalcoholic
fatty liver disease: A cohort study. Hepatology. 65:828–835. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He J, Zeng YL, Li W, Guo EE, Li JL, Kang Y
and Shang J: Clinical study of non-alcoholic fatty liver disease
and its combined the chronic HBV infection. Zhonghua Gan Zang Bing
Za Zhi. 25:618–622. 2017.(In Chinese). PubMed/NCBI
|
|
11
|
Lin HJ, Ku KL, Lin IH and Yeh CC:
Naringenin attenuates hepatitis B virus X protein-induced hepatic
steatosis. BMC Complement Altern Med. 17:5052017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bouchard MJ and Schneider RJ: The
enigmatic X gene of hepatitis B virus. J Virol. 78:12725–12734.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen HY, Tang NH, Lin N, Chen ZX and Wang
XZ: Hepatitis B virus X protein induces apoptosis and cell cycle
deregulation through interfering with DNA repair and checkpoint
responses. Hepatol Res. 38:174–182. 2008.PubMed/NCBI
|
|
14
|
Na TY, Shin YK, Roh KJ, Kang SA, Hong I,
Oh SJ, Seong JK, Park CK, Choi YL and Lee MO: Liver X receptor
mediates hepatitis B virus X protein-induced lipogenesis in
hepatitis B virus-associated hepatocellular carcinoma. Hepatology.
49:1122–1131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim KH, Shin HJ, Kim K, Choi HM, Rhee SH,
Moon HB, Kim HH, Yang US, Yu DY and Cheong J: Hepatitis B virus X
protein induces hepatic steatosis via transcriptional activation of
SREBP1 and PPARgamma. Gastroenterology. 132:1955–1967. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim SJ, Khan M, Quan J, Till A, Subramani
S and Siddiqui A: Hepatitis B virus disrupts mitochondrial
dynamics: Induces fission and mitophagy to attenuate apoptosis.
PLoS Pathog. 9:e10037222013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Valkov I, Ivanova R, Alexiev A, Antonov K
and Mateva L: Association of serum lipids with hepatic steatosis,
stage of liver fibrosis and viral load in chronic Hepatitis C. J
Clin Diagn Res. 11:OC15–OC20. 2017.PubMed/NCBI
|
|
18
|
Chen J, Zhang W, Lin J, Wang F, Wu M, Chen
C, Zheng Y, Peng X, Li J and Yuan Z: An efficient antiviral
strategy for targeting hepatitis B virus genome using transcription
activator-like effector nucleases. Mol Ther. 22:303–311. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li CH, Xu F, Chow S, Feng L, Yin D, Ng TB
and Chen Y: Hepatitis B virus X protein promotes hepatocellular
carcinoma transformation through interleukin-6 activation of
microRNA-21 expression. Eur J Cancer. 50:2560–2569. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong S, Peng Y, Jiang P, Wang M, Fan M,
Wang X, Zhou H, Li H, Yan Q, Huang T and Guan MX: A
deafness-associated tRNAHis mutation alters the mitochondrial
function, ROS production and membrane potential. Nucleic Acids Res.
42:8039–8048. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park JS, Sharma LK, Li H, Xiang R,
Holstein D, Wu J, Lechleiter J, Naylor SL, Deng JJ, Lu J and Bai Y:
A heteroplasmic, not homoplasmic, mitochondrial DNA mutation
promotes tumorigenesis via alteration in reactive oxygen species
generation and apoptosis. Hum Mol Genet. 18:1578–1589. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sharma LK, Fang H, Liu J, Vartak R, Deng J
and Bai Y: Mitochondrial respiratory complex I dysfunction promotes
tumorigenesis through ROS alteration and AKT activation. Hum Mol
Genet. 20:4605–4616. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shen L, Fan JG, Shao Y, Zeng MD, Wang JR,
Luo GH, Li JQ and Chen SY: Prevalence of nonalcoholic fatty liver
among administrative officers in Shanghai: An epidemiological
survey. World J Gastroenterol. 9:1106–1110. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou YJ, Li YY, Nie YQ, Ma JX, Lu LG, Shi
SL, Chen MH and Hu PJ: Prevalence of fatty liver disease and its
risk factors in the population of South China. World J
Gastroenterol. 13:6419–6424. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim HY, Cho HK, Yoo SK and Cheong JH:
Hepatic STAMP2 decreases hepatitis B virus X protein-associated
metabolic deregulation. Exp Mol Med. 44:622–632. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Satapati S, Kucejova B, Duarte JA,
Fletcher JA, Reynolds L, Sunny NE, He T, Nair LA, Livingston KA, Fu
X, et al: Mitochondrial metabolism mediates oxidative stress and
inflammation in fatty liver. J Clin Invest. 125:4447–4462. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ringelhan M, O'Connor T, Protzer U and
Heikenwalder M: The direct and indirect roles of HBV in liver
cancer: Prospective markers for HCC screening and potential
therapeutic targets. J Pathol. 235:355–367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ryerson AB, Eheman CR, Altekruse SF, Ward
JW, Jemal A, Sherman RL, Henley SJ, Holtzman D, Lake A, Noone AM,
et al: Annual report to the nation on the status of cancer,
1975-2012, featuring the increasing incidence of liver cancer.
Cancer. 122:1312–1337. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar R and Boon-Bee Goh G: Chronic
hepatitis B and fatty liver: Issues in clinical management. Clin
Res Hepatol Gastroenterol. 40:755–759. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang CC and Kao JH: Hepatitis B virus
infection and decreased risk of nonalcoholic fatty liver disease: A
cohort study. Hepatology. 66:6812017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wong VW, Wong GL, Chu WC, Chim AM, Ong A,
Yeung DK, Yiu KK, Chu SH, Chan HY, Woo J, et al: Hepatitis B virus
infection and fatty liver in the general population. J Hepatol.
56:533–540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng YL, Wang YJ, Kao WY, Chen PH, Huo
TI, Huang YH, Lan KH, Su CW, Chan WL, Lin HC, et al: Inverse
association between hepatitis B virus infection and fatty liver
disease: A large-scale study in populations seeking for check-up.
PLoS One. 8:e720492013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Z, Xu M, Hu Z and Shrestha UK:
Prevalence of nonalcoholic fatty liver disease and its metabolic
risk factors in women of different ages and body mass index.
Menopause. 22:667–673. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu YL, Peng XE, Zhu YB, Yan XL, Chen WN
and Lin X: Hepatitis B Virus X protein induces hepatic steatosis by
enhancing the expression of liver fatty acid binding protein. J
Virol. 90:1729–1740. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim K, Kim KH, Kim HH and Cheong J:
Hepatitis B virus X protein induces lipogenic transcription factor
SREBP1 and fatty acid synthase through the activation of nuclear
receptor LXRalpha. Biochem J. 416:219–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nassir F and Ibdah JA: Role of
mitochondria in nonalcoholic fatty liver disease. Int J Mol Sci.
15:8713–8742. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu L, Zhang K, Sandoval H, Yamamoto S,
Jaiswal M, Sanz E, Li Z, Hui J, Graham BH, Quintana A and Bellen
HJ: Glial lipid droplets and ROS induced by mitochondrial defects
promote neurodegeneration. Cell. 160:177–190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Weinberg F, Hamanaka R, Wheaton WW,
Weinberg S, Joseph J, Lopez M, Kalyanaraman B, Mutlu GM, Budinger
GR and Chandel NS: Mitochondrial metabolism and ROS generation are
essential for Kras-mediated tumorigenicity. Proc Natl Acad Sci USA.
107:8788–8793. 2010. View Article : Google Scholar : PubMed/NCBI
|