Introduction
Chronic lymphocytic leukemia (CLL) is a clonal
proliferative tumor of mature B lymphocytes characterized by the
accumulation of lymphocytes in the peripheral blood, bone marrow,
lymph nodes and spleen (1). It is
the most common type of leukemia in western populations, causing
~5,000 mortalities in the United States of America annually
(2). Unfortunately, the exact
molecular mechanism underlying the development of CLL remains
unclear, which limits early diagnosis and timely treatment.
Therefore, exploring the molecular mechanism of CLL progression and
its biomarkers to improve the likelihood of early diagnosis and
treatment have become the focus in CLL research. Increasing
evidence suggests that numerous genes, miRNAs and cellular pathways
are involved in the occurrence and progression of CLL (3,4).
In recent years, the rapid development of
computer-based technologies, such as molecular dynamics simulation,
has gradually revealed novel molecular mechanisms of the disease
(5,6). Through the combination of Gene
Expression Omnibus (GEO) chip, The Cancer Genome Atlas data mining
(cancergenome.nih.gov/) and
computer-based analysis techniques, a series of highly specific and
sensitive markers have been reported (7–9).
Gene chips are widely used as a genetic testing tool as they can be
used to detect rapid gene expression of a sample at a certain time
point and are particularly suitable for the screening of
differentially expressed genes (10). MicroRNAs (miRNAs/miRs) are
endogenous non-coding small RNAs which have several important
regulatory roles in cells (11).
Computer-based analysis techniques have demonstrated a mutation of
a miRNA binding site that is associated with cancer and revealed
its mechanism of gene regulation (12).
In the present study, two original gene microarray
datasets (GSE22529 and GSE39411) and one original miRNA microarray
dataset (GSE622137) were downloaded from the National Center for
Biotechnology Information-Gene Expression Omnibus database
(NCBI-GEO). The raw gene microarray datasets were initially
processed online using the Gene-Cloud of Biotechnology Information
(GCBI) software to screen for differentially expressed genes (DEGs)
in CLL and normal samples. In addition, the Gene Ontology (GO)
Consortium and the Kyoto Encyclopedia of Genes and Genomes (KEGG),
in conjunction with the Database for Annotation, Visualization, and
Integrated Discovery (DAVID) software program were used to analyze
the pathway enrichment of DEGs. The STRING online database and
Cytoscape software were then used to develop the protein-protein
interaction (PPI) network of the DEGs and for modular analysis to
identify CLL central genes. In addition, GEO2R analysis of the
miRNA dataset was employed to obtain the differentially expressed
miRNAs (DEMs). Cytoscape software was used to analyze the
association between the predicted target mRNAs of DEMs and DEGs,
which helped to define the regulatory network of miRNA-mRNA in
relation to the development of CLL. Our findings suggest that data
mining and integration may be useful methods for predicting and
understanding the mechanisms of CLL disease development and
progression. Ultimately, the early diagnosis and personalized
treatment of CLL may be facilitated by identifying new biomarkers
(3).
Materials and methods
Microarray data
Data of gene chips GSE22529 (13), GSE39411 (14) and GSE62137 (15) were obtained from the GEO database
(www.ncbi.nlm.nih.gov/geo). GSE22529 was
based on Agilent GPL96 (Agilent Technologies, Inc., Santa Clara,
CA, USA; [HG-U133A] Affymetrix Human Genome U133A Array,
Affymetrix; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
GPL97 [(HG-U133B) Affymetrix Human Genome U133B Array, Affymetrix;
Thermo Fisher Scientific, Inc.)]. To reduce the offset, based on
the GCBI online laboratory (www.gcbi.com.cn/gclib/html/index), the GPL96
annotation data set as the research object, was screened. The
GSE22529 dataset included 52 samples, containing 41 CLL samples and
11 normal samples. The GSE39411 data were based on GPL570
[(HG-U133_Plus_2) Affymetrix Human Genome U133 Plus 2.0 Array;
Affymetrix, Thermo Fisher Scientific, Inc.], consisting of 41 CLL
samples and 11 normal samples. The GSE62137 data were based on
GPL14767 [(Agilent-021827 Human miRNA Microarray G4470C)],
consisting of 15 CLL samples and 3 normal samples.
Identification of DEGs and DEMs
To identify DEGs in the CLL and normal samples, a
web-based online tool, GCBI, was utilized. GCBI is an interactive
online tool that allows users to compare two or more groups of
samples in a GEO series and analyzes the majority of GEO series
with gene symbols. DEGs were screened according to P<0.05 and
|log fold change (FC)|>1.5. DEMs were obtained by analyzing
miRNA expression profiles using the GEO2R tool (https://www.ncbi.nlm.nih.gov/geo/geo2r/)
and screening according to the |log FC|>2 and P<0.05. In
addition, DEG heat maps from datasets GSE22529 and GSE39411, were
created using Morpheus online software (software.broadinstitute.org/morpheus).
GO and pathway enrichment analysis of
DEGs
DAVID version 6.8 (david.abcc.ncifcrf.gov/), an open source platform, was
utilized to determine the association among target molecules
(16). To gain insight into the
biological functions of these misaligned genes, GO and (KEGG)
pathway enrichment analyses were performed on the DEGs based on
DAVID, and P<0.05 was set as the cut-off criterion. The
molecular function, biological processes, cell component and KEGG
pathways of these DEGs and target genes were investigated using
DAVID.
Protein-protein interaction (PPI)
network and module analysis
As a search tool for the retrieval of interacting
genes, STRING version 10.5 (string-db.org)
was used to assess PPI information (17). To further examine the potential
association between these DEGs, STRING was used to characterize the
PPI network, and a confidence score >0.4 was set as the cut-off
criterion. Cytoscape_v3.5.1 software was used to visualize the
resulting PPI network (18). The
Molecular Complex Detection (MCODE version 1.4.2) application
(http://apps.cytoscape.org/apps/mcode)
was used to select significant modules from the PPI network in
Cytoscape with MCODE scores >5 and number of nodes >10 as the
standard. Furthermore, a topological analysis of the PPI network
was performed and the key genes were screened for further
biological analysis of the node degree of these DEGs.
Target mRNA prediction of DEMs and
construction of miRNA-mRNA regulatory networks
The target mRNAs of DEMs were predicted using
TargetScan (www.targetscan.org/), miRDB (www.mirdb.org/miRDB/) (19), and miRTarBase version 7 (mirtarbase.mbc.nctu.edu.tw/) (20) online analysis tools. To further
improve the reliability of bioinformatics analysis, overlapping
target genes between DEGs and predicted target mRNAs were
identified using a Venn diagram. The miRNA-mRNA networks were
visualized using the Cytoscape platform software.
Results
Identification of DEGs and DEMs
NCBI-GEO is a free database containing
microarray/gene profiles and next-generation sequences. Data for
gene expression in CLL patients and normal samples was obtained
from GSE22529 and GSE39411 along with miRNA expression data from
GSE62137. The original gene data files were uploaded to the GCBI
online laboratory to screen for the DEGs in CLL and normal samples.
Using P<0.05 and |log FC|>1.5 as cut-off criteria, 781 and
1,607 DEGs were extracted from the expression profile datasets
GSE22529 and GSE39411, respectively. Following integrated
bioinformatics analysis, a total of 488 consistently expressed
genes were identified from the two profile datasets, including 204
upregulated and 284 downregulated genes in the CLL samples compared
with the normal population. Using a novel web-based tool, Morpheus,
heat maps (illustrating the top 100 upregulated and top 100
downregulated genes) of DEGs were constructed from the GSE22529 and
GSE39411 datasets (Fig. 1A and B).
In addition, the miRNA profile dataset, GSE62137, was analyzed to
screen for DEMs in CLL using the GEO2R tool. According to the
cut-off criteria (P<0.05 and |logFC|>2.0), a total of 34
differentially expressed miRNAs were identified from this
microarray dataset, including 29 upregulated and 5 downregulated
miRNAs (Table I). Among them,
miR-144 and miR-181a were the most downregulated miRNAs in the
GSE62137 dataset, whereas miR-582 was the most upregulated. The 488
consistently expressed genes and 34 differentially expressed miRNAs
were used for further functional analysis.
 | Table I.Main dysregulated miRNAs in chronic
lymphocytic leukemia. |
Table I.
Main dysregulated miRNAs in chronic
lymphocytic leukemia.
| miRNA | LogFC | P-value |
|---|
| hsa-miR-582-5p | 8.23 |
2.78×10−21 |
| hsa-miR-451 | −6.25 |
1.91×10−5 |
|
hsa-miR-548c-3p | 3.65 |
9.27×10−5 |
|
hsa-miR-181c-3p | 3.22 |
1.03×10−4 |
| hsa-miR-181a | −8.66 |
1.12×10−4 |
| hsa-miR-95 | 4.34 |
1.16×10−4 |
| hsa-miR-132 | 6.46 |
2.84×10−4 |
| hsa-miR-144 | −8.52 |
3.25×10−4 |
| hsa-miR-486-5p | −5.57 |
3.88×10−4 |
| hsa-miR-885-3p | 3.23 |
5.86×10−3 |
|
hsa-miR-199a-3p | 5.76 |
7.00×10−3 |
| hsa-miR-126 | 6.45 |
9.25×10−3 |
| hsa-miR-145 | 2.59 |
1.98×10−2 |
| hsa-miR-136 | 2.78 |
1.98×10−2 |
| hsa-miR-326 | 2.02 |
1.98×10−2 |
| hsa-miR-346 | 2.03 |
1.98×10−2 |
| hsa-miR-376c | 2.96 |
1.98×10−2 |
| hsa-miR-139-5p | 2.14 |
1.98×10−2 |
| hsa-miR-410 | 2.33 |
1.98×10−2 |
| hsa-miR-339-5p | 2.05 |
1.98×10−2 |
| hsa-miR-584 | 2.44 |
1.98×10−2 |
| hsa-miR-1 | 2.24 |
1.98×10−2 |
| hsa-miR-495 | 2.11 |
1.98×10−2 |
| hsa-miR-376a | 2.89 |
1.98×10−2 |
| hsa-miR-377 | 2.76 |
1.98×10−2 |
| hsa-miR-26b-3p | 2.1 |
1.98×10−2 |
| hsa-miR-181c | 2.09 |
1.98×10−2 |
| hsa-miR-381 | 2.37 |
1.98×10−2 |
| hsa-miR-296-3p | 2.16 |
1.98×10−2 |
| hsa-miR-409-3p | 2.36 |
1.98×10−2 |
|
hsa-miR-199a-5p | 3.42 |
1.98×10−2 |
| hsa-miR-126-5p | 3.11 |
1.98×10−2 |
|
hsa-miR-664a-5p | −5.32 |
1.98×10−2 |
| hsa-miR-501-5p | 4.2 |
1.98×10−2 |
GO function and KEGG pathway
enrichment analysis of the DEGs
To gain a deeper understanding of the selected DEGs,
GO and KEGG pathway analyses were performed using DAVID. GO
analysis demonstrated that upregulated and downregulated DEGs were
particularly abundant in the ‘molecular function’, ‘biological
processes’ and ‘cell component classification’ (Table II). As demonstrated in this Table,
in the ‘molecular function group’, upregulated genes were mainly
enriched in ‘protein binding’, ‘protein kinase binding’ and
‘non-membrane spanning protein tyrosine kinase activity’, while the
downregulated DEGs were enriched in various combinations, such as
‘antigen binding’, ‘immunoglobulin receptor binding’ and ‘cell
adhesion molecule binding’. In the ‘biological processes’ group,
upregulated DEGs were mainly associated with ‘intracellular
signaling transduction’, the ‘transmembrane receptor protein
tyrosine kinase signaling pathway’ and ‘positive regulation of
granulocyte differentiation’, and downregulated DEGs were
significantly enriched in ‘immune response’ and the ‘B cell
receptor signaling pathway’. In addition, GO cell component
analysis demonstrated that the upregulated DEGs were enriched in
‘cytosol and plasma membrane’, and downregulated DEGs were enriched
in ‘cell surface’, ‘extracellular exosome’ and ‘cell-cell adherent
junction’. These results demonstrated that most DEGs are
significantly enriched in ‘binding’, ‘adhesion’ and ‘cell
cycle’.
| Table II.Gene ontology analysis of
differentially expressed genes associated with chronic lymphocytic
leukemiaa. |
Table II.
Gene ontology analysis of
differentially expressed genes associated with chronic lymphocytic
leukemiaa.
| Category | Term | Count | P-value |
|---|
| Upregulated |
|
|
|
|
GOTERM_BP_DIRECT |
GO:0035556~intracellular signal
transduction | 16 |
2.64×10−5 |
|
|
GO:0007169~transmembrane receptor protein
tyrosine kinase signaling pathway | 7 |
5.49×10−4 |
|
| GO:0008283~cell
proliferation | 12 |
1.89×10−3 |
|
| GO:0030854~positive
regulation of granulocyte differentiation | 3 |
2.27×10−3 |
|
|
GO:1901385~regulation of voltage-gated
calcium channel activity | 3 |
2.27×10−3 |
|
GOTERM_MF | GO:0005515~protein
binding | 122 |
6.86×10−6 |
| _DIRECT | GO:0019901~protein
kinase binding | 14 |
1.72×10−4 |
|
| GO:0005524~ATP
binding | 31 |
4.04×10−4 |
|
|
GO:0004715~non-membrane spanning protein
tyrosine kinase activity | 5 |
1.35×10−3 |
|
| GO:0042169~SH2
domain binding | 4 |
3.39×10−3 |
|
GOTERM_CC | GO:0005886~plasma
membrane | 65 |
1.81×10−4 |
| _DIRECT |
GO:0005829~cytosol | 53 |
7.60×10−4 |
|
| GO:0009897~external
side of plasma membrane | 9 |
1.63×10−3 |
|
| GO:0045121~membrane
raft | 8 |
5.46×10−3 |
|
|
GO:0030175~filopodium | 5 |
6.11×10−3 |
| Downregulated |
GO:0006955~immune | 31 |
1.58×10−12 |
|
GOTERM_BP | response |
|
|
| _DIRECT |
GO:0050776~regulation of immune
response | 21 |
2.84×10−12 |
|
|
GO:0006958~complement activation,
classical pathway | 14 |
2.97×10−9 |
|
|
GO:0006911~phagocytosis, engulfment | 8 |
7.61×10−7 |
|
| GO:0050853~B cell
receptor signaling pathway | 9 |
1.36×10−6 |
|
GOTERM_MF | GO:0003823~antigen
binding | 15 |
3.62×10−10 |
| _DIRECT | GO:0042803~protein
homodimerization activity | 29 |
4.07×10−6 |
|
|
GO:0034987~immunoglobulin receptor
binding | 5 |
5.43×10−4 |
|
| GO:0004872~receptor
activity | 11 |
1.51×10−3 |
|
| GO:0005515~protein
binding | 154 |
2.01×10−3 |
|
GOTERM_CC |
GO:0070062~extracellular exosome | 97 |
8.36×10−18 |
| _DIRECT | GO:0005886~plasma
membrane | 113 |
1.27×10−13 |
|
| GO:0009986~cell
surface | 24 |
3.43×10−6 |
|
| GO:0072562~blood
microparticle | 12 |
1.19×10−5 |
|
| GO:0005887~integral
component of plasma membrane | 37 |
5.02×10−4 |
The results of the enrichment of DEG function and
signaling pathways (Table III)
demonstrated that the downregulated genes were mainly associated
with the complex biological behavior of CLL. Using P<0.05 as a
cut-off criterion, 17 KEGG pathways were over-represented in
downregulated DEGs, including ‘hematopoietic cell lineage’,
‘transcriptional dysregulation in cancer’ and ‘pathways in cancer’,
while the four KEGG pathways involved in the upregulated DEGs were
enriched in multiple signaling pathways, the ‘cyclic adenosine
monophosphate (cAMP) signaling pathway’ and the ‘mitogen-activated
protein kinase (MAPK) signaling pathway’. These most significantly
enriched GO terms and KEGG pathways demonstrated the interactions
of DEGs at the functional level.
| Table III.Kyoto Encyclopedia of Genes and
Genomes pathway analysis of differentially expressed genes
associated with chronic lymphocytic leukemiaa. |
Table III.
Kyoto Encyclopedia of Genes and
Genomes pathway analysis of differentially expressed genes
associated with chronic lymphocytic leukemiaa.
| Category | Term | Count | P-value | Genes |
|---|
| Upregulated
DEGs | hsa04261:Adrenergic
signaling in cardiomyocytes | 7 |
9.44×10−3 | ADRB2, ATP2B4,
BCL2, PPP2R5C, CREB3L2, CACNB2, RAPGEF3 |
|
|
hsa00561:Glycerolipid metabolism | 4 |
3.52×10−2 | DGKA, LPL, AGPAT5,
ALDH3A2 |
|
| hsa04024:cAMP
signaling pathway | 7 |
3.63×10−2 | ADRB2, ATP2B4,
PDE4A, RRAS2, CREB3L2, RAPGEF3, MYL9 |
|
| hsa04010:MAPK
signaling pathway | 7 |
3.89×10−2 | PTPN7, MAP3K5,
RASGRF1, RRAS2, CACNB2, MYC, RASA1, CDC25B |
| Downregulated
DEGs |
hsa05144:Malaria | 8 |
3.32×10−5 | KLRB1, GYPC, CR1,
KLRC4-KLRK1, ITGB2, HBA1, HBB, TGFB |
|
|
hsa04640:Hematopoietic cell lineage | 9 |
1.90×10−4 | CR1, CR2, CD44,
CD1C, CD22, ITGA4, IL7R, ITGAM, CD1D |
|
|
hsa05202:Transcriptional dysregulation in
cancer | 12 |
3.22×10−4 | JUP, CD86, CEBPB,
ID2, REL, LMO2, BCL2A1, AFF1, ZBTB16, MYC, ITGAM, KLF3 |
|
|
hsa05140:Leishmaniasis | 8 |
3.64×10−4 | FOS, CR1, NFKBIA,
ITGB2, ITGA4, ITGAM, TGFB1, IFNGR1 |
|
| hsa05323:Rheumatoid
arthritis | 7 |
6.30×10−3 | FOS, CD86, CCL3,
ITGB2, IL15, LTB, TGFB1 |
PPI network construction and module
selection
Using the STRING online database and Cytoscape
software, a total of 354 DEGs were filtered into the complex DEG
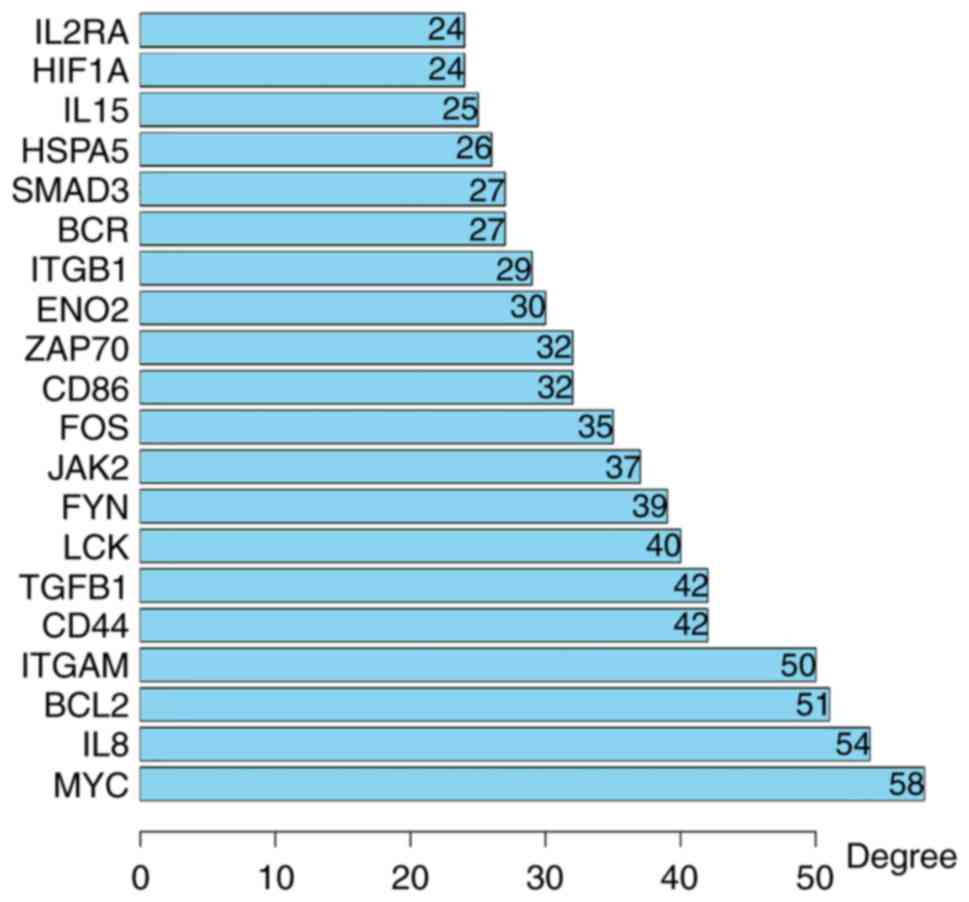
PPI network, containing 354 nodes and 1,392 edges (Fig. 2); 134 of the 488 DEGs did not fall
into the DEG PPI network. Among the 354 nodes, 12 differentially
expressed genes with a high degree of node were screened from the
PPI network, and the most significant 12 node degree genes were
MYC, interleukin (IL)8, B-cell lymphoma 2
(BCL2), integrin subunit alpha M (ITGAM),
transforming growth factor (TGF)B1, cluster of
differentiation (CD)44, LCK, FYN, Janus kinase (JAK)2,
FOS, CD86 and z chain of T cell receptor associated protein
kinase (ZAP)-70 (Fig. 3).
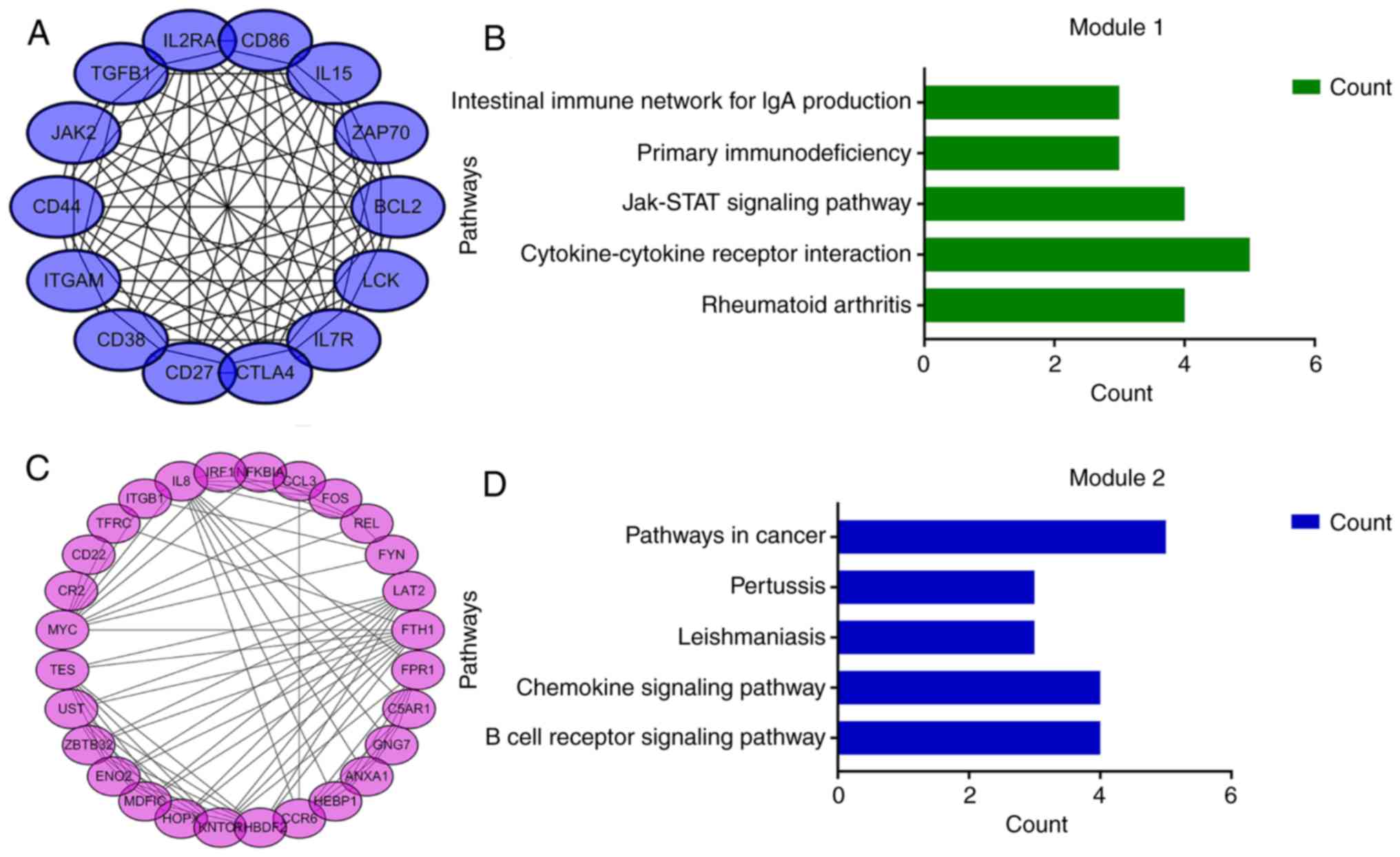
According to their degree of importance, 2 important modules from
the PPI network complex were selected for further analysis based on
Cytoscape MCODE. Functional and pathway enrichment analysis of the
genes in these two modules were performed using DAVID. The results
revealed that Module 1 consisted of 14 nodes and 82 edges (Fig. 4A and B), which were mainly
associated with cytokine-cytokine receptor interaction, the
JAK/signal transducer and activator of transcription (Jak-STAT)
signaling pathway, and that Module 2 consisted of 28 nodes and 101
edges (Fig. 4C and D), which were
enriched in the B cell receptor signaling pathway and the chemokine
signaling pathway. The results of the module analysis revealed that
eight of the 12 DEGs with a high degree were clustered in Module 1,
and the other four genes were in Module 2. In order to further
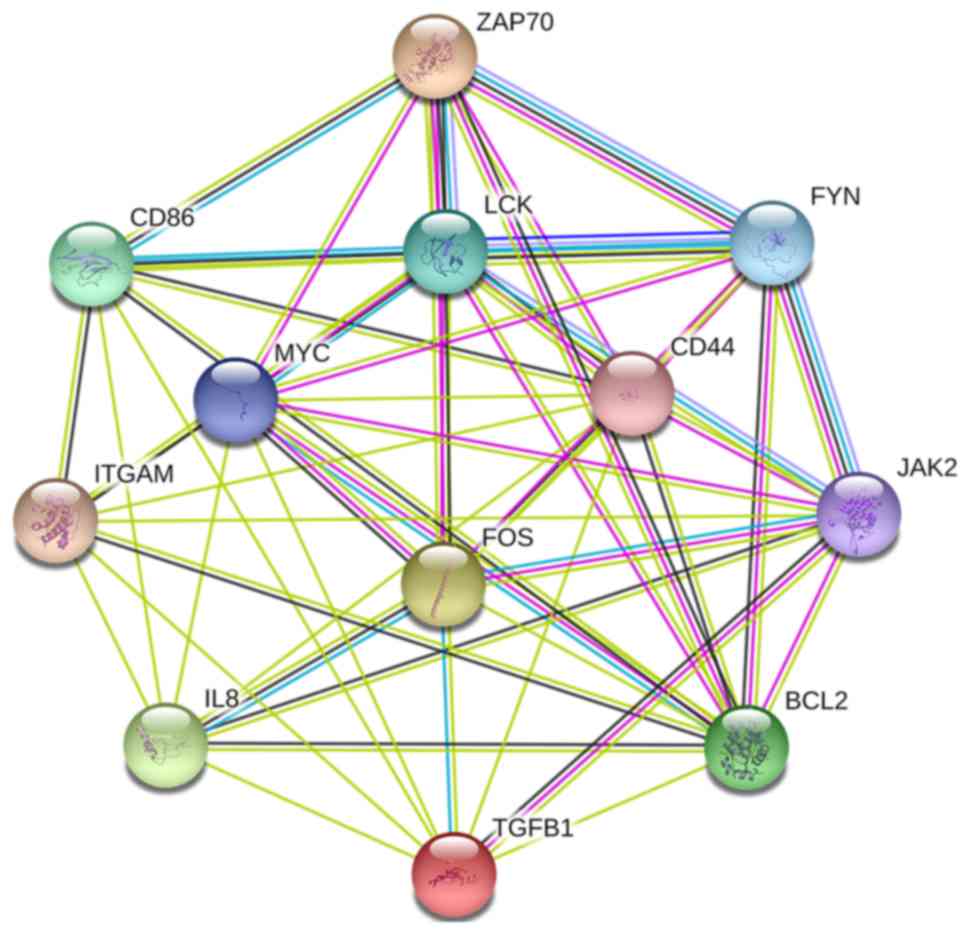
understand the interaction of these 12 key genes, the PPI network
of these genes was constructed by STRING (Fig. 5). These results suggested that
these central genes are closely associated with CLL and interact to
promote the development of disease, which may suggest novel
therapeutic approaches against CLL.
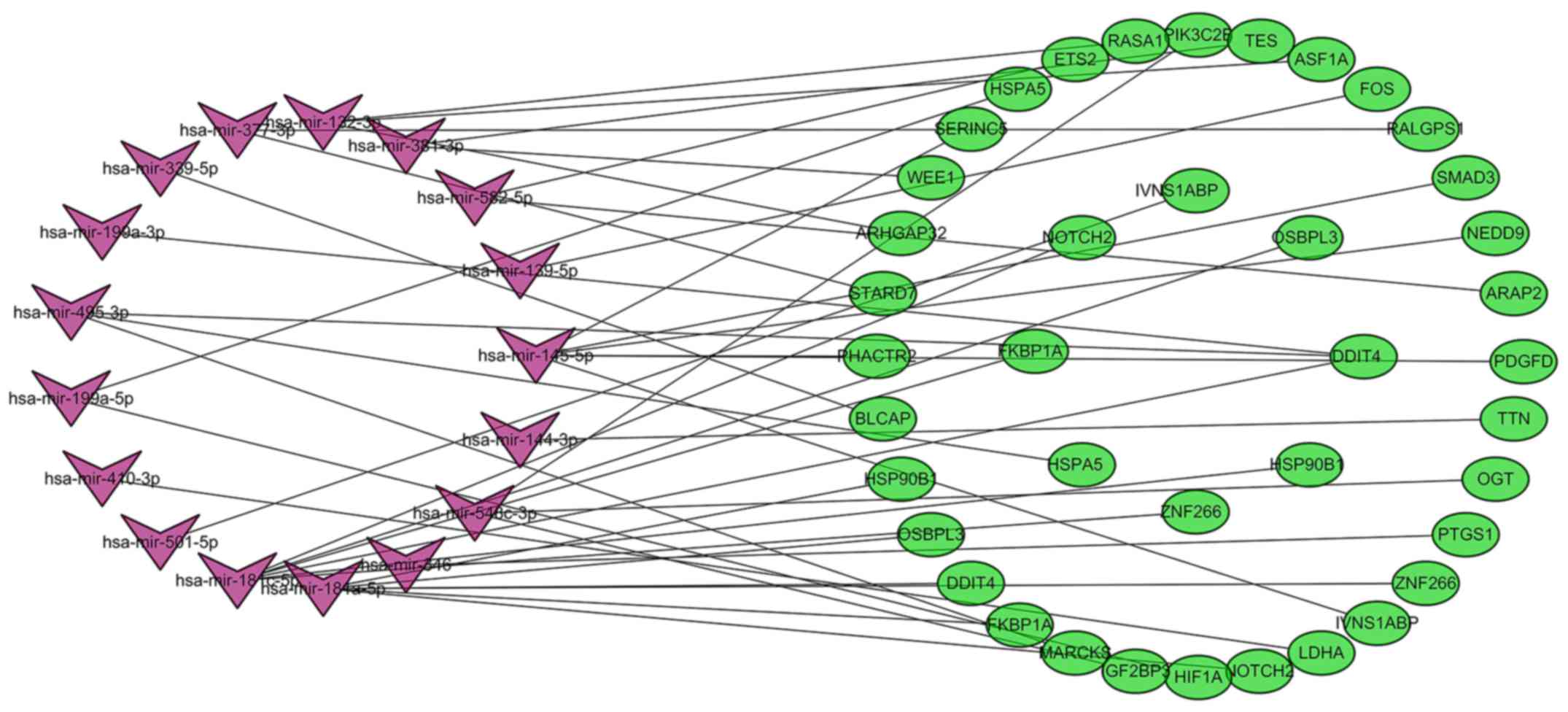
Construction of miRNA-mRNA regulatory
network in the pathogenesis of CLL
The target mRNAs of DEMs were predicted using the
TargetScan, miRDB and miRTarBase online analysis tools. A total of
621 target mRNAs were predicted, and 32 target mRNAs associated
with CLL were screened further by analyzing the association between
them and the corresponding DEGs. In the present study, miR-582 was
the most significantly upregulated miRNA and was predicted to
target the ETS2 and ArfGAP with RhoGAP domain, ankyrin
repeat and PH domain (ARAP)2 genes. MiR-144 and miR-181a
were the most significantly downregulated miRNAs. MiR-144 was
predicted to target the titin (TTN) gene, and miR-188a was
predicted to target the zinc finger (ZNF)266, NOTCH2, FK506
binding protein (FKBP)1A, DNA damage inducible transcript
(DDIT)4, oxysterol binding protein like (OSBPL)3 and
heat shock protein 90 β family member (HSP90B)1 genes. The
results also demonstrated that miR-181c and miR-145 targeted six
different differentially expressed genes, while miR-584, miR-132
and miR-548c-3p targeted 3 different genes. In addition,
DDIT4 could be regulated by hsa-miR-181a, miR-181c,
miR-199a-3p and miR-495, while ZNF266, influenza virus NS1A
binding protein (IVNS1ABP), OSBPL3, HSP90B1, HSPA5 and
NOTCH2 could be regulated by two different miRNAs. A total
of 32 target mRNAs corresponded to 17 differentially expressed
miRNAs. To better understand the pathogenesis of CLL, miRNA-mRNA
regulatory networks (Fig. 6) were
constructed and analyzed, and a total of 42 miRNA-mRNA pairs were
identified, including 16 positively associated target pairs and 26
negatively associated target pairs (Table IV).
| Table IV.A total of 42 miRNA-mRNA pairs that
identified, including 16 positively associated target pairs and 26
negatively associated target pairs. |
Table IV.
A total of 42 miRNA-mRNA pairs that
identified, including 16 positively associated target pairs and 26
negatively associated target pairs.
| Category | miRNA | mRNA |
|---|
| Positively | hsa-mir-145 | PDGFD |
| associated | hsa-mir-145 | PHACTR2 |
| target pairs |
hsa-mir-548c-3p | OGT |
|
|
hsa-mir-548c-3p | IGF2BP3 |
|
| hsa-mir-181a | NOTCH2 |
|
| hsa-mir-377 | STARD7 |
|
| hsa-mir-381 | WEE1 |
|
| hsa-mir-145 | SERINC5 |
|
| hsa-mir-132 | ASF1A |
|
| hsa-mir-377 | RALGPS1 |
|
| hsa-mir-132 | RASA1 |
|
|
hsa-miR-181c-3p | ZNF266 |
|
| hsa-mir-181a | FKBP1A |
|
| hsa-mir-181a | DDIT4 |
|
| hsa-mir-181a | OSBPL3 |
|
| hsa-mir-181a | HSP90B1 |
| Negatively | hsa-mir-145 | IVNS1ABP |
| associated | hsa-mir-501-5p | IVNS1ABP |
| IVNS1ABP | hsa-mir-410 | LDHA |
| target pairs | hsa-mir-181a | ZNF266 |
|
|
hsa-miR-181c-3p | NOTCH2 |
|
|
hsa-mir-199a-5p | HIF1A |
|
| hsa-mir-495-3p | MARCKS |
|
|
hsa-miR-181c-3p | FKBP1A |
|
|
hsa-miR-181c-3p | DDIT4 |
|
|
hsa-mir-199a-3p | DDIT4 |
|
| hsa-mir-495 | DDIT4 |
|
|
hsa-miR-181c-3p | OSBPL3 |
|
|
hsa-miR-181c-3p | HSP90B1 |
|
| hsa-mir-339-5p | BLCAP |
|
| hsa-mir-132 | ARHGAP3 |
|
|
hsa-mir-199a-5p | HSPA5 |
|
| hsa-mir-495 | HSPA5 |
|
| hsa-mir-582-5p | ETS2 |
|
|
hsa-mir-548c-3p | PIK3C2B |
|
| hsa-mir-381 | TES |
|
| hsa-mir-139-5p | FOS |
|
| hsa-mir-145 | SMAD3 |
|
| hsa-mir-145 | NEDD9 |
|
| hsa-mir-582-5p | ARAP2 |
|
| hsa-mir-144 | TTN |
|
| hsa-mir-346 | PTGS1 |
Discussion
As an incurable B-cell malignancy, CLL is the most
common leukemia in the western hemisphere (21). Recently, methods such as chemical
compounds, stem cell transplantation and monoclonal antibody
treatment have demonstrated high remission rates in the treatment
of CLL (22,23); however, these treatments have a
weak effect on the overall survival of CLL (24). Therefore, there is an urgent need
to identify sensitive and specific biomarkers of CLL and to find
novel therapeutic targets.
Recently, the rapid development of microarray
technology has provided a research platform for further
investigation of disease progression and identification of tumor
biomarkers, which may facilitate the identification of targets for
diagnosis, therapeutics, and prognosis of tumors (25,26).
In the present study, genes whose expression was significantly
different between CLL and normal samples were identified, and a
series of bioinformatics analyses were conducted. A total of 42
miRNA-mRNA pairs, including 17 differential miRNAs and 32
differential mRNAs were identified, and a miRNA-mRNA network
analysis was performed to explore the pathogenesis of CLL.
GO and KEGG analysis demonstrated that the DEGs
enrichment pathway reflected the complex biological behavior of
CLL, including ‘transcriptional dysregulation in cancer’ and
‘hematopoietic cell lineage’. Upregulated DEGs were associated with
multiple signaling pathways, such as cAMP and MAPK. MAPK signaling
pathway is fundamental for the maintenance of basic cellular
processes including proliferation, differentiation, migration and
survival (27). A previous study
demonstrated that dysregulation of the MAPK signaling pathway is a
common mechanism in the pathogenesis of CLL (28,29).
These results indicate that the development and progression of
cancer cells are consistent with the close association between cell
cycle/cell proliferation regulation and the dysfunction of cancer
cells.
In addition, a study reported that nodes with a high
degree of connectivity serve an important role in maintaining the
entire PPI network and are indispensable (30–32).
The 12 hub genes screened from the PPI network are the 12 most
central nodes in the network and are considered to be the key genes
associated with the development of CLL. Based on the significant
association between the node structure and its functional
importance in the PPI network (32), the 12 hub genes could be used to
gain further insight into therapeutic studies of CLL at the
molecular level.
As a gene involved in the induction of apoptosis,
MYC was identified as one of the mutant genes that
demonstrated the highest degree of connectivity in the present
study. It is known that MYC serves an oncogenic role in many
cancer types, including prostate and bladder cancers (33,34),
and serves an important role in leukemia and lymphomas (35,36).
In the present study, MYC was differentially expressed and
was enriched in the transcriptional dysregulation of cancer and
chronic myelogenous leukemia. BCL2 is the first discovered
inhibitor of apoptosis and serves a key role in promoting cell
survival. Many studies have demonstrated that BCL2 is one of
the most important oncogenes involved in cancer (37–39),
inducing the development of lymphoma, together with c-MYC
overexpression, in particular (40). In the present study, MYC and
BCL2 were identified as the top-ranking regulated candidates
among the differentially expressed genes in CLL, as previously
demonstrated (41,42).
A persistent inflammatory response is the result of
excessive production of inflammatory mediators, leading to
inflammatory diseases and cancer (43). A study demonstrated that several
CLL-associated cytokines, such as IL8, are associated with
disease stage, and the correlation of IL-8 expression levels
has been described as one of the prognostic factors for CLL
(44). It was also demonstrated
that IL8 is differentially expressed in CLL and can serve as
a biomarker, further demonstrating that the present results have a
certain degree of accuracy. CD44, another member of the
adhesion factor family, participates in the process of cell
proliferation, differentiation, adhesion and migration (45). Studies have demonstrated that
anti-CD44 antibodies can interfere with the formation of B
lymphocytes and that the downregulation of CD44 leading to
the arrest of cell cycle at the G0/G1 phase, resulting in the
decrease of proliferation of K562 cells (46–48).
In addition, the expression of CD86 in many types of
cancerous tissues was significantly lower than that in
paracancerous tissues and normal tissues (49). As a member of the Janus family of
tyrosine kinases, JAK2 is a key component of chemokine
signaling of B lymphocytes in CLL (50). Previous studies demonstrated that
inhibition of JAK2 activity in leukemia cells can promote
oncogene silencing (51). The
mutation of JAK2 activates the JAK-STAT signaling pathway
and is involved in the development of myeloproliferative neoplasms
(52), suggesting that
upregulation of the JAK pathway may be a key factor in the
progression of CLL. In addition, a series of studies demonstrated
that dysregulated expression of TGFB1, ITGAM, LCK and
FYN are involved in the occurrence of cancer and play an
important role in tumor progression (53–56).
As a tyrosine protein kinase, ZAP-70 is
involved in the signal transduction of T cells (57). Under normal conditions, B cells
have a very low expression of ZAP-70, but the expression of
ZAP-70 in B cells can be used as a specific and sensitive
indicator of B cell diseases, particularly CLL (58). In the present study, the expression
of ZAP-70 was upregulated, which is consistent with previous
studies (57,59). Gene transcription is regulated by
the transcription factor complex, which is widely involved in cell
proliferation, differentiation and regulation of transformation
(60). As a nuclear
proto-oncogene, FOS encoded protein is an important
transcription factor, which can induce the transcription and
protein expression of its downstream gene that is involved in the
regulation of cell proliferation and apoptosis (61). The abnormal expression of
FOS can induce cell transformation and tumor formation
(62). In the present study, the
dysregulation of FOS expression was observed in CLL
patients, which may be involved in abnormal gene transcription in
CLL.
Module analysis of the 12 central genes from the PPI
network revealed that 8 genes belonged to module 1, and 4 belonged
to module 2. Functional and pathway enrichment analysis of genes in
module 1 indicated that multiple pathways contribute to the
development of CLL, including the NF-κB signaling pathway, which is
involved in immune and inflammatory responses, and the
phosphoinositide 3-kinase (PI3K)-protein kinase B (Akt) signaling
pathway, which is related to chronic myelocytic leukemia resistance
and cell proliferation regulation (63). In module 2, the genes were mainly
involved in the B cell receptor signaling pathway and the Toll-like
receptor signaling pathway. The results suggest that dysregulation
of these molecular pathways in CLL may serve a role in its
pathogenesis and may eventually serve as additional biomarkers to
facilitate early diagnosis and treatment.
The aforementioned analyses demonstrated that these
genes have a variety of biological functions in tumorigenesis and
progression. A previous study demonstrated the presence of a
regulatory control network between miRNAs and mRNAs (64), in which miRNAs could silence mRNA
translation through sequence-specific targeting; therefore, it was
reasonable to investigate the pathogenesis and treatment of tumors
by studying the specific miRNA-mRNA co-regulation effect. In the
present study, 34 DEMs were identified, including 29 upregulated
and 5 downregulated miRNAs in CLL. Among them, miR-582 was the most
significantly upregulated miRNA in the GSE62137 dataset, whereas
miR-144 and miR-181a were the most significantly downregulated. A
total of 42 miRNA-mRNA pairs were identified, including 17
differential miRNAs and 32 differential mRNAs and comprising 16
positively correlated and 26 negatively correlated target
pairs.
miR-144 is expressed at low levels in various
tumors, such as hepatocellular carcinoma (65), B-cell lymphoma (66) and chronic lymphocytic leukemia
(67). In the present study,
TTN was predicted to be the target of miR-144, which was
upregulated in CLL according to the GSE22529 and GSE39411 datasets.
TTN may be a good predictor of disease progression,
survival, and treatment response in metastatic prostate cancer
(68). Other studies have
demonstrated that miR-582 was highly expressed in a variety of
tumors, such as colorectal cancer (69) and non-small cell lung carcinoma
(70), and that the upregulation
of miR-582 contributed to an increase in the proliferation of
prostate cancer cells (71) and to
the reduction of human bladder cancer proliferation and invasion by
suppressing the expression of target genes (72). In the present study, miR-582 was
significantly overexpressed in CLL cells compared to normal samples
and that it targeted the downregulated ETS2 and ARAP2
genes. There are few reports regarding the role of ETS2 and
ARAP2 in cancer. Some studies indicated that the expression
of ETS2 might be associated with the progression of CML
(73). Further study is needed to
determine the role of ETS2 and ARAP2 in the
development of CLL. The results of a previous study confirmed that
miR-181a was significantly downregulated in CLL (74). NOTCH2, one of the target genes of
miR-181a, is involved in the regulation of cellular processes
including differentiation, proliferation and apoptosis (75), and dysregulation of NOTCH2
signaling is involved in aberrant CD23 expression in B-cell
CLL (76).
As miRNAs can participate in complex biological
functions in the body, they can influence a variety of biological
processes and pathways through miRNA-mRNA regulatory networks
(77,78). Providing that each miRNA
potentially regulates a large number of targets, understanding the
function of miRNAs in tumorigenesis may provide insight into the
key questions that remain in cancer research. The miRNA-mRNA
regulatory network highlighted in the present study provides new
theoretical guidance for further exploring the mechanism of CLL and
provides a novel perspective for understanding the underlying
biological processes of CLL and miRNA-targeted therapy.
Taken together, the present data demonstrated that
the molecular mechanisms of CLL can be understood and that
biomarker prediction can be achieved through data mining and
integration. Overall, 488 candidate genes and 32 different miRNAs
from multiple datasets were screened by bioinformatics analysis.
From the PPI network of DEGs, 12 major altered hub genes and
multiple pathways associated with the development of CLL were
identified, such as the Jak-STAT signaling pathway, the PI3K-Akt
signaling pathway and the MAPK signaling pathway. In addition, the
comprehensive bioinformatics analysis of DEGs and the construction
of miRNA-mRNA regulatory network provides a bioinformatics approach
to further study the pathogenesis of CLL and lay a solid foundation
for the personalized treatment of CLL in the future. However,
further studies are needed to validate these results.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81673799)
and the National Natural Science Foundation of China Youth Fund
(grant no. 81703915).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
CG, CS and CZ conceived and designed the study. JZ
and LL performed data analysis. JW, CL, and HL integrated the data
and created the figures. CZ reviewed and revised the article. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chiorazzi N, Rai KR and Ferrarini M:
Chronic lymphocytic leukemia. N Engl J Med. 352:804–815. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fernando TR, Rodriguez-Malave NI and Rao
DS: MicroRNAs in B cell development and malignancy. J Hematol
Oncol. 5:72012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martini V, Frezzato F, Severin F, Raggi F,
Trimarco V, Martinello L, Molfetta R, Visentin A, Facco M,
Semenzato G, et al: Abnormal regulation of BCR signalling by c-Cbl
in chronic lymphocytic leukaemia. Oncotarget. 9:32219–32231. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rajendran V: Structural analysis of
oncogenic mutation of isocitrate dehydrogenase 1. Mol Biosyst.
12:2276–2287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rajendran V, Gopalakrishnan C and
Sethumadhavan R: Pathological role of a point mutation (T315I) in
BCR-ABL1 protein-A computational insight. J Cell Biochem.
119:918–925. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luo P, Yang Q, Cong LL, Wang XF, Li YS,
Zhong XM, Xie RT, Jia CY, Yang HQ, Li WP, et al: Identification of
miR124a as a novel diagnostic and prognostic biomarker in nonsmall
cell lung cancer for chemotherapy. Mol Med Rep. 16:238–246. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kamaraj B, Gopalakrishnan C and Purohit R:
In silico analysis of miRNA-mediated gene regulation in OCA and OA
genes. Cell Biochem Biophys. 70:1923–1932. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Han X, Wu H and Zhou Y:
Bioinformatics analysis of transcription profiling of solid
pseudopapillary neoplasm of the pancreas. Mol Med Rep.
16:1635–1642. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vogelstein B, Papadopoulos N, Velculescu
VE, Zhou S, Diaz LA Jr and Kinzler KW: Cancer genome landscapes.
Science. 339:1546–1558. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lim L, Lau N, Garrett-Engele P, Grimson A,
Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bhaumik P, Gopalakrishnan C, Kamaraj B and
Purohit R: Single nucleotide polymorphisms in microRNA binding
sites: Implications in colorectal cancer. ScientificWorldJournal.
2014:5471542014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gutierrez A Jr, Tschumper RC, Wu X,
Shanafelt TD, Eckel-Passow J, Huddleston PM III, Slager SL, Kay NE
and Jelinek DF: LEF-1 is a prosurvival factor in chronic
lymphocytic leukemia and is expressed in the preleukemic state of
monoclonal B-cell lymphocytosis. Blood. 116:2975–2983. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vallat L, Kemper CA, Jung N,
Maumy-Bertrand M, Bertrand F, Meyer N, Pocheville A, Fisher JW III,
Gribben JG and Bahram S: Reverse-engineering the genetic circuitry
of a cancer cell with predicted intervention in chronic lymphocytic
leukemia. Proc Natl Acad Sci USA. 110:459–464. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ruiz-Lafuente N, Alcaraz-García MJ,
Sebastián-Ruiz S, García-Serna AM, Gómez-Espuch J, Moraleda JM,
Minguela A, García-Alonso AM and Parrado A: IL-4 up-regulates
MiR-21 and the MiRNAs hosted in the CLCN5 gene in chronic
lymphocytic leukemia. PLoS One. 10:e01249362015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43:D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wong N and Wang X: miRDB: An online
resource for microRNA target prediction and functional annotations.
Nucleic Acids Res. 43:D146–D152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chou CH, Shrestha S, Yang CD, Chang NW,
Lin YL, Liao KW, Huang WC, Sun TH, Tu SJ, Lee WH, et al: miRTarBase
update 2018: A resource for experimentally validated
microRNA-target interactions. Nucleic Acids Res. 46:D296–D302.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nabhan C and Rosen ST: Chronic lymphocytic
leukemia: A clinical review. JAMA. 312:2265–2276. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cang S, Iragavarapu C, Savooji J, Song Y
and Liu D: ABT-199 (venetoclax) and BCL-2 inhibitors in clinical
development. J Hematol Oncol. 8:1292015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rai KR: Therapeutic potential of new B
cell-targeted agents in the treatment of elderly and unfit patients
with chronic lymphocytic leukemia. J Hematol Oncol. 8:852015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kwok M, Davies N, Agathanggelou A, Smith
E, Oldreive C, Petermann E, Stewart G, Brown J, Lau A, Pratt G, et
al: ATR inhibition induces synthetic lethality and overcomes
chemoresistance in TP53 or ATM defective chronic lymphocytic
leukemia cells. Blood. 127:582–595. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen F, Shen C, Wang X, Wang H, Liu Y, Yu
C, Lv J, He J and Wen Z: Identification of genes and pathways in
nasopharyngeal carcinoma by bioinformatics analysis. Oncotarget.
8:63738–63749. 2017.PubMed/NCBI
|
|
26
|
Sun C, Yuan Q, Wu D, Meng X and Wang B:
Identification of core genes and outcome in gastric cancer using
bioinformatics analysis. Oncotarget. 8:70271–70280. 2017.PubMed/NCBI
|
|
27
|
Zhou YY, Li Y, Jiang WQ and Zhou LF:
MAPK/JNK signalling: A potential autophagy regulation pathway.
Biosci Rep. 35:e001992015.PubMed/NCBI
|
|
28
|
Shukla A, Shukla V and Joshi SS:
Regulation of MAPK signaling and implications in chronic
lymphocytic leukemia. Leuk Lymphoma. 59:1565–1573. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pandzic T, Larsson J, He L, Kundu S, Ban
K, Akhtar-Ali M, Hellström AR, Schuh A, Clifford R, Blakemore SJ,
et al: Transposon mutagenesis reveals fludarabine resistance
mechanisms in chronic lymphocytic leukemia. Clin Cancer Res.
22:6217–6227. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He X and Zhang J: Why do hubs tend to be
essential in protein networks? PLoS Genet. 2:e882006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barabási A and Oltvai Z: Network biology:
Understanding the cell's functional organization. Nat Rev Genet.
5:101–113. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hahn M and Kern A: Comparative genomics of
centrality and essentiality in three eukaryotic protein-interaction
networks. Mol Biol Evol. 22:803–806. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fisher KW, Montironi R, Lopez Beltran A,
Moch H, Wang L, Scarpelli M, Williamson SR, Koch MO and Cheng L:
Molecular foundations for personalized therapy in prostate cancer.
Curr Drug Targets. 16:103–114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu X, Liu D, Tao D, Xiang W, Xiao X, Wang
M, Wang L, Luo G, Li Y, Zeng F and Jiang G: BRD4 regulates EZH2
transcription through upregulation of C-MYC and represents a novel
therapeutic target in bladder cancer. Mol Cancer Ther.
15:1029–1042. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sánchez-Beato M, Sánchez-Aguilera A and
Piris MA: Cell cycle deregulation in B-cell lymphomas. Blood.
101:1220–1235. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Delgado MD and León J: Myc roles in
hematopoiesis and leukemia. Genes Cancer. 1:605–616. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vaux DL, Cory S and Adams JM: Bcl-2 gene
promotes haemopoietic cell survival and cooperates with c-myc to
immortalize pre-B cells. Nature. 335:440–442. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dawson SJ, Makretsov N, Blows FM, Driver
KE, Provenzano E, Le Quesne J, Baglietto L, Severi G, Giles GG,
McLean CA, et al: BCL2 in breast cancer: A favourable prognostic
marker across molecular subtypes and independent of adjuvant
therapy received. Br J Cancer. 103:668–675. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cheng H, Wang X, Li T and Chen L: Bcl-2
expression and patient survival in gastric cancer: A systematic
review of the literature with meta-analysis. Med Oncol. 32:3892015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Strasser A, Harris AW, Bath ML and Cory S:
Novel primitive lymphoid tumours induced in transgenic mice by
cooperation between myc and bcl-2. Nature. 348:331–333. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Landau D, Tausch E, Taylor-Weiner A,
Stewart C, Reiter JG, Bahlo J, Kluth S, Bozic I, Lawrence M,
Böttcher S, et al: Mutations driving CLL and their evolution in
progression and relapse. Nature. 526:525–530. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Robertson L, Plunkett W, McConnell K,
Keating M and McDonnell T: Bcl-2 expression in chronic lymphocytic
leukemia and its correlation with the induction of apoptosis and
clinical outcome. Leukemia. 10:456–459. 1996.PubMed/NCBI
|
|
43
|
Kaser A, Zeissig S and Blumberg RS:
Inflammatory bowel disease. Annu Rev Immunol. 28:573–621. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wierda WG, Johnson MM, Do KA, Manshouri T,
Dey A, O'Brien S, Giles FJ, Kantarjian H, Thomas D, Faderl S, et
al: Plasma interleukin 8 level predicts for survival in chronic
lymphocytic leukaemia. Br J Haematol. 120:452–456. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Slevin M, Krupinski J, Gaffney J, Matou S,
West D, Delisser H, Savani RC and Kumar S: Hyaluronan-mediated
angiogenesis in vascular disease: Uncovering RHAMM and CD44
receptor signaling pathways. Matrix Biol. 26:58–68. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chang G, Zhang H, Wang J, Zhang Y, Xu H,
Wang C, Zhang H, Ma L, Li Q and Pang T: CD44 targets Wnt/β-catenin
pathway to mediate the proliferation of K562 cells. Cancer Cell
Int. 13:1172013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Krause DS, Lazarides K, von Andrian UH and
Van Etten RA: Requirement for CD44 in homing and engraftment of
BCR-ABL-expressing leukemic stem cells. Nat Med. 12:1175–1180.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang Y, Yang J, Li J, Wang RC, Yuan J, Li
Y, Wang SY, Wang C and Hao HL: Effect of arsenic trioxide on K562
cell proliferation and its mechanism. Zhongguo Shi Yan Xue Ye Xue
Za Zhi. 25:90–93. 2017.(In Chinese). PubMed/NCBI
|
|
49
|
Yang WF and WANG FZ: Expression of CD1a,
CD80, CD86 in dendritic cell of tumor tissue and regional lymph
node in esophageal carcinoma. Ai Zheng. 23:189–192. 2004.(In
Chinese). PubMed/NCBI
|
|
50
|
Montresor A, Toffali L, Mirenda M, Rigo A,
Vinante F and Laudanna C: JAK2 tyrosine kinase mediates integrin
activation induced by CXCL12 in B-cell chronic lymphocytic
leukemia. Oncotarget. 6:34245–34257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tefferi A and Pardanani A: JAK inhibitors
in myeloproliferative neoplasms: Rationale, current data and
perspective. Blood Rev. 25:229–237. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang HY and Zhai XW: Research advances in
the role of JAK2 mutations in acute leukemia. Zhongguo Dang Dai Er
Ke Za Zhi. 17:644–649. 2015.(In Chinese). PubMed/NCBI
|
|
53
|
Blahna MT and Hata A: Smad-mediated
regulation of microRNA biosynthesis. FEBS Lett. 586:1906–1912.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fabregat I, Moreno-Càceres J, Sànchez A,
Dooley S, Dewidar B, Giannelli G and Ten Dijke P: IT-LIVER
Consortium: TGF-β signalling and liver disease. FEBS J.
283:2219–2232. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Serafin V, Lissandron V, Buldini B,
Bresolin S, Paganin M, Grillo F, Andriano N, Palmi C, Cazzaniga G,
Marmiroli S, et al: Phosphoproteomic analysis reveals
hyperactivation of mTOR/STAT3 and LCK/Calcineurin axes in pediatric
early T-cell precursor ALL. Leukemia. 31:1007–1011. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ban K, Gao Y, Amin HM, Howard A, Miller C,
Lin Q, Leng X, Munsell M, Bar-Eli M, Arlinghaus RB and Chandra J:
BCR-ABL1 mediates up-regulation of Fyn in chronic myelogenous
leukemia. Blood. 111:2904–2908. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen L, Widhopf G, Huynh L, Rassenti L,
Rai KR, Weiss A and Kipps TJ: Expression of ZAP-70 is associated
with increased B-cell receptor signaling in chronic lymphocytic
leukemia. Blood. 100:4609–4614. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Orchard JA, Ibbotson RE, Davis Z, Wiestner
A, Rosenwald A, Thomas PW, Hamblin TJ, Staudt LM and Oscier DG:
ZAP-70 expression and prognosis in chronic lymphocytic leukaemia.
Lancet. 363:105–111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Purroy N, Abrisqueta P, Carabia J, Carpio
C, Palacio C, Bosch F and Crespo M: Co-culture of primary CLL cells
with bone marrow mesenchymal cells, CD40 ligand and CpG ODN
promotes proliferation of chemoresistant CLL cells phenotypically
comparable to those proliferating in vivo. Oncotarget. 6:7632–7643.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kolla V, Gonzales L, Gonzales J, Wang P,
Angampalli S, Feinstein SI and Ballard PL: Thyroid transcription
factor in differentiating type II cells: Regulation, isoforms, and
target genes. Am J Respir Cell Mol Biol. 36:213–225. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Durchdewald M, Angel P and Hess J: The
transcription factor Fos: A Janus-type regulator in health and
disease. Histol Histopathol. 24:1451–1461. 2009.PubMed/NCBI
|
|
62
|
Wagstaff SC, Bowler WB, Gallagher JA and
Hipskind RA: Extracellular ATP activates multiple signalling
pathways and potentiates growth factor-induced c-fos gene
expression in MCF-7 breast cancer cells. Carcinogenesis. 2175–2181.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhao L, Shan Y, Liu B, Li Y and Jia L:
Functional screen analysis reveals miR-3142 as central regulator in
chemoresistance and proliferation through activation of the
PTEN-AKT pathway in CML. Cell Death Dis. 8:e28302017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chiang HR, Schoenfeld LW, Ruby JG, Auyeung
VC, Spies N, Baek D, Johnston WK, Russ C, Luo S, Babiarz JE, et al:
Mammalian microRNAs: Experimental evaluation of novel and
previously annotated genes. Genes Dev. 24:992–1009. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Cao T, Li H, Hu Y, Ma D and Cai X: miR-144
suppresses the proliferation and metastasis of hepatocellular
carcinoma by targeting E2F3. Tumour Biol. 35:10759–10764. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang H, Wang A, Hu Z, Xu X, Liu Z and Wang
Z: A critical role of miR-144 in diffuse large B-cell lymphoma
proliferation and invasion. Cancer Immunol Res. 4:337–344. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Machová Poláková K, Lopotová T, Klamová H,
Burda P, Trněný M, Stopka T and Moravcová J: Expression patterns of
microRNAs associated with CML phases and their disease related
targets. Mol Cancer. 10:412011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Tomioka A, Tanaka N, Yoshikawa M, Miyake
M, Anai S, Chihara Y, Okajima E, Hirayama A, Hirao Y and Fujimoto
K: Nadir PSA level and time to nadir PSA are prognostic factors in
patients with metastatic prostate cancer. BMC Urol. 14:332014.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Song B, Long Y, Liu D, Zhang W and Liu C:
MicroRNA-582 promotes tumorigenesis by targeting phosphatase and
tensin homologue in colorectal cancer. Int J Mol Med. 40:867–874.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fang L, Cai J, Chen B, Wu S, Li R, Xu X,
Yang Y, Guan H, Zhu X, Zhang L, et al: Aberrantly expressed
miR-582-3p maintains lung cancer stem cell-like traits by
activating Wnt/β-catenin signalling. Nat Commun. 6:86402015.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Maeno A, Terada N, Uegaki M, Goto T, Okada
Y, Kobayashi T, Kamba T, Ogawa O and Inoue T: Up-regulation of
miR-582-5p regulates cellular proliferation of prostate cancer
cells under androgen-deprived conditions. Prostate. 74:1604–1612.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Uchino K, Takeshita F, Takahashi RU,
Kosaka N, Fujiwara K, Naruoka H, Sonoke S, Yano J, Sasaki H, Nozawa
S, et al: Therapeutic effects of microRNA-582-5p and −3p on the
inhibition of bladder cancer progression. Mol Ther. 21:610–619.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lafage-Pochitaloff M, Courcoul M,
Simonetti J, Sainty D, Dastugue N, Tabilio A, Hagemeijer A and Birg
F: Expression of the ETS2 and transferrin receptor genes in
Philadelphia-positive chronic myeloid leukemia patients with a
reciprocal t(3;21). Genes Chromosomes Cancer. 5:1–13. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhu DX, Miao KR, Fang C, Fan L, Zhu W, Zhu
HY, Zhuang Y, Hong M, Liu P, Xu W and Li JY: Aberrant microRNA
expression in Chinese patients with chronic lymphocytic leukemia.
Leuk Res. 35:730–734. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hubmann R, Schwarzmeier JD, Shehata M,
Hilgarth M, Duechler M, Dettke M and Berger R: Notch2 is involved
in the overexpression of CD23 in B-cell chronic lymphocytic
leukemia. Blood. 99:3742–3747. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Song HM, Luo Y, Li DF, Wei CK, Hua KY,
Song JL, Xu H, Maskey N and Fang L: MicroRNA-96 plays an oncogenic
role by targeting FOXO1 and regulating AKT/FOXO1/Bim pathway in
papillary thyroid carcinoma cells. Int J Clin Exp Pathol.
8:9889–9900. 2015.PubMed/NCBI
|
|
78
|
Chruścik A and Lam AK: Clinical
pathological impacts of microRNAs in papillary thyroid carcinoma: A
crucial review. Exp Mol Pathol. 99:393–398. 2015. View Article : Google Scholar : PubMed/NCBI
|