Introduction
Visfatin, additionally termed pre-B-cell
colony-enhancing factor or nicotinamide phosphoribosyltransferase
(Nampt), is an adipocytokine that is predominantly produced in
visceral adipose tissue (1–3);
however, during pregnancy, visfatin is additionally expressed and
secreted by the placental tissue (4,5).
Visfatin is involved in the pathogenesis of various metabolic
disorders; increased plasma concentrations of visfatin have been
reported in individual with obesity, as well as in patients with
gestational diabetes mellitus (GDM) or metabolic syndromes
(6–8).
Visfatin functions as an immunomodulatory cytokine
involved in the inflammatory responses, and is a factor associated
with obesity, inflammation and insulin resistance (3,6).
Upregulation of inflammatory cytokines, including tumor necrosis
factor-α (TNF-α) and interleukin 6 (IL-6), serves important roles
in the induction of insulin resistance (9). Numerous previous studies indicated
that visfatin was associated with inflammation (10–12);
however, the underlying molecular mechanisms remain unknown.
Furthermore, visfatin functions as the rate-limiting enzyme of
NAD+ biosynthesis and converts nicotinamide into
nicotinamide mononucleotide, a key NAD+ intermediate
(13). In addition,
visfatin-mediated NAD+ biosynthesis in adipocytes is an
important physiological regulator of the metabolic function of
adipose tissue and whole-body (13). Although still controversial,
visfatin was reported to exert insulin mimetic or sensitizing
effects; similar to insulin, visfatin increases lipogenesis,
enhances glucose uptake of myocytes and adipocytes, and inhibits
hepatocyte glucose release, consequently contributing to glucose
and lipid metabolism (12,14,15).
Previous studies identified the upregulation of IL-6 in the
placenta of patients with GDM (16,17).
In addition, it was indicated that the expression of visfatin in
placenta was unchanged in patients with GDM (18). IL-6 (16,17)
and visfatin (8,9) are associated with GDM. Kralisch et
al (19) observed that IL-6
suppressed the mRNA synthesis of visfatin in 3T3-L1 adipocytes;
however, in human adipocytes and amniotic epithelial cells, IL-6
significantly promoted visfatin protein expression (20,21).
Therefore, the present study hypothesized that the modulation of
visfatin expression by IL-6 may be tissue specific; however, how
pro-inflammatory IL-6 regulates the expression of visfatin in
placenta remains unknown.
Peroxisome proliferator-activated receptor-γ
(PPAR-γ) is a nuclear receptor that is abundantly expressed in
adipocytes, where it regulates adipogenesis and lipid homeostasis
(22). PPAR-γ functions in other
tissues by promoting anti-immune responses and lipid metabolism in
macrophages, increasing lipid storage in the liver and enhancing
glucose-stimulated insulin secretion in pancreatic β-cells
(22,23). PPAR-γ is involved in the
pathophysiology of GDM and serves key roles in the regulation of
glucose metabolism, lipid homeostasis and anti-inflammatory
processes (22,23). Previous studies identified the
downregulation of PPAR-γ in the trophoblast cells of patients with
GDM (24–26). Mayi et al (12) observed that synthetic PPAR-γ
ligands upregulated the expression of visfatin gene in a
PPAR-γ-dependent manner in resting primary human macrophages and
adipose tissue macrophages, but not in adipocytes; however, whether
PPAR-γ ligands induce the gene expression of visfatin in placenta
remains unknown.
BeWo cells possess characteristics of the
syncytiotrophoblast and extravillous trophoblast, and serve as an
in vitro model to investigate trophoblast fusion (27,28).
BeWo cells are frequently used to investigate the regulation of
transplacental transport and cellular activities and uptake
mechanisms (28). The aim of the
present study was to determine the effect of IL-6 on the expression
and secretion of visfatin in BeWo cells, and to determine whether
the PPAR-γ pathway is involved in the regulation of visfatin by
IL-6.
Materials and methods
Cell culture and treatment
All materials were purchased from Gibco (Thermo
Fisher Scientific, Inc., Waltham, MA, USA) unless otherwise stated.
BeWo cells were obtained from the National Infrastructure of Cell
Line Resource (Beijing, China). The cells were cultured in Ham's
F12 medium supplemented with 10% fetal bovine serum (ScienCell
Research Laboratories, Inc., San Diego, CA, USA), 1% L-glutamine,
1% penicillin and 1% streptomycin (Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China) at 37°C in a humidified
atmosphere of 5% CO2. BeWo cells were seeded at a
density of 2×106 cells/ml in six-well plates or 25
cm2 culture flasks (Corning Incorporated, Corning, NY,
USA). Cells in the logarithmic growth phase were incubated at 37°C
for 12, 24, 48 and 72 h with recombinant human IL-6 (cat. no.
200-06; PeproTech, Inc.) at serial concentrations (0, 1, 5 and 10
ng/ml). Additionally, BeWo cells were treated with the PPAR-γ
ligand pioglitazone (cat. no. HY-14601; MedChem Express Co., Ltd.,
Monmouth Junction, NJ, USA) at serial concentrations (0.1, 1 and 10
µM) or dimethyl sulfoxide as a control at 37°C for 48 h.
Subsequently, BeWo cells were incubated with recombinant human IL-6
(5 ng/ml) and/or the PPAR-γ agonist pioglitazone (10 µM), or
dimethyl sulfoxide as a control at 37°C for 48 h. The supernatants
were collected, and levels of secreted visfatin were determined
using an ELISA kit (cat. no. KE1720; ImmunoWay Biotechnology
Company, Plano, TX, USA) according to the manufacturer's protocols.
Additionally, BeWo cells were treated with recombinant human
visfatin (cat. no. 130-09; PeproTech, Inc., Rocky Hill, NJ, USA) at
serial concentrations (0, 10, 50 and 100 ng/ml) at 37°C for 48 h.
The secretion levels of IL-6 and TNF-α were determined using an
IL-6 ELISA kit (cat. no. EHC007; Xinbosheng Biotechnology Co.,
Ltd., Beijing, China) and TNF-α ELISA kit (cat. no. EHC103a;
Xinbosheng Biotechnology Co., Ltd.) respectively, according to the
manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from BeWo cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). RNA was reverse transcribed into cDNA using the Easy Script
First Strand cDNA Synthesis Super Mix kit (Beijing Transgen Biotech
Co., Ltd., Beijing, China) according to the manufacturer's
protocol. RT was performed with a total of 20 µl reaction mixture
at 42°C for 15 min and 95°C for 3 min. Specific primers designed
for the amplification of visfatin, PPAR-γ, and GAPDH were verified
by NCBI BLAST (http://blast.ncbi.nlm.gov/Blast.cgi). The sequences
were as follows: Human visfatin, forward,
5′-GCCAGCAGGGAATTTTGTTA-3′ and reverse, 5′-TGATGTGCTGCTTCCAGTTC-3′;
human PPAR-γ, forward, 5′-GCCCTTCACTACTGTTGACTTCT-3′ and reverse,
5′-CAGGCTCCACTTTGATTGC-3′; and human GAPDH, forward,
5′-TGAACGGGAAGCTCACTG-3′ and reverse, 5′-GCTTCACCACCTTCTTGATG-3′.
Reactions were performed on an ABI PRISM 7300 PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using
PowerUp™ SYBR Green Master Mix (Applied Biosystems;
Thermo Fisher Scientific, Inc.). qPCR was performed with a total of
20 µl reaction mixture at 50°C for 2 min and 95°C for 2 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. Gene
expression levels were analyzed in triplicate and normalized to
GAPDH using the 2−∆∆Cq method (29).
Western blot analysis
Treated BeWo cells were washed twice using ice-cold
PBS, and incubated using lysis buffer [1% NP-40, 150 mM NaCl, 50
mMTris (pH 8.0), 0.1% aprotinin, 0.1% leupeptin, 0.035% pepstatin A
and 100 µg/ml PMSF] supplemented with protease and phosphatase
inhibitor cocktail (Beijing Solarbio Science & Technology Co.,
Ltd.) at 4°C for 30 min. The samples were centrifuged at 11,600 × g
at 4°C for 30 min, and protein concentrations were determined using
a bicinchoninic acid assay kit (Thermo Fisher Scientific, Inc.).
The protein samples were subsequently denatured in SDS sample
buffer (125 mM Tris-HCl, pH 6.8, 50% glycerol, 2% SDS, 5%
β-mercaptoethanol and 0.01% bromophenol blue) at 100°C for 10 min.
Equal amounts of protein (30 µg/lane) were separated by 10%
SDS-PAGE and transferred onto polyvinylidene difluoride membranes
(EMD Millipore, Billerica, MA, USA). Subsequently, the membranes
were blocked in Tris-buffered saline with 5% non-fat milk at room
temperature for 4 h. The membranes were incubated with rabbit
anti-human primary antibodies raised against visfatin (1:250; cat.
no. ab45890; Abcam, Cambridge, MA, USA) and GAPDH (1:5,000; cat.
no. AP0063; Bioworld Technology, Inc., Louis Park, MN, USA)
overnight at 4°C, followed by incubation with a horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G secondary
antibody (1:5,000; cat. no. L3012-2; Signalway Antibody, LLC,
College Park, MD, USA) at room temperature for 2 h. The bands were
visualized using an enhanced chemiluminescence detection kit (GE
Healthcare, Chicago, IL, USA). Band densities were quantified using
Bio-Rad ChemiDoc™ XRS+System with Image Lab™
software version 4.1 (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). The experiments were repeated at least three times.
Statistical analysis
The experiments were repeated at least three times.
Data were presented as the mean ± standard deviation and analyzed
using SPSS 21.0 (IBM Corp., Armonk, NY, USA). Groups were compared
by one-way analysis of variance and a Student-Newman-Keuls post-hoc
test for normally distributed data. Correlation analysis was
performed using Spearman's or Pearson's correlation coefficients.
P<0.05 was considered to indicate a statistically significant
difference.
Results
IL-6 reduces the expression of
visfatin gene in BeWo cells
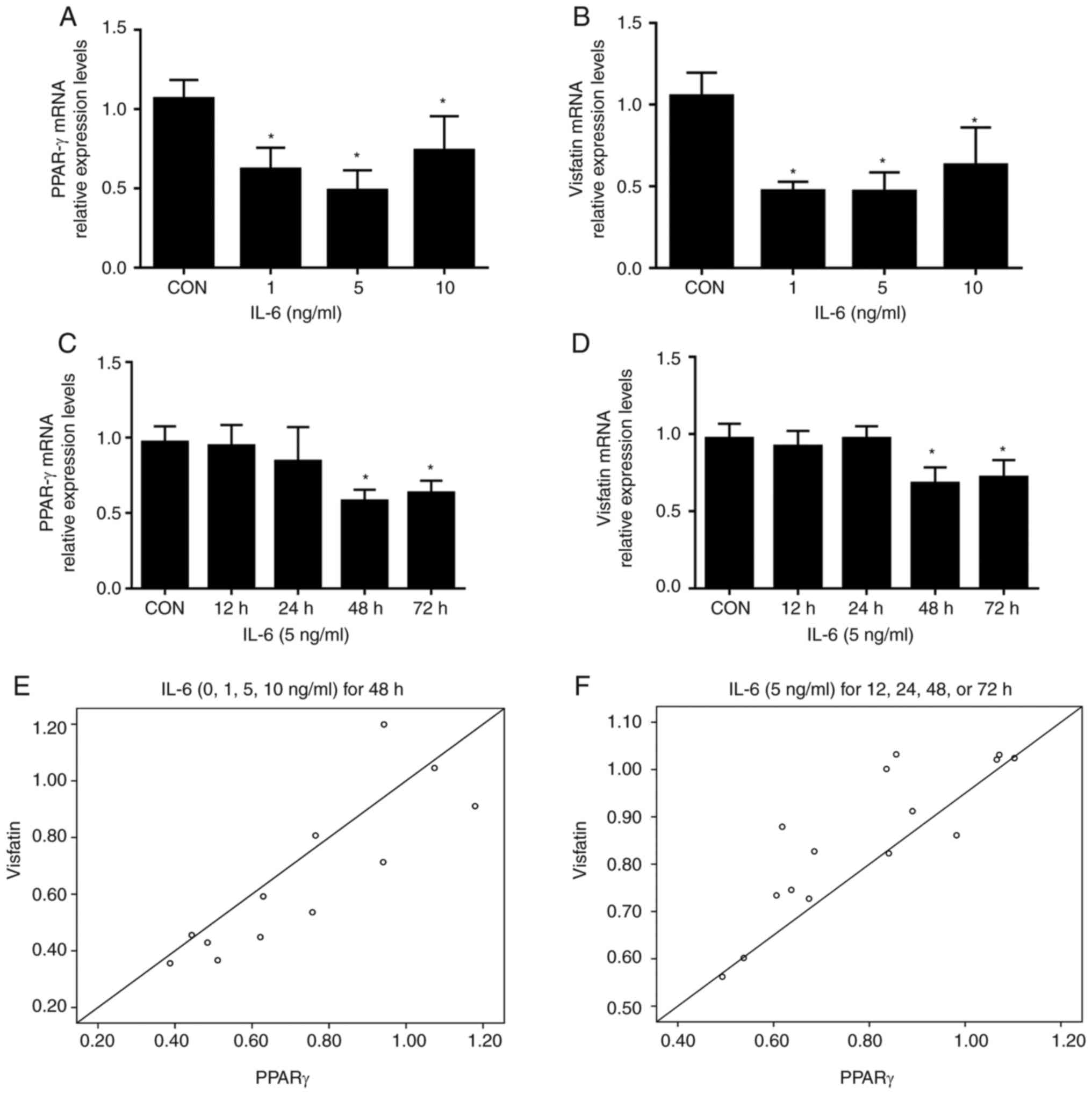
Treatment of BeWo cells with increasing
concentrations of IL-6 (1, 5, and 10 ng/ml) resulted in
significantly reduced levels of PPAR-γ and visfatin mRNA compared
with the control; no further decrease was detected with 10 ng/ml
IL-6 (Fig. 1A and B). Time course
experiments demonstrated that the inhibitory effects of IL-6 (5
ng/ml) on visfatin and PPAR-γ mRNA peaked at 48 h; no further
reductions were observed at 72 h (Fig.
1C and D). Pearson correlation analysis demonstrated that the
mRNA levels of visfatin were positively correlated with the
expression of PPAR-γ (r=0.857, P<0.05; r=0.854, P<0.001,
Fig. 1E and F, respectively). In
summary, the data of the present study demonstrated that IL-6
inhibited the expression of visfatin in BeWo cells, which was
associated with PPAR-γ.
PPAR-γ agonist pioglitazone induces
the expression of visfatin gene of BeWo cells
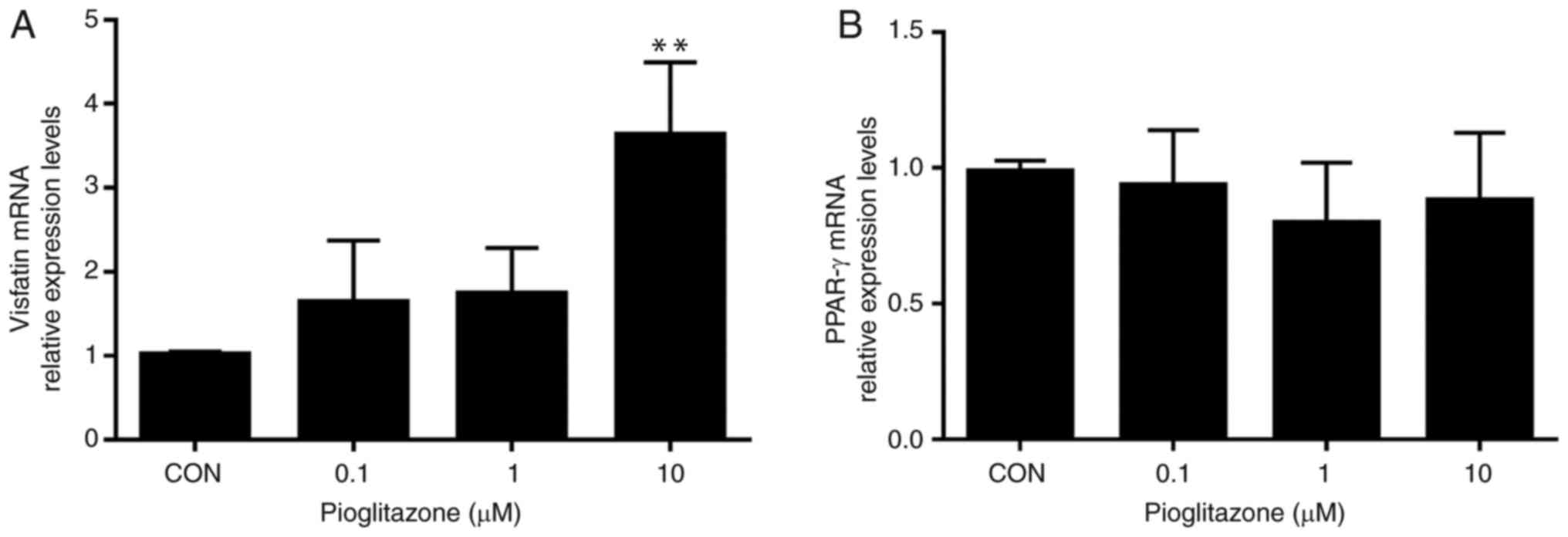
Treatment with increasing concentrations of
pioglitazone (0.1, 1 and 10 µM) upregulated the mRNA expression
levels of visfatin in a dose-dependent manner, whereas, no
significant differences were observed in the mRNA levels of PPAR-γ
among the treatment groups (Fig.
2). Furthermore, activation of PPAR-γ by exposure to
pioglitazone (10 µM) for 48 h resulted in significantly increased
expression of visfatin compared with the control, which abrogated
the inhibitory effects of IL-6 on visfatin in BeWo cells (Fig. 3A). In summary, the results of the
present study demonstrated that visfatin may be a target gene of
PPAR-γ in BeWo cells and that the PPAR-γ pathway may be involved in
the regulation of visfatin by IL-6.
PPAR-γ activation regulates the
expression and secretion of visfatin protein of BeWo cells
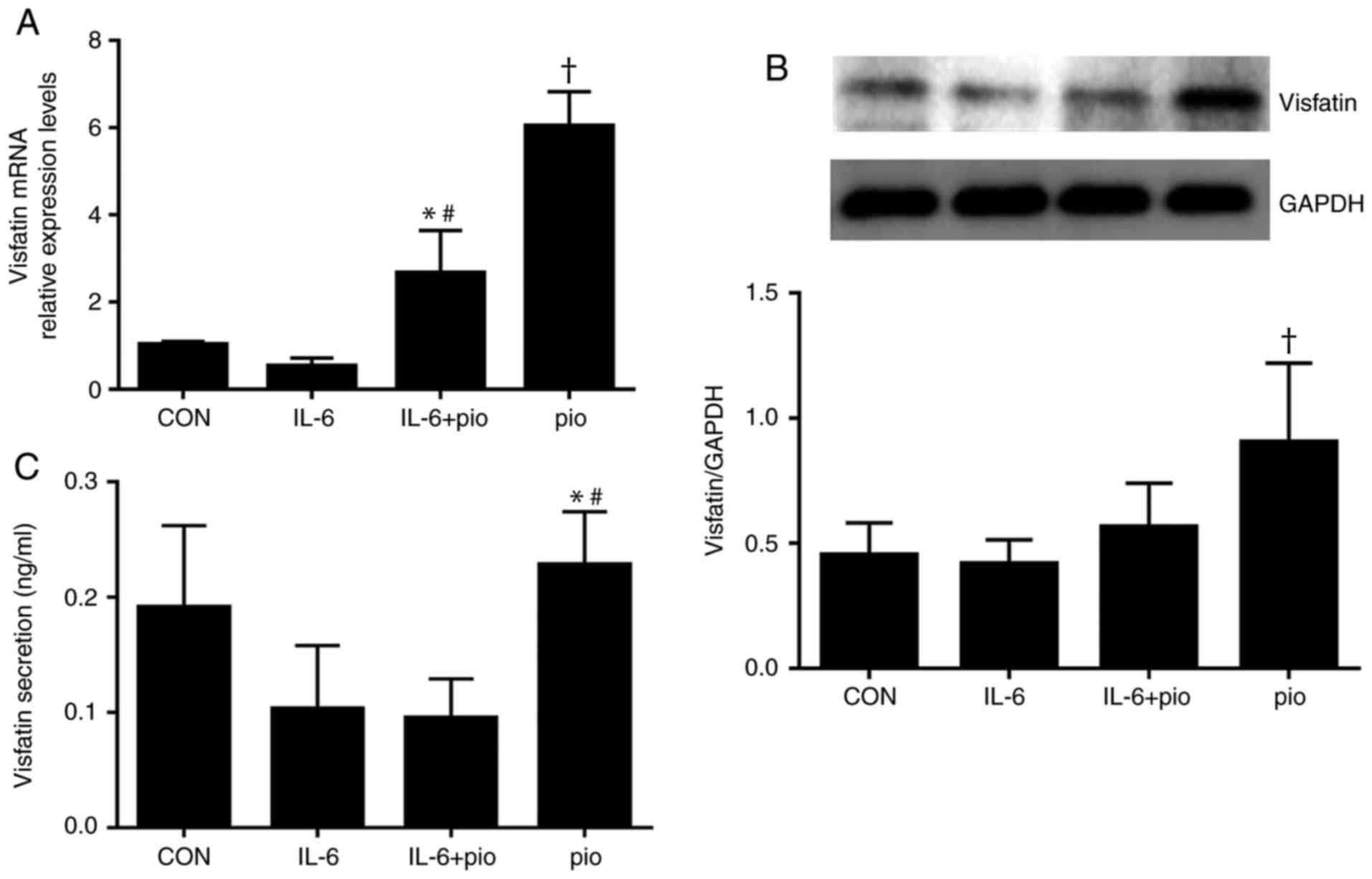
To investigate whether pioglitazone-induced
expression of visfatin resulted in increased protein levels, BeWo
cells treated with IL-6 (5 ng/ml) and/or pioglitazone (10 µM) for
48 h were subjected to western blot analysis. IL-6 exhibited no
significant effect on the protein expression of visfatin, while
activation of PPAR-γ resulted in significantly increased protein
expression levels of visfatin compared with the control, IL-6
treatment alone or in combination with pioglitazone (Fig. 3B); however, the expression levels
of PPAR-γ protein were not affected by IL-6 or pioglitazone (data
not shown). To examine whether induction by pioglitazone promoted
the secretion of visfatin secretion, the ability of PPAR-γ to
stimulate the release of visfatin was evaluated. As presented in
Fig. 3C, treatment with
pioglitazone (10 µM) significantly increased the levels of visfatin
in BeWo cell compared with IL-6 alone (5 ng/ml) or in combination
with pioglitazone. The findings suggested that pioglitazone
upregulated the expression level of visfatin by activating the
PPAR-γ pathway and the PPAR-γ pathway may be involved in the
regulation of visfatin by IL-6.
Discussion
In the present study, the expression of visfatin in
BeWo cells was evaluated in the presence of IL-6. Downregulation of
visfatin was observed in BeWo cells treated with IL-6, which was
consistent with the findings in 3T3-L1 adipocytes by Kralisch et
al (19). The present study
additionally identified that IL-6 reduced the mRNA expression level
of PPAR-γ in BeWo cells and was associated with the expression of
visfatin. The PPAR-γ agonist pioglitazone induced the expression of
visfatin in BeWo cells, possibly in a PPAR-γ-dependent manner,
which was consistent with the findings by Mayi et al
(12) in resting primary human
macrophages and adipose tissue macrophages. To the best of our
knowledge, the present study is the first to demonstrate that
visfatin may be a target gene of PPAR-γ in BeWo cells. In addition,
the PPAR-γ pathway could be involved in the regulation of visfatin
by IL-6; however, Lv et al (15) reported that pioglitazone
ameliorates insulin resistance by downregulating visfatin
expression in visceral adipose tissue and reducing circulating
visfatin in high-fat-fed rats, whereas, no effect of pioglitazone
was observed during the differentiation of 3T3-L1 adipocytes. This
discrepancy may be due to the different sources of cells and
tissues, which indicates that the effects of pioglitazone on
visfatin expression could be cell- or tissue-specific.
PPARs are a member of the nuclear hormone receptor
superfamily consisting of three isotypes: PPAR-α, PPAR-δ
and PPAR-γ. All three PPAR isotypes are expressed in the placenta
(30). The expression levels of
PPAR-γ were downregulated in the placenta of females with GDM;
however, that of PPAR-δ remained unchanged. A previous
study demonstrated that PPAR-α agonist fenofibrate stimulated the
expression of PPAR-α and induced the mRNA expression of visfatin in
visceral fat deposits of Otsuka Long-Evans Tokushima fatty rats
(31). The protein expression
levels of PPAR-α in placenta were reduced, whereas, the mRNA levels
were unchanged in females with GDM (24). Therefore, PPAR-γ was selected as
the potential target in the present study. A chemical agonist of
PPARs was easier to obtain compared with overexpression vectors
and/or PPAR small interfering RNAs, thus, pioglitazone was used in
the present study.
The results of the present study demonstrated that
IL-6 downregulated PPAR-γ and visfatin at the transcriptional
level; the expression levels of visfatin and PPAR-γ were strongly
correlated in BeWo cells. In addition, visfatin and PPAR-γ
expression was reduced as the concentration of IL-6 increased
between 0 and 5 ng/ml in BeWo cells, but their expression levels
increased as the concentration of IL-6 reached 10 ng/ml; however,
the expression levels of visfatin and PPAR-γ were significantly
decreased compared with control group. The results of the present
study suggested that the inhibitory effect of IL-6 on visfatin and
PPAR-γ did not occur in a dose-dependent manner. The underlying
mechanisms require further investigation.
Following treatment with pioglitazone, the mRNA and
protein expression levels of visfatin were significantly increased
in a dose-dependent manner in the present study. Therefore,
visfatin may serve within trophoblastic cells as a protective
factor, and may be involved in the regulation of energy metabolism
and inflammatory reactions. It is well-established that
visfatin/Nampt promotes intracellular NAD+ synthesis and
induces the activation of sirtuin1 (12,32–34).
Furthermore, placental visfatin/Nampt may additionally serve as the
Nampt enzyme within trophoblasts (34). The overexpression of visfatin/Nampt
may increase intracellular NAD+ and stimulate sirtuin1,
consequently affecting DNA repair, protecting telomeres and
reducing inflammation (34).
NAD+ is an important coenzyme in numerous
oxidation-reduction reactions (34). NAD+ accepts and donates
electrons in reactions, thus leading to the production of ATP,
which is required by cells for most energy-consuming processes
(34). Sirtuin1, additionally
termed NAD+-dependent deacetylase, is a negative
regulator of pro-inflammatory cytokines (35–37).
In summary, visfatin/Nampt may be the regulatory factor underlying
the survival of trophoblastic cells and maintenance of placental
function in a hostile environment, including the inflammation
response or hyperglycemia. Upon inflammation, reduced expression of
visfatin may lead to disrupted placental function and increased
risk of abortion, preterm birth, and/or fetal distress (33).
However, treatment with the PPAR-γ agonist
pioglitazone induced the mRNA and protein expression of visfatin by
increasing the secretion of visfatin and offsetting the inhibitory
effect of IL-6 at the mRNA level. The findings of the present study
suggested that visfatin was a potential target gene of PPAR-γ in
BeWo cells, and pioglitazone upregulated the expression level of
visfatin by activating the PPAR-γ pathway. Additionally, the mRNA
expression levels of PPAR-γ remained unchanged following the
treatment of pioglitazone. Subsequent to being activated by its
ligands, PPAR-γ forms a heterodimer with the retinoic X receptor,
which transactivates the PPAR-response elements of target genes
involved in insulin sensitivity, glucose metabolism and immune
responses (12,38,39).
Therefore, upon activation, PPAR-γ upregulates its target genes,
including visfatin, rather than promoting its own transcription.
Furthermore, treatment with pioglitazone alone for 48 h resulted in
increased secretion of visfatin compared with IL-6 alone or in
combination with pioglitazone; however, the expression levels of
visfatin in all groups remained low, indicating that visfatin may
serve as an autocrine/paracrine factor rather than a typical
endocrine agent in trophoblastic cells. Therefore, the present
study hypothesized that visfatin was able to exert local effects in
the placenta. Additionally, the activation of PPAR-γ suppressed the
activity of nuclear factor-κB, consequently regulating the balance
between cytokine production and lipid metabolism (40); however, the present study indicated
that treatment with visfatin for 48 h did not alter the secretion
levels of inflammatory cytokines, including IL-6 and TNF-α (data
not shown). In summary, the results of the present study provide
novel insight into the roles of visfatin in trophoblastic cells.
Placental visfatin may serve as the Nampt enzyme by increasing
NAD+ biosynthesis and activating sirtuin1, consequently
functioning as an anti-inflammatory factor rather than a
pro-inflammatory factor, and serving an important role in
maintaining the energy metabolism of trophoblastic cells and the
function of the placenta.
In conclusion, the findings of the present study
suggested that IL-6 downregulated visfatin and PPAR-γ at the
transcriptional level; in addition, activation of PPAR-γ induced
the expression of visfatin in BeWo cells. The thiazolidinedione
pioglitazone may promote the energy metabolism of trophoblastic
cells, maintain the function of the placenta and improve the
outcome of pregnancy; however, further study is required to confirm
whether visfatin may be a target gene of PPAR-γ in BeWo cells. The
roles of visfatin as a Nampt enzyme, as well as its downstream
signaling pathway and underlying mechanisms in trophoblastic cells
or placenta, require further investigation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Hebei Natural
Science Foundation (grant no. H2016307035).
Availability of data and materials
The datasets used and/or analyzed in the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YZ and SL conducted the experiments, were involved
in data collection and drafted the manuscript. YH and WH performed
the statistical analyses and contributed to the study design. HL
and LL helped with the literature review and collected the data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GDM
|
gestational diabetes mellitus
|
|
Nampt
|
nicotinamide
phosphoribosyltransferase
|
|
PPAR-γ
|
peroxisome proliferator-activated
receptor γ
|
|
IL-6
|
interleukin 6
|
|
TNF-α
|
tumor necrosis factor-α
|
References
|
1
|
Shi KL, Qian JY, Qi L, Mao DB, Chen Y, Zhu
Y and Guo XG: Atorvastatin antagonizes the visfatin-induced
expression of inflammatory mediators via the upregulation of NF-κB
activation in HCAECs. Oncol Lett. 12:1438–1444. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kocełak P, Olszanecka-Glinianowicz M,
Owczarek AJ, Krupa W, Obirek P, Bożentowicz-Wikarek M, Brzozowska
A, Mossakowska M, Zdrojewski T, Skalska A, et al: Plasma
visfatin/nicotinamide phosphoribosyltransferase (visfatin/NAMPT)
concentration in elderly subjects with metabolic syndrome. Pol Arch
Med Wewn. 125:402–413. 2015.PubMed/NCBI
|
|
3
|
Owczarek AJ, Olszanecka-Glinianowicz M,
Kocełak P, Bożentowicz-Wikarek M, Brzozowska A, Mossakowska M,
Puzianowska-Kuźnicka M, Grodzicki T, Więcek A and Chudek J: The
relationship between circulating visfatin/nicotinamide
phosphoribosyltransferase, obesity, inflammation and lipids profile
in elderly population, determined by structural equation modeling.
Scand J Clin Lab Invest. 76:632–640. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma Y, Cheng Y, Wang J, Cheng H, Zhou S and
Li X: The changes of visfatin in serum and its expression in fat
and placental tissue in pregnant women with gestational diabetes.
Diabetes Res Clin Pract. 90:60–65. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Porter B, Babbar S, Ye SQ and Maulik D:
The role of nicotinamide phosphoribosyltransferase in pregnancy: A
review. Am J Perinatol. 33:1327–1336. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun L, Chen S, Gao H, Ren L and Song G:
Visfatin induces the apoptosis of endothelial progenitor cells via
the induction of pro-inflammatory mediators through the NF-κB
pathway. Int J Mol Med. 40:637–646. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vejrazkova D, Lischkova O, Vankova M,
Stanicka S, Vrbikova J, Lukasova P, Vcelak J, Vacinova G and
Bendlova B: Distinct response of fat and gastrointestinal tissue to
glucose in gestational diabetes mellitus and polycystic ovary
syndrome. Physiol Res. 66:283–292. 2017.PubMed/NCBI
|
|
8
|
Liang Z, Wu Y, Xu J, Fang Q and Chen D:
Correlations of serum visfatin and metabolisms of glucose and lipid
in women with gestational diabetes mellitus. J Diabetes Investig.
7:247–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hosseinzadeh-Attar MJ, Golpaie A, Foroughi
M, Hosseinpanah F, Zahediasl S and Azizi F: The relationship
between visfatin and serum concentrations of c-reactive protein,
interleukin 6 in patients with metabolic syndrome. J Endocrinol
Invest. 39:917–922. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moschen AR, Kaser A, Enrich B, Mosheimer
B, Theurl M, Niederegger H and Tilg H: Visfatin, an adipocytokine
with proinflammatory and immunomodulating properties. J Immunol.
178:1748–1758. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kendal CE and Bryant-Greenwood GD:
Pre-B-cell colony-enhancing factor (Pbef/Visfatin) gene expression
is modulated by NF-kappaB and AP-1 in human amniotic epithelial
cells. Placenta. 28:305–314. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mayi TH, Duhem C, Copin C, Bouhlel MA,
Rigamonti E, Pattou F, Staels B and Chinetti-Gbaguidi G: Visfatin
is induced by peroxisome proliferator-activated receptor gamma in
human macrophages. FEBS J. 277:3308–3320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stromsdorfer KL, Yamaguchi S, Yoon MJ,
Moseley AC, Franczyk MP, Kelly SC, Qi N, Imai S and Yoshino J:
NAMPT-mediated NAD(+) biosynthesis in adipocytes regulates adipose
tissue function and multi-organ insulin sensitivity in mice. Cell
Rep. 16:1851–1860. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim HS, Han SY, Sung HY, Park SH, Kang MK,
Han SJ and Kang YH: Blockade of visfatin induction by oleanolic
acid via disturbing Il-6-TRAF6-NF-κB signaling of adipocytes. Exp
Biol Med (Maywood). 239:284–292. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lv Q, Wang Y, Wang W, Wang L and Zhou X:
Effect of pioglitazone on visfatin expression in 3T3-L1 adipocytes
and SD rats. Endocr Res. 34:130–141. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mrizak I, Grissa O, Henault B, Fekih M,
Bouslema A, Boumaiza I, Zaouali M, Tabka Z and Khan NA: Placental
infiltration of inflammatory markers in gestational diabetic women.
Gen Physiol Biophys. 33:169–176. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang J, Chi H, Xiao H, Tian X, Wang Y,
Yun X and Xu Y: Interleukin 6 (IL-6) and tumor necrosis factor α
(TNF-α) single nucleotide polymorphisms (SNPs), inflammation and
metabolism in gestational diabetes mellitus in inner Mongolia. Med
Sci Monit. 23:4149–4157. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Telejko B, Kuzmicki M, Zonenberg A,
Szamatowicz J, Wawrusiewicz-Kurylonek N, Nikolajuk A, Kretowski A
and Gorska M: Visfatin in gestational diabetes: Serum level and
mRNA expression in fat and placental tissue. Diabetes Res Clin
Pract. 84:68–75. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kralisch S, Klein J, Lossner U, Bluher M,
Paschke R, Stumvoll M and Fasshauer M: Interlenkin-6 is a negative
regulator of visfatin gene expression in 3T3-L1 adipocyts. Am J
Physicol Endocrinol Metab. 289:E586–E590. 2005. View Article : Google Scholar
|
|
20
|
McGee KC, Harte AL, da Silva NF, Al-Daghri
N, Creely SJ, Kusminski CM, Tripathi G, Levick PL, Khanolkar M,
Evans M, et al: Visfatin is regulated by rosiglitazone in Type 2
diabetes mellitus and influenced by NFκB and JNK in human abdominal
subcutaneous adipocytes. PLoS One. 6:e202872011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ognjanovic S, Bao S, Yamamoto SY,
Garibay-Tupas J, Samal B and Bryant-Greenwood GD: Genomic
organization of the gene coding for human pre-B-cell colony
enhancing factor and expression in human fetal membranes. J Mol
Endocrinol. 26:107–117. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahmadian M, Suh JM, Hah N, Liddle C,
Atkins AR, Downes M and Evans RM: PPARγ signaling and metabolism:
The good, the bad and the future. Nat Med. 19:557–566. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Charrier A, Wang L, Stephenson EJ, Ghanta
SV, Ko CW, Croniger CM, Bridges D and Buchner DA: Zinc finger
protein 407 overexpression upregulates PPAR target gene expression
and improves glucose homeostasis in mice. Am J Physiol Endocrinol
Metab. 311:E869–E880. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Holdsworth-Carson SJ, Lim R, Mitton A,
Whitehead C, Rice GE, Permezel M and Lappas M: Peroxisome
proliferator-activated receptors are altered in pathologies of the
human placenta: Gestational diabetes mellitus, intrauterine growth
restriction and preeclampsia. Placenta. 31:222–229. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suwaki N, Masuyama H, Masumoto A, Takamoto
N and Hiramatsu Y: Expression and potential role of peroxisome
proliferator-activated receptor gamma in the placenta of diabetic
pregnancy. Placenta. 28:315–323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Capobianco E, Jawerbaum A, Romanini MC,
White V, Pustovrh C, Higa R, Martinez N, Mugnaini MT, Soñez C and
Gonzalez E: 15-Deoxy-∆12,14-prostaglandin J2 and peroxisome
proliferator-activated receptor γ (PPARγ) levels in term placental
tissues from control and diabetic rats: Modulatory effects of a
PPARγ agonist on nitridergic and lipid placental metabolism. Reprod
Fertil Dev. 17:423–433. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Szklanna PB, Wynne K, Nolan M, Egan K,
Áinle FN and Maguire PB: Comparative proteomic analysis of
trophoblast cell models reveals their differential phenotypes,
potential uses and limitations. Proteomics. Mar 20–2017;(Epub ahead
of print).
|
|
28
|
Orendi K, Gauster M, Moser G, Meiri H and
Huppertz B: The choriocarcinoma cell line BeWo: Syncytial fusion
and expression of syncytium-specific proteins. Reproduction.
140:759–766. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schimittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Barak Y, Sadovsky Y and Shalom-Barak T:
PPAR signaling in placental development and function. PPAR Res.
2008:1420822008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Choi KC, Ryu OH, Lee KW, Kim HY, Seo JA,
Kim SG, Kim NH, Choi DS, Baik SH and Choi KM: Effect of PPAR-alpha
and -gamma agonist on the expression of visfatin, adiponectin, and
TNF-alpha in visceral fat of OLETF rats. Biochem Biophys Res
Commun. 336:747–753. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bermudez B, Dahl TB, Medina I, Groeneweg
M, Holm S, Montserrat-de la Paz S, Rousch M, Otten J, Herias V,
Varela LM, et al: Leukocyte overexpression of intracellular NAMPT
attenuates atherosclerosis by regulating PPARγ-dependent monocyte
differentiation and function. Arterioscler Thromb Vasc Biol.
37:1157–1167. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang SJ, Choi JM, Kim L, Park SE, Rhee EJ,
Lee WY, Oh KW, Park SW and Park CY: Nicotinamide improves glucose
metabolism and affects the hepatic NAD-sirtuin pathway in a rodent
model of obesity and type 2 diabetes. J Nutr Biochem. 25:66–72.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsai PJ, Davis J, Thompson K and
Bryant-Greenwood G: Visfatin/Nampt and SIRT1: Roles in postterm
delivery in pregnancies associated with obesity. Reprod Sci.
22:1028–1036. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin QQ, Geng YW, Jiang ZW and Tian ZJ:
SIRT1 regulates lipopolysaccharide-induced CD40 expression in renal
medullary collecting duct cells by suppressing the TLR4-NF-κB
signaling pathway. Life Sci. 170:100–107. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li L, Sun Q, Li Y, Yang Y, Yang Y, Chang
T, Man M and Zheng L: Overexpression of sirt1 induced by
resveratrol and inhibitor of miR-204 suppresses activation and
proliferation of microglia. J Mol Neurosci. 56:858–867. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tian Y, Ma J, Wang W, Zhang L, Xu J, Wang
K and Li D: Resveratrol supplement inhibited the NF-κB inflammation
pathway through activating AMPKα-SIRT1 pathway in mice with fatty
liver. Mol Cell Biochem. 422:75–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chigurupati S, Dhanaraj SA and Balakumar
P: A step ahead of PPARγ full agonists to PPARγ partial agonists:
Therapeutic perspectives in the management of diabetic insulin
resistance. Eur J Pharmacol. 755:50–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mayi TH, Rigamonti E, Pattou F, Staels B
and Chinetti-Gbaguidi G: Liver X receptor (LXR) activation
negatively regulates visfatin expression in macrophages. Biochem
Biophys Res Commun. 404:458–462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kirwan AM, Lenighan YM, O'Reilly ME,
McGillicuddy FC and Roche HM: Nutritional modulation of metabolic
inflammation. Biochem Soc Trans. 45:979–985. 2017. View Article : Google Scholar : PubMed/NCBI
|