Introduction
Hearing loss is a common communication disorder that
approximately 360 million people have worldwide. There are two
types of hearing loss: Non-syndromic and syndromic (1). The molecular mechanism behind
deafness has not been well characterized; but it is generally
thought to occur through a combination of genetic, environmental
and lifestyle factors. In fact, the genetic aspects may account for
>50% of cases of deafness (2).
Specifically, variants in mitochondrial (mt)DNA have been reported
to be associated with aminoglycoside-induced hearing loss (3,4).
Human mtDNA is a 16,569 bp molecule that encodes 13
oxidative phosphorylation (OXPHOS)-related protein subunits, 2
ribosomal (r)RNAs, as well as 22 transfer (t)RNAs (5). Of the mutations in human mtDNA,
A1555G is one of the most common pathogenic mutations associated
with aminoglycoside ototoxicity (6). However, increasing evidence has shown
that the A1555G mutation cannot directly lead to hearing loss, as
among pedigrees with maternally inherited deafness, a certain
number of members (despite carrying the A1555G mutation) exhibit
different levels of hearing, ranging from normal to profound
hearing loss (7–11). Therefore, aminoglycosides,
nuclear-encoded genes and the mtDNA genetic background may actively
function in the occurrence of deafness (12,13).
In the present study, to determine the contribution
of mtDNA mutations/polymorphisms to the occurrence of deafness,
mtDNA gene mutations were screened from a cohort of deaf patients.
The present study reported the molecular features of a deaf Chinese
pedigree. Sequence analysis of the mtDNA genes identified the
co-existence of tRNAAsp A7551G and 12S rRNA A1555G
mutations.
Materials and methods
Samples and clinical examinations
The current study was approved by the Ethics
Committee of Zhoushan Hospital, and each participant provided
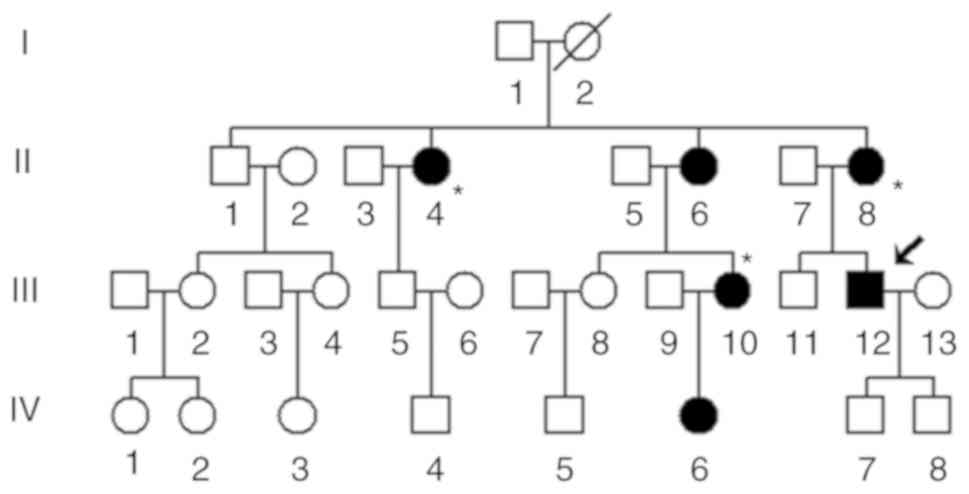
written informed consent. A four-generation Han Chinese family
(Fig. 1), comprising 30
individuals and 6 deaf patients, was enrolled from the Department
of Otolaryngology, Zhoushan Hospital, the age, sex of each patient
was summarized in Table I. First,
a comprehensive physical examination of each patient was performed,
including the determination of whether they were taking
aminoglycosides. In addition, the levels of hearing loss were
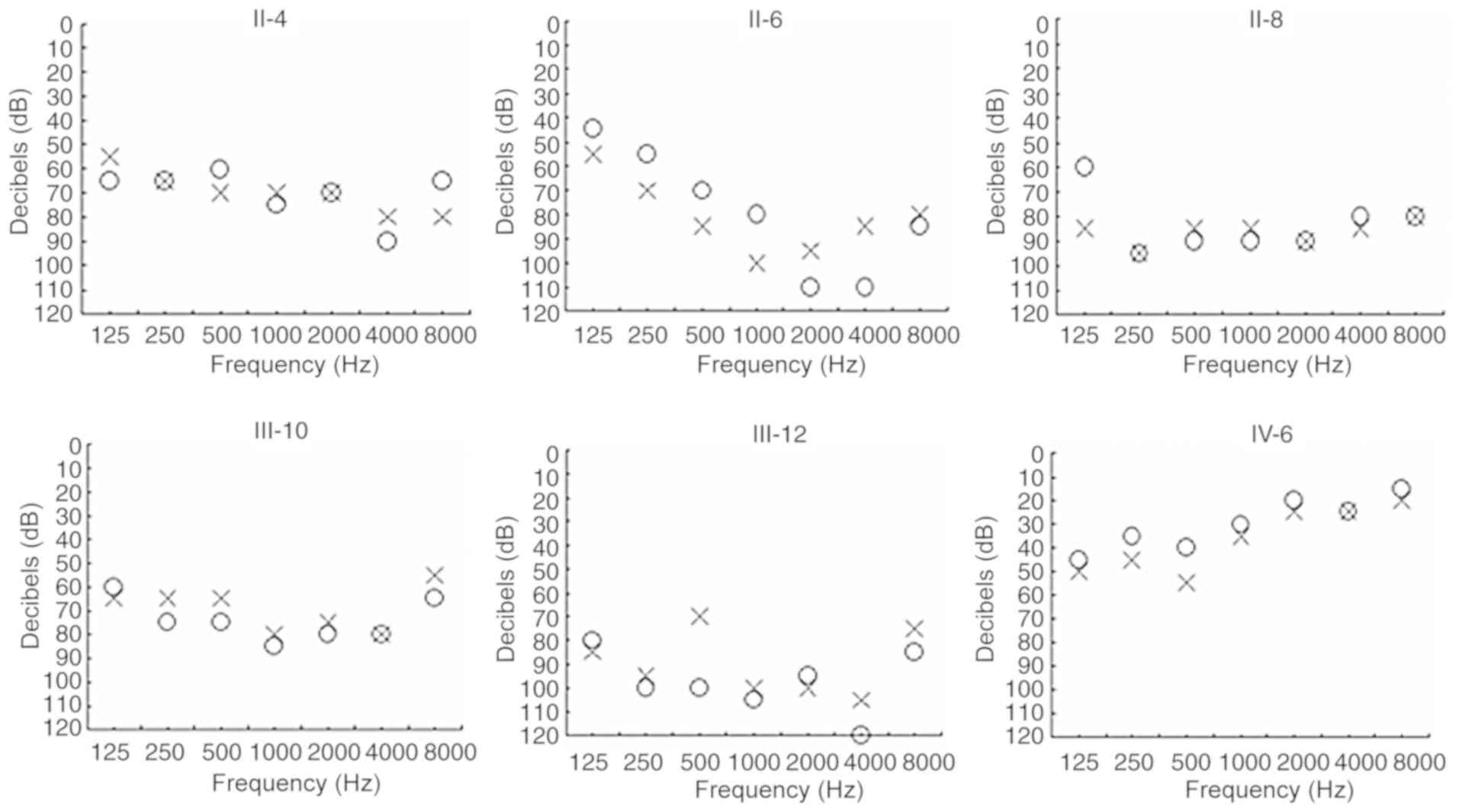
tested in each participant using pure-tone audiometry (PTA) as
described (3). The average PTA
level was measured using the total audiometric thresholds at 125,
250, 500, 1,000, 2,000, 4,000 and 8,000 Hz. The levels of hearing
loss were divided into the following degrees: i) Normal hearing
(PTA <26 dB); ii) mild hearing loss (PTA 26–40 dB); iii)
moderate hearing loss (PTA 41–70 dB); iv) severe hearing loss (PTA
71–90 dB); and v) profound hearing loss (PTA >90 dB). In
addition, 200 healthy subjects were enrolled from the Physical
Examination Center of Zhoushan Hospital. The inclusion criteria for
the healthy subjects were that they were age- and sex-matched,
healthy, and had no family history of mitochondrial disorders.
 | Table I.Summary of the clinical and molecular
data of several deafness patients of a Chinese family. |
Table I.
Summary of the clinical and molecular
data of several deafness patients of a Chinese family.
| Subject | Sex | Age at test
(year) | Age at onset
(year) | PTA (dB) right
ear | PTA (dB) left
ear | Level of hearing
loss |
|---|
| II-4 | Female | 68 | 60 | 68 | 70 | Severe |
| II-6 | Female | 59 | 55 | 87 | 65 | Severe |
| II-8 | Female | 61 | 58 | 90 | 87 | Severe |
| III-10 | Female | 33 | 28 | 80 | 73 | Severe |
| III-12 | Male | 35 | 30 | 100 | 90 | Profound |
| IV-6 | Female | 8 | 4 | 30 | 38 | Mild |
Screening for mutations in mtDNA
genes
To analyze the mtDNA mutations, PCR and direct
sequencing analysis were performed. The genomic DNA was first
isolated from blood samples using Puregene DNA Isolation kits
(Gentra Systems; Thermo Fisher Scientific, Inc.). The complete
mtDNA of the matrilineal relatives from this family were then PCR
amplified using 24 oligonucleotides, as previously described
(14). The PCR product was
purified and analyzed by Sanger sequencing as described (3). The data were then compared with the
updated mitochondrial genome sequence to detect mtDNA mutations
(GenBank accession no. NC_001807) (15).
Genotyping analysis of gap junction
protein β 2 (GJB2) variants
Variants in the GJB2 gene were further
investigated in matrilineal relatives. For amplification of the
GJB2 coding region, PCR was performed using the following
primers: Forward, 5′-TTGGTGTTTGCTCAGGAAGA-3′ and reverse,
5′-GGCCTACAGGGGTTTCAAAT-3′, as previously described (16). PCR fragments were sequenced and the
data were analyzed with the wild-type GJB2 sequence
(accession no. NM_004004.5).
Statistical analysis
SPSS 19.0 software (IBM Corp., Armonk, NY, USA) and
Student's t-test were used to perform the statistical analysis; we
compared the significance between deaf patients carrying
mitochondrial A1555G and A7551G mutations and control subjects.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Pedigree information
The proband (III-12), aged 35, lived in Zhoushan
(Zhejiang, China). He attended to the Otology Clinic of Zhoushan
Hospital for deafness treatment. Clinical examination suggested
that he had manifested bilateral hearing loss at 30 years old. As
shown in Fig. 2, it was noted that
the proband exhibited profound hearing loss (right ear, 100 dB;
left ear, 90 dB). However, he did not have any associated medical
history.
In addition, a physical examination was performed
for each deaf patient from this pedigree, including the level of
hearing loss, aminoglycosides usage, as well as other disorders. It
was revealed that the family members did not have other common
disorders, such as cardiovascular events, diabetes mellitus, visual
impairment or neurological disorders. It was also noted that
several individuals had a history of using aminoglycosides, such as
individual II-8. She received kanamycin for fever when she was 58
years old, unfortunately, three weeks later, she developed
bilateral deafness. As presented in Table I and Fig. 2, a variable degree of hearing loss
was determined in this Chinese pedigree, ranging from mild (IV-6)
to profound (III-12). Furthermore, the deafness exhibited a pattern
of maternal transmission (Fig.
1).
Mutational analysis of mitochondrial
genome
Owing to the apparent maternal transmission of
deafness in this family, the proband's complete mtDNA was analyzed
for mutations, together with those of other matrilineal relatives.
A total of 24 overlapping DNA fragments spanning the 16,569 bp of
mtDNA were PCR amplified and sequenced. It was shown that compared
with the standard mitochondrial genome sequences, the proband and
other matrilineal relatives exhibited 24 genetic polymorphisms
(Table II), which were as
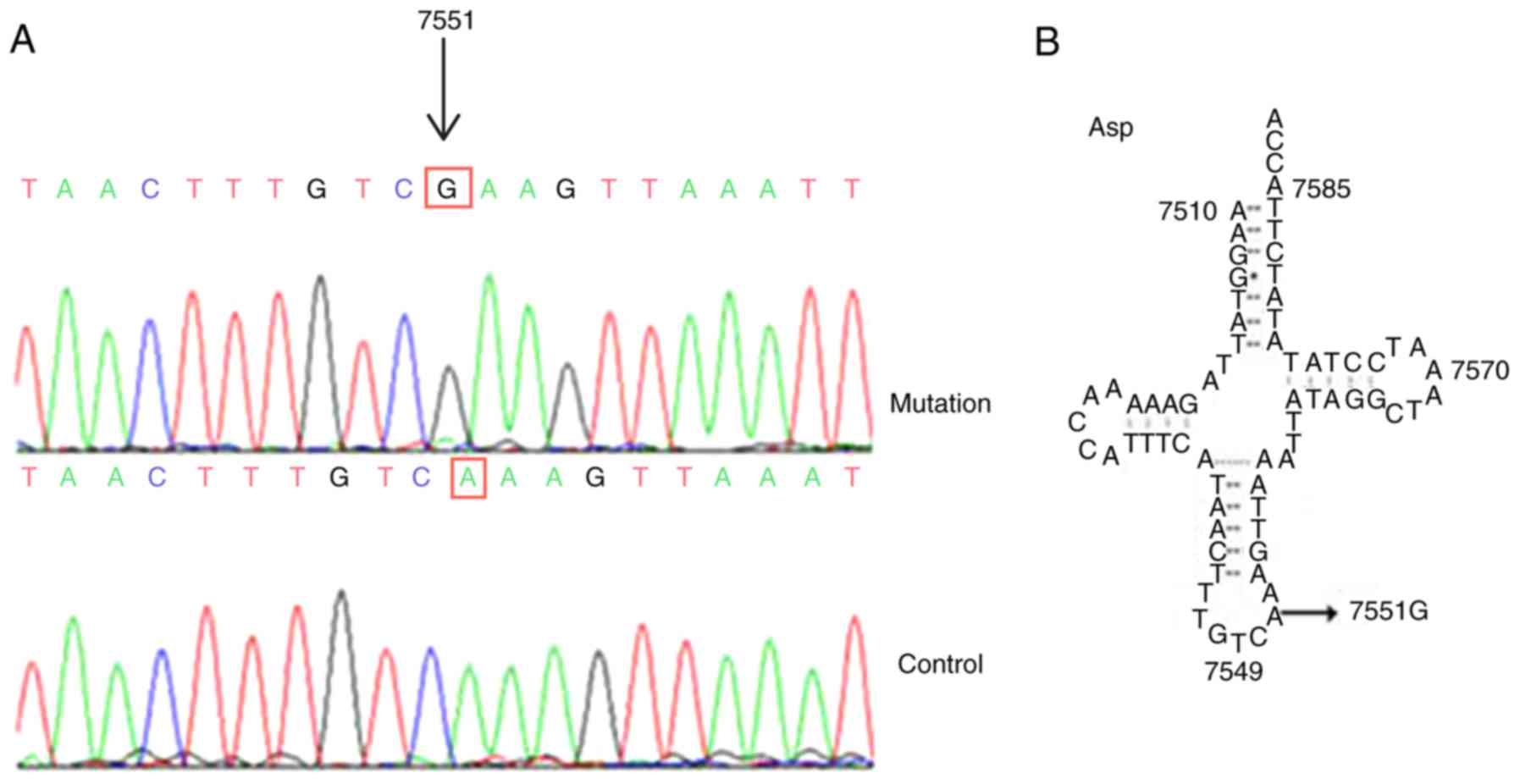
follows: Seven variants in the D-loop gene, three in the 12S rRNA
gene, two in the 16S rRNA gene and one in a tRNA gene (Fig. 3A and B), as well as a nine bp
deletion in the non-coding region between tRNALys and
cytochrome C oxidase II (CO2). These polymorphisms have all
been previously reported (www.mitomap.org). The other variants were located in
oxidative phosphorylation (OXPHOS) encoding genes. In addition, six
missense mutations were identified, namely, the NADH:Ubiquinone
(ND2) C5178A (Leu→Met) and T5442C (Phe→Leu) mutations, the
ATPase 6 A8701G (Thr→Ala) and A8860G (Thr→Ala) mutations,
the NADH:Ubiquinone oxidoreductase core subunit 5 (ND5)
G13928C (Ser→Thr) mutation, and the cytochrome B (Cyt b)
A15326G (Thr→Ala) mutation. Phylogenetic analysis of these
mutations in 11 vertebrates was performed, including mouse
(17), bovine (18), and Xenopus laevis (19) (Table
III). The results revealed that only the A1555G and A7551G
mutations exhibited a high level of conservation. Further, the
A1555G and A7551G mutations were not detected in the 200 control
subjects, suggesting that these mutations may have a functional
impact associated with deafness (P<0.05; Table IV). Notably, the homoplasmic
A7551G mutation occurred at position 37 in the anticodon stem of
tRNAAsp, which may have caused tRNA metabolism
failure.
| Table II.Sequence analysis of the family's
complete mitochondrial genome. |
Table II.
Sequence analysis of the family's
complete mitochondrial genome.
| Gene | Position | Alternation | Conservation
(H/B/M/X) |
|---|
| D-loop | 73 | A to G |
|
|
| 152 | T to C |
|
|
| 310 | T to CTC |
|
|
| 489 | T to C |
|
|
| 16189 | T to C |
|
|
| 16261 | C to T |
|
|
| 16519 | T to C |
|
| 12S rRNA | 750 | A to G | A/A/G/- |
|
| 1438 | A to G | A/A/A/G |
|
| 1555 | A to G | A/A/A/A |
| 16S rRNA | 2706 | A to G | A/G/A/A |
|
| 3010 | G to A | G/G/A/A |
| ND1 | 3970 | C to T |
|
| ND2 | 5178 | C to A (Leu to
Met) | L/T/T/T |
|
| 5442 | T to C (Phe to
Leu) | F/F/M/L |
| CO1 | 7028 | C to T |
|
|
tRNAAsp | 7551 | A to G | A/A/A/A |
| NC_7 | 8271-8279 | 9-bp del |
|
| A6 | 8701 | A to G (Thr to
Ala) | T/S/L/Q |
|
| 8860 | A to G (Thr to
Ala) | T/A/A/T |
| ND4 | 11719 | G to A |
|
| ND5 | 13928 | G to C (Ser to
Thr) | S/T/S/T |
| Cyt b | 15301 | G to A |
|
|
| 15326 | A to G (Thr to
Ala) | T/M/I/I |
| Table III.Alignment of mt-tRNAAsp
gene from different species, arrow indicated the position 37,
corresponding to the A7551G mutation. |
Table III.
Alignment of mt-tRNAAsp
gene from different species, arrow indicated the position 37,
corresponding to the A7551G mutation.
| Organism | Acc-stem | D-stem | D-loo | D-stem | Ac-Stem | Anti CD-loop | Ac-stem | V-region | T-stem | T-loop | T-stem | Acc-stem |
|---|
| Homo
sapiens | AAGGTATTA | GAAA | AACCA | TTTCA | TAACT | TTGTCAA | AGTTA | AATT | ATAGG | CTAAAT | CCTAT | ATATCTTA |
| Gorilla
gorilla | AAGATATTA | GAAA | AACTA | TTTCA | TAACT | TTGTCAA | GGTTA | AATT | ACGGG | TTAAAC | CCCGT | ATATCTTA |
| Lemur
catta | AAGATATTA | GTAA | AATCA | TTACA | TAACT | TTGTCAA | AGTTA | ATTT | ATAGG | TTAAACC | CCTAT | GTATCTTT |
| Pan
troglodytes | AAGATATTA | GAAA | AACTA | TTTCA | TAACT | TTGTCAA | AGTTA | AATT | ACAGG | TTAACC | CCCGT | ATATCTTA |
| Hylobates
lar | AAGATATTA | GAAA | AACTA | TTTCA | TAACT | TTGTCAA | AGTTA | AGTT | ACAGG | TTCAAAC | CCCGT | ATATCTTA |
| Trachypithecus
obscurus | GAGATATTA | GAAA | AATTA | TTTCA | TAGCT | TTGTCAA | AGCTA | AATT | ATAGG | ACATAT | CCTAT | ATATCTTA |
| Muntiacus
reevesi | GAGGTATTA | GTAA | AATA | TTACA | TAACT | TTGTCAA | AGTTA | AGTT | ACAGG | TGAAAAT | CCCGT | ATATCTCA |
| Mus
musculus | AAGATATTA | GTAA | AATCAA | TTACA | TAACT | TTGTCAA | AGTTA | AATT | ATAGA | TCAATAA | TCTAT | ATATCTTA |
| Balaenoptera
bonaerensis | GAGATATTA | GTAA | AATC | TTATA | TAACT | TTGTCAA | AGTTA | AGTT | ACAAG | TGAGAAT | CCTGT | ATATCTCC |
| Cynocephalus
variegatus | GAGATATTA | GTAA | AAACA | TTACA | TGACT | TCGTCAG | AGTCA | ACTT | ATAGG | TCTAAT | CCTTT | ATATCTTT |
| Pongo pygmaeus
abelii | GAGATACTA | GAAA | AACCA | TTTCA | TGACT | TTGTCAA | AGTTA | AGTT | ACAGG | CCAAAC | CCTGT | GTATCTTA |
| Table IV.Mitochondrial A1555G and A7551G
mutations in affected individuals and controls. |
Table IV.
Mitochondrial A1555G and A7551G
mutations in affected individuals and controls.
| Gene | Position | Replacement | Number of patients
(%) | Number of 200
controls (%) | P-value |
|---|
| 12S rRNA | 1555 | A to G | 6 (60) | 0 (0) | <0.05 |
|
tRNAAsp | 7551 | A to G | 6 (60) | 0 (0) | <0.05 |
Screening for GJB2 mutations
To examine whether the GJB2 gene had critical
function in deafness expression, the coding regions of GJB2
were analyzed. However, no functional variants in this gene were
identified (data not shown).
Discussion
The present paper reported the molecular features of
a Chinese pedigree with maternally inherited hearing loss. Overall,
a total of six matrilineal relatives of this family exhibited
hearing loss of various degrees. Besides the deafness phenotype, no
other clinical disorders were detected in this family. As the
deafness was shown to be maternally transmitted in this family, the
mtDNA variants of the proband and other matrilineal relatives were
analyzed in this pedigree. There were six deaf patients in total,
while the number of matrilineal relatives was nine, therefore, the
penetrance of hearing loss was 6/9=66.7% when the aminoglycoside
was included, however, in the absence of aminoglycoside, there were
three deaf patients, thus, the penetrance of hearing loss was
3/9=33.3%.
The maternal inheritance of deafness suggested that
mitochondrial dysfunction may be the molecular basis of this
hearing loss. Therefore, the sequence variants of mtDNA from the
matrilineal relatives and the proband were detected. Screening of
the mtDNA genes from the members of this family identified 24
genetic polymorphisms, belonging to human mitochondrial haplogroup
B5a (20). Among these variants,
the homoplasmic tRNAAsp A7551G mutation was of
particular interest. At the molecular level, this mutation was
localized at the 3′ end to the anticodon stem (position 37). The
nucleotide at position 37 in tRNAAsp is highly conserved
among different vertebrates (21).
In fact, A37 in tRNAs is often chemically modified (by thiolation
or methylation) (22), therefore,
the A7551G mutation may contribute to maintenance of the level of
tRNAAsp (23). Notably,
the well-known A4435G mutation occurring at the same position in
the tRNAMet gene was previously found to be a risk
factor for hypertension (24) and
Leber's hereditary optic neuropathy (LHON) (25). Thus, the A7551G mutation, which is
similar to the A4435G mutation, may have also resulted in failure
of tRNA metabolism. A previous study also reported that the A7551G
mutation, combined with GJB2 235delC, was involved in the
progression of deafness in a Chinese family (26). Consequently, a decrease in the
tRNAAsp steady-state level may affect mitochondrial
protein translation. Thus, the A7551G mutation would lead to
defects in the synthesis of components of the mitochondrial
respiration chain and subsequently, a decline in ATP production and
a decrease in reactive oxygen species (ROS), which was similar to
the tRNAIle A4317G mutation (27). These mitochondrial dysfunctions,
caused by the A7551G mutation, may be involved in the pathogenesis
of hearing loss (28,29). However, the penetrance of hearing
loss and haplogroup in the present study differed from those
reported by Wu et al (26).
The current study identified the co-existence of 12S rRNA A1555G
and tRNAAsp A7551G mutations in a Chinese family,
indicating that the A7551G mutation may increase the penetrance and
expression of the deafness-associated A1555G mutation.
However, the incomplete penetrance of hearing loss,
the homoplasmic form of the mtDNA mutations, the variable degrees
of hearing impairment, as well as the absence of any functional
variants in the GJB2 gene suggested that the A7551G mutation
itself was insufficient to produce the clinical phenotypes.
Therefore, other risk-modifying factors, such as nuclear DNA, the
use of aminoglycosides and environmental factors may have
contributed to the clinical expression of the deafness-associated
A1555G mutation in this family. Taking together, the findings of
the present study indicated that the tRNAAsp A7551G
mutation may have contributed to the clinical manifestation of
hearing loss-associated mitochondrial A1555G mutation in this
family. The main limitations of the current study was the
relatively small sample size. Further assays including more
deafness patients are required to verify the results. In addition,
functional experiments, such as the measurement of ROS production,
ATP and oxygen consumption levels, should be performed.
Acknowledgements
We thank Liwen Bianji for editing the English text
of a draft of this manuscript.
Funding
The present study was supported by the Zhoushan
Ministry of Public Health (grant no. 2017B16).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
JZ, BL, W-WX and BF made substantial contributions
to the design of the present study. G-WH collected the samples and
performed the clinical analysis. X-XD performed the molecular and
statistical analysis. G-WH wrote the paper. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The current study was approved by the Ethics
Committee of Zhoushan Hospital, and each participant provided
written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fitzpatrick EM, Whittingham J and
Durieux-Smith A: Mild bilateral and unilateral hearing loss in
childhood: A 20-year view of hearing characteristics, and
audiologic practices before and after newborn hearing screening.
Ear Hear. 35:10–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ghogomu N, Umansky A and Lieu JE:
Epidemiology of unilateral sensorineural hearing loss with
universal newborn hearing screening. Laryngoscope. 124:295–300.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ding Y, Xia BH, Liu Q, Li MY, Huang SX and
Zhuo GC: Allele-specific PCR for detecting the deafness-associated
mitochondrial 12S rRNA mutations. Gene. 591:148–152. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ding Y, Leng J, Fan F, Xia B and Xu P: The
role of mitochondrial DNA mutations in hearing loss. Biochem Genet.
51:588–602. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Anderson S, Bankier AT, Barrell BG, de
Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA,
Sanger F, et al: Sequence and organization of the human
mitochondrial genome. Nature. 290:457–465. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prezant TR, Agapian JV, Bohlman MC, Bu X,
Oztas S, Qiu WQ, Arnos KS, Cortopassi GA, Jaber L, Rotter JI, et
al: Mitochondrial ribosomal RNA mutation associated with both
antibiotic-induced and non-syndromic deafness. Nat Genet.
4:289–294. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guan MX: Mitochondrial 12S rRNA mutations
associated with aminoglycoside ototoxicity. Mitochondrion.
11:237–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park MK, Sagong B, Lee JD, Bae SH, Lee B,
Choi KS, Choo YS, Lee KY and Kim UK: A1555G homoplasmic mutation
from A1555G heteroplasmic mother with pendred syndrome. Int J
Pediatr Otorhinolaryngol. 78:1996–1999. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Skou AS, Tranebjærg L, Jensen T and Hasle
H: Mitochondrial 12S ribosomal RNA A1555G mutation associated with
cardiomyopathy and hearing loss following high-dose chemotherapy
and repeated aminoglycoside exposure. J Pediatr. 164:413–415. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Salomão KB, Ayo CM and Della-Rosa VA:
Investigation of the A1555G mutation in mitochondrial DNA (MT-RNR1)
in groups of Brazilian individuals with nonsyndromic deafness and
normal-hearing. Indian J Hum Genet. 19:54–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bae JW, Kim DB, Choi JY, Park HJ, Lee JD,
Hur DG, Bae SH, Jung DJ, Lee SH, Kim UK and Lee KY: Molecular and
clinical characterization of the variable phenotype in Korean
families with hearing loss associated with the mitochondrial A1555G
mutation. PLoS One. 7:e424632012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guan MX, Fischel-Ghodsian N and Attardi G:
Nuclear background determines biochemical phenotype in the
deafness-associated mitochondrial 12S rRNA mutation. Hum Mol Genet.
10:573–580. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tekin D, Yan D, Bademci G, Feng Y, Guo S,
Foster J II, Blanton S, Tekin M and Liu X: A next-generation
sequencing gene panel (MiamiOtoGenes) for comprehensive analysis of
deafness genes. Hear Res. 333:179–184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rieder MJ, Taylor SL, Tobe VO and
Nickerson DA: Automating the identification of DNA variations using
quality-based fluorescence re-sequencing: Analysis of the human
mitochondrial genome. Nucleic Acids Res. 26:967–973. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Andrews RM, Kubacka I, Chinnery PF,
Lightowlers RN, Turnbull DM and Howell N: Reanalysis and revision
of the Cambridge reference sequence for human mitochondrial DNA.
Nat Genet. 23:1471999. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leclère JC, Le Gac MS, Le Maréchal C,
Ferec C and Marianowski R: GJB2 mutations: Genotypic and phenotypic
correlation in a cohort of 690 hearing-impaired patients, toward a
new mutation? Int J Pediatr Otorhinolaryngol. 102:80–85. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bibb MJ, Van Etten RA, Wright CT, Walberg
MW and Clayton DA: Sequence and gene organization of mouse
mitochondrial DNA. Cell. 26:167–180. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gadaleta G, Pepe G, De Candia G,
Quagliariello C, Sbisà E and Saccone C: The complete nucleotide
sequence of the Rattus norvegicus mitochondrial genome: Cryptic
signals revealed by comparative analysis between vertebrates. J Mol
Evol. 28:497–516. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Roe BA, Ma DP, Wilson RK and Wong JF: The
complete nucleotide sequence of the Xenopus laevis
mitochondrial genome. J Biol Chem. 260:9759–9774. 1985.PubMed/NCBI
|
|
20
|
Kong QP, Bandelt HJ, Sun C, Yao YG, Salas
A, Achilli A, Wang CY, Zhong L, Zhu CL, Wu SF, et al: Updating the
east asian mtDNA phylogeny: A prerequisite for the identification
of pathogenic mutations. Hum Mol Genet. 15:2076–2086. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sprinzl M and Vassilenko KS: Compilation
of tRNA sequences and sequences of tRNA genes. Nucleic Acids Res.
33:D139–D140. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Asano K, Suzuki T, Saito A, Wei FY,
Ikeuchi Y, Numata T, Tanaka R, Yamane Y, Yamamoto T, Goto T, et al:
Metabolic and chemical regulation of tRNA modification associated
with taurine deficiency and human disease. Nucleic Acids Res.
46:1565–1583. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sloan KE, Höbartner C and Bohnsack MT: How
RNA modification allows non-conventional decoding in mitochondria.
Cell Cycle. 16:145–146. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou M, Xue L, Chen Y, Li H, He Q, Wang B,
Meng F, Wang M and Guan MX: A hypertension-associated mitochondrial
DNA mutation introduces an m1G37 modification into tRNAMet,
altering its structure and function. J Biol Chem. 293:1425–1438.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qu J, Li R, Zhou X, Tong Y, Lu F, Qian Y,
Hu Y, Mo JQ, West CE and Guan MX: The novel 4435A>G mutation in
the mitochondrial tRNAMet may modulate the phenotypic expression of
the LHON-associated ND4 G11778A mutation. Invest Ophthalmol Vis
Sci. 47:475–483. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu Y, Liang LZ, Xiao HL, Yang YL, Yu X,
Zheng J, Fang F, Zheng BJ, Tang XW, Jin LJ and Guan MX: Hearing
loss may be associated with the novel mitochondrial tRNA(Asp)
A7551G mutation in a Chinese family. Zhonghua Er Bi Yan Hou Tou
Jing Wai Ke Za Zhi. 48:978–984. 2013.(In Chinese). PubMed/NCBI
|
|
27
|
Meng F, He Z, Tang X, Zheng J, Jin X, Zhu
Y, Ren X, Zhou M, Wang M, Gong S, et al: Contribution of the
tRNAIle 4317A→G mutation to the phenotypic manifestation of the
deafness-associated mitochondrial 12S rRNA 1555A→G mutation. J Biol
Chem. 293:3321–3334. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Böttger EC and Schacht J: The
mitochondrion: A perpetrator of acquired hearing loss. Hear Res.
303:12–19. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Riva C, Donadieu E, Magnan J and Lavieille
JP: Age-related hearing loss in CD/1 mice is associated to ROS
formation and HIF target proteins up-regulation in the cochlea. Exp
Gerontol. 42:327–336. 2007. View Article : Google Scholar : PubMed/NCBI
|