Introduction
Gastric cancer (GC) is the fourth most frequently
occurring cancer and the second most common cause of
cancer-associated mortality worldwide (1), with approximately 1,000,000 newly
diagnosed cases and >700,000 mortalities due to GC estimated to
occur annually worldwide. To date, several risk factors influencing
the formation and progression of GC have been identified, including
Helicobacter pylori infection, smoking, obesity, dietary
habits and chronic atrophic gastritis (2,3).
Despite significant progress in treatment development, the clinical
outcome of patients with advanced-stage GC remains poor, with a
5-year survival rate of only 30–50% (4). Tumor formation and development in GC
is multifactorial and significant genetic and epigenetic changes
have been demonstrated to contribute to GC (5). However, the molecular mechanism
associated with GC occurrence and development remains largely
unknown. Therefore, further insight into the mechanisms that
regulate GC progression may be advantageous in the discovery and
development of novel therapies for patients with this disease.
MicroRNAs (miRNAs/miRs) are a group of short
non-coding RNA molecules that are critical regulators of
oncogenesis and cancer progression (6). miRNAs silence the expression of their
target genes through direct binding to the 3′untranslated regions
(UTRs), resulting in translation inhibition and/or mRNA degradation
(7). Previous studies have
reported that miRNAs modulate the expression of >50% of human
protein-coding genes involved in the regulation of a wide range of
physiological and pathological processes, including cell
proliferation, cycle, apoptosis, differentiation, metabolism and
tumorigenesis (8). Several miRNAs
have been demonstrated to be aberrantly expressed in the majority
of human malignancies (9–11). Abnormally expressed miRNAs may
result in the progression or inhibition of normal cell growth
patterns, thus leading to cancer initiation and progression
(12). miRNAs may have roles in
tumor suppression or oncogenesis, depending on the functional roles
of their target genes (13,14).
Hence, investigation of the regulatory roles of miRNAs in human
cancers may provide promising therapeutic targets for antitumor
therapy.
The aberrant expression of miR-18b has been widely
reported to occur in several types of human cancer (15–17).
However, the expression pattern, biological role and specific
functional mechanism of miR-18b in GC remains to be fully
elucidated. A better understanding of miR-18b in GC may provide
novel therapeutic targets for the treatment of patients with
GC.
Materials and methods
Human tissue samples
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Guangxi Medical
University (Nanning, China). Written informed consent was obtained
from all participants. A total of 49 GC and matched adjacent normal
gastric tissue samples were collected from patients (28 males, 21
females; age range, 52–79 years) who underwent surgery resection at
The First Affiliated Hospital of Guangxi Medical University between
July 2014 and November 2016. No patients received chemotherapy,
radiotherapy or other treatments prior to surgery. Upon resection,
all tissues were immediately frozen in liquid nitrogen and
subsequently stored at −80°C.
Cell culture and transfection
GC cell lines (AGS, SGC-7901, BGC-823 and MGC-803)
and the human gastric epithelial immortalized GES-1 cell line were
purchased from American Type Culture Collection (Manassas, VA,
USA). All cell lines were cultured in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1%
penicillin-streptomycin solution at 37°C in a humidified incubator
with 5% CO2. DMEM, FBS and antibiotic solution were all
purchased from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA).
miR-18b inhibitor, miRNA inhibitor negative control
(NC inhibitor), KLF6-specific small interfering RNA (siRNA) and
negative control siRNA (NC siRNA) were synthesized by Shanghai
GenePharma Co., Ltd., (Shanghai, China). The miR-18b inhibitor
sequence was 5′-CUAACUGCACUAGAUGCACCUUA-3′ and the NC inhibitor
sequence was 5′-CAGUACUUUUGUGUAGUACAAA-3′.
The KLF6 siRNA sequence was
5′-GCAGGAAAGUUUACACCAATT-3′ and the NC siRNA sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. Cells were plated into 6-well plates
at a density of 8×105 cells/well. Following incubation
overnight, cells were transfected with miR-18b inhibitor (100
pmol), NC inhibitor (100 pmol), KLF6 siRNA (100 pmol) or NC siRNA
(100 pmol) using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
A total of 48 h following transfection, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) was
performed to determine miR-18b expression. Cell Counting kit-8
(CCK8) and cell invasion assays were performed at 24 and 48 h post
transfection. Western blot analysis was carried out at 72 h
following transfection.
RT-qPCR
Total tissue or cell RNA was isolated using
TRIzol® reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. To quantify miR-18b
expression, total RNA was converted into first-strand complementary
DNA (cDNA) using a TaqMan microRNA reverse transcription kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
temperature protocol for reverse transcription was as follows: 16°C
for 30 min, 42°C for 30 min and 85°C for 5 min. Subsequently, PCR
amplification was performed to detect miR-18b expression with the
TaqMan microRNA PCR kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The cycling conditions were as follows: 50°C for
2 min, 95°C for 10 min; 40 cycles of denaturation at 95°C for 15
sec; and annealing/extension at 60°C for 60 sec. To quantify KLF6
mRNA expression, cDNA was synthesized from total RNA using a
Moloney Murine Leukemia Virus reverse transcriptase kit (Promega
Corporation, Madison, WI, USA). The temperature protocol for
reverse transcription was as follows: 95°C for 2 min; 20 cycles of
94°C for 1 min, 55°C for 1 min and 72°C for 2 min; and 72°C for 5
min. cDNA was subsequently subjected to qPCR using SYBR®
Premix Ex TaqTM II (Takara Biotechnology Co., Ltd., Dalian, China).
The cycling conditions were as follows: 5 min at 95°C, followed by
40 cycles of 95°C for 30 sec and 65°C for 45 sec. U6 snRNA and
GAPDH were used as internal reference for miR-18b and KLF6 mRNA,
respectively. The primers were designed as follows: miR-18b,
5′-GGGTAAGGTGCATCTAGTGC-3′ (forward) and 5′-CAGTGCGTGTCGTGGAGT-3′
(reverse); U6, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ (forward) and
5′-CGCTTCACGAATTTGCGTGTCAT-3′ (reverse); KLF6,
5′-CGGACGCACACAGGAGAAAA-3′ (forward) and 5′-CGGTGTGCTTTCGGAAGTG-3′
(reverse); and GAPDH, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ (forward) and
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′ (reverse). Data were analyzed using
the 2−ΔΔCq method (18).
Cell Counting kit-8 (CCK8) assay
A CCK8 assay was performed to detect cell
proliferation. Cells were plated into 96-well plates at a density
of 3×103 cells/well. Following transfection, cells were
incubated at 37°C in a humidified incubator with 5% CO2
for 0, 24, 48 and 72 h. At each time point, the CCK8 assay was
conducted according to the manufacturer's protocol. Briefly, 10 µl
CCK8 solution (Dojindo Molecular Technologies, Inc., Kumamoto,
Japan) was added to each well and incubated at 37°C with 5%
CO2 for further 2 h. Optical density values were
measured at a wavelength of 450 nm using a microplate
spectrophotometer (Bio-Tek Instruments, Inc., Winooski, VT,
USA).
Cell invasion assay
Matrigel coated Transwell chambers (pore size, 8-µm;
BD Biosciences, Franklin Lakes, NJ, USA) were used to examine cell
invasive ability. Transfected cells were collected 48 h
post-transfection. A total of 1×105 cells in 200 µl
FBS-free DMEM medium were added to the upper chamber. The lower
chambers were filled with 500 µl DMEM supplemented with 10% FBS.
Following incubation for 24 h, cells remaining on the upper surface
were removed using cotton swabs. The invaded cells were fixed with
100% methanol at room temperature for 15 min, stained with 0.1%
crystal violet at room temperature for 15 min and washed with PBS.
The number of invasive cells was counted under an inverted light
microscope (Olympus Corporation, Tokyo, Japan) in five randomly
selected fields.
Bioinformatics analysis and luciferase
reporter assay
The putative target genes of miR-18b were predicted
using miRanda (August 2010 Release, Last Update: 2010-11-01;
www.microrna.org) and TargetScan (version 7.1;
www.targetscan.org). KLF6 was predicted as a
potential target of miR-18b. A luciferase reporter assay was
utilized to investigate if KLF6 is a direct target of miR-18b.
Luciferase plasmids pmirGLO-KLF6-3′-UTR wild type (Wt) and
pmirGLO-KLF6-3′-UTR mutant (Mut), were chemically synthesized by
Shanghai GenePharma Co., Ltd. Cells were seeded in 24-well plates
at an initial density of 1.5×105 cells/well. Following
an overnight incubation, cells were transfected with miR-18b
inhibitor or NC inhibitor, in addition to pmirGLO-KLF6-3′-UTR Wt or
pmirGLO-KLF6-3′-UTR Mut using Lipofectamine® 2000.
Following incubation at 37°C with 5% CO2 for 48 h, cells
were harvested and the luciferase activity was detected using the
Dual-Luciferase Reporter assay system (Promega Corporation)
according to the manufacturer's protocol. Firefly luciferase
activity was normalized Renilla luciferase activity.
Western blot analysis
The primary antibodies used in the present study
were mouse anti-human KLF6 primary antibody (1:1,000; cat. no.
sc-134374) and mouse anti-human GAPDH primary antibody (1:1,000;
cat. no. sc-32233), purchased from Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA. Total protein was extracted from tissues or cells
using a radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China). The concentration of
total protein was quantified with a bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology). Equal amounts of
protein (30 µg) were loaded into each lane and separated by 10%
SDS-PAGE, followed by transfer onto polyvinylidene difluoride
membranes. Membranes were blocked with 5% skimmed milk in
Tris-buffered saline containing 0.1% Tween-20 (TBST) at room
temperature for 1 h, followed by incubation with primary antibodies
at 4°C overnight. Membranes were subsequently washed with TBST
three times and incubated with goat anti-mouse horseradish
peroxidase-conjugated secondary IgG goat anti-mouse (1:5,000; cat.
no. sc-2005; Santa Cruz Biotechnology, Inc.) at room temperature
for 2 h. Protein bands were visualized using an enhanced
chemiluminescence plus reagent (GE Healthcare Life Sciences, Little
Chalfont, UK). Protein expression was quantified using Quantity One
software version 4.62 (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Statistical analysis
Data is expressed as the mean ± standard deviation
of at least 3 independent experiments and was analyzed using the
Student's t-test or one-way analysis of variance followed by the
Student-Newman-Keuls multiple comparisons test. The association
between miR-18b and clinicopathological characteristics of patients
with GC was analyzed using a univariate χ2 test.
Spearman's correlation analysis was performed to evaluate the
association between miR-18b and KLF6 mRNA levels in GC tissues.
SPSS software, version 13.0 (SPSS, Inc., Chicago, IL, USA) was used
for statistical analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
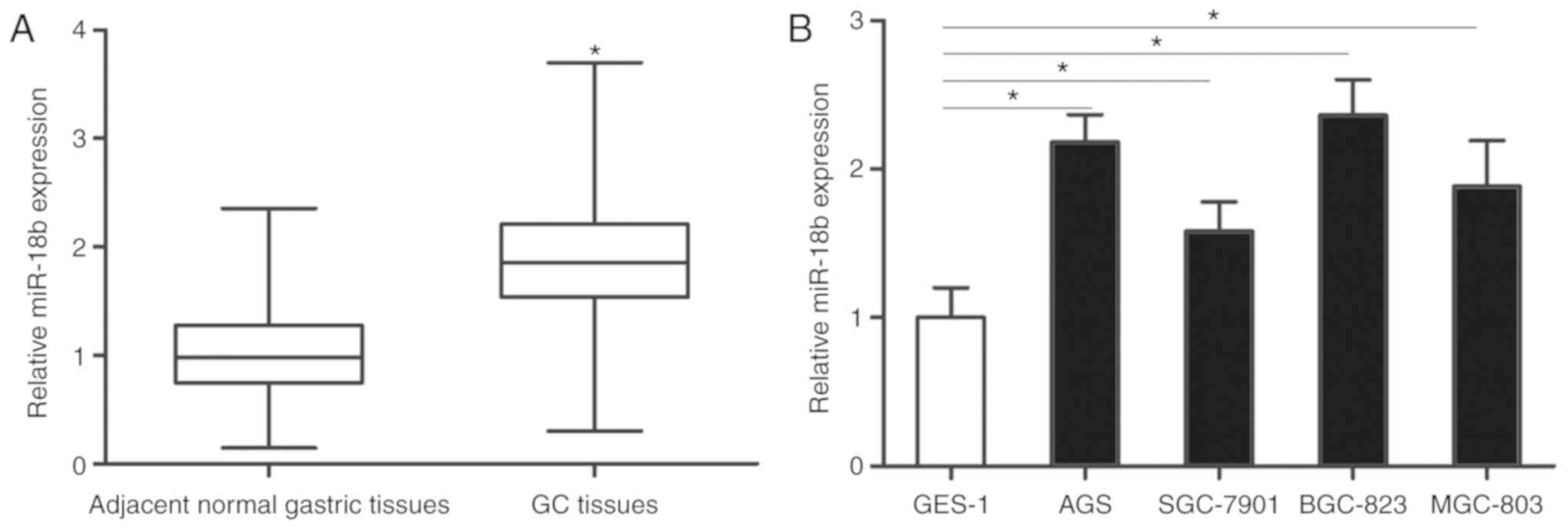
MiR-18b is upregulated in GC tissues
and cell lines
miR-18b expression was detected in 49 GC and matched
adjacent normal gastric tissue samples. RT-qPCR analysis revealed
that miR-18b expression was significantly upregulated in GC tissues
compared with the adjacent normal gastric tissues (P<0.05;
Fig. 1A). Additionally, the
expression levels of miR-18b in GC cell lines and the human gastric
epithelial immortalized cell line GES-1 were also determined using
RT-qPCR. Compared with GES-1, higher expression levels of miR-18b
were detected in all four GC cell lines (P<0.05; Fig. 1B). As AGS and BGC-823 cells
expressed relatively higher miR-18b expression amongst the four GC
cell lines, these two cell lines were selected for the subsequent
experiments. These results indicated that miR-18b is upregulated in
both GC tissues and cell lines.
Association between miR-18b and
clinicopathological factors of patients with GC
To explore the clinical value of miR-18b in GC,
patients were divided into miR-18b high-expression group (n=25) and
low-expression groups (n=24) based on the median expression of
miR-18b (1.86). As presented in Table
I, high miR-18b expression was associated with lymph node
metastasis (P=0.032), invasive depth (P=0.015) and Tumor Node
Metastasis (TNM) stage (P=0.015) in patients with GC. However, no
significant difference was observed between miR-18b expression and
other clinicopathological factors, including age, sex, tumor size
and differentiation (P>0.05). These results suggested that
miR-18b may be associated with the malignant progression of GC.
 | Table I.Association between miR-18b
expression and clinicopathological features of patients with
gastric cancer. |
Table I.
Association between miR-18b
expression and clinicopathological features of patients with
gastric cancer.
|
|
| miR-18b
expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | Number of
cases | High | Low | P-value |
|---|
| Age (years) |
|
|
| 0.282 |
|
<60 | 18 | 11 | 7 |
|
|
≥60 | 31 | 14 | 17 |
|
| Sex |
|
|
| 0.322 |
|
Male | 28 | 16 | 12 |
|
|
Female | 21 | 9 | 12 |
|
| Tumor size
(cm) |
|
|
| 0.458 |
|
<4 | 21 | 12 | 9 |
|
| ≥4 | 28 | 13 | 15 |
|
|
Differentiation |
|
|
| 0.477 |
| Well
and moderate | 24 | 11 | 13 |
|
| Poor
and signet | 25 | 14 | 11 |
|
| Lymph node
metastasis |
|
|
| 0.032a |
| No | 23 | 8 | 15 |
|
|
Yes | 26 | 17 | 9 |
|
| Invasive depth |
|
|
| 0.015b |
|
T1+T2 | 26 | 9 | 17 |
|
|
T3+T4 | 23 | 16 | 7 |
|
| TNM stage |
|
|
| 0.015c |
|
I–II | 20 | 6 | 14 |
|
|
III–IV | 29 | 19 | 10 |
|
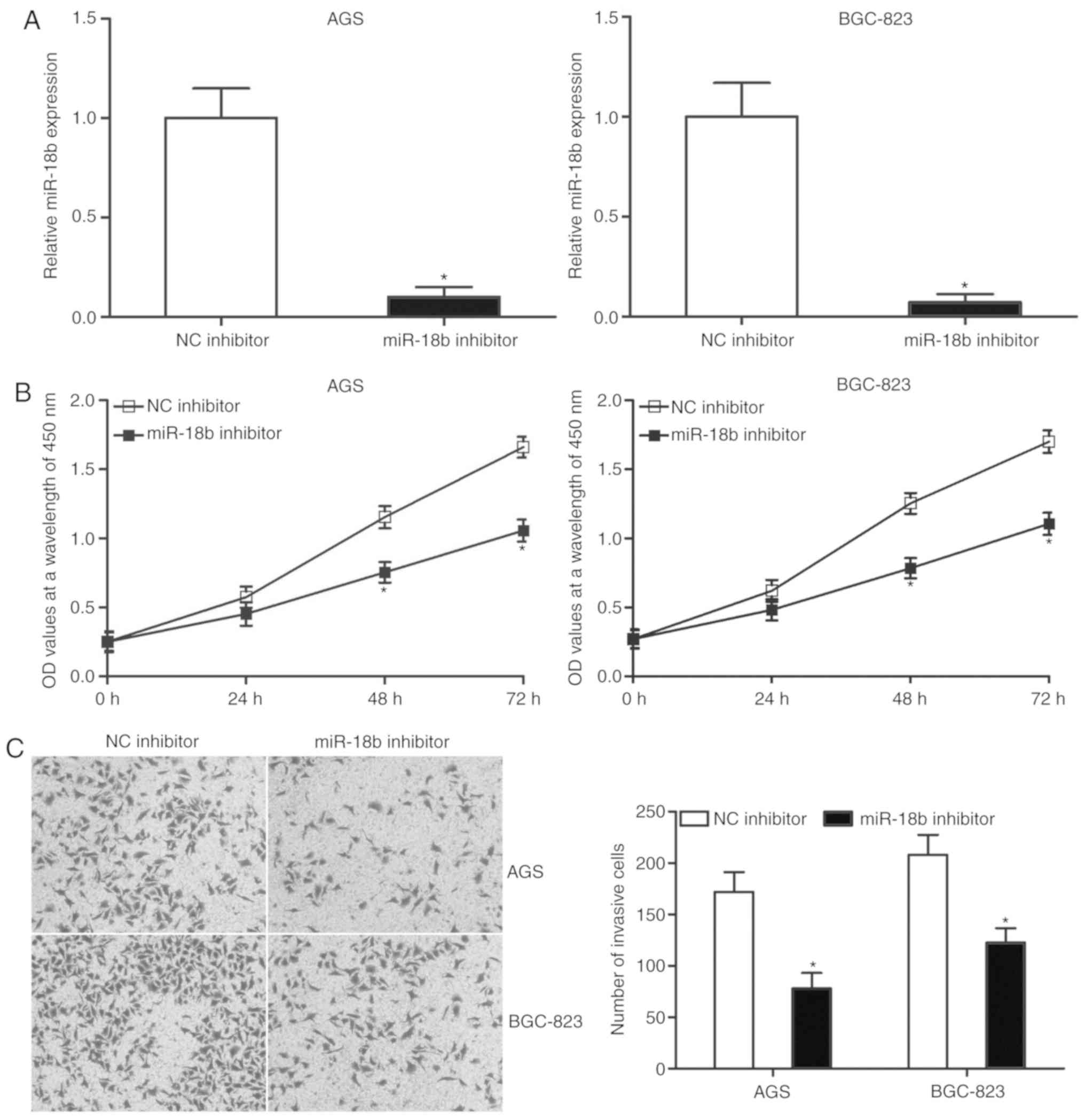
Downregulation of miR-18b inhibits
proliferation and invasion of AGS and BGC-823 cells
To investigate the role of miR-18b in GC, AGS and
BGC-823 cells were transfected with miR-18b inhibitor to decrease
its expression. Following transfection, RT-qPCR revealed that
miR-18b was significantly downregulated in cells transfected with
miR-18b inhibitor compared with the cells transfected with NC
inhibitor (Fig. 2A; P<0.05). A
CCK-8 assay was subsequently conducted to examine the effect of
miR-18b knockdown on GC cell proliferation. The downregulation of
miR-18b significantly suppressed the proliferation of AGS and
BGC-823 cells (Fig. 2B;
P<0.05). Additionally, a cell invasion assay was performed to
evaluate cell invasion ability in cells transfected with miR-18b
inhibitor or NC inhibitor. Transfection with miR-18b inhibitor
markedly attenuated the invasion capacities of AGS and BGC-823
cells compared with the NC inhibitor group (Fig. 2C; P<0.05). Taken together, this
indicated that miR-18b may have an oncogenic role in GC
progression.
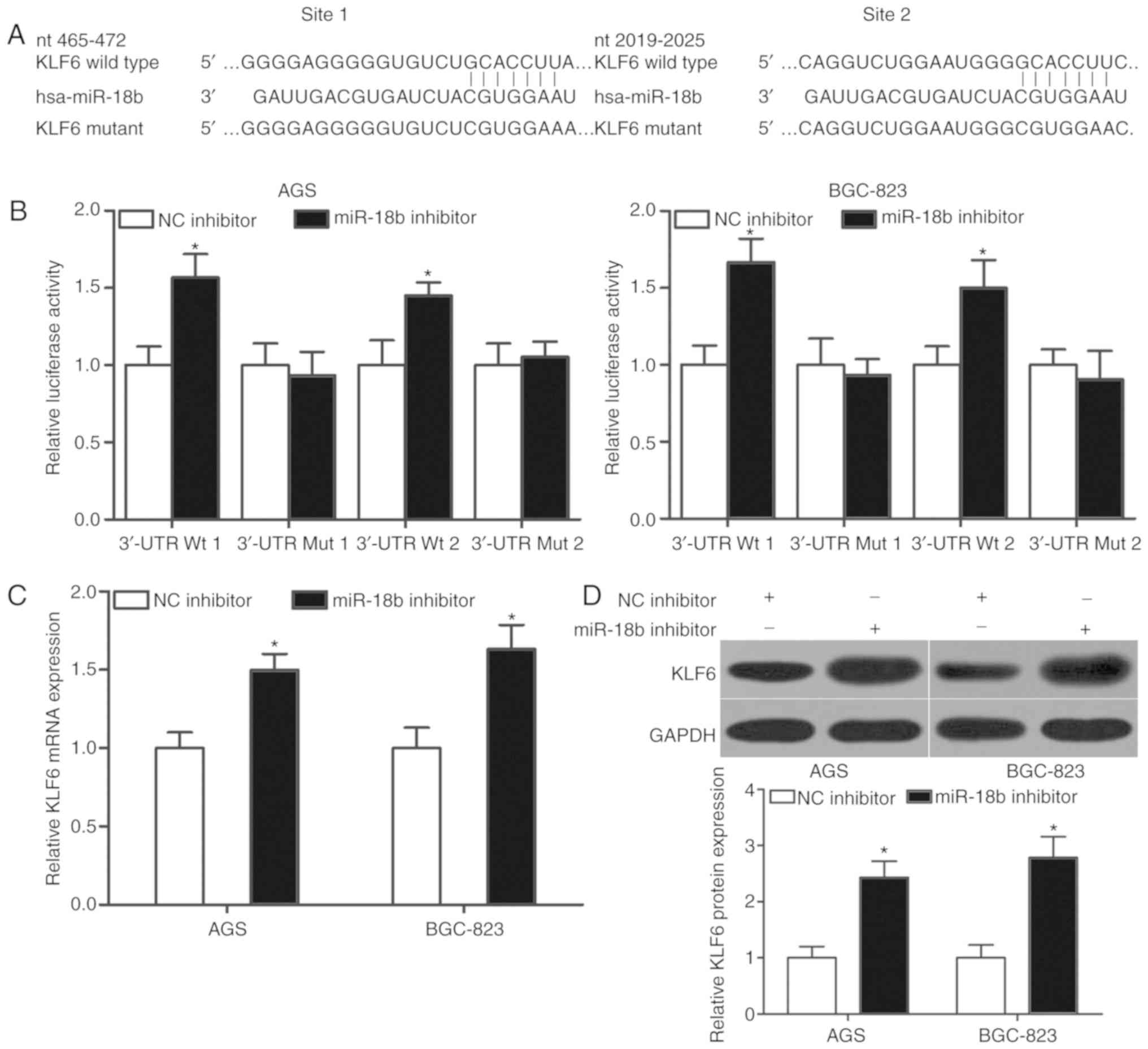
KLF6 is a direct target of miR-18b in
GC
To elucidate the mechanism underlying the oncogenic
role of miR-18b in GC, bioinformatics analysis was used to predict
the potential targets of miR-18b. KLF6, which has previously been
reported to affect GC initiation and progression (19–21),
was predicted as a putative target of miR-18b. As presented in
Fig. 3A, the 3′-UTR of KLF6
contains two miR-18b binding sites (1 and 2). A luciferase reporter
assay was performed to investigate if miR-18b interacted with the
3′-UTR of KLF6. miR-18b downregulation increased the luciferase
activity of pmirGLO-KLF6-3′-UTR Wt (1 and 2) in AGS and BGC-823
cells (Fig. 3B; P<0.05).
However, no significant changes in luciferase activity were
detected in cells transfected with pmirGLO-KLF6-3′-UTR Mut (1 and
2) in the presence or absence of miR-18b inhibitor. Furthermore,
RT-qPCR and western blot analysis were performed to investigate if
miR-18b regulated KLF6 expression in GC cell lines. Transfection of
miR-18b inhibitor increased the expression of KLF6 expression at
both mRNA (P<0.05; Fig. 3C) and
protein (P<0.05; Fig. 3D)
levels in AGS and BGC-823 cells. Collectively, this data indicated
that KLF6 may be a direct target of miR-18b in GC.
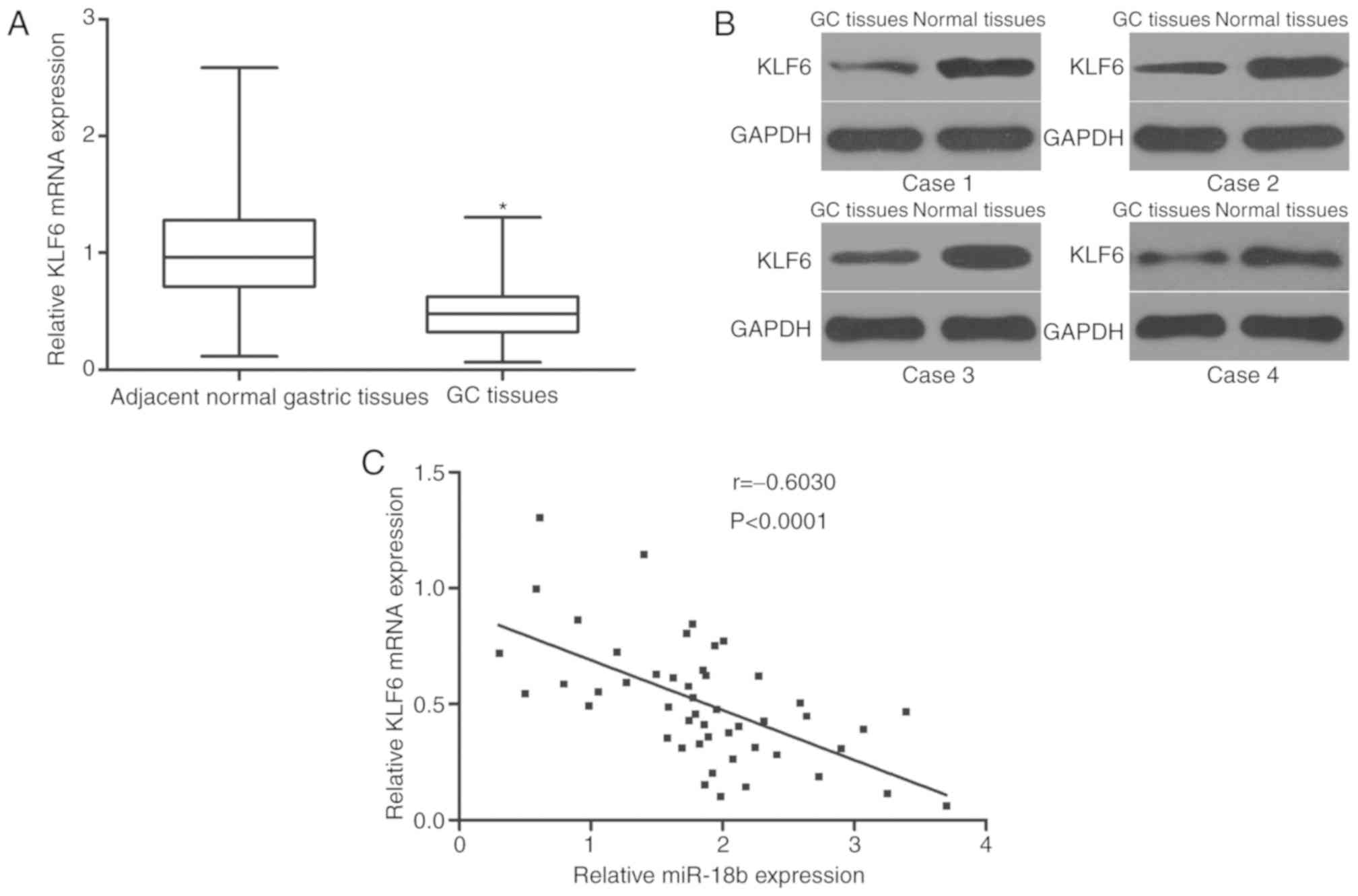
KLF6 is downregulated in GC tissues
and inversely correlates with miR-18b expression
To further explore the association between miR-18b
and KLF6 in GC, the expression of KLF6 in 49 pairs of GC and
matched adjacent normal gastric tissues was detected. RT-qPCR and
western blot analysis revealed that KLF6 expression was
downregulated in GC tissues compared with adjacent normal gastric
tissues at the mRNA (Fig. 4A;
P<0.05) and protein (Fig. 4B)
level. Additionally, a negative association between miR-18b and
KLF6 mRNA expression in GC tissues was confirmed by Spearman's
correlation analysis (Fig. 4C;
r=−0.6030; P<0.0001). These results suggested that the
downregulation of KLF6 in GC may be attributed at least in part to
the upregulation of miR-18b.
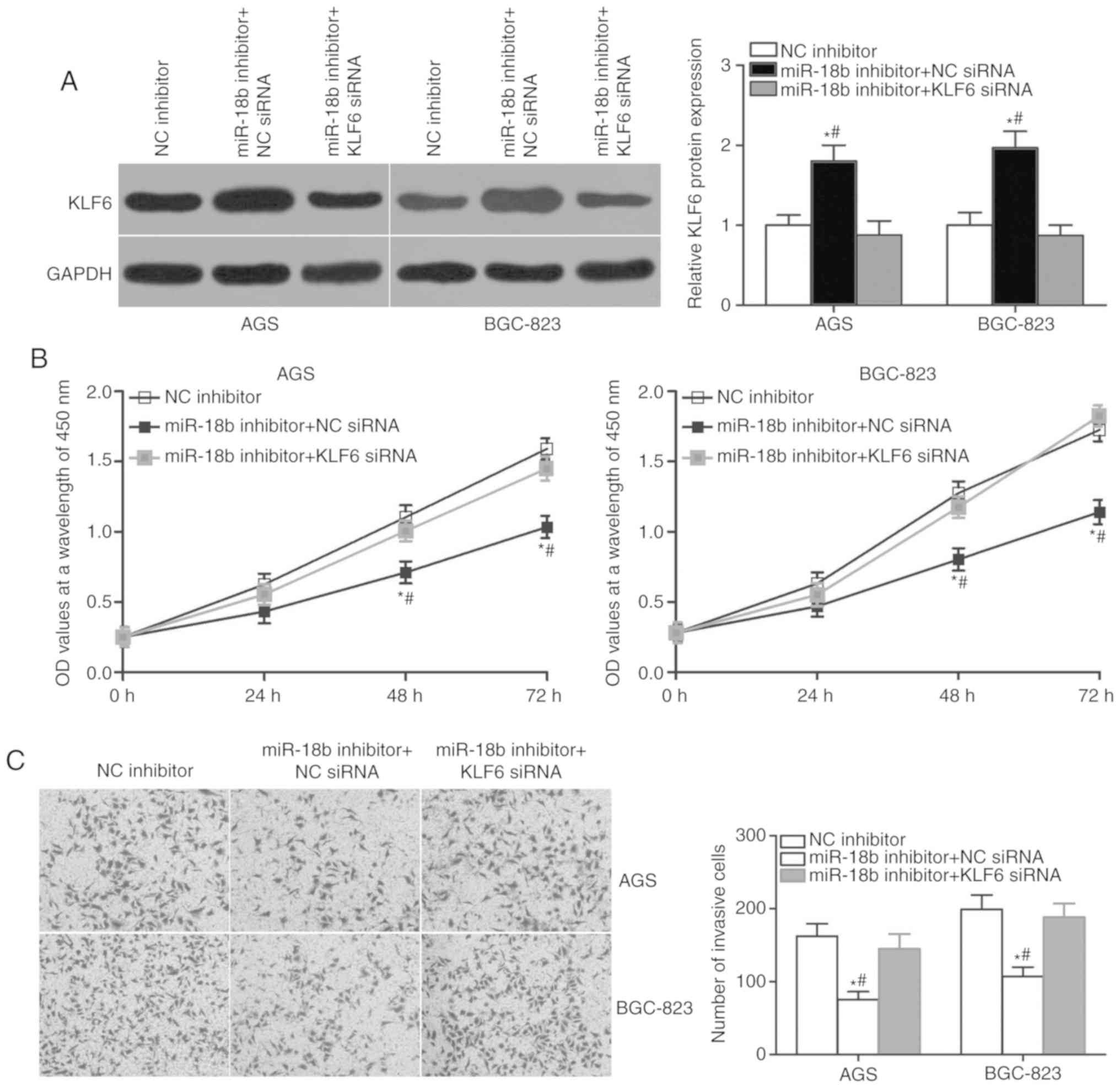
KLF6 knockdown partially rescues the
effects of miR-18b inhibitor on proliferation and invasion of AGS
and BGC-823 cells
Rescue experiments were performed to further confirm
that the effect of miR-18b knockdown on GC cell proliferation and
invasion is mediated by the regulation of KLF6 expression. AGS and
BGC-823 cells were transfected with miR-18b inhibitor in
combination with KLF6 siRNA or NC siRNA. Western blot analysis
indicated that co-transfection of KLF6 siRNA partially abrogated
the miR-18b inhibitor-mediated upregulation of KLF6 (Fig. 5A; P<0.05). Additionally,
functional experiments demonstrated that KLF6 restoration rescued
the inhibition of cell proliferation (Fig. 5B; P<0.05) and invasion (Fig. 5C; P<0.05) caused by miR-18b
knockdown in AGS and BGC-823 cells. Collectively, these results
suggested that the oncogenic roles of miR-18b in GC are achieved,
at least in part, by regulation of KLF6 expression.
Discussion
An increasing number of studies have demonstrated
that miRNAs contribute to gastric cancer development and
progression (22–24). Therefore, investigation of the
miRNAs involved in the development of GC may provide an effective
therapeutic target for patients with this malignancy. In the
present study, miR-18b was significantly upregulated in GC tissues
and cell lines. High miR-18b expression was associated with lymph
node metastasis, invasive depth and TNM stage in patients with GC.
Inhibition of miR-18b prohibited the proliferation and invasion of
GC cells. Additionally, KLF6 was identified as a direct target of
miR-18b in GC. KLF6 was downregulated in GC and negatively
correlated with miR-18b expression level. Furthermore, KLF6
restoration partially rescued the effects of miR-18b inhibition on
the proliferation and invasion of GC cells. Therefore, targeting of
miR-18b may be developed as a potential therapeutic strategy in the
treatment of patients with GC.
miR-18b expression is dysregulated in several types
of human cancer. miR-18b is downregulated in melanoma tissues and
cell lines, and decreased levels are also significantly associated
with tumor thickness and stage (15). Additionally, miR-18b has been
reported to be upregulated in colorectal cancer. High expression of
miR-18b is significantly associated with lymph node and distant
metastasis of patients with colorectal cancer (16). Expression of miR-18b is also higher
in poorly differentiated hepatocellular carcinoma (HCC), compared
with that in well-differentiated HCC. Following surgery resection,
patients with HCC and high miR-18b expression have a shorter
relapse-free period than those patients with low expression
(17). In mantle cell lymphoma,
miR-18b is overexpressed and associated with a poor survival rate
(25). Elevated miR-18b expression
is also reported in nasopharyngeal carcinoma (26) and breast cancer (27). These findings indicate that the
expression pattern of miR-18b possesses tissue specificity and may
serve as a useful prognostic marker in human cancers.
Numerous studies have reported that miR-18b
contributes to the malignant phenotype of cancers. Upregulation of
miR-18b has been demonstrated to inhibit melanoma cell
proliferation, migration, invasion, glycolysis and
epithelial-to-mesenchymal transition in vitro, increase
apoptosis in vitro and reduce tumor growth in vivo
(15,28). miR-18b has been validated as an
oncogene in colorectal cancer via participation in the regulation
of cell cycle, proliferation and migration (16). Murakami et al (17) reported that the upregulation of
miR-18b promotes cell proliferation and inhibits cell adhesion
capacity in HCC. Fonseca-Sanchéz et al (27) demonstrated that the downregulation
of miR-18b attenuates breast cancer cell migration in vitro.
These results suggest that biological roles of miR-18b also have
tissue specificity and further indicate the potential of miR-18b as
a therapeutic target for the treatment of certain cancer types.
Several targets of miR-18b have been identified,
including hypoxia inducible factor 1 a (15) and MDM2 (28) in melanoma, cyclin dependent kinase
inhibitor 2B (16) in colorectal
cancer and trinucleotide repeat containing 6B (17) in HCC. In the present study, KLF6
was validated as a novel target of miR-18b in GC. KLF6, a zinc
finger transcription factor, is frequently downregulated in several
types of human cancer, including lung (29), ovarian (30), prostate (31), glioma (32) and colorectal cancer (33). Previous studies have demonstrated
that KLF6 contributes to the regulation of various biological
processes, including cell proliferation, apoptosis,
differentiation, invasion and metastasis (34,35).
In GC, the expression level of KLF6 is decreased in tumor tissues
compared with normal gastric mucosa (18). Low KLF6 expression is strongly
associated with histological differentiation, TNM stage, lymph node
metastasis and distant metastasis (19). KLF6 overexpression reduces cell
proliferation, colony formation, metastasis and increases apoptosis
in GC (20,21). Considering the important roles of
KLF6 in GC, targeting KLF6 may provide an opportunity to inhibit
tumor formation and development in GC.
In conclusion, miR-18b was upregulated in GC tissues
and cell lines. Its high expression was significantly associated
with lymph node metastasis, invasive depth and TNM stage in
patients with GC. miR-18b may have an oncogenic role in GC partly
through direct targeting of KLF6, suggesting that the miR-18b/KLF6
axis may be a potential therapeutic target for treating patients
with GC.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81360370), the Natural
Science Foundation of Guangxi Zhuang Autonomous Region (grant no.
2017AB45153), the Education Department Foundation for Innovation
Team of Guangxi Zhuang Autonomous Region and the self-financing
project of the Guangxi Zhuang Autonomous Region Health and Family
Planning Commission (grant no. Z 20170908).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
DL and JC designed the present study. DL, SH, JX, XS
and PY performed functional experiments. All authors have read and
approved the final draft.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of The First Affiliated Hospital of Guangxi
Medical University (Nanning, China), and was performed in
accordance with the Declaration of Helsinki and the guidelines of
the Ethics Committee of The First Affiliated Hospital of Guangxi
Medical University.
Consent for publication
Written informed consent was obtained from all
patients for the use of their clinical tissues.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheng XJ, Lin JC and Tu SP: Etiology and
prevention of gastric cancer. Gastrointest Tumors. 3:25–36. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Compare D, Rocco A and Nardone G: Risk
factors in gastric cancer. Eur Rev Med Pharmacol Sci. 14:302–308.
2010.PubMed/NCBI
|
|
4
|
Newton AD, Datta J, Loaiza-Bonilla A,
Karakousis GC and Roses RE: Neoadjuvant therapy for gastric cancer:
Current evidence and future directions. J Gastrointest Oncol.
6:534–543. 2015.PubMed/NCBI
|
|
5
|
Otani K, Li X, Arakawa T, Chan FK and Yu
J: Epigenetic-mediated tumor suppressor genes as diagnostic or
prognostic biomarkers in gastric cancer. Expert Rev Mol Diagn.
13:445–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Monroig Pdel C, Chen L, Zhang S and Calin
GA: Small molecule compounds targeting miRNAs for cancer therapy.
Adv Drug Deliv Rev. 81:104–116. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding X, Liu J, Liu T, Ma Z, Wen D and Zhu
J: miR-148b inhibits glycolysis in gastric cancer through targeting
SLC2A1. Cancer Med. 6:1301–1310. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bai X, Meng L, Sun H, Li Z, Zhang X and
Hua S: MicroRNA-196b inhibits cell growth and metastasis of lung
cancer cells by targeting Runx2. Cell Physiol Biochem. 43:757–767.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Karatas OF, Wang J, Shao L, Ozen M, Zhang
Y, Creighton CJ and Ittmann M: miR-33a is a tumor suppressor
microRNA that is decreased in prostate cancer. Oncotarget.
8:60243–60256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pillai RS: MicroRNA function: Multiple
mechanisms for a tiny RNA? RNA. 11:1753–1761. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Long MJ, Wu FX, Li P, Liu M, Li X and Tang
H: MicroRNA-10a targets CHL1 and promotes cell growth, migration
and invasion in human cervical cancer cells. Cancer Lett.
324:186–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Zhang H, Dong Y, Fan Y, Li Y, Zhao
C, Wang C, Liu J, Li X, Dong M, et al: MiR-146b-5p functions as a
suppressor miRNA and prognosis predictor in non-small cell lung
cancer. J Cancer. 8:1704–1716. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen Y, Zhang Z, Luo C, Chen Z and Zhou J:
MicroRNA-18b inhibits the growth of malignant melanoma via
inhibition of HIF-1α-mediated glycolysis. Oncol Rep. 36:471–479.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Y, Chen M, Liu J, Li L, Yang X, Zhao J,
Wu M and Ye M: Upregulation of microRNA-18b contributes to the
development of colorectal cancer through inhibiting CDKN2B. Mol
Cell Biol. 37(pii): e00391–17. 2017.PubMed/NCBI
|
|
17
|
Murakami Y, Tamori A, Itami S, Tanahashi
T, Toyoda H, Tanaka M, Wu W, Brojigin N, Kaneoka Y, Maeda A, et al:
The expression level of miR-18b in hepatocellular carcinoma is
associated with the grade of malignancy and prognosis. BMC Cancer.
13:992013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Q, Tan XP, Yuan YS, Hu CM, He CH,
Wang WZ, Li JC, Zhao Q and Liu NZ: Decreased expression of KLF6 and
its significance in gastric carcinoma. Med Oncol. 27:1295–1302.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sangodkar J, Shi J, DiFeo A, Schwartz R,
Bromberg R, Choudhri A, McClinch K, Hatami R, Scheer E, Kremer-Tal
S, et al: Functional role of the KLF6 tumour suppressor gene in
gastric cancer. Eur J Cancer. 45:666–676. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang X, Nie Y, Du Y, Cao J, Shen B and Li
Y: MicroRNA-181a promotes gastric cancer by negatively regulating
tumor suppressor KLF6. Tumour Biol. 33:1589–1597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shrestha S, Hsu SD, Huang WY, Huang HY,
Chen W, Weng SL and Huang HD: A systematic review of microRNA
expression profiling studies in human gastric cancer. Cancer Med.
3:878–888. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu X, Lv M, Wang H and Guan W:
Identification of circulating microRNAs as novel potential
biomarkers for gastric cancer detection: A systematic review and
meta-analysis. Dig Dis Sci. 59:911–919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rao M, Zhu Y, Zhou Y, Cong X and Feng L:
MicroRNA-122 inhibits proliferation and invasion in gastric cancer
by targeting CREB1. Am J Cancer Res. 7:323–333. 2017.PubMed/NCBI
|
|
25
|
Husby S, Ralfkiaer U, Garde C, Zandi R, Ek
S, Kolstad A, Jerkeman M, Laurell A, Räty R, Pedersen LB, et al:
miR-18b overexpression identifies mantle cell lymphoma patients
with poor outcome and improves the MIPI-B prognosticator. Blood.
125:2669–2677. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu X, Zhen Y, Yang H, Wang H, Zhou Y, Wang
E, Marincola FM, Mai C, Chen Y, Wei H, et al: Loss of connective
tissue growth factor as an unfavorable prognosis factor activates
miR-18b by PI3K/AKT/C-Jun and C-Myc and promotes cell growth in
nasopharyngeal carcinoma. Cell Death Dis. 4:e6342013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fonseca-Sanchéz MA, Pérez-Plasencia C,
Fernandez-Retana J, Arechaga-Ocampo E, Marchat LA, Rodríguez-Cuevas
S, Bautista-Piña V, Arellano-Anaya ZE, Flores-Pérez A, Diaz-Chávez
J and López-Camarillo C: microRNA-18b is upregulated in breast
cancer and modulates genes involved in cell migration. Oncol Rep.
30:2399–2410. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dar AA, Majid S, Rittsteuer C, de Semir D,
Bezrookove V, Tong S, Nosrati M, Sagebiel R, Miller JR III and
Kashani-Sabet M: The role of miR-18b in MDM2-p53 pathway signaling
and melanoma progression. J Natl Cancer Inst. 105:433–442. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ito G, Uchiyama M, Kondo M, Mori S, Usami
N, Maeda O, Kawabe T, Hasegawa Y, Shimokata K and Sekido Y:
Kruppel-like factor 6 is frequently down-regulated and induces
apoptosis in non-small cell lung cancer cells. Cancer Res.
64:3838–3843. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
DiFeo A, Narla G and Martignetti JA:
Emerging roles of Kruppel-like factor 6 and Kruppel-like factor 6
splice variant 1 in ovarian cancer progression and treatment. Mt
Sinai J Med. 76:557–566. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang X, Li X and Guo B: KLF6 induces
apoptosis in prostate cancer cells through up-regulation of ATF3. J
Biol Chem. 283:29795–29801. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kimmelman AC, Qiao RF, Narla G, Banno A,
Lau N, Bos PD, Nuñez Rodriguez N, Liang BC, Guha A, Martignetti JA,
et al: Suppression of glioblastoma tumorigenicity by the
Kruppel-like transcription factor KLF6. Oncogene. 23:5077–5083.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cho YG, Choi BJ, Song JW, Kim SY, Nam SW,
Lee SH, Yoo NJ, Lee JY and Park WS: Aberrant expression of
krUppel-like factor 6 protein in colorectal cancers. World J
Gastroenterol. 12:2250–2253. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gao Y, Li H, Ma X, Fan Y, Ni D, Zhang Y,
Huang Q, Liu K, Li X, Wang L, et al: KLF6 suppresses metastasis of
clear cell renal cell carcinoma via transcriptional repression of
E2F1. Cancer Res. 77:330–342. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gehrau RC, D'Astolfo DS, Andreoli V, Bocco
JL and Koritschoner NP: Differential expression of the klf6 tumor
suppressor gene upon cell damaging treatments in cancer cells.
Mutat Res. 707:15–23. 2011. View Article : Google Scholar : PubMed/NCBI
|