Introduction
The hippocampus is one of the most intensely studied
brain regions given its function in learning and memory, providing
that it is a site of neurogenesis and that it has a unique cellular
organization and development (1).
In the mouse hippocampus, a small number (15%) of granule cells are
generated during embryogenesis, while the others are produced
within 2 weeks after birth (2–4).
Although we demonstrated in previous studies that the structural
lamination of the granule cell layer is accomplished by 2 weeks
after birth (4,5), maturation was observed at 3 weeks
after birth (6).
Cyclooxygenases (COXs) are enzymes that catalyze the
production of prostaglandins from the arachidonic acid.
Specifically, two COX isoforms similar in their amino acid sequence
exist, namely COX-1 and COX-2. While COX-1 is constitutively
expressed in cells, although its expression levels vary in
different cell types, COX-2 expression is induced by mitogens and
it is nearly undetectable in most tissues under normal conditions
(7,8). In fact, the arachidonic acid is
converted in the cell membrane into prostaglandin H2
(PGH2) by COX-2, which is a rate-limiting enzyme in
inflammatory processes. Subsequently, it is further metabolized
into prostanoids, including PGE2, PGI2, and
thromboxane A2 (9),
most of which are involved in the inflammatory response; PGE2 is
associated with excitatory synaptic transmission by the release of
glutamate instead (10).
For example, brain damage induced by transient
forebrain ischemia activates the inflammatory responses by
upregulating COX-2 expression and the subsequent increases in
prostaglandin synthesis (11).
Furthermore, pharmacological or genetic blockade of COX-2 confers
neuroprotection against ischemic damage (12). Nevertheless, COX-2 is
constitutively expressed in certain tissues, including the brain,
thymus, gut, and kidney (13).
While constitutive COX-2 is believed to play a major role in
homeostatic function, the constitutive expression of neuronal COX-2
is regulated by synaptic activity (14–16).
We demonstrated in a previous study that COX-2 immunoreactivity is
mainly found in both the granule cells of the dentate gyrus and the
pyramidal cells of the hippocampal CA3 region and that COX-2
immunoreactivity is decreased in the hippocampus with age (16). In addition, COX-2 deficiency
achieved by genetic and pharmacological inhibition decreased cell
proliferation and neuroblast differentiation in the dentate gyrus
(17–19). However, although COX-2 mRNA is
expressed in the brain, including in the hippocampus (20–22),
little is known about COX-2 expression during the structural
lamination of the hippocampus and its role in synaptic plasticity.
Therefore, in the present study, we investigated the postnatal
changes in both COX-2 immunoreactivity and protein levels in the
mice hippocampus to observe the localization of constitutive COX-2
during structural lamination. Furthermore, we examined the role of
COX-2 in the synaptic plasticity of the hippocampal dentate gyrus
using wild-type (WT) and COX-2 KO mice.
Experimental procedures
Experimental animals
Young male C57BL/6J mice at postnatal days (P) 1, 7,
14, 28, and 56 (n=10 for each group) were used in the
current study. The day of birth was considered day 0. Litters were
culled to a maximum of 7 pups at the day of birth, as described in
previous studies (4–6). From any one litter, a maximum of two
animals were taken for each age group, ensuring that animals of a
given age originated from at least four different litters. To
elucidate the role of COX-2 in the hippocampus, COX-2 KO and WT
mice (8-week-old) were purchased from Taconic (Hudson, NY, USA).
Animal handling and care conformed to the guidelines established by
the current international laws and policies (NIH Guide for the Care
and Use of Laboratory Animals, NIH Publication No. 85-23, 1985,
revised 2011) and were approved by both the Institutional Animal
Care and Use Committee (IACUC) of Seoul National University
(approval no.: SNU-120210-1 and SNU-140313-1) and Kangwon National
University (approval no.: KW-161226-1). All of the experiments were
conducted with an effort to minimize the number of animals used and
the suffering caused by the procedures employed in the present
study.
Tissue processing
Animals from each age group as well as COX-2 KO and
WT mice of 12 weeks of age were euthanized with a urethane
injection (1.5 g/kg body weight, intraperitoneally, Sigma; Merck
KGaA, Darmstadt, Germany). While the brains from each age group
were fixed for 24 h in 10% neutral buffered formalin at 25°C, COX-2
KO and WT mice were perfused transcardially with 0.1 M PBS (pH 7.4)
followed by 4% paraformaldehyde in a 0.1 M phosphate-buffer (pH
7.4), as described previously (4–6).
Subsequently, their brains were dissected and post-fixed for 12 h
in the same fixative; subsequently, they were dehydrated with
graded concentrations of alcohol to be embedded in paraffin.
Finally, paraffin-embedded tissues were sectioned on a microtome
(Leica Microsystems GmbH, Wetzlar, Germany) into 3-µm coronal
sections and mounted onto silane-coated slides.
Immunohistochemistry for COX-2, Arc,
GluN1, GluN2A/2B, and pCREB
All sections were processed under the same
conditions to ensure that the immunohistochemical data would be
comparable among the groups, as described in previous studies
(4–6). Specifically, they were hydrated and
treated with 0.3% H2O2 in phosphate-buffered
saline (PBS) for 30 min. Thereafter, they were placed in 100-ml
jars filled with citrate buffer (pH 6.0) and heated in a
2100-retriever (Prestige Medical, Lancashire, UK) for antigen
retrieval, after which slides were allowed to cool at room
temperature and were then washed in PBS. Subsequently, all sections
were incubated in 5% normal goat serum in PBS at 25°C for 30 min
and in diluted rabbit anti-COX-2 (1:200, Cayman Chemical Company,
Ann Arbor, MI, USA), mouse anti-Arc (activity-regulated
cytoskeletal, 1:500; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA), rabbit anti-GluN1 (N-methyl-d-aspartate receptor 1,
1:250; EMD Millipore, Billerica, MA, USA), rabbit anti-GluN2A/2B
(1:100; EMD Millipore), and rabbit anti-pCREB (phosphorylated cAMP
response element-binding protein, 1:400; Cell Signaling Technology,
Inc., Danvers, MA, USA) antibodies at 4°C for 72 h. Successively,
the sections were treated with biotinylated goat anti-rabbit IgG
antibody and a streptavidin-peroxidase complex (1:200; Vector
Laboratories, Inc., Burlingame, CA, USA) for 2 h at 25°C. All
sections were visualized through the reaction with
3,3′-diaminobenzidine tetrachloride (Sigma; Merck KGaA) in a 0.1 M
Tris-HCl buffer (pH 7.2) and mounted on gelatin-coated slides.
Sections were dehydrated and mounted in Canada balsam (Kanto,
Tokyo, Japan).
Western blot analysis
To quantify the changes in COX-2 levels within the
hippocampus, animals were euthanized at P1, 7, 14, 28, and 56
(n=5 per each group) and their brains were removed.
Furthermore, tissues were dissected and processed for use in the
western blot analysis, as described in a previous study (16). In brief, the hippocampus was
dissected using a surgical blade and homogenized in 50 mM PBS (pH
7.4) containing 0.1 mM ethylene glycol-bis
(2-aminoethylether)-N,N,N′,N′-tetraacetic acid (pH 8.0), 0.2%
Nonidet P-40, 10 mM ethylenediaminetetraacetic acid (pH 8.0), 15 mM
sodium pyrophosphate, 100 mM β-glycerophosphate, 50 mM NaF, 150 mM
NaCl, 2 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl
fluoride, and 1 mM dithiothreitol (DTT). After centrifugation, the
protein levels in the supernatant were determined using a Micro BCA
Protein assay kit according to the manufacturer's instructions
(Pierce Chemical, Rockford, IL, USA). Aliquots containing 20 µg of
total protein were denatured by boiling in the loading buffer
containing 150 mM Tris (pH 6.8), 3 mM DTT, 6% sodium dodecyl
sulfate, 0.3% bromophenol blue, and 30% glycerol and were then
loaded onto a polyacrylamide gel. Successively to electrophoresis,
proteins were transferred to nitrocellulose membranes (Pall Crop,
East Hills, NY, USA), which were then blocked in 5% non-fat dry
milk in PBS/0.1% Tween 20 for 45 min, prior to incubation with a
peroxidase-conjugated rabbit anti-COX-2 antibody (diluted 1:1,000).
Detection was performed using the peroxidase-conjugated anti-rabbit
IgG antibody (1:200; Vector Laboratories, Inc.) and an enhanced
luminol-based chemiluminescent kit (Pierce Chemical). Blots were
scanned and densitometry was conducted using the Scion Image
software (Scion Corp., Frederick, MD, USA). Finally, they were
stripped and reprobed with an antibody against β-actin as an
internal loading control. Data were normalized to the β-actin level
in each lane.
Data analysis
The analyses for COX-2 in both the hippocampal
CA1/CA3 regions and the dentate gyrus and for GluN1 and GluN2A/2B
in the dentate gyrus were performed using an image analysis system
and the ImageJ software v. 1.50 (National Institutes of Health,
Bethesda, MD, USA). Furthermore, data analysis was conducted under
the same conditions by two observers for each experiment to ensure
objectivity in the blind conditions, as described in a previous
study (16). Digital images of the
midpoint of the dentate gyrus were captured with a BX51 light
microscope (Olympus, Tokyo, Japan) equipped with a digital camera
(DP72, Olympus) connected to a computer monitor. Images were
calibrated into an array of 512×512 pixels corresponding to a
tissue area of 1,200×900 µm (×100 primary magnification). The
resolution of each pixel was 256 gray levels, while the intensity
of COX-2, GluN1, and GluN2A/2B pCREB immunoreactivity was evaluated
by relative optical density (ROD), which was obtained after the
transformation of the mean gray level using the following formula:
ROD=log (256/mean gray level). After determining the background
staining ROD in unlabeled portions of the sections using the
Photoshop CS6 software (Adobe Systems, Inc., San Jose, CA, USA),
such a value was subtracted to correct for non-specific staining,
using the ImageJ v. 1.50 software (National Institutes of Health).
Data are expressed as a percentage of either P1 or the WT group
(which was set at 100%).
Additionally, Arc and pCREB immunoreactive nuclei in
the whole dentate gyrus were counted with an analysis system
equipped with a computer-based CCD camera (OPTIMAS software version
6.5; CyberMetrics® Corporation, Phoenix, AZ, USA;
magnification, ×100), as described in a previous study (11). Thereafter, the image was converted
into a gray-scale image and Arc and pCREB immunoreactive nuclei
were automatically selected according to the intensity of Arc and
pCREB immunohistochemical staining, respectively.
Finally, tissue sections located at 90 µm from each
other were selected from an area between 1.82 and 2.30 mm posterior
to the bregma, as defined by a mouse atlas (23). Cell counts and ROD were averaged
using 5 sections from each WT and COX-2 KO mice.
Statistical analysis
The data were expressed as the mean of the
experiments performed for each experimental investigation. To
determine both the changes in COX-2 expression during postnatal
development and the effects of COX-2 depletion on Arc, pCREB,
GluN1, and GluN2A/2B, the mean differences among the groups were
statistically analyzed through one-way analyses of variance
followed by either the Bonferroni's post-hoc test or an unpaired
student's t-test using the GraphPad Prism 5.01 software (GraphPad
Software, Inc., La Jolla, CA, USA). The results were considered
statistically significant when P<0.05.
Results
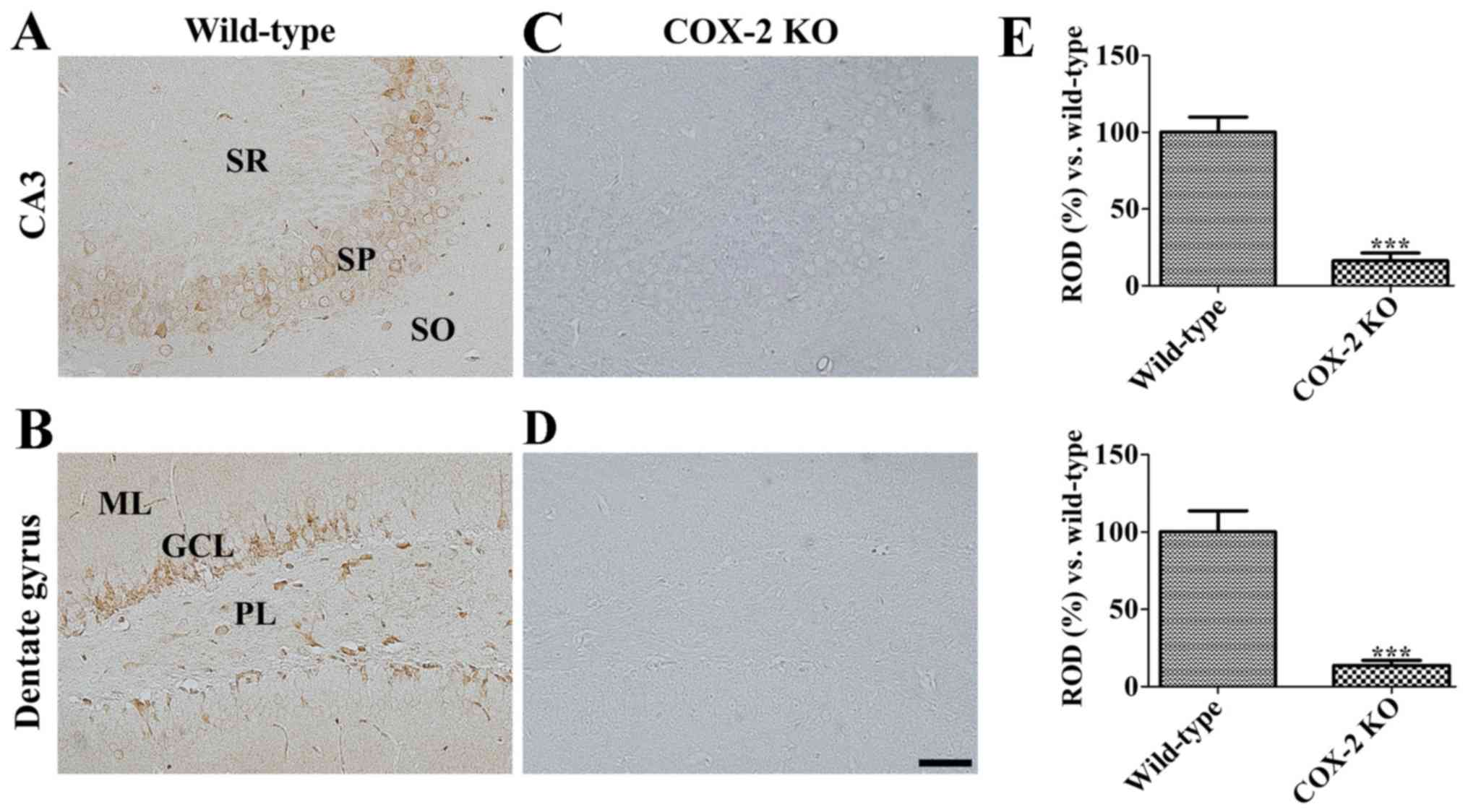
Confirmation of COX-2 antibody
specificity
Initially, COX-2 antibody specificity was assessed
by COX-2 immunohistochemical analysis performed on COX-2 KO and WT
mice. As a result, COX-2 immunoreactivity was found in both the
stratum pyramidale of the CA3 region and the granule cell layer of
the dentate gyrus in WT mice (Fig. 1A
and B). In contrast, the same was not detected in COX-2 KO mice
(Fig. 1C and D), who reported a
significantly decreased COX-2 immunoreactivity in both the
hippocampal CA3 region and the dentate gyrus when compared to WT
mice (Fig. 1E).
COX-2 immunoreactivity
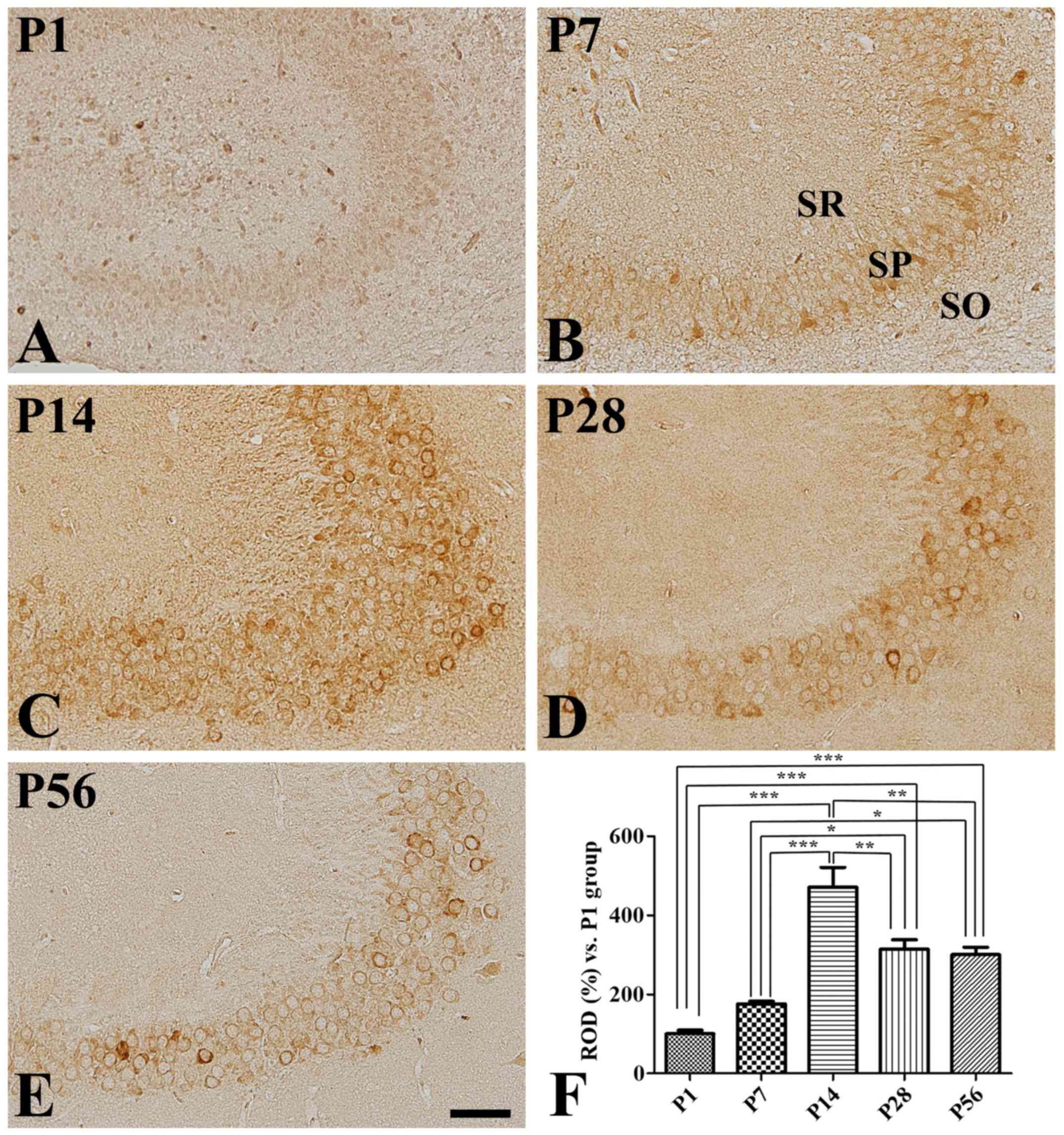
CA1 region
Although extremely weak COX-2 immunoreactivity was
detected in the hippocampal CA1 region at P1 (Fig. 2A), it was observed in the stratum
pyramidale of the CA1 region at P7 and P14, with an increase in
intensity with age (Fig. 2B, C and
F). In fact, strong COX-2 immunoreactivity was found in the
stratum pyramidale of the CA1 region at P14. In contrast, it
significantly decreased at P28 (Fig.
2F), during which it was only found in a few cells in the
stratum pyramidale (Fig. 2D).
However, COX-2 distribution pattern and immunoreactivity in the CA1
region at P56 was similar to those reported at P28 (Fig. 2E and F).
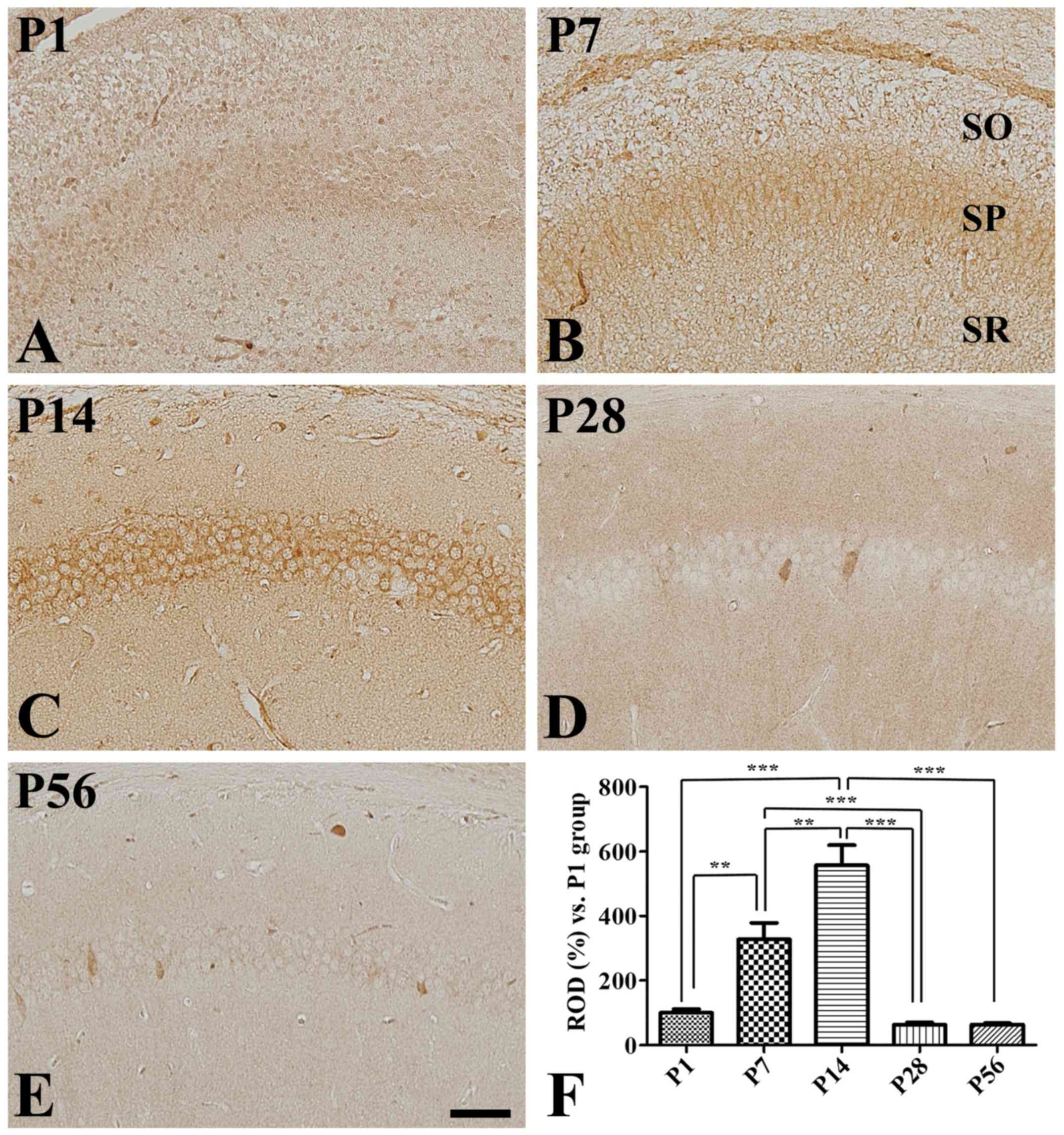
CA3 region
Although COX-2 immunoreactivity was weakly detected
in the CA3 region at P1 (Fig. 3A),
some strongly COX-2-immunoreactive cells were found at P7 in the
outer layer of the stratum pyramidale (Fig. 3B), where COX-2 immunoreactivity was
significantly increased compared to that at P1 (Fig. 3F). In concordance, strong COX-2
immunoreactive cells were found at P14 in most of the pyramidal
cells in this region (Fig. 3C).
However, COX-2 immunoreactivity was significantly decreased in the
CA3 region at P28 compared to that seen at P14, although it was
significantly higher than that found at either P1 or P7 (Fig. 3D and F). Finally, COX-2
distribution pattern and immunoreactivity at P56 were similar to
those reported at P28 (Fig. 3E and
F).
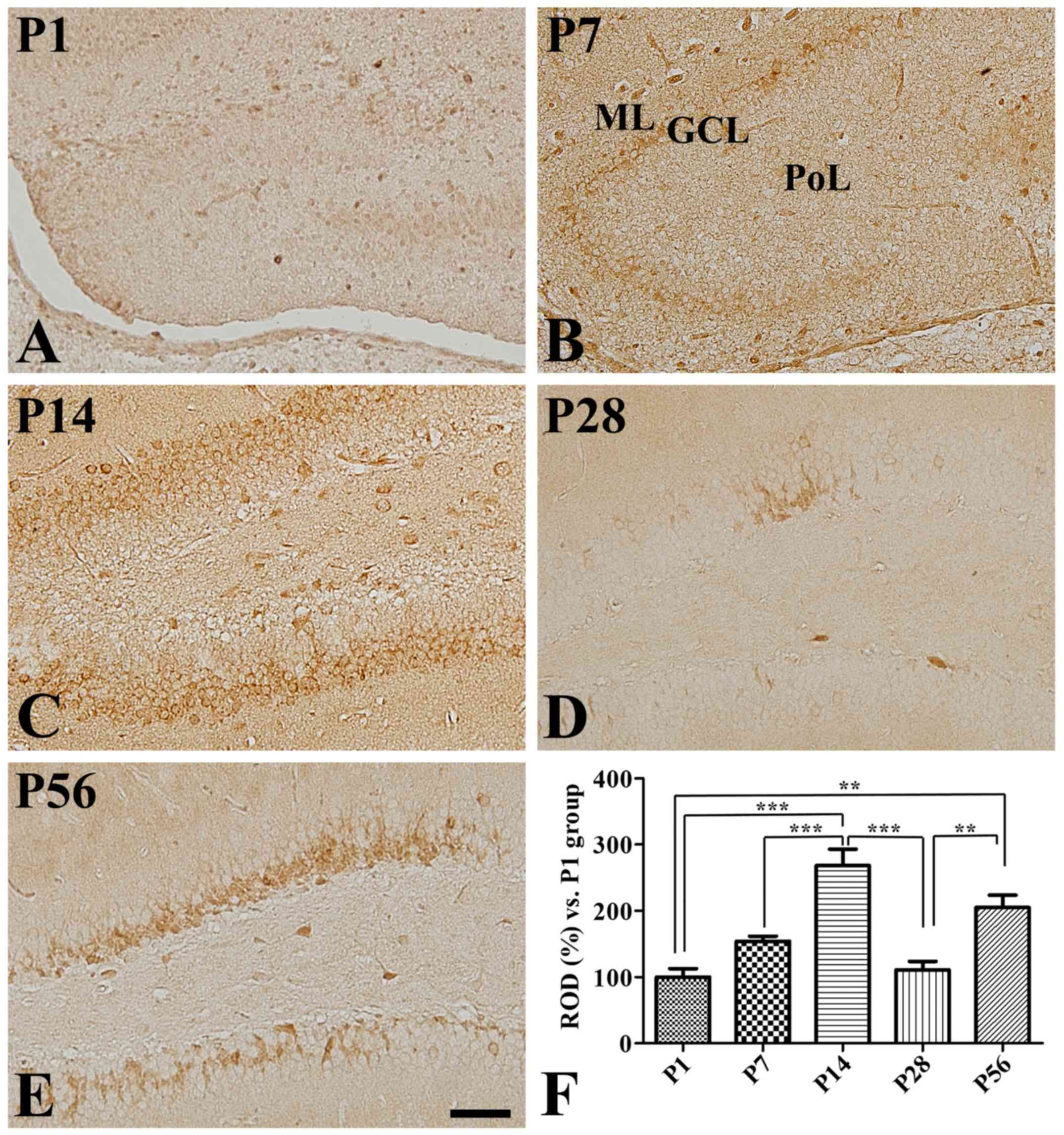
Dentate gyrus
Some COX-2 immunoreactivity was detected at P1 in
the dentate gyrus (Fig. 4A).
Furthermore, it was found in the outer granule cell layer of the
dentate gyrus at both P7 (Fig. 4B)
and P14 (Fig. 4C). Additionally, a
significant age-dependent increase in COX-2 immunoreactivity was
observed in the dentate gyrus at P14 (Fig. 4F). In contrast, COX-2
immunoreactivity was significantly decreased in the dentate gyrus
at P28 compared to that seen at P14, while immunoreactive neurons
were found in the inner half of the granule cell layer (Fig. 4D and F). However, COX-2
immunoreactivity was found in the inner granule cell layer at P56
and its expression levels were significantly increased in this
region compared to those found at P28 and were similar to those
seen at P14 (Fig. 4E and F).
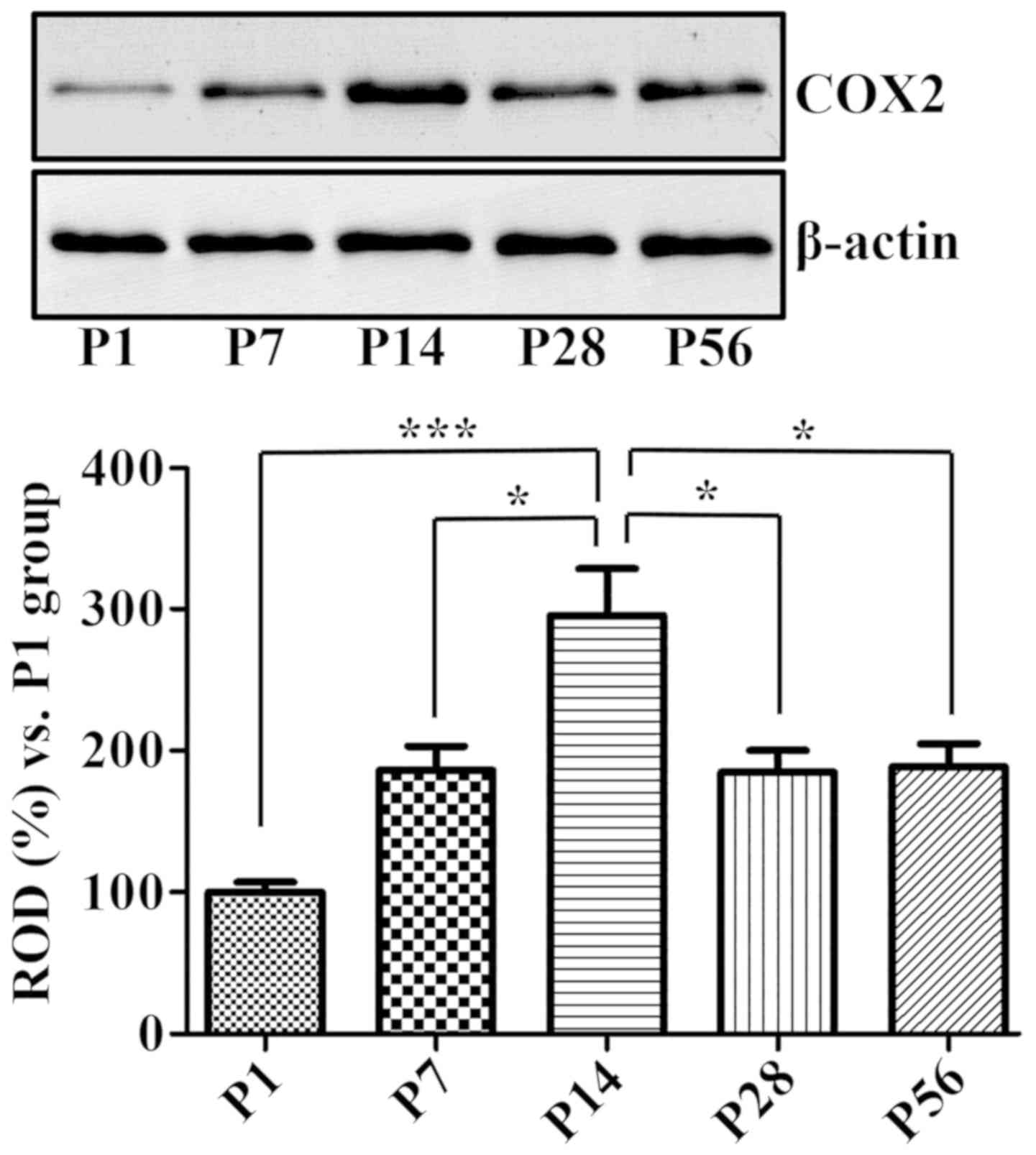
COX-2 protein levels
Western blot analysis produced similar results,
considering that the COX-2 protein levels in the hippocampal
homogenates were significantly increased at P14 and decreased at
both P28 and P56 (Fig. 5).
Effects of COX-2 depletion on the
synaptic plasticity
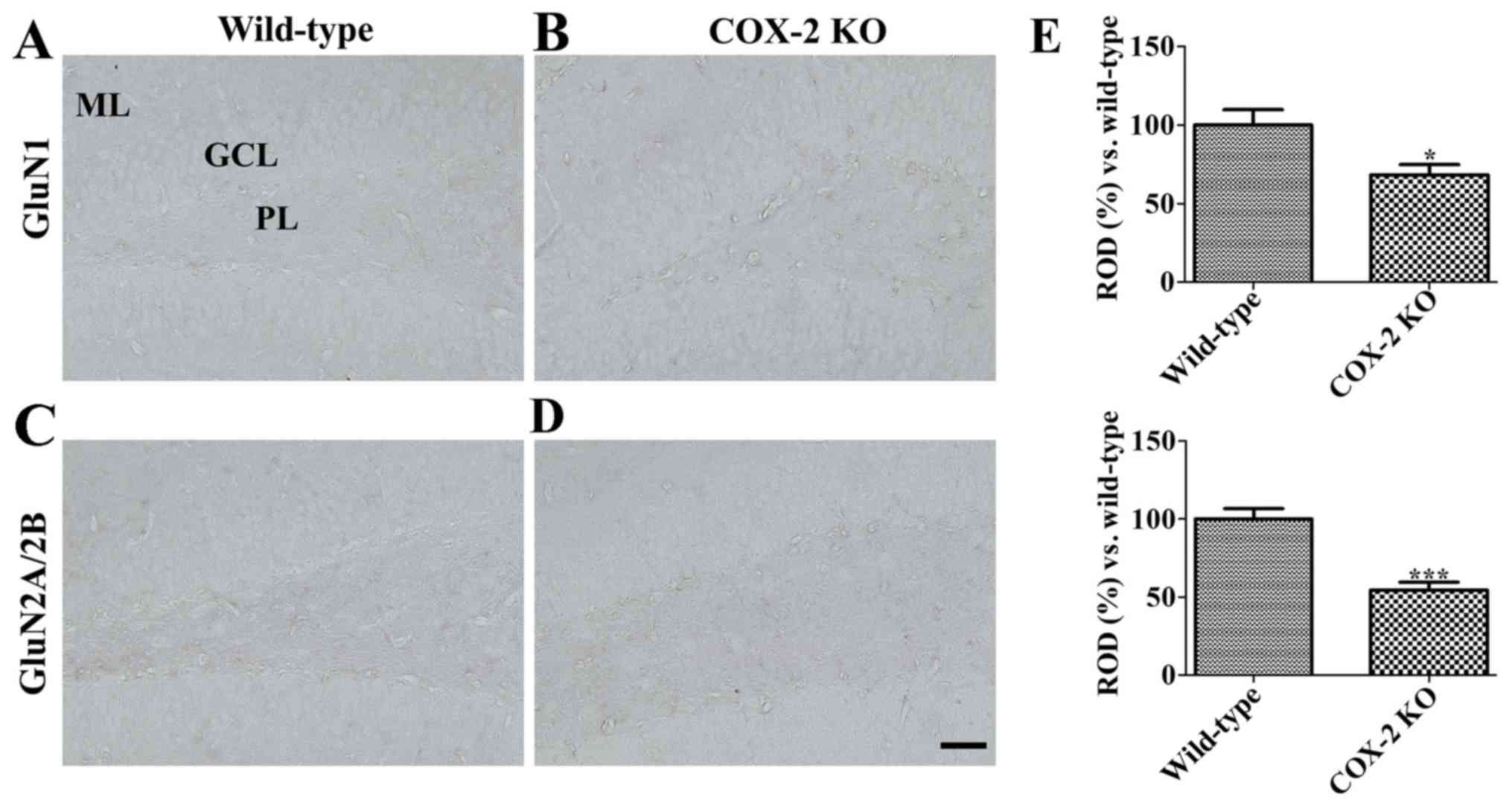
While GluN1 immunoreactivity was observed in the
granule cell layer of the dentate gyrus in WT mice (Fig. 6A), it was decreased in KO mice,
although it was still observed (Fig.
6B and E).
In concordance, while GluN2A/2B immunoreactivity was
mainly found in the polymorphic layer and the molecular layer of
the dentate gyrus in WT mice (Fig.
6C), it was weakly detected in KO mice (Fig. 6D), given that it was significantly
decreased (Fig. 6E).
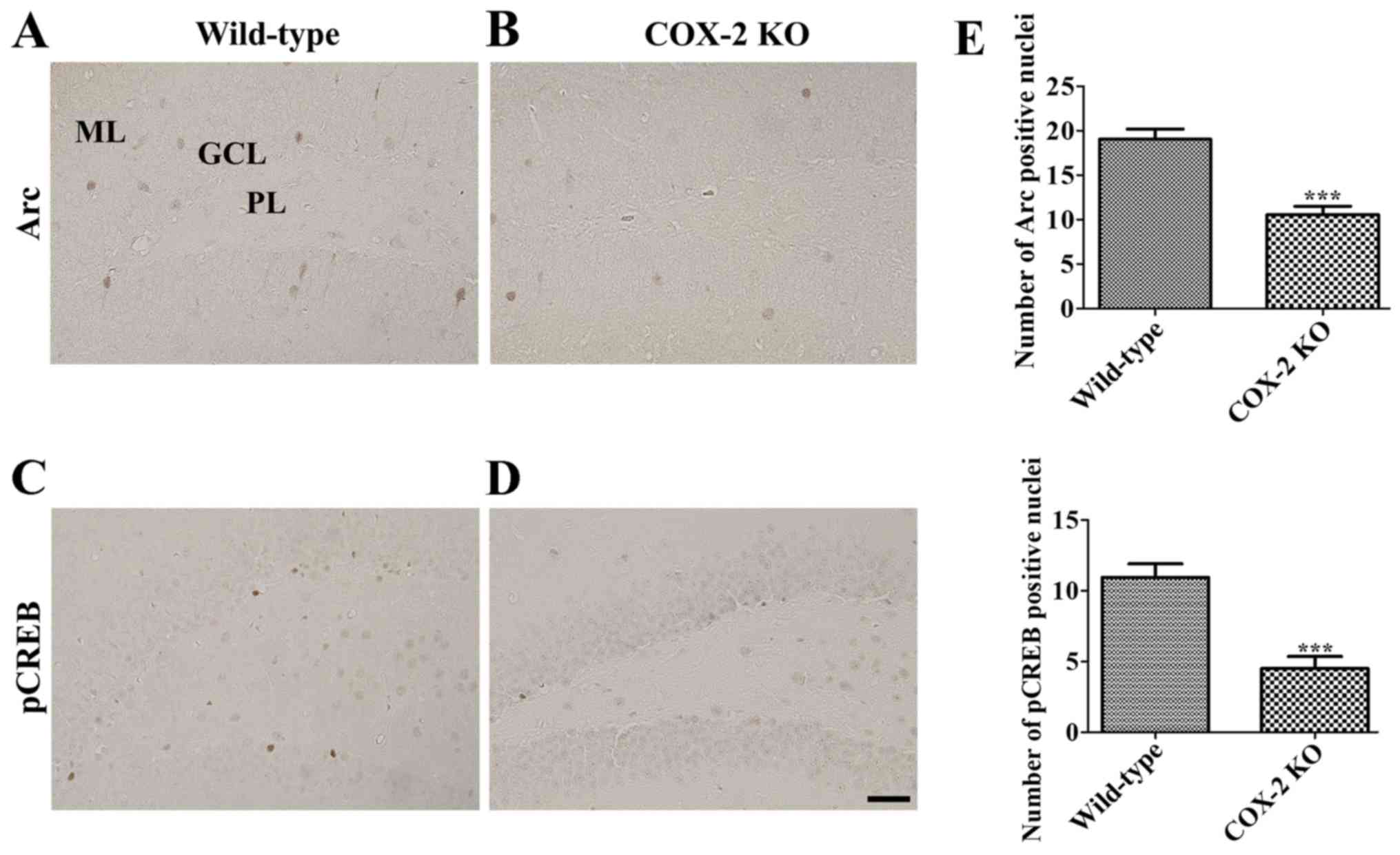
Furthermore, although Arc immunoreactivity was
observed in the granule cell layer of the dentate gyrus in WT mice
(Fig. 7A), Arc immunoreactive
cells were fewer in the dentate gyrus of KO mice than in that of WT
mice (Fig. 7B). In fact, the
number of Arc immunoreactive cells in the dentate gyrus of KO mice
was decreased by 55.5% than that of WT mice (Fig. 7E).
Finally, several pCREB immunoreactive nuclei were
detected in the subgranular zone of the dentate gyrus of WT mice
(Fig. 7C); however, fewer pCREB
immunoreactive nuclei were observed in the same area in KO mice
(Fig. 7D), who reported the number
of pCREB nuclei to be 41.3% of WT mice (Fig. 7E).
Discussion
Although the expression of COX-2, a rate-limiting
enzyme involved in prostanoid synthesis, is upregulated in
neurological disorders, such as ischemia and seizures, COX-2 is
constitutively expressed in some brain regions, including the
hippocampus. In the present study, we assessed COX-2
immunoreactivity and protein levels in the hippocampus considering
that COX-2 is correlated with the neuronal activity through the
GluN (14,15). Furthermore, we tested COX-2
antibody specificity based on the immunohistochemistry in the
hippocampus using COX-2 KO mice. As a result, while COX-2
immunoreactivity was found in both the stratum pyramidale of the
CA3 region and the granule cell layer of the dentate gyrus in WT
mice, it nearly disappeared in KO mice. In the present study, we
did not observe COX-2 immunoreactivity in either the dentate gyrus
or the hippocampal CA3 region at P7. This result is supported by
previous studies on rats that showed extremely little COX-2
expression immediately after birth (20–22).
Additionally, we observed a significant increase in COX-2
immunoreactivity at P14 in these regions compared to P7. This
result is in concordance with previous studies reporting that COX-2
is primarily expressed in neurons of intact animals, although it is
inducible by inflammation (15,24,25).
Moreover, a greater than twofold increase in constitutive COX-2
mRNA expression was observed in both the granule cell layer of the
dentate gyrus and the CA3 hippocampal region by P21, identifying
COX-2 adult levels (22). Conflict
evidence was demonstrated that COX-2 mRNA levels were similarly
detected by P11 in the hippocampus (26). In fact, in the rat cerebral cortex,
COX-2 expression increases two- to threefold between P14 and P30
when compared to P7 (20), while
no significant COX-2 mRNA expression was found in the cortex by P11
(26). In addition, we found COX-2
immunoreactivity in the hippocampal subregions of the mouse brain
and we observed that COX-2 expression peaked in the hippocampus at
P14 to decrease again at P28 and P56. This result is supported by a
previous study suggesting that COX-2 expression peaks during the
second postnatal week (27).
However, most studies have been investigated constitutive COX-2
mRNA expression, not protein expression in the hippocampus during
postnatal development. Furthermore, although we previously
demonstrated that doublecortin-positive neuroblasts are abundantly
detected in the somatosensory cortex by P14 and rarely detected at
P21 (4), we observed differential
localization of COX-2 with age between P7 and P56, shifting from
the outer to the inner granule cell layer.
Arc (activity-regulated cytoskeleton-associated
protein) is known to be rapidly up-regulated by external stimuli in
excitatory projection neurons to induce neural plasticity (28–30)
and is a factor established to be responsible for modulating memory
consolidation in a GluNs activation-dependent manner (28,31,32).
Arc is also closely related to adult neurogenesis in the
hippocampus (33,34). To elucidate the role of COX-2 on
hippocampal synaptic plasticity, we investigated Arc expression in
the hippocampus of COX-2 KO and WT mice. As a result, we observed
Arc expression in the granule cell layer of the dentate gyrus and
found the number of Arc-positive nuclei to be significantly
decreased in the dentate gyrus of COX-2 KO mice compared to that of
WT mice. This result suggests that COX-2 plays an important role in
the synaptic plasticity of the dentate gyrus. Furthermore, these
COX-2 changes may be associated with the first-time space
navigation ability of mice after birth. In fact, Langston et
al (35) demonstrated that
strong directional tuning was already apparent at P15 and P16 in
pups exploring for the first time the outside of their nest.
Considering that both COX-2 (36)
and Arc (29) are coupled to
neural activity in the hippocampus, hippocampal neural activity is
strongly coupled to navigation through the firing of place
cells.
Hippocampal GluNs are primarily composed of the
GluN1 and GluN2A or GluN2B subunits. In the current study, we
observed a reduction in GluN1 and GluN2A/2B immunoreactivity in the
hippocampal dentate gyrus of COX-2 KO mice compared to that seen in
WT mice. This result suggests that COX-2 has a role in modulating
the basal levels of GluN1 and GluN2A/2B in the hippocampus. While
GluN stimulation activates CREB and BDNF expression, extrasynaptic
GluN stimulation decreases it (37–39).
Furthermore, GluN participates in the formation of neuronal spines
and facilitates the proliferation of cells into neurons (40). GluN1 transgenic mice were shown to
report increases in proliferating neural progenitor cells in the
adult sugranular zone of dentate gyrus (41), whereas GluN2A KO mice were
described to present better exercise-induced amelioration of
proliferating neural progenitor cells compared to WT mice (42). In addition, GluN1 null mutant
embryo, COX-2 mRNA expression was significantly decreased compared
to that in the wide-type embryo (43). Therefore, the reduction in GluN1
and GluN2A/2B observed in COX-2 KO mice may be closely associated
with decreases in both the proliferating activity and synaptic
plasticity.
Moreover, we observed the effects of COX-2 depletion
on pCREB expression in the hippocampus given that GluNs stimulation
increases pCREB levels, acquiring a neuroprotective role (38). In addition, activation of both the
GluNs and BDNF signaling pathways can activate the phosphorylation
of CREB at Ser133 (pCREB), which is a rate-limiting step in CREB
signaling (44). Our results
reported that COX-2 depletion significantly decreased pCREB
immunoreactive nuclei in the subgranular zone of the dentate gyrus
of KO mice compared to the age-matched WT mice. This result is
consistent with a previous study indicating that genetic and
pharmacological inhibition of COX-2 prominently decreased the
number of pCREB-immunoreactive nuclei in the dentate gyrus
(17–19). In addition, luciferase assay
demonstrated that CREB was involved in the constitutive expression
of COX-2 in neurons (43).
Another possible reason behind such a phenomenon may
be that COX-2 plays a role in the Wnt-5a-dependent regulation of
the hippocampal synaptic structure and function; specifically,
Wnt-5a targets COX-2 via microRNA-101b (45) and has a key role in both the
synaptic structure and function of the adult nervous system
(46,47). In addition, COX-2 is associated
with stress-related functions in the brain, as endogenous
glucocorticoids modulate the levels of COX-2 and its downstream
target (i.e., microsomal PGE synthase-1) under conditions of stress
(48). Furthermore, blockade of
the PGE2 receptor 2 as well as COX-2 significantly
reduces the number of neural progenitor cells in the dentate gyrus
(49). However, more studies need
to be elucidated about the changes of proteins related to synaptic
plasticity. Overall, the modulation of COX-2 is closely related
with the synaptic plasticity in postnatal development and
hippocampal neurogenesis in adult brain. Long-term treatment of
COX-2 inhibitor may reduce the basal levels of hippocampal
neurogenesis and synaptic plasticity in the naïve young and adult
brain.
In conclusion, COX-2 is constitutively expressed in
both the granule cells of the dentate gyrus and the pyramidal cells
of the hippocampal CA1-3 regions at P14, although it thereafter
decreases. In particular, the localization of COX-2 expression
shifts from the outer to the inner granule cell layer of the
dentate gyrus with age. Furthermore, COX-2 depletion significantly
decreases the expression of synaptic markers, including Arc, GluN1,
GluN2A/2B, and pCREB, in the hippocampal dentate gyrus. Overall,
constitutively expressed COX-2 may be associated with synaptic
plasticity in the hippocampus during postnatal development.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea Government
(Ministry of Science, ICT and Future Planning) (grant no.
NRF-2016R1A2B4009156) and by the Korea Mouse Phenotyping Project
(grant nos. NRF-2015M3A9D5A01076747). In addition, this work was
supported by the Promising-Pioneering Researcher Program through
Seoul National University (SNU) in 2015 and by the Research
Institute for Veterinary Science, Seoul National University.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
HYJ and IKH designed the experiments and the study.
HYJ, DYY, SMN, JWK, WK, KYL and JHC looked after the animals and
performed the morphological experiments. HJK and DWK conducted
western blot analysis. YSY and JKS participated in designing and
discussing the study. HYJ and IKH wrote this manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal handling and care conformed to the guidelines
established by the current international laws and policies
[National Institutes of Health (NIH) Guide for the Care and Use of
Laboratory Animals; NIH Publication no. 85-23, 1985, revised 2011],
and were approved by both the Institutional Animal Care and Use
Committee (IACUC) of Seoul National University (approval no.:
SNU-120210-1 and SNU-140313-1) and Kangwon National University
(approval no.: KW-161226-1).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Isaacson RL and Pribram KH: The
Hippocampus. 3 and 4. Plenum; New York: 1985
|
|
2
|
Bayer SA: Development of the hippocampal
region in the rat. II. Morphogenesis during embryonic and early
postnatal life. J Comp Neurol. 190:115–134. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schlessinger AR, Cowan WM and Gottlieb DI:
An autoradiographic study of the time of origin and the pattern of
granule cell migration in the dentate gyrus of the rat. J Comp
Neurol. 159:149–175. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yoo DY, Yoo KY, Choi JW, Kim W, Lee CH,
Choi JH, Park JH, Won MH and Hwang IK: Time course of postnatal
distribution of doublecortin immunoreactive developing/maturing
neurons in the somatosensory cortex and hippocampal CA1 region of
C57BL/6 mice. Cell Mol Neurobiol. 31:729–736. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jung HY, Yim HS, Yoo DY, Kim JW, Chung JY,
Seong JK, Yoon YS, Kim DW and Hwang IK: Postnatal changes in
glucose transporter 3 expression in the dentate gyrus of the
C57BL/6 mouse model. Lab Anim Res. 32:1–7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yoo DY, Yoo KY, Park JH, Choi JW, Kim W,
Hwang IK and Won MH: Detailed differentiation of calbindin
D-28k-immunoreactive cells in the dentate gyrus in C57BL/6 mice at
early postnatal stages. Lab Anim Res. 27:153–159. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kujubu DA, Fletcher BS, Varnum BC, Lim RW
and Herschman HR: TIS10, a phorbol ester tumor promoter-inducible
mRNA from Swiss 3T3 cells, encodes a novel prostaglandin
synthase/cyclooxygenase homologue. J Biol Chem. 266:12866–12872.
1991.PubMed/NCBI
|
|
8
|
O'Banion MK, Winn VD and Young DA: cDNA
cloning and functional activity of a glucocorticoid-regulated
inflammatory cyclooxygenase. Proc Natl Acad Sci USA. 89:4888–4892.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yagami T, Koma H and Yamamoto Y:
Pathophysiological roles of cyclooxygenases and prostaglandins in
the central nervous system. Mol Neurobiol. 53:4754–4771. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sang N, Zhang J, Marcheselli V, Bazan NG
and Chen C: Postsynaptically synthesized prostaglandin E2 (PGE2)
modulates hippocampal synaptic transmission via a presynaptic PGE2
EP2 receptor. J Neurosci. 25:9858–9870. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Endres M, Dirnagl U and Moskowitz MA: The
ischemic cascade and mediators of ischemic injury. Handb Clin
Neurol. 92:31–41. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iadecola C, Niwa K, Nogawa S, Zhao X,
Nagayama M, Araki E, Morham S and Ross ME: Reduced susceptibility
to ischemic brain injury and N-methyl-d-aspartate-mediated
neurotoxicity in cyclooxygenase-2-deficient mice. Proc Natl Acad
Sci USA. 98:1294–1299. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kirkby NS, Zaiss AK, Urquhart P, Jiao J,
Austin PJ, Al-Yamani M, Lundberg MH, MacKenzie LS, Warner TD,
Nicolaou A, et al: LC-MS/MS confirms that COX-1 drives vascular
prostacyclin whilst gene expression pattern reveals non-vascular
sites of COX-2 expression. PLoS One. 8:e695242013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hewett SJ, Bell SC and Hewett JA:
Contributions of cyclooxygenase-2 to neuroplasticity and
neuropathology of the central nervous system. Pharmacol Ther.
112:335–357. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamagata K, Andreasson KI, Kaufmann WE,
Barnes CA and Worley PF: Expression of a mitogen-inducible
cyclooxygenase in brain neurons: Regulation by synaptic activity
and glucocorticoids. Neuron. 11:371–386. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jung HY, Yoo DY, Kim JW, Kwon HJ, Lee KY,
Choi JH, Kim DW, Chung JY, Yoon YS and Hwang IK: Age-associated
alterations in constitutively expressed cyclooxygenase-2
immunoreactivity and protein levels in the hippocampus. Mol Med
Rep. 15:4333–4337. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hwang IK, Yi SS, Yoo KY, Park OK, Yan B,
Kim IY, Kim YN, Song W, Moon SM, Won MH, et al: Effects of
treadmill exercise on cyclooxygenase-2 in the hippocampus in type 2
diabetic rats: Correlation with the neuroblasts. Brain Res 1341.
84–92. 2010. View Article : Google Scholar
|
|
18
|
Nam SM, Kim JW, Yoo DY, Choi JH, Kim W,
Jung HY, Won MH, Hwang IK, Seong JK and Yoon YS: Effects of
treadmill exercise on neural stem cells, cell proliferation, and
neuroblast differentiation in the subgranular zone of the dentate
gyrus in cyclooxygenase-2 knockout mice. Neurochem Res.
38:2559–2569. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nam SM, Kim JW, Yoo DY, Choi JH, Kim W,
Jung HY, Won MH, Hwang IK, Seong JK and Yoon YS: Comparison of
pharmacological and genetic inhibition of cyclooxygenase-2: Effects
on adult neurogenesis in the hippocampal dentate gyrus. J Vet Sci.
16:245–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hickey RW, Adelson PD, Johnnides MJ, Davis
DS, Yu Z, Rose ME, Chang YF and Graham SH: Cyclooxygenase-2
activity following traumatic brain injury in the developing rat.
Pediatr Res. 62:271–276. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kaufmann WE, Worley PF, Taylor CV, Bremer
M and Isakson PC: Cyclooxygenase-2 expression during rat
neocortical development and in Rett syndrome. Brain Dev. 19:25–34.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tocco G, Freire-Moar J, Schreiber SS,
Sakhi SH, Aisen PS and Pasinetti GM: Maturational regulation and
regional induction of cyclooxygenase-2 in rat brain: Implications
for Alzheimer's disease. Exp Neurol. 144:339–349. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Paxinos G and Franklin KBJ: The Mouse
Brain in Stereotaxic Coordinates. Academic Press; San Diego:
2001
|
|
24
|
Breder CD, Dewitt D and Kraig RP:
Characterization of inducible cyclooxygenase in rat brain. J Comp
Neurol. 355:296–315. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Breder CD: Cyclooxygenase systems in the
mammalian brain. Ann NY Acad Sci. 813:296–301. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Piscopo P, Bernardo A, Calamandrei G,
Venerosi A, Valanzano A, Bianchi D, Confaloni A and Minghetti L:
Altered expression of cyclooxygenase-2, presenilins and oxygen
radical scavenging enzymes in a rat model of global perinatal
asphyxia. Exp Neurol. 209:192–198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang MZ, Wang JL, Cheng HF, Harris RC and
McKanna JA: Cyclooxygenase-2 in rat nephron development. Am J
Physiol. 273:F994–F1002. 1997.PubMed/NCBI
|
|
28
|
Bloomer WA, VanDongen HM and VanDongen AM:
Arc/Arg3.1 translation is controlled by convergent
N-methyl-D-aspartate and Gs-coupled receptor signaling pathways. J
Biol Chem. 283:582–592. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guzowski JF, McNaughton BL, Barnes CA and
Worley PF: Environment-specific expression of the immediate-early
gene Arc in hippocampal neuronal ensembles. Nat Neurosci.
2:1120–1124. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guzowski JF, Lyford GL, Stevenson GD,
Houston FP, McGaugh JL, Worley PF and Barnes CA: Inhibition of
activity-dependent arc protein expression in the rat hippocampus
impairs the maintenance of long-term potentiation and the
consolidation of long-term memory. J Neurosci. 20:3993–4001. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Holloway CM and McIntyre CK: Post-training
disruption of Arc protein expression in the anterior cingulate
cortex impairs long-term memory for inhibitory avoidance training.
Neurobiol Learn Mem. 95:425–432. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Steward O and Worley P: Local synthesis of
proteins at synaptic sites on dendrites: Role in synaptic
plasticity and memory consolidation? Neurobiol Learn Mem.
78:508–527. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kuipers SD, Trentani A, Tiron A, Mao X,
Kuhl D and Bramham CR: BDNF-induced LTP is associated with rapid
Arc/Arg3.1-dependent enhancement in adult hippocampal neurogenesis.
Sci Rep. 6:212222016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meconi A, Lui E and Marrone DF: Sustained
Arc expression in adult-generated granule cells. Neurosci Lett.
603:66–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Langston RF, Ainge JA, Couey JJ, Canto CB,
Bjerknes TL, Witter MP, Moser EI and Moser MB: Development of the
spatial representation system in the rat. Science. 328:1576–1580.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kaufmann WE, Worley PF, Pegg J, Bremer M
and Isakson P: COX-2, a synaptically induced enzyme, is expressed
by excitatory neurons at postsynaptic sites in rat cerebral cortex.
Proc Natl Acad Sci USA. 93:2317–2321. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hardingham GE, Fukunaga Y and Bading H:
Extrasynaptic NMDARs oppose synaptic NMDARs by triggering CREB
shut-off and cell death pathways. Nat Neurosci. 5:405–414. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hardingham GE and Bading H: Synaptic
versus extrasynaptic NMDA receptor signalling: Implications for
neurodegenerative disorders. Nat Rev Neurosci. 11:682–696. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Parsons MP and Raymond LA: Extrasynaptic
NMDA receptor involvement in central nervous system disorders.
Neuron. 82:279–293. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nacher J and McEwen BS: The role of
N-methyl-d-aspartate receptors in neurogenesis. Hippocampus.
16:267–270. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bursztajn S, Falls WA, Berman SA and
Friedman MJ: Cell proliferation in the brains of NMDAR NR1
transgenic mice. Brain Res 1172. 10–20. 2007. View Article : Google Scholar
|
|
42
|
Kitamura T, Mishina M and Sugiyama H:
Enhancement of neurogenesis by running wheel exercises is
suppressed in mice lacking NMDA receptor epsilon 1 subunit.
Neurosci Res. 47:55–63. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hewett SJ, Shi J, Gong Y, Dhandapani K,
Pilbeam C and Hewett JA: Spontaneous glutamatergic synaptic
activity regulates constitutive COX-2 expression in neurons:
Opposing roles for the transcription factors CREB (cAMP response
element binding) protein and Sp1 (stimulatory protein-1). J Biol
Chem. 291:27279–27288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gonzalez GA, Yamamoto KK, Fischer WH, Karr
D, Menzel P, Biggs W III, Vale WW and Montminy MR: A cluster of
phosphorylation sites on the cyclic AMP-regulated nuclear factor
CREB predicted by its sequence. Nature. 337:749–752. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Codocedo JF and Inestrosa NC:
Wnt-5a-regulated miR-101b controls COX2 expression in hippocampal
neurons. Biol Res. 49:92016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bian WJ, Miao WY, He SJ, Wan ZF, Luo ZG
and Yu X: A novel Wnt5a-Frizzled4 signaling pathway mediates
activity-independent dendrite morphogenesis via the distal PDZ
motif of Frizzled 4. Dev Neurobiol. 75:805–822. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Farías GG, Alfaro IE, Cerpa W, Grabowski
CP, Godoy JA, Bonansco C and Inestrosa NC: Wnt-5a/JNK signaling
promotes the clustering of PSD-95 in hippocampal neurons. J Biol
Chem. 284:15857–15866. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ma Y, Matsuwaki T, Yamanouchi K and
Nishihara M: Cyclooxygenase-2-related signaling in the hypothalamus
plays differential roles in response to various acute stresses.
Brain Res 1508. 23–33. 2013. View Article : Google Scholar
|
|
49
|
Ma Y, Matsuwaki T, Yamanouchi K and
Nishihara M: Glucocorticoids suppress the protective effect of
Cyclooxygenase-2-related signaling on hippocampal neurogenesis
under acute immune stress. Mol Neurobiol. 54:1953–1966. 2017.
View Article : Google Scholar : PubMed/NCBI
|