Introduction
Recently, more studies investigating natural
materials have been conducted in response to the increased public
interest in healthy skin. There is particular interest in the
prevention and treatment of various skin diseases, including atopic
dermatitis and inflammatory acne (1). Atopic dermatitis (AD), also known as
inflammatory dermatitis, is characterized by severe itching,
chronic edema and skin rashes (2).
Although the etiology is not clearly understood, it is known that
AD is associated with immunological abnormalities, abnormal skin
barriers, and genetic and environmental factors (3). Exposure to allergens induces atopy,
leading to an imbalance between T helper cell (Th) 1 and Th2
cytokines, and the overproduction of Th2 cytokines stimulates
B-cells to increase the generation of immunoglobulin E (4). Increased immunoglobulin E (IgE)
induces the exocytosis of compounds such as histamine in skin mast
cells, causing edema and itching, thereby aggravating AD (4). Treatment methods for dermatitis
include moisturizing dry skin, the use of steroids as
anti-inflammatory agents, the application or administration of
antihistamines and immunosuppressive agents. Although these drugs
may relieve certain symptoms, long-term use of topical steroids may
additionally lead to thinning of the skin with subsequent bleeding
(5,6). Therefore, a number of ongoing studies
are attempting identify functional substances from safe and
effective natural products to prevent AD without side effects
(7,8).
The inflammatory response is a physiological
protective activity in the human body that recognizes external
physical and chemical stimuli, and employs a defensive mechanism to
restore damaged tissues (9).
Inflammatory reactions are categorized as acute or chronic
depending on their activation and duration (10). When an inflammatory reaction
occurs, inflammatory mediators such as nuclear factor-κB (NF-κB),
nitric oxide (NO) and inflammatory cytokines are secreted. NF-κB is
widely distributed and is known as a regulator of immune and
inflammatory responses (11,12).
NO, an indicator of the inflammatory response, is synthesized by NO
synthase (NOS) from L-arginine. There are three types of NOS:
Endothelial, neuronal and inducible NOS (iNOS). Of these, NO
produced by iNOS serves an important pathological role (13). Excessive NO production by iNOS
induced by lipopolysaccharide (LPS) or inflammatory cytokines
exacerbates the inflammatory response, resulting in tissue damage,
gene mutations and nerve damage (14). Cyclooxygenase (COX) is an enzyme
that catalyzes the conversion of arachidonic acid to prostaglandin.
COX-2 is induced by various stimuli including inflammation, growth
factors and cytokines released by cancer cells (15). The proinflammatory cytokines that
are enriched during inflammation include tumor necrosis factor
(TNF)-α, interleukin (IL)-6 and IL-1β, and are secreted by
macrophages to mediate various inflammatory responses that induce
NO generation. Excessive production of NO and cytokines can lead to
genetic alterations, and tissue and nerve damage, with fatal
consequences for the host (16).
Therefore, regulation of the inflammatory response is important for
maintaining homeostasis, and an anti-inflammatory effect may be
achieved by inhibiting the production of mediators such as NO and
cytokines, which are produced in inflammatory reactions (17).
Other signaling molecules involved in inflammatory
reactions include mitogen-activated protein kinases (MAPKs), which
can be characterized as three different types of serine/threonine
protein kinases: Extracellular signal-regulated protein kinase
(ERK), stress-activated c-Jun N-terminal kinase (JNK) and P38
kinases. Each type of kinase has a different activity, and all 3
types are known to carry out signal transduction in various
cellular activities, including proliferation, differentiation, cell
death and inflammation (18).
Efficient inhibition of these inflammatory mediators may be an
important strategy in the development of anti-inflammatory
drugs.
Dendropanax morbifera (D. morbifera)
is one of the world's rarest warm climate trees. There is only one
species in one genus in Korea, and it is found across a broad area
of western and southern coastal Korea encompassing Jeju Island,
Wando, Geomundo and Haenam; however, the native distribution has
been impacted by overharvesting and urbanization, and D.
morbifera now only grows in a limited area (19). D. morbifera, known as
Dendro, means ‘omnipotent drug’ (Panax), and has been called the
panacea tree. Various ongoing studies on D. morbifera are
aiming to determine its medicinal uses (20–24).
D. morbifera leaves are composed of 70.2% water, 1.2%
protein, 2.7% fat, 1.7% ash, 56.9 mg% vitamin C and 746 mg% water
soluble tannins; 32 types of substances have been identified with
gas chromatography-mass spectrometry. Among these substances are
β-selinene and capnellene-8-one, which are sesquiterpenes with two
ring structures, as well as many unidentified volatile components
(20). When D. morbifera
bark is scarred, it produces a golden resinous solution that
contains benzoic acid, a unique aromatic component (21,22).
Recent studies have revealed that polyacetylene
compounds isolated from D. morbifera leaves have
anti-complement activity, whereas ethanol extracts promote the
actions of B- and T-cells (20,23).
D. morbifera sap inhibited melanin biosynthesis by reducing
the expression of tyrosinase in melanin synthesis, and the ethyl
acetate fraction extracted from D. morbifera leaves also
exhibited cell protective and skin whitening effects, such as
antioxidant properties and tyrosinase inhibition (24). However, few studies or animal
experiments have been conducted on the mechanism underlying its
anti-inflammatory properties.
In the present study, the effects of D.
morbifera leaf (DPL) extract on the production of NO, cytokine
secretion and inflammation-associated protein expression was
investigated in activated RAW264.7 macrophages by inducing
inflammation with LPS. The anti-inflammatory effect of DPL was also
determined in a 2,4-dinitrochlorobenzene (DNCB)-induced AD animal
model.
Materials and methods
Chemicals, drugs and antibodies
Dulbecco's modified Eagle's medium (DMEM),
penicillin-streptomycin, and 10% fetal bovine serum (FBS) were
purchased from Welgene (Gyeongsan, Korea). LPS, MTT and dimethyl
sulfoxide (DMSO) were purchased from Merck KGaA (Darmstadt,
Germany). The nitrate/nitrite colorimetric assay kit was obtained
from Cayman Chemical Company (Ann Arbor, MI, USA). The mouse TNF
ELISA (cat. no. 560478) and mouse IL-6 ELISA kits (cat. no. 555240)
were purchased from BD Biosciences (San Jose, CA, USA). β-actin
(cat. no. 4967), iNOS (cat. no. 2982), COX-2 (cat. no. 4842),
phosphorylated (p)-NF-κB-p65 (cat. no. 3033), NF-κB-p65 (cat. no.
8242), p-NF-κB inhibitor-α (IκB-α; cat. no. 4812), IκB-α (cat. no.
5209) p-ERK1/2 (cat. no. 4376), ERK1/2 (cat. no. 9194), p-P38 MAPK
(cat. no. 9211), P38 MAPK (cat. no. 8690), p-JNK (cat. no. 4668),
JNK (cat. no. 9252) and anti-rabbit horseradish peroxidase (HRP;
cat. no. 7074) antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA).
Plant materials and extraction
The D. morbifera leaves used in the
experiments were collected from the native areas of Jeju Island.
The collected D. morbifera leaves (100 g) were pulverized
and then fermented with 70% ethanol and tertiary distilled water at
a ratio of 4:6 for 10 days at room temperature, then subjected to
hydrothermal extraction. The extracted solution was filtered using
Whatman No. 1 disc paper. Then the extract was concentrated under
reduced pressure using a rotary vacuum evaporator (R-220; BUCHI
Corporation, New Castle, DE, USA), and the remaining solution was
boiled again to obtain an extract (25). The extracts were mixed with
distilled water (DW) and refrigerated.
Cell culture and stimulation
The RAW264.7 macrophage line was obtained from the
Korean Cell Line Bank (Korean Cell Line Research Foundation, Seoul,
Korea), and maintained in DMEM supplemented with 5% FBS/1%
penicillin-streptomycin at 37°C in a 5% CO2 humidified
air environment. The cells were incubated for 24 h in medium
supplemented with 10% FBS. Subsequently, the RAW 264.7 cells were
pre-treated with doses of DPL (0, 100, 200, 300, 400 and 500 µg/ml)
for 2 h at 37°C and treated with LPS (1 µg/ml) for 24 h at 37°C, in
serum-free media.
Cell viability assay
RAW264.7 cells were seeded in a 96-well plate at a
density of 1×105 cells/ml and a volume of 200 µl/well.
Following incubation for 24 h at 37°C, the cells were treated with
DPL extract (mixed with DW) and negative control (DW) at various
concentrations (DPL 0, 100, 200, 300, 400 and 500 µg/ml) for 24 h
at 37°C, followed by the addition of 5 mg/ml MTT solution to each
well, and the plates were further incubated for 2 h at 37°C. The
supernatant was removed and 200 µl DMSO was added to each well to
solubilize the water-insoluble purple formazan crystals. The
absorbance at a wavelength of 595 nm was measured using a
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The percentage of viable cells compared with that of untreated
control cells was then estimated.
Measurement of NO
RAW264.7 cells were seeded (1×105/ml) and
cultured in 96-well plates. Following incubation for 24 h at 37°C,
the cells were treated with DPL extract at the indicated
concentrations (DPL 0, 100, 200, 300 and 400 µg/ml) for 2 h in
serum-free medium prior to the addition of LPS (1 µg/ml). Following
a 24-h incubation at 37°C, the supernatants were measured for NO
production using the nitrate/nitrite assay kit (Cayman Chemical
Company). NO was measured as the accumulation of nitrite and
nitrate reductase, which were determined spectrophotometrically
using Griess reagent, included in the nitrate/nitrite assay kit, at
an optical density of 540 nm.
Determination of TNF-α and IL-6
production
RAW 264.7 cells were pre-treated with doses of DPL
(0, 100, 200, 300 and 400 µg/ml) for 2 h and treated with LPS (1
µg/ml) for 24 h at 37°C. Subsequently, production of the
proinflammatory cytokines TNF-α and IL-6 in the culture medium was
determined using commercially available ELISA kits (BD
Biosciences), according to the manufacturer's protocol.
Western blot analysis
Cells were preincubated with various concentrations
of DPL extract (0, 200 and 400 µg/ml) for 2 h prior to a 24-h
incubation at 37°C with LPS (1 µg/ml), and subsequently harvested.
Western blot assays were performed as previously described
(26). The membranes were
incubated with the primary antibodies specific for β-actin
(1:10,000), p-NF-κB-p65 (1:1,000), NF κB-p65 (1:1,000), p-IκB-α
(1:1,000), IκB-α (1:1,000) p-ERK1/2 (1:1,000), ERK1/2 (1:1,000),
p-P38 MAPK (1:1,000), P38 MAPK (1:1,000), p-JNK (1:1,000) and JNK
(1:1,000) overnight at 4°C with gentle shaking. Following
incubation with the primary antibodies, the membranes were
incubated with HRP-conjugated anti-rabbit IgG secondary antibodies
(1:1,000) for 2 h at room temperature with gentle shaking. The
membranes were washed three times for 10 min in TBS containing 0.1%
Tween-20. The bands were detected using enhanced chemiluminescence
western blotting detection reagents (Pierce; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. β-actin was used as a loading control.
Band density was measured using ImageJ software (version 1.48;
National Institutes of Health, Bethesda, MD, USA).
Animals
A total of 30 BALB/c female mice (age, 4 weeks; body
weight, 16–18 g) were purchased from the Nara Biotech, Co., Ltd.
(Seoul, South Korea) and maintained at 23±5°C at 40±10% relative
humidity with a 12-h light/dark cycle (artificial lighting from
8:00 a.m. to 8:00 p.m.) in facilities approved by the Companion and
Laboratory Animal Science Department of Kongju National University
(Chungnam, Korea). The animals were housed in cages and allowed
access to sterilized water and commercial rodent chow (Biopia,
Seoul, Korea) ad libitum. All animal experiments were
performed with the approval of the Institutional Animal Care and
Use Committee of Kong-Ju National University (approval no.
KNU_2017-10; Yesan, Korea) and the institutional guidelines were
adhered to.
Induction of AD
AD was induced in BALB/c mice as previously
described, with minor modifications (27). Briefly, BALB/c mice were divided
into seven groups (n=5/group): Group I (control treatment; 3:1
acetone/olive oil solution; 20 µl/ear), group II (1% DNCB; 20
µl/ear), groups III (dexamethasone; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany; 50 µg/20 µl/ear; positive control), group IV
(DPL high; 20 µl/ear, undiluted solution), group V (DPL medium; 20
µl/ear, 0.5× undiluted solution/DW) and group VI (DPL low; 20
µl/ear, 0.25× undiluted solution/DW) received DPL extract or
combinations as indicated. To induce AD, the surfaces of both ears
of the mice (not anesthetized) were stripped with surgical tape.
Following this, 1% DNCB dissolved in a 3:1 acetone/olive oil
solution was painted on each ear. The DPL extract was applied at
three concentrations: High (undiluted solution mixed with an equal
volume of 3:1 solution of acetone and olive oil, v/v), medium (0.5×
undiluted solution/DW mixed with an equal volume of 3:1 solution of
acetone and olive oil, v/v) or low (0.25× undiluted solution/DW
mixed with an equal volume of 3:1 solution of acetone and olive
oil, v/v). DNCB was applied every 2 days starting from 3 days prior
to measurements. Dexamethasone and DPL extract were applied every 2
days from day 0. Following 24 h of DNCB application, the thickness
of the ears was measured with a vernier caliper (Mitutoyo,
Kawasaki, Japan). The mice were sacrificed on Day 10. Blood samples
were then collected from the abdominal aorta, and the plasma was
stored at −70°C until further analysis. Following sacrifice, the
ears were excised and subjected to histopathological analysis.
Measurement of Ig levels
Blood samples were obtained from each treatment
group 10 days following AD induction. Total serum IgE levels were
measured using an ELISA kit (cat. no. K3231082; Komabiotech, Seoul,
Korea), according to the manufacturer's protocol.
Histological observations
The ears were immediately fixed in 10% formaldehyde
(1 week at room temperature) following excision, and embedded in
paraffin. Blocks were then cut into 5-µm-thick slices. To measure
epidermal thickening, hematoxylin and eosin (H&E) staining was
performed (6 h at room temperature). To evaluate mast cells, the
skin sections were stained with toluidine blue (TB; 6 h at room
temperature). The sections were examined under a light microscope
(magnification, ×200; Olympus CH30; Olympus Corporation, Tokyo,
Japan).
Statistical analysis
The results are expressed as the mean ± standard
deviation. The experiments were repeated three times. Statistical
analyses were performed using Prism (version 5; GraphPad Software,
Inc., La Jolla, CA, USA) Differences between the mean values for
the individual groups were assessed by one-way analysis of variance
with Dunnett's post hoc test. P<0.05 was considered to indicate
a statistically significant difference.
Results
Effects of DPL extract on the
viability of RAW264.7 macrophages
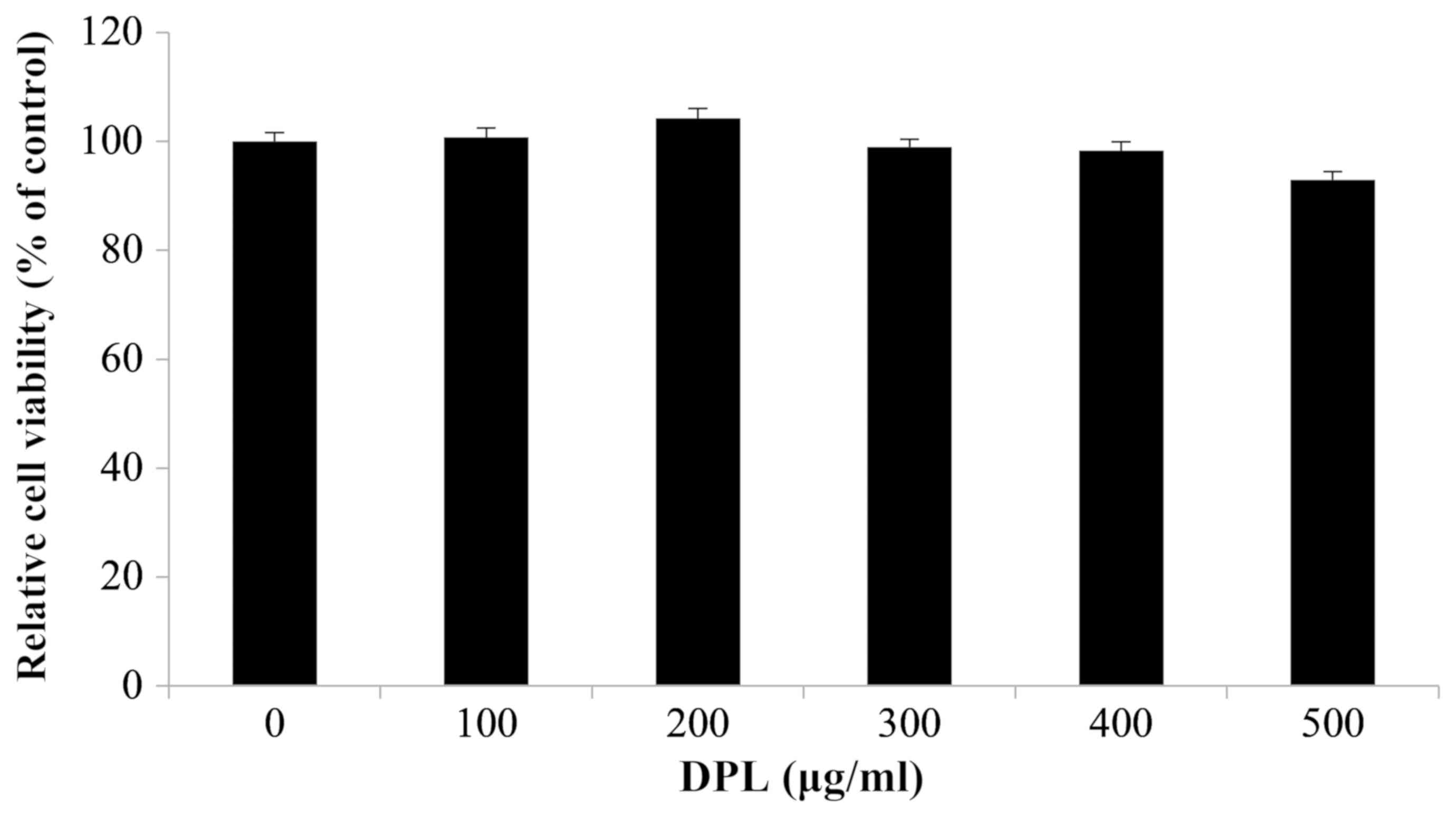
To investigate the effect of DPL extract on the
survival of RAW264.7 macrophages, an MTT assay was performed. Cell
viability was measured once RAW264.7 cells were treated with 0,
100, 200, 300, 400 and 500 µg/ml DPL extract and cultured for 24 h.
There were no differences in cell viability at any concentration,
confirming that the DPL extract did not affect the survival of
RAW264.7 cells (Fig. 1). However,
cell viability following treatment with DPL at a concentration of
500 µg/ml was decreased compared with other concentrations.
Therefore, concentrations of 100, 200, 300 and 400 µg/ml were
used.
Effects of DPL extract on LPS-induced
NO production
NO is an inorganic compound produced by NO synthase.
It is involved in many biological processes including the immune
response, cytotoxicity and vascular relaxation, and it also
maintains cell function and cytotoxicity depending on the
concentration (28). The present
study selected the concentration of DPL extract that had no effect
on the survival of RAW264.7 macrophages and performed a NO assay.
RAW264.7 cells were treated with DPL extract (100, 200, 300 and 400
µg/ml) for 2 h before treatment with LPS (1 µg/ml), and the
concentration of NO was measured (Fig.
2A). The NO concentration in RAW264.7 cells stimulated with LPS
was ~5 times higher compared with that observed in the untreated
control group. In the groups treated with 100, 200, 300 and 400
µg/ml DPL extract with LPS, the NO concentrations were 68.9, 51.2,
28.1 and 24.4%, respectively, indicating that the NO concentration
decreased in a DPL concentration-dependent manner when compared
with the group treated with LPS alone (Fig. 2A).
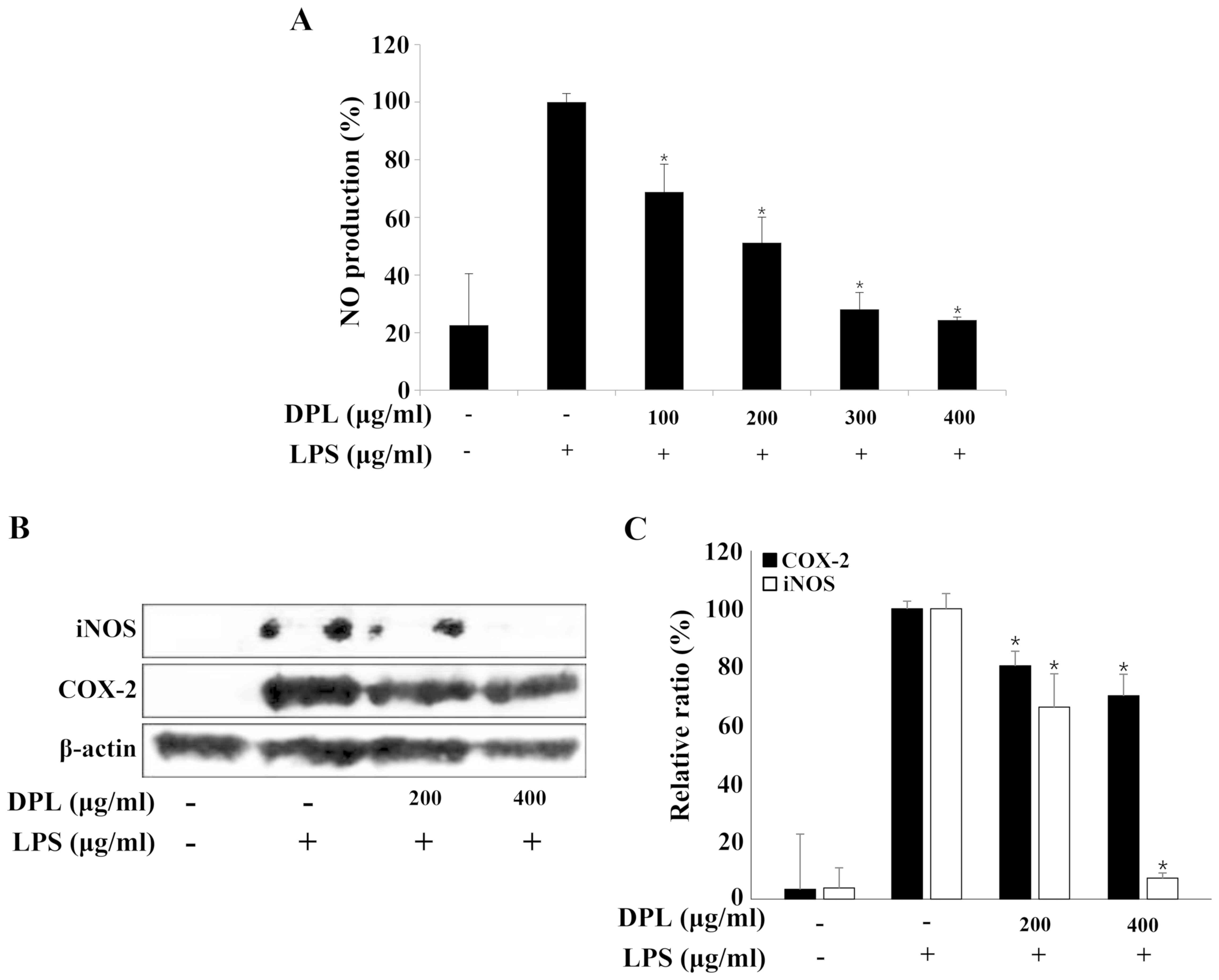 | Figure 2.Effects of DPL extract on LPS-induced
iNOS and COX-2 protein levels and NO production in RAW264.7 cells.
(A) RAW264.7 cells were pre-incubated with 100, 200, 300, and 400
µg/ml DPL extract for 2 h, and then treated with 1 µg/ml LPS for an
additional 24 h. NO was measured using the Griess reaction. (B) The
cells were sampled and lysed following 24 h treatment, and iNOS and
COX-2 protein levels were determined by western blotting. β-actin
was used as a loading control. (C) Data analysis was performed
using ImageJ software by measuring the integrated band densities
following background subtraction. Each bar represents the mean ±
standard deviation calculated from three independent experiments.
*P<0.05 vs. LPS only treatment. DPL, Dendropanax
morbifera leaf; LPS, lipopolysaccharide; iNOS, inducible nitric
oxide synthase; NO, nitric oxide; COX2, cyclooxygenase 2. |
Effects of DPL extract on LPS-induced
iNOS and COX-2 protein expression
The protein expression levels of iNOS and COX-2,
which are known to activate inflammation, were assessed by western
blotting. The protein expression levels of iNOS and COX-2 were
increased in the LPS-treated group when compared with the control
group, and these levels were significantly decreased in a
concentration-dependent manner in the groups treated with different
concentrations of DPL (Fig. 2B and
C).
Effects of DPL extract on LPS-induced
proinflammatory cytokine production
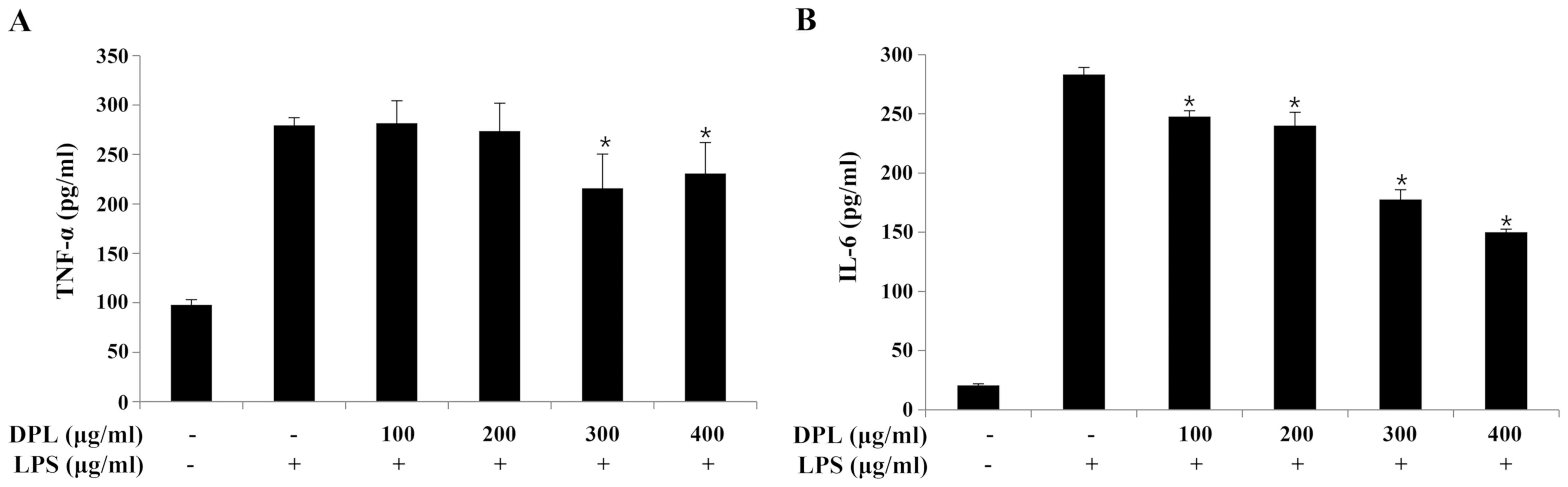
The expression of TNF-α and IL-6 was quantified by
ELISA to assess the anti-inflammatory effects of DPL extract. When
compared with the expression levels in the group treated with LPS
alone, the expression of TNF-α was significantly decreased
following treatment with 300 and 400 µg/ml DPL, and the expression
of IL-6 was significantly decreased in a concentration-dependent
manner (Fig. 3).
Effects of DPL extract on the
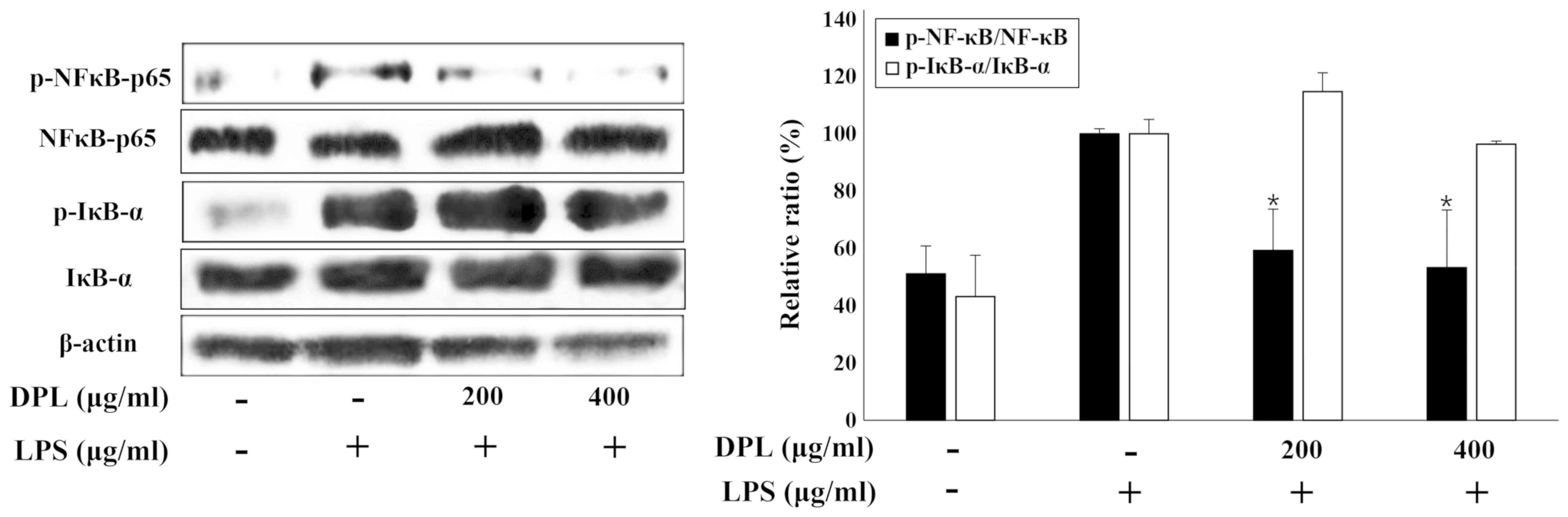
activation of NF-κB in LPS-stimulated RAW264.7 macrophages
NF-κB is activated by IκB-α and acts as an
inflammatory mediator (11). To
investigate the activation of the well-known inflammatory
mediators, NF-κB and IκB-α, the present study performed western
blot analysis. The expression of NF-κB and IκB-α was increased in
the LPS-treated group when compared with the untreated control
group. Comparisons of the LPS and 200 or 400 µg/ml DPL extract
groups and the LPS-only group revealed that the activation of NF-κB
was significantly decreased with DPL extract treatment, whereas the
activation of IκB-α was not significantly changed (Fig. 4).
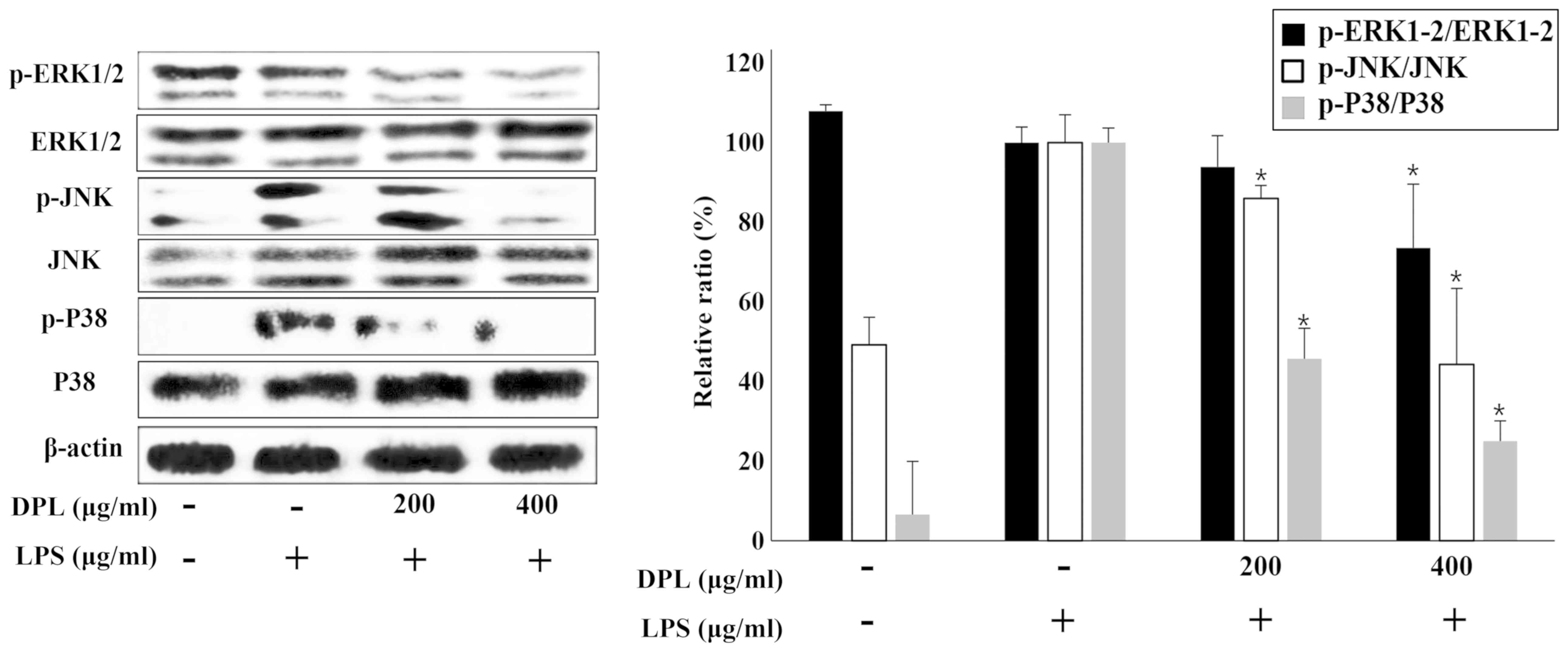
Effects of DPL extract on LPS-induced
NF-κB and MAPK phosphorylation
MAPK is a typical signaling molecule that affects
the activation of NF-κB. When MAPK is phosphorylated and activated
in the cell, it influences the production of various inflammatory
mediators. When inflammation occurs, MAPKs such as P38 and JNK are
activated in macrophages, further activating the inflammatory
response (18). To confirm if the
expression of MAPKs is involved in the inflammatory response, the
present study performed western blot analysis. Comparing the
LPS-only group with the untreated control group, the expression of
p-ERK was not significantly changed, but the expression levels of
p-JNK and p-P38 were increased (Fig.
5). When assessing DPL extract treatment, compared with the
group treated with LPS-only, the expression levels of p-ERK, p-JNK
and p-P38 were significantly downregulated in the 400 µg/ml
DPL-treated group; the levels of p-JNK and p-P38 were also
significantly downregulated in the 200 µg/ml DPL-treated group.
Therefore, p-JNK and p-P38, in particular, exhibited more
significant decreases in expression than p-ERK.
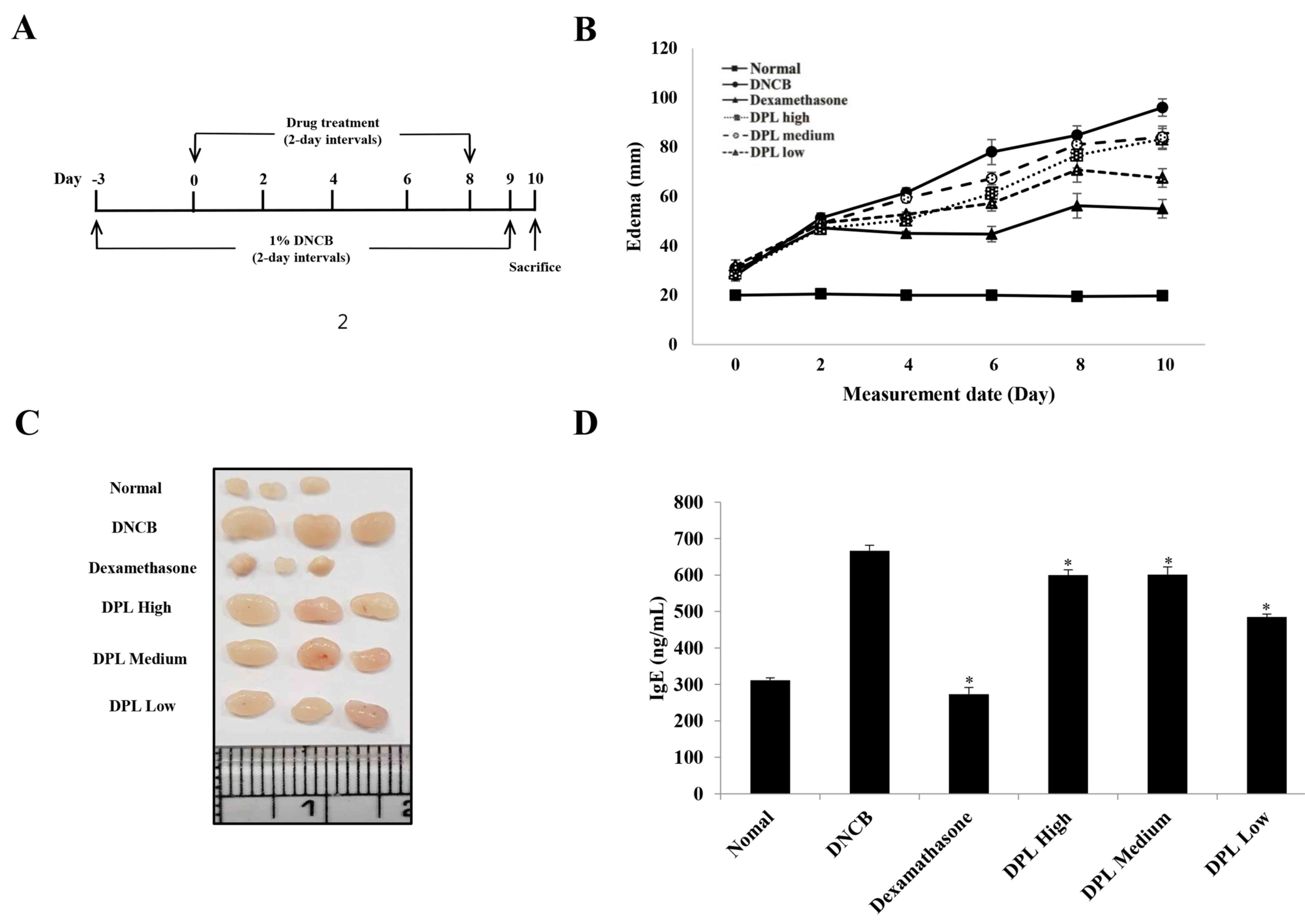
Effect of DPL extract on DNCB-induced
AD-like skin lesions in BALB/c mice
DNCB is a typical compound with a benzene ring that
triggers a local immune response to induce inflammatory dermatitis
(25,29,30).
Therefore, to determine the effect of DPL extract on induced AD,
the present study measured ear thickness in mice following
treatment with DNCB and DPL extract. The DPL extract was applied to
the ears at three concentrations: High (undiluted solution), medium
(1:2 dilution), or low (1:4 dilution). A schematic of the
experimental procedure is presented in Fig. 6A. The ear thickness of the
DPL-treated group was decreased when compared with that of the
DNCB-induced AD group (Fig. 6B).
In addition, the AD symptoms were more effectively reduced in the
DPL low group than in the DPL high and medium groups. Next, the
present study measured the morphological changes in the lymph nodes
of induced AD mice. The results revealed that the lymph nodes were
increased in the DNCB-treated group when compared with the
untreated control group, whereas, the DPL-treated groups exhibited
a decreased size compared with DNCB, particularly in the DPL low
group (Fig. 6C). In the induced AD
mice, IgE levels were significantly decreased in the DPL-treated
groups when compared with the DNCB-treated group (Fig. 6D). These results confirmed that DPL
extracts attenuated the symptoms of AD in mice.
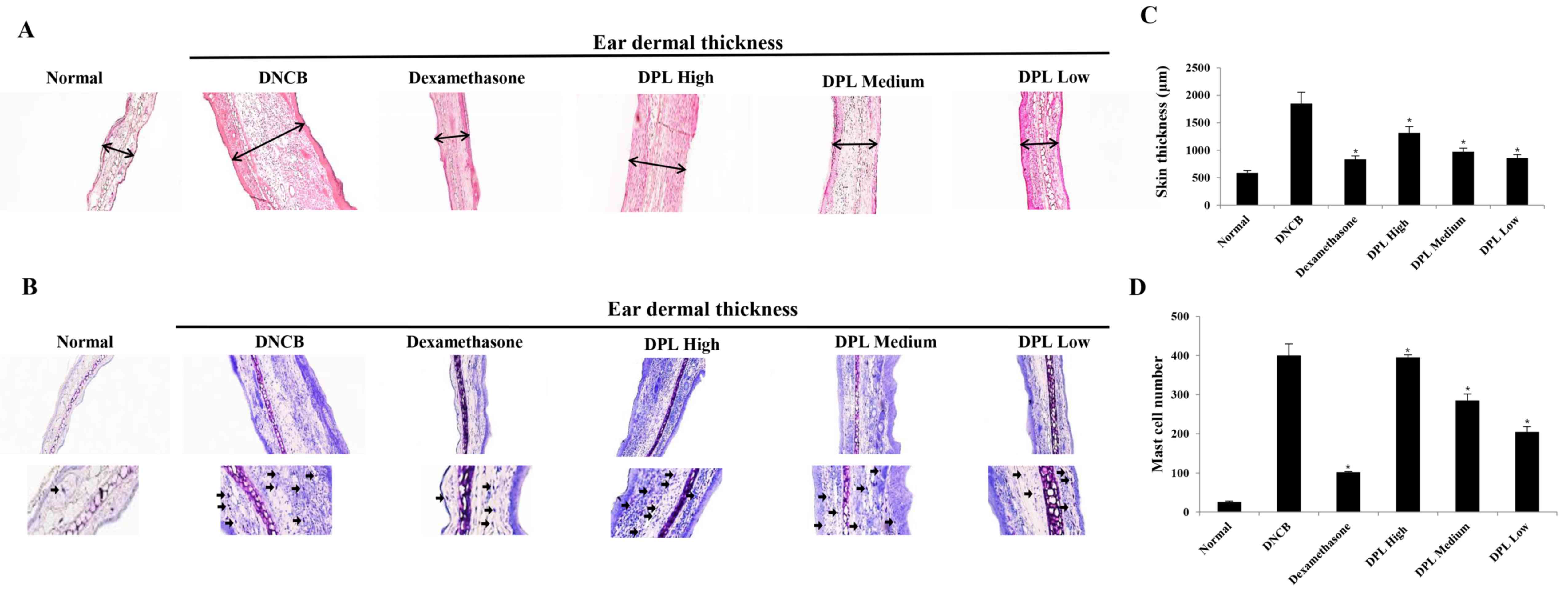
Effects of DPL extract on DNCB-induced
immune cell infiltration in BALB/c mice
When AD is induced, the secretion of vasodilators
such as proteases and histamine by mast cells increase (31). The ears of the induced AD mice were
excised to examine the thickness of the ear tissue and mast cell
secretion. Firstly, the morphological changes in the ear tissues
were evaluated by H&E staining. The ear dermal thickness of
induced AD mice were increased, whereas the ear dermal thickness
was significantly decreased in the DPL-treated group (Fig. 7A and C). Additionally, the present
study measured mast cell secretion by TB staining, which revealed
that mast cell secretions were increased in AD mice, whereas mast
cell secretions were significantly decreased in the DPL-treated
group (Fig. 7B and D). These
results indicate that DPL reduces inflammation and mast cell
secretions, and suppresses AD symptoms.
Discussion
AD is an inflammatory skin disease that causes
symptoms such as severe itching, chronic edema and skin rash. The
pathogenesis of this disease has not been elucidated to date, but
immunological abnormalities, and environmental and genetic factors
are known to be involved (3).
Contact with atopic allergens alters the balance of inflammatory
cytokines and stimulates B-cells to produce IgE, which mediates the
mast cell secretion of histamine in the skin, causing edema, itchy
skin, inflammatory reactions and the release of inflammatory
mediators such as NF-κB, NO and inflammatory cytokines (4). Immunosuppressive agents such as
steroids or antihistamines are used to treat inflammatory skin
diseases, but owing to the induced side effects, these agents are
inadequate for long-term treatment. In this regard, studies are
being conducted to identify potentially safe and effective natural
products with no side effects (5,6).
Among such natural sources, D. morbifera,
which means ‘omnipotent drug (Panax)’, has been called a panacea,
and has been the subject of various studies investigating medicinal
plants (20–24). As a result, 32 different substances
have been identified in D. morbifera. Extracts from D.
morbifera leaves are known to activate B- and T-cells (20,23),
inhibit the synthesis of melanin, have antioxidant properties, and
protect and whiten skin (24).
Although D. morbifera has exhibited potential
anti-inflammatory effects, few studies have been conducted to
identify a clear mechanism relevant to AD. The present study
investigated the effects of DPL extract on the production of NO,
cytokine secretion and inflammation-associated protein expression
in activated RAW264.7 macrophages by inducing inflammation with
LPS, and investigated its anti-inflammatory effects in DNCB-induced
AD.
Firstly, the toxicity of DPL was evaluated in
RAW264.7 macrophages by MTT assay. No cytotoxicity was observed at
a concentration of 500 µg/ml. This has already been reported
(32), and further experiments
were conducted at concentrations of 400 µg/ml or less.
NO, a known indicator of the inflammatory response,
is synthesized by NOS from L-arginine. There are three types of
NOS: Endothelial, neuronal, and iNOS, and among these, NO produced
by iNOS serves an important pathological role (13). In the present study, the NO
production in LPS-induced RAW264.7 cells was significantly reduced
by treatment with different concentrations of DPL. A previous study
also demonstrated that NO expression was inhibited in extracts of
D. morbifera stems (25).
These results suggest that DPL extract inhibited the LPS-induced
expression of NO in RAW264.7 cells.
iNOS and COX-2 serve important roles in the
production of NO, and the regulation of iNOS and COX-2 expression
is a known strategy for alleviating inflammatory diseases (14,15).
In the present study, western blotting was performed to examine the
expression of these proteins. The expression levels of iNOS and
COX-2 were significantly increased in LPS-stimulated RAW264.7 cells
when compared with untreated cells, whereas in the group treated
with 200 and 400 µg/ml DPL the expression levels of iNOS and COX-2
were decreased. In particular, the expression of iNOS was decreased
in a concentration-dependent manner. A previous study also reported
significant inhibition of COX-2 and iNOS expression, consistent
with the results obtained in the present study (33). These results suggest that DPL
extracts may regulate the expression of the inflammatory mediators
iNOS and COX-2, and regulate the expression of iNOS, thereby
suppressing NO production and alleviating the inflammatory
response.
The inflammatory cytokines TNF-α and IL-6 serve
important roles in the inflammatory process, causing inflammation
through trauma and stress, and regulating the expression of
inflammatory mediators (34). In
the present study, the expression of TNF-α and IL-6 was measured,
revealing that TNF-α expression was significantly decreased with
high concentrations of DPL, and the expression of IL-6 was
decreased in a concentration-dependent manner when compared with
RAW264.7 cells stimulated with LPS and DPL extract. Previous
studies have also reported significantly reduced IL-6 expression
following treatment with different concentrations of D.
morbifera extract (20,25).
These results indicate that DPL extract may inhibit the
inflammatory response by regulating the expression of IL-6 in
RAW264.7 cells stimulated with LPS.
TNF-α and IL-6 activate inflammation by activating
NF-κB attached to IκB-α (11).
Western blotting was performed to examine the activation of NF-κB
and IκB-α. The induction of p-NF-κB appeared to be increased
following treatment with LPS; by contrast, the induction of p-NF-κB
was lower in the groups treated with 200 and 400 µg/ml DPL than in
the group stimulated with LPS. Yu et al (34) demonstrated that olefolioside A, a
component of the D. morbifera extract, inhibited the
activation of NF-κB and IκB-α. These results suggest that DPL
inhibited the activation of NF-κB, which induced an inflammatory
response in RAW264.7 macrophages stimulated with LPS.
MAPK, a typical signaling molecule that affects the
activation of NF-κB, is phosphorylated and activated in cells, and
influences various inflammatory mediators. When inflammatory
responses are induced, MAPK activators such as P38 and JNK are
activated in macrophages (18). In
the present study, RAW264.7 cells stimulated with LPS were treated
with different concentrations of DPL extract and the activation of
ERK, P38, and JNK was assessed. The expression of p-ERK, p-P38 and
p-JNK was significantly decreased by 400 µg/ml DPL treatment. A
previous study also revealed that D. morbifera extract
modulates the expression of MAPK (34). These results suggest that DPL
extract may modulate the signaling pathway of MAPK and inhibit the
inflammatory response in LPS-stimulated macrophages.
BALB/c mice were treated with DNCB, which is known
to induce inflammatory dermatitis, to induce AD. DPL extract was
then applied to the ear and the thickness of the ear was assessed.
Ear thickness was significantly lower in the DPL-treated group than
in the group treated with DNCB. The ears in the DPL low-dose group
exhibited a larger decrease in thickness than the medium- and
high-dose groups. This suggested that the higher the concentration
of DPL extract (the closer to undiluted solution), the higher the
viscosity, which decreased its penetration; thus, groups treated
with lower concentrations of DPL exhibited higher effects owing to
the low viscosity. DPL was less effective than dexamethasone, which
was used as a positive control. However, the dexamethasone groups
observed cutaneous atrophy and erythema. This indicated that DPL
may have potential for therapeutic use with fewer side effects
(cutaneous atrophy and erythema) than dexamethasone with repeated
use. There were significantly fewer morphological changes in the
lymph nodes of the DPL-treated group than in the DNCB-treated
group, and IgE was significantly decreased in the DPL-treated
group. Additionally, morphological changes in the epidermis and
corium of ear tissues, and the secretion of immune-activated mast
cells were measured by H&E and TB staining. The ears of
DNCB-treated mice were thicker than those of DPL-treated mice, and
the mast cell distribution was significantly decreased in the
DPL-treated group. Lee et al (35) reported that Dendropanax
exhibited anti-inflammatory effects in pneumonia, and Jung et
al (36) confirmed that
Dendropanax relieves nephritis. These results suggest that
DPL extract alleviated DNCB-induced atopic skin disease.
In conclusion, the present study investigated the
anti-inflammatory effects of DPL extract. It was observed that
D. morbifera regulated the expression of NO and IL-6, and
the signaling pathway of NF-κB and MAPK in LPS-stimulated RAW264.7
macrophages in vitro. The present study also confirmed that
DNCB-induced inflammatory dermatitis was mitigated by DPL in
vivo, without side effects. These results suggest that DPL
extract merits future research and development as a natural
anti-inflammatory or functional drug. In addition, the
investigation of the components of D. morbifera will be
required in the future as well as studying the mechanisms
underlying its anti-inflammatory effects.
Acknowledgements
Not applicable.
Funding
The present study was supported by a research grant
from MBG group (Daejeon, Republic of Korea).
Availability of data and materials
Not applicable.
Authors' contributions
GSC, DPL, SMK, CHK and JYJ conceived and designed
the experiments. GSC, ESY, SHK and JSW performed the experiments.
GSC, DPL, ESY and JYJ analyzed the data. DPL, CHK, HJK and SMK
contributed in analyzing the data and provided reagents and
materials. GS wrote the manuscript.
Ethics approval and consent to
participate
All animal experiments were performed with the
approval of the Institutional Animal Care and Use Committee of
Kong-Ju National University (approval no. KNU_2017-10; Yesan,
Korea) and the institutional guidelines were adhered to.
Patient consent for publication
Not applicable.
Competing interests
The authors Dong-pyo Lim, Sae-man Kim and Chang-hyun
Kim are all affiliated with MBG group (Daejeon, Republic of Korea);
this company also provided financial support for the present study.
The authors declare that they have no competing interests.
References
|
1
|
Urabe K, Aroca P, Tsukamoto K, Mascagna D,
Paulumbo A, Prota G and Hearing VJ: The inherent cytotoxicity of
melanin precursors: A revision. Biochim Biophys Acta 1221. 272–278.
1994.
|
|
2
|
Voss GT, Oliveira RL, de Souza JF, Duarte
LFB, Fajardo AR, Alves D, Luchese C and Wilhelm EA: Therapeutic and
technological potential of 7-chloro-4-phenylselanyl quinoline for
the treatment of atopic dermatitis-like skin lesions in mice. Mater
Sci Eng C Mater Biol Appl. 84:90–98. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hwang SW, Kang JH, Seol JE, Seo JK, Lee
DBR and Sung HS: The correlation between SCORAD index and
instrumental assessment in evaluation of atopic dermatitis
severity. Korean J Dermatol. 48:266–271. 2010.(In Korean).
|
|
4
|
Leung DY: Pathogenesis of atopic
dermatitis. J Allergy Clin Immun. 104:S99–S108. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leung V, Hartwell R, Yang H, Ghahary A and
Ko F: Bioactive nanofibres for wound healing applications. Journal
of Fiber Bioengineering and Informatics. 4:1–14. 2011. View Article : Google Scholar
|
|
6
|
Guglielmetti S, Dart JK and Calder V:
Atopic keratoconjunctivitis and atopic dermatitis. Curr Opin
Allergy Clin Immunol. 10:478–485. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim BJ, Son WR, Choi MO, Jo SK, Jung HK,
Lee JT, Kim HY and Kwoen DJ: Anti-atopic effects of Castanea
crenata inner shell extracts fermented by Lactobacillus
bifermentans. J Korean Soc Food Sci Nutr. 42:1378–1386. 2013.
View Article : Google Scholar
|
|
8
|
Kang BK, Kim KBWR, Kim MJ, Bark SW, Pak
WM, Kim BR, Ahn NK, Choi YU, Bae NY, Park JH, et al: Anti-atopic
activity of tuna heart ethanol extract. J Korean Soc Food Sci Nutr.
44:1–6. 2015. View Article : Google Scholar
|
|
9
|
Ferero-Miliani L, Nielsen OH, Andersen PS
and Girardin SE: Chronic inflammation: Importance of NOD2 and NALP3
in interleukin-1beta generation. Clin Exp Immunol. 147:227–235.
2007.PubMed/NCBI
|
|
10
|
Zamora R, Vodovotz Y and Billiar TR:
Inducible nitric oxide synthase and inflammatory diseases. Mol Med.
6:347–373. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bierhaus A, Schiekofer S, Schwaninger M,
Andrassy M, Humpert PM, Chen J, Hong M, Luther T, Henle T, Klöting
I, et al: Diabetes-associated sustained activation of the
transcription factor nuclear factor-kappaB. Diabetes. 50:2792–2808.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hattori Y, Hattori S, Sato N and Kasai K:
High-glucose-induced nuclear factor kappaB activation in vascular
smooth muscle cells. Cardiovasc Res. 46:188–197. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yun HY, Dawson VL and Dawson TM:
Neurobiology of nitric oxide. Crit Rev Neurobiol. 10:291–316. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nathan C: Nitric oxide as a secretory
product of mammalian cells. FASEB J. 6:3051–3064. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Seibert K, Zhang Y, Leahy K, Hauser S,
Masferrer J, Perkins W, Lee L and Isakson P: Pharmacological and
biochemical demonstration of the role of cyclooxygenase 2 in
inflammation and pain. Proc Natl Acad Sci USA. 91:12013–12017.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hippeli S and Elastner EF: Inhibition of
biochemical model reactions for inflammatory processes by plant
extracts: A review on recent developments. Free Radic Res. 31
Suppl:S81–S87. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yoon YI, Chung MY, Hwang JS, Goo TW, Ahn
MY, Lee YB, Han MS and Yun EY: Anti-inflammatory effect of Oxya
chinensis sinuosa ethanol extract in LPS-induced RAW 264.7 cells. J
Life Sci. 24:370–376. 2014. View Article : Google Scholar
|
|
18
|
Cobb MH: MAP kinase pathways. Prog Biophys
Mol Biol. 71:479–500. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee MK, Lee IS and Lee JS: For the
utilization of native plant resources as high-value materials:
Evaluation of demelanizing activity of Dendropanax morbifera in
Bogildo. J Korean Island. 25:227–240. 2013.(In Korean).
|
|
20
|
Lee SH, Lee HS, Park YS, Hwang B, Kim JH
and Lee HY: Screening of immune activation activities in the leaves
of Dendropanax morbifera Lev. Korean J Medicinal Crop Sci.
10:109–115. 2002.(In Korean).
|
|
21
|
Castro Aceituno V, Ahn S, Simu SY, Wang C,
Mathiyalagan R and Yang DC: Silver nanoparticles from Dendropanax
morbifera Léveille inhibit cell migration, induce apoptosis, and
increase generation of reactive oxygen species in A549 lung cancer
cells. In Vitro Cell Dev Biol Anim. 52:1012–1019. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Choi SK: Growth characteristics of
Dendropanax morbifera LEV. in Wando area of Korea. Korean J Crop
Sci. 48:434–437. 2003.(In Korean).
|
|
23
|
Park BY, Min BS, Oh SR, Kim JH, Kim TJ,
Kim DH, Bae KH and Lee HK: Isolation and anticomplement activity of
compounds from Dendropanax morbifera. J Ethnopharmacol. 90:403–408.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park SA, Park J, Park CI, Jie YJ, Hwang
YC, Kim YH, Jeon SH, Lee HM, Ha JH, Kim KJ and Park SN: Cellular
antioxidant activity and whitening effects of Dendropanax morbifera
leaf extracts. Korean J Microbiol Biotechnol. 41:407–415. 2013.
View Article : Google Scholar
|
|
25
|
Im KJ, Jang SB and Yoo DY: Anti-cancer
effects of Dendropanax Morbifera extract in MCF-7 and MDA-MB-231
cells. J Korean Obstet Gynecol. 28:26–39. 2015. View Article : Google Scholar
|
|
26
|
Lee HN, Shin SA, Choo GS, Kim HJ, Park YS,
Kim BS, Kim SK, Cho SD, Nam JS, Choi CS, et al: Anti-inflammatory
effect of quercetin and galangin in LPS-stimulated RAW264.7
macrophages and DNCB-induced atopic dermatitis animal models. Int J
Mol Med. 41:888–898. 2018.PubMed/NCBI
|
|
27
|
Liu YN, Zha WJ, Ma Y, Chen FF, Zhu W, Ge
A, Zeng XN and Huang M: Galangin attenuates airway remodelling by
inhibiting TGF-β1-mediated ROS generation and MAPK/Akt
phosphorylation in asthma. Sci Rep. 5:117582015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jeong DH, Kim KB, Kim MJ, Kang BK and Ahn
DH: Anti-inflammatory activity of methanol extract and n-hexane
fraction mojabanchromanol b from Myagropsis myagroides. Life Sci.
114:12–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim BA, Yang JC and Park CI: Effect of
Hwangryunhaedok-tang extracts on DNCB-induced allergic contact
dermatitis. Kor J Herbology. 24:1–5. 2009.(In Korean).
|
|
30
|
Park SO, Park BS, Ryu CM and Ahn YS:
Effect of herb extracts mixed with Houttuynia cordata on antiatopic
dermatitis in DNCB-induced BALB/c mouse. J Kor Oil Chemists Soc.
2:175–183. 2012.(In Korean).
|
|
31
|
Lim SJ, Kim M, Randy A, Nam EJ and Nho CW:
Effects of Hovenia dulcis Thunb. extract and methyl vanillate on
atopic dermatitis-like skin lesions and TNF-α/IFN-γ-induced
chemokines production in HaCaT cells. J Pharm Pharmacol.
68:1465–1479. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park SY, Karthivashan G, Ko HM, Cho DY,
Kim J, Cho DJ, Ganesan P, Su-Kim I and Choi DK: Aqueous extract of
Dendropanax morbiferus leaves effectively alleviated
neuroinflammation and behavioral impediments in MPTP-induced
parkinson's mouse model. Oxid Med Cell Longev 2018.
31752142018.
|
|
33
|
Hyun TK, Ko YJ, Kim EH, Chung IM and Kim
JS: Anti-inflammatory activity and phenolic composition of
Dendropanax morbifera leaf extracts. Ind Crop Pro. 74:263–270.
2015. View Article : Google Scholar
|
|
34
|
Yu HY, Kim KS, Lee YC, Moon HI and Lee JH:
Oleifolioside A, a new active compound, attenuates LPS-Stimulated
iNOS and COX-2 expression through the downregulation of NF-κB and
MAPK activities in RAW 264.7 macrophages. Evid Based Complement
Alternat Med 2012. 6375122012.
|
|
35
|
Lee JW, Ryu HW, Lee SU, Son TH, Park HA,
Kim MO, Yuk HJ, Ahn KS and Oh SR: Protective effect of
polyacetylene from Dendropanax morbifera Leveille leaves on
pulmonary inflammation induced by cigarette smoke and
lipopolysaccharide. J Fun Foods. 32:358–366. 2017. View Article : Google Scholar
|
|
36
|
Jung HY, Kwon HJ, Hahn KR, Yoo DY, Kim W,
Kim JW, Kim YJ, Yoon YS, Kim DW and Hwang IK: Dendropanax morbifera
Léveille extract ameliorates cesium-induced inflammation in the
kidney and decreases antioxidant enzyme levels in the hippocampus.
Mol Cel Toxicol. 14:193–199. 2018. View Article : Google Scholar
|