Introduction
Arrhythmogenic right ventricular cardiomyopathy
(ARVC) is an autosomal dominant inherited disease. The
distinguishing feature of ARVC is the replacement of the myocardium
with fibrous fatty tissue, which leads to ventricular tachycardia
(VT) and ultimately results in sudden cardiac death (SCD) (1). As reported in previous studies, the
majority of patients with ARVC are male (2–5). In
addition, the sex disparity in phenotypic presentations of ARVC has
been demonstrated by numerous studies. Naneix et al
(6) indicated that the cause of
SCD was more often the cardiomyopathy in women, particularly
arrythmogenic right ventricular cardiomyopathy (ARVC). In
life-threatening arrhythmias, numerous studies have demonstrated
that the male sex is associated with a significantly higher risk of
VT (7–9). In order to provide individualized
therapy for patients with ARVC, the present study aimed to identify
key genes and pathways that may be associated with the diversity of
phenotypes between female and male patients.
In the present study, the GSE29819 gene expression
dataset was downloaded from the Gene Expression Omnibus (GEO)
database. Differentially expressed genes (DEGs) were defined as
genes with significantly different expression levels between female
and male patients with ARVC. The biological functions of the DEGs
were identified using Gene Ontology (GO) and Kyoto Encyclopedia of
Genes and Genomes (KEGG) analyses (10,11).
The protein-protein interaction (PPI) network was constructed using
the Search Tool for the Retrieval of Interacting Genes (STRING)
database, in order to detect the hub genes and modules (12). The results of the present
bioinformatics analysis may aid elucidation of the sex differences
of ARVC by identifying the key genes and pathways that contribute
to the phenotypes.
Materials and methods
Microarray data
The GSE29819 gene expression dataset (13) is based on the gene chip of the
Human Genome U133 Plus 2.0 Array platform (Affymetrix; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and was retrieved from
the GEO database (http://www.ncbi.nlm.nih.gov/geo/). The GSE29819
dataset contains 38 ventricular myocardial samples, including 12
tissues from patients with ARVC, 14 tissues from patients with
dilated cardiomyopathy and 12 tissues from individuals with
non-failing hearts. The 12 tissues from patients with ARVC were
divided into two groups according to sex: The female patients group
(4 samples) and the male patients group (8 samples). The
identification of DEGs was conducted using GEO2R (http://www.ncbi.nlm.nih.gov/geo/geo2r/),
which is an online analytical tool for performing comparisons on
raw data with the limma R package and GEO-query (14). The results of the comparisons were
extracted in file format, and the cut-off criteria for DEGs were
set as follows: P<0.05 and |log fold-change|>1.5.
GO and KEGG enrichment analyses
GO and KEGG enrichment analyses of the upregulated
and downregulated DEGs were conducted separately with the Cytoscape
plugin ClueGO (version 2.5.2) (15). The criteria for statistically
significant differences were set as follows: P<0.01 and a κ
score of 1.0.
Integration of the PPI network
Interactions among the DEGs were evaluated using the
STRING (version 10.5) database; a combined score of >0.4 was
considered to indicate a statistically significant interaction
(16). The integrated networks
were visualized with Cytoscape software (version 3.6.1) (17). In addition, the Cytoscape plugin
Molecular Complex Detection (MCODE; version 1.31), with a cutoff
criterion of MCODE score >5, was used to identify molecular
modules in the PPI network (18).
Further functional annotation of the modules was achieved with
STRING (version 10.5) (16). The
Cytoscape plugin cytoHubba (version 0.1), with the ranking methods
of maximal clique centrality (MCC), was used to identify the hub
genes (19).
Results
Identification of DEGs in patients
with ARVC of different sexes
The datasets were analyzed using GEO2R and 1,188
DEGs were identified, of which 915 were upregulated and 273 were
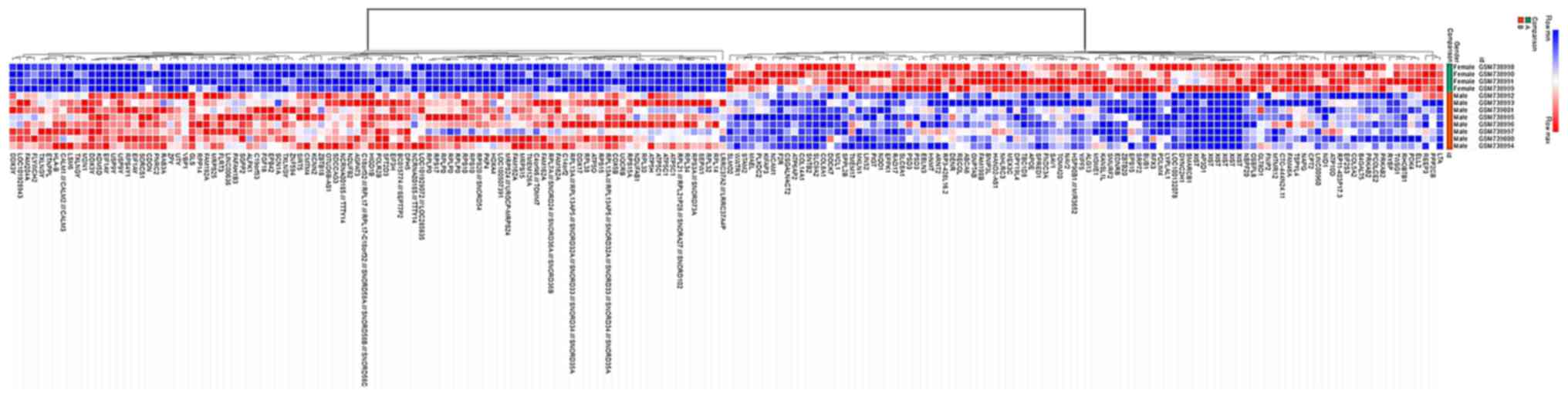
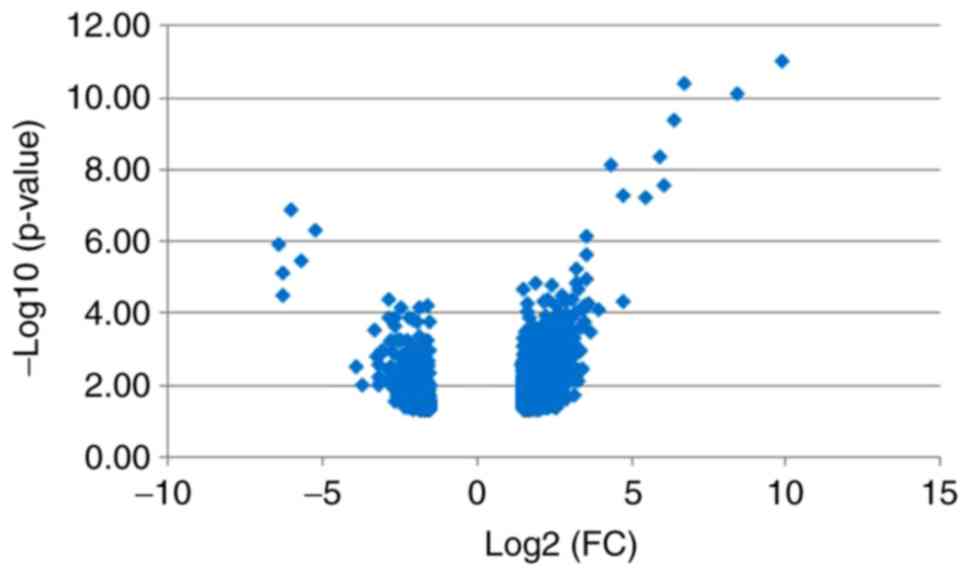
downregulated in the male group. The DEGs were used to construct a
heat map (Fig. 1) and a volcano
plot (Fig. 2).
Functional enrichment analysis
The upregulated and downregulated DEGs were
processed separately with ClueGO for GO and KEGG pathway analyses.
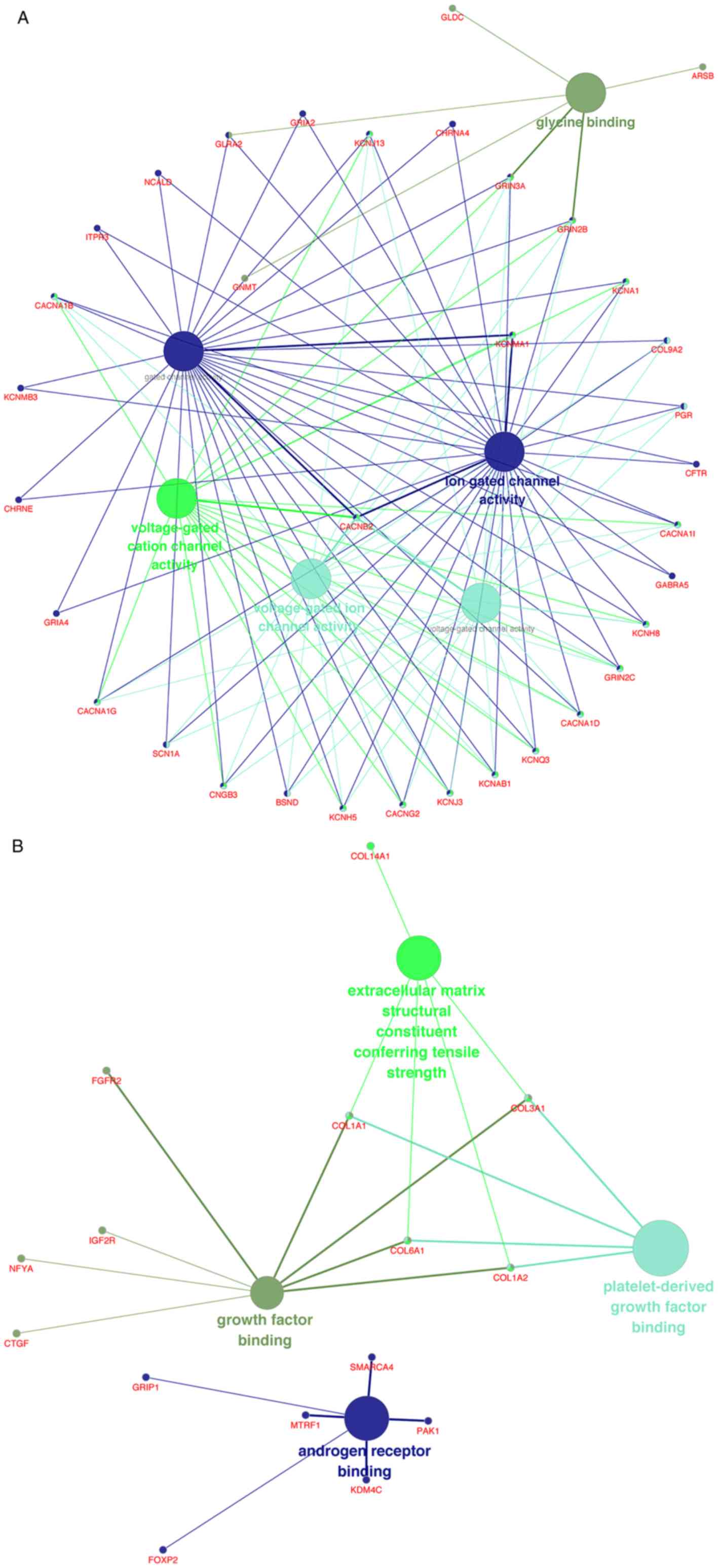
In terms of molecular function (MF), the upregulated DEGs were
mostly enriched in ‘glycine binding’, ‘voltage-gated cation channel
activity’, ‘voltage-gated ion channel activity’, ‘voltage-gated
channel activity’, ‘gated channel activity’ and ‘ion gated channel
activity’ (Fig. 3A), whereas the
downregulated DEGs were mainly enriched in ‘growth factor binding’,
‘extracellular matrix structural constituent conferring tensile
strength’, ‘platelet-derived growth factor binding’ and ‘androgen
receptor binding’ (Fig. 3B).
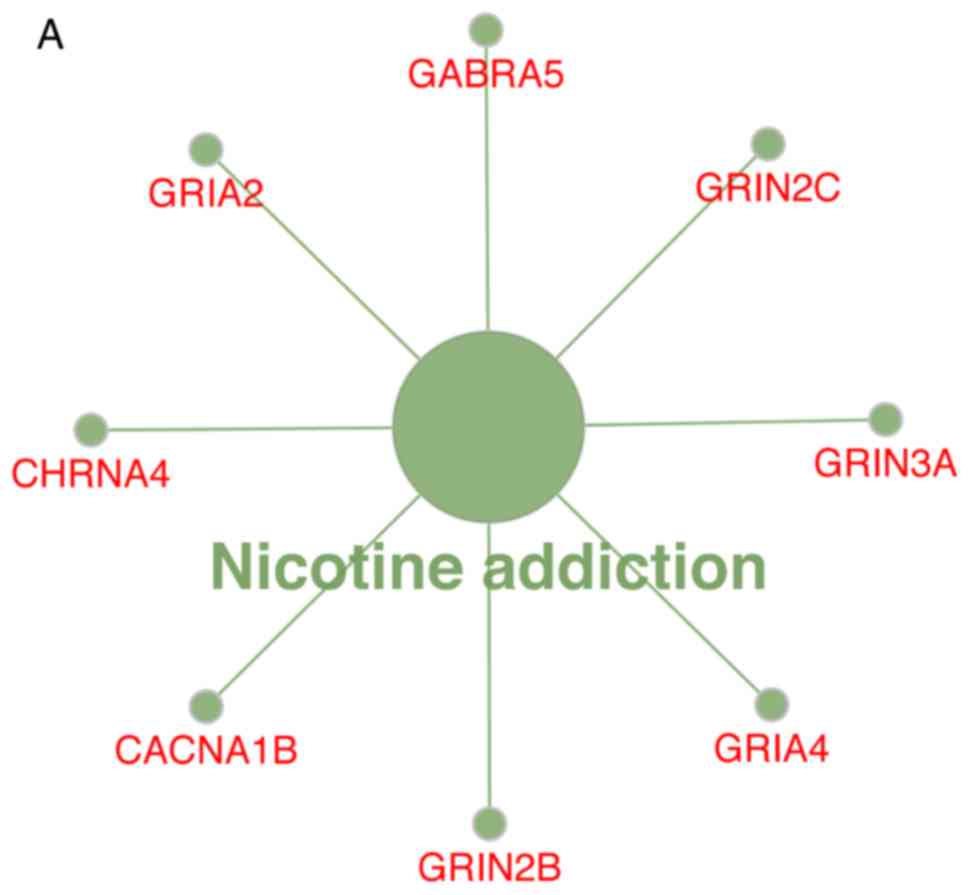
The KEGG pathway analysis revealed that the
upregulated DEGs were substantially enriched in the ‘nicotine’
pathways (Fig. 4A), whereas the
downregulated DEGs were substantially enriched in the ‘ECM-receptor
interaction’ and ‘protein digestion and absorption’ pathways
(Fig. 4B).
PPI network construction, and module
and hub gene identification
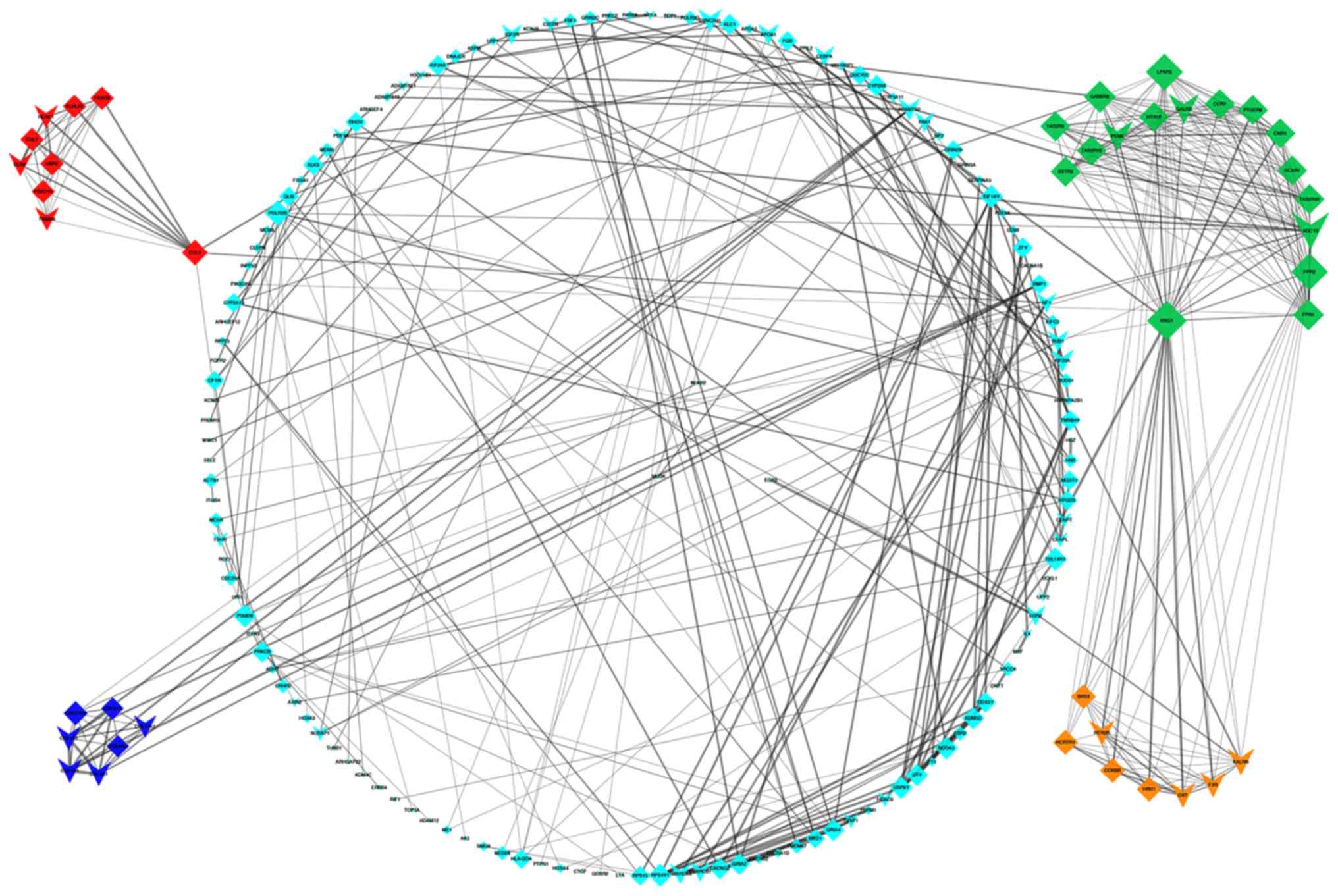
Detection of interactions among the DEGs was
performed by constructing a PPI network, which contained 899 nodes
and 1,627 edges (Fig. 5). With a
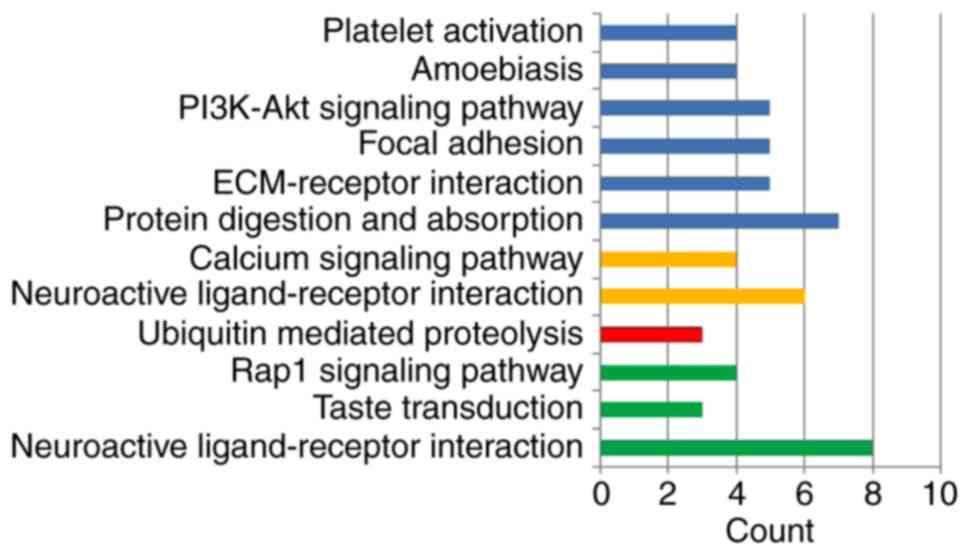
cutoff criterion of MCODE score >5, four modules were
identified, which were significantly enriched in the following
pathways: DEGs in module 1, including LPAR5, FPR2, ADCY2, GABBR2,
FPR1, HCAR1, PTGER3 and CNR1, were enriched in ‘neuroactive
ligand-receptor interaction’, ‘taste transduction’ and ‘Rap1
signaling pathway’; DEGs in module 2, including KLHL25, CUL3,
HERC1, CUL7, CCNF, UBR2, FBXO9, FBXO10 and TRIM36, were enriched in
‘ubiquitin mediated proteolysis’; DEGs in module 3, including OXT,
HRH1, F2R, KALRN, CCKBR, HTR2B, HCRTR1 and BRS3, were enriched in
‘neuroactive ligand-receptor interaction’ and ‘calcium signaling
pathway’; and DEGs in module 4, including COL1A2, COL1A1, COL3A1,
COL6A1, COL14A1, COL27A1 and COL9A2, were enriched in ‘protein
digestion and absorption’, ‘ECM-receptor interaction’, ‘focal
adhesion’, ‘PI3K-Akt signaling pathway’, ‘amoebiasis’ and ‘platelet
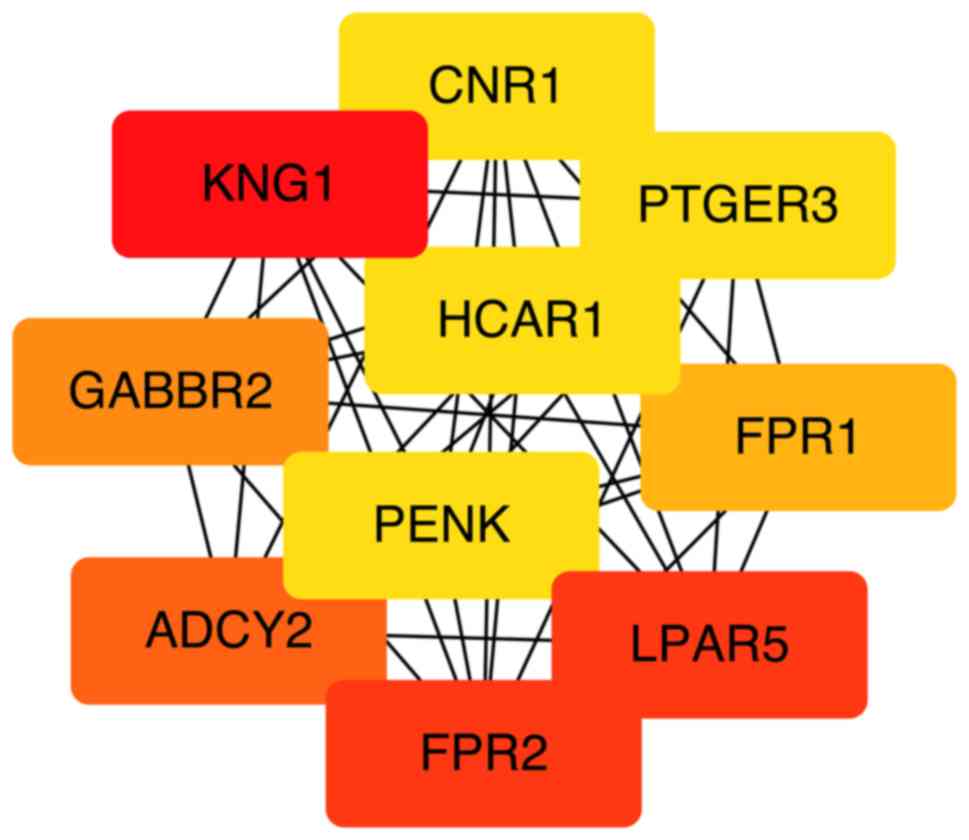
activation’ (Fig. 6). Based on the
ranking methods of MCC, the following genes were identified as the
top 10 hub genes (Fig. 7):
Kininogen 1 (KNG1), lysophosphatidic acid receptor 5 (LPAR5),
formyl peptide receptor (FPR) 2, adenylate cyclase 2 (ADCY2),
γ-aminobutyric acid type B receptor subunit 2 (GABBR2), FPR1,
hydroxycarboxylic acid receptor 1 (HCAR1), prostaglandin E receptor
3 (PTGER3), cannabinoid receptor 1 (CNR1) and proenkephalin
(PENK).
Discussion
Numerous studies have focused on the sex differences
in the phenotypes of ARVC and have revealed substantial diversity
in clinical presentations, including SCD or VT, between female and
male patients (6–9). The aim of the present study was to
aid identification of the cause of these differences using
bioinformatics analysis.
ARVC is an inherited disease with 12 confirmed
related genes. Notably, none of these 12 genes exhibited
statistical sex differences in the present study, indicating the
possible involvement of other symptom-associated genes. In the
present study, the genetic profile data were extracted from the GEO
database and analyzed with GEO2R. The analysis revealed that 1,188
genes were differentially expressed between the female and male
patients with ARVC, of which 273 were downregulated and 915 were
upregulated.
A previous study identified a splice site homozygous
deletion in solute carrier family 6 member 6 (SLC6A6) in patients
with idiopathic dilated cardiomyopathy, indicating the possible
role of SLC6A6 in the pathophysiology of heart failure (HF)
(20). Natriuretic peptide (NPP) A
and NPPB were two significantly downregulated DEGs in the present
study. The NPPB promoter regulates the dynamic expression of NPPA,
and NPPA has previously been demonstrated to be highly expressed
during ventricular stress (21).
The disparity detected in NPPA expression suggests markedly
different levels of ventricular stress between the groups, which
may leads to diverse phenotypes. Ubiquitin specific peptidase 9
Y-linked (USP9Y), ribosomal protein S4 Y-linked 1 (RPS4Y1) and
eukaryotic translation initiation factor 1A Y-linked (EIF1AY) were
significantly upregulated DEGs in the present study. Similar
patterns of expression were demonstrated by Heidecker et al
(22), thus suggesting that
overexpression of USP9Y, RPS4Y1 and EIF1AY may contribute to sex
differences in the pathophysiology of HF.
The GO analysis revealed that, in the molecular
function domain, the downregulated DEGs were significantly enriched
in extracellular matrix (ECM) structural constituent. A previous
study demonstrated that proteolysis-mediated signaling can enhance
accumulation of the ECM, which contributes to negative remodeling
of the left ventricle and may eventually lead to severe HF
(23). This may be a possible
mechanism in certain patients that present with severe HF. In the
MF domain, the majority of DEGs in the present study were enriched
in ‘glycine binding’, ‘voltage-gated cation channel activity’,
‘voltage-gated ion channel activity’, ‘growth factor binding’,
‘extracellular matrix structural constituent conferring tensile
strength’, ‘platelet-derived growth factor binding’ and ‘androgen
receptor binding’. A number of these results are consistent with
those of previous studies. Qi et al (24) suggested that glycine protects
cardiomyocytes from lipopolysaccharide and hypoxia/reoxygenation
injury by blocking the influx of calcium. Numerous studies have
demonstrated that ‘voltage-gated channel’ markedly contributes to
VT (25–27). Therefore, dysregulation in these
DEGs may cause different phenotypes between the sexes.
Using MCODE, four modules were identified that were
enriched in the following pathways: ‘neuroactive ligand-receptor
interaction’, ‘taste transduction’, ‘Rap1 signaling pathway’,
‘ubiquitin mediated proteolysis’, ‘calcium signaling pathway’,
‘protein digestion and absorption’, ‘ECM-receptor interaction’,
‘focal adhesion’, ‘PI3K-Akt signaling pathway’, ‘amoebiasis’ and
‘platelet activation’. These pathways may have served a role in the
diversity of clinical presentations between the sexes. Using the
KEGG database (28), the majority
of pathways enriched in modules 1 and 3 were mapped in
‘environmental information processing’, including ‘signaling
molecules and interaction’ and ‘Signal transduction’. The DEGs in
module 1, including LPAR5, FPR2, ADCY2, GABBR2, FPR1, HCAR1, PTGER3
and CNR1, were involved in the ‘signaling molecules and
interaction’ and ‘signal transduction’ pathways, suggesting that
these hubs contribute to the sex differences of phenotypes via
‘environmental information processing’. Pathway of ‘ubiquitin
mediated proteolysis’ was enriched in module 2, indicating the
possible involvement of the well-known protein degradation systems
‘ubiquitin proteasome’ in the development of ARVC (29). In addition to ‘environmental
information processing’, the pathways of ‘amoebiasis’, ‘platelet
activation’ and ‘protein digestion and absorption’ were
significantly enriched in module 4. Since both the ‘amoebiasis’ and
‘platelet activation’ pathways participate in the activity of the
immune system, it may be hypothesized that the DEGs (collagen type
I α1 chain, collagen type I α2 chain, collagen type XXVII α1 chain
and collagen type III α1 chain) in module 4 function in the various
clinical presentations of ARVC via the immune mechanism (30,31).
The following 10 hub genes with a high level of
connectivity were identified using cytoHubba: KNG1, LPAR5, FPR2,
ADCY2, GABBR2, FPR1, HCAR1, PTGER3, CNR1 and PENK. Variations of
the KNG gene can impact the aldosterone response, which markedly
contributes to structural alterations in the heart (32,33).
FPR agonists act in favor of cardioprotection (34). Therefore upregulation of the DEGs
KNG1, FPR1 and FPR2 in female patients may reduce pathological
cardiac reconstruction and increase cardiac protection, resulting
in a lower chance of SCD. A similar deduction can be made for
LPAR5, which is expressed in H9c2 cardiomyocytes. Ectonucleotide
pyrophosphatase/phosphodiesterase 2/LPAR5 signaling regulates
ferroptosis-related genes (glutathione peroxidase 4, acyl-CoA
synthetase long chain family member 4 and nuclear factor, erythroid
2 like 2) and adjusts the survival signals of cardiomyocytes in
order to protect cardiomyocytes from ferroptosis. PENK encodes the
protein proenkephalin, which is mainly located in skeletal and
cardiac muscle (35). In a
previous study, the level of PENK was used to assess the
progression of HF (36).
Treskatsch et al (37)
demonstrated that rats with severe HF develop overexpression of
PENK. Furthermore, PENK is considered an independent risk predictor
in the prognosis of patients with HF (38). Therefore, downregulation of PENK in
female patients indicated that relatively preserved heart function
may be the reason for a lower incidence of SCD. A limitation of the
present study is that verification was not conducted, as ARVC is a
relatively rare disease and the number of available genetic data
profiles is limited.
The present study used bioinformatics analysis as a
tool to identify genes and pathways that may be responsible for the
sex differences in ARVC. The results of this study provided genetic
insights into the diversity in the phenotype and clinical
characteristics between female and male patients. However, further
studies that verify these results are required. Only a few of the
candidate DEGs identified in this study have been associated with
HF or VT in previous studies. Further studies are required to
establish the specific molecular mechanisms underlying the effects
of the DEGs and pathways involved in different clinical
presentations of ARVC between the sexes.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the Gene Expression Omnibus repository
(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE29819).
Authors' contributions
LTC analyzed and interpreted the data, and wrote the
manuscript. CYJ made substantial contributions to the conception
and study design, revised the manuscript critically for important
intellectual content and gave the final approval of the version to
be published. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ARVC
|
arrhythmogenic right ventricular
cardiomyopathy
|
|
GEO
|
Gene Expression Omnibus
|
|
DEGs
|
differentially expressed genes
|
|
PPI
|
protein-protein interaction
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
VT
|
ventricular tachycardia
|
|
SCD
|
sudden cardiac death
|
|
MF
|
molecular function
|
|
MCC
|
maximal clique centrality
|
|
ECM
|
extracellular matrix
|
|
HF
|
heart failure
|
References
|
1
|
Maupain C, Badenco N, Pousset F, Waintraub
X, Duthoit G, Chastre T, Himbert C, Hébert JL, Frank R,
Hidden-Lucet F and Gandjbakhch E: Risk stratification in
arrhythmogenic right ventricular cardiomyopathy/dysplasia without
an implantable cardioverter-defibrillator. JACC Clin
Electrophysiol. 4:757–768. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gerull B, Heuser A, Wichter T, Paul M,
Basson CT, McDermott DA, Lerman BB, Markowitz SM, Ellinor PT,
MacRae CA, et al: Mutations in the desmosomal protein plakophilin-2
are common in arrhythmogenic right ventricular cardiomyopathy. Nat
Genet. 36:1162–1164. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Beffagna G, Occhi G, Nava A, Vitiello L,
Ditadi A, Basso C, Bauce B, Carraro G, Thiene G, Towbin JA, et al:
Regulatory mutations in transforming growth factor-beta3 gene cause
arrhythmogenic right ventricular cardiomyopathy type 1. Cardiovasc
Res. 65:366–373. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pilichou K, Nava A, Basso C, Beffagna G,
Bauce B, Lorenzon A, Frigo G, Vettori A, Valente M, Towbin J, et
al: Mutations in desmoglein-2 gene are associated with
arrhythmogenic right ventricular cardiomyopathy. Circulation.
113:1171–1179. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Syrris P, Ward D, Evans A, Asimaki A,
Gandjbakhch E, Sen Chowdhry S and McKenna WJ: Arrhythmogenic right
ventricular dysplasia/cardiomyopathy associated with mutations in
the desmosomal gene desmocollin-2. Am J Hum Genet. 79:978–984.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Naneix AL, Périer MC, Beganton F, Jouven X
and Lorin de la Grandmaison G: Sudden adult death: An autopsy
series of 534 cases with gender and control comparison. J Forensic
Leg Med. 32:10–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kimura Y, Noda T, Otsuka Y, Wada M,
Nakajima I, Ishibashi K, Miyamoto K, Okamura H, Aiba T, Kamakura S,
et al: Potentially lethal ventricular arrhythmias and heart failure
in arrhythmogenic right ventricular cardiomyopathy: What are the
differences between men and women? JACC Clin Electrophysiol.
2:546–555. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mazzanti A, Ng K, Faragli A, Maragna R,
Chiodaroli E, Orphanou N, Monteforte N, Memmi M, Gambelli P,
Novelli V, et al: Arrhythmogenic right ventricular cardiomyopathy:
Clinical course and predictors of arrhythmic risk. J Am Coll
Cardiol. 68:2540–2550. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Protonotarios A, Anastasakis A,
Panagiotakos DB, Antoniades L, Syrris P, Vouliotis A, Stefanadis C,
Tsatsopoulou A, McKenna WJ and Protonotarios N: Arrhythmic risk
assessment in genotyped families with arrhythmogenic right
ventricular cardiomyopathy. Europace. 18:610–616. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gan M: Correlating information contents of
gene ontology terms to infer semantic similarity of gene products.
Comput Math Methods Med 2014. 8918422014.
|
|
11
|
Du J, Yuan Z, Ma Z, Song J, Xie X and Chen
Y: KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway
analysis using a path analysis model. Mol Biosyst. 10:2441–2447.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hasan MM and Kahveci T: Indexing a
protein-protein interaction network expedites network alignment.
BMC Bioinformaticss. 16:3262015. View Article : Google Scholar
|
|
13
|
Gaertner A, Schwientek P, Ellinghaus P,
Summer H, Golz S, Kassner A, Schulz U, Gummert J and Milting H:
Myocardial transcriptome analysis of human arrhythmogenic right
ventricular cardiomyopathy. Physiol Genomics. 44:99–109. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:(Database Issue).
D991–D995. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bindea G, Mlecnik B, Hackl H, Charoentong
P, Tosolini M, Kirilovsky A, Fridman WH, Pagès F, Trajanoski Z and
Galon J: ClueGO: A cytoscape plug-in to decipher functionally
grouped gene ontology and pathway annotation networks.
Bioinformaticss. 25:1091–1093. 2009. View Article : Google Scholar
|
|
16
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43:(Database Issue). D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformaticss. 4:22003. View Article : Google Scholar
|
|
19
|
Chin CH, Chen SH, Wu HH, Ho CW, Ko MT and
Lin CY: cytoHubba: Identifying hub objects and sub-networks from
complex interactome. BMC Syst Biol. 8 Suppl 4:S112014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shakeel M, Irfan M and Khan IA: Rare
genetic mutations in Pakistani patients with dilated
cardiomyopathy. Gene. 673:134–139. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Man J, Barnett P and Christoffels VM:
Structure and function of the Nppa-Nppb cluster locus during heart
development and disease. Cell Mol Life Sci. 75:1435–1444. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Heidecker B, Lamirault G, Kasper EK,
Wittstein IS, Champion HC, Breton E, Russell SD, Hall J, Kittleson
MM, Baughman KL and Hare JM: The gene expression profile of
patients with new-onset heart failure reveals important
gender-specific differences. Eur Heart J. 31:1188–1196. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Spinale FG, Janicki JS and Zile MR:
Membrane-associated matrix proteolysis and heart failure. Circ Res.
112:195–208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qi RB, Zhang JY, Lu DX, Wang HD, Wang HH
and Li CJ: Glycine receptors contribute to cytoprotection of
glycine in myocardial cells. Chin Med J (Engl). 120:915–921. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wagner S, Maier LS and Bers DM: Role of
sodium and calcium dysregulation in tachyarrhythmias in sudden
cardiac death. Circ Res. 116:1956–1970. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Papadatos GA, Wallerstein PM, Head CE,
Ratcliff R, Brady PA, Benndorf K, Saumarez RC, Trezise AE, Huang
CL, Vandenberg JI, et al: Slowed conduction and ventricular
tachycardia after targeted disruption of the cardiac sodium channel
gene Scn5a. Proc Natl Acad Sci USA. 99:6210–6215. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang X, Tang H, Wei EQ, Wang Z, Yang J,
Yang R, Wang S, Zhang Y, Pitt GS, Zhang H and Wang C: Conditional
knockout of Fgf13 in murine hearts increases arrhythmia
susceptibility and reveals novel ion channel modulatory roles. J
Mol Cell Cardiol. 104:63–74. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kanehisa M, Sato Y, Kawashima M, Furumichi
M and Tanabe M: KEGG as a reference resource for gene and protein
annotation. Nucleic Acids Res. 44((D1)): D457–D462. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nishida K, Yamaguchi O and Otsu K:
Degradation systems in heart failure. J Mol Cell Cardiol.
84:212–222. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hidron A, Vogenthaler N, Santos-Preciado
JI, Rodriguez-Morales AJ, Franco-Paredes C and Rassi A Jr: Cardiac
involvement with parasitic infections. Clin Microbiol Rev.
23:324–349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Herter JM, Rossaint J and Zarbock A:
Platelets in inflammation and immunity. J Thromb Haemost.
12:1764–1775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Barbalic M, Schwartz GL, Chapman AB,
Turner ST and Boerwinkle E: Kininogen gene (KNG) variation has a
consistent effect on aldosterone response to antihypertensive drug
therapy: The GERA study. Physiol Genomics. 39:56–60. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gomez-Sanchez CE: Non renal effects of
aldosterone. Steroids. 91:1–2. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qin CX, May LT, Li R, Cao N, Rosli S, Deo
M, Alexander AE, Horlock D, Bourke JE, Yang YH, et al:
Small-molecule-biased formyl peptide receptor agonist compound 17b
protects against myocardial ischaemia-reperfusion injury in mice.
Nat Commun. 8:142322017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bojnik E, Kleczkowska P, Marron Fernandez
de Velasco E, Corbani M, Babos F, Lipkowski AW, Magyar A and Benyhe
S: Bioactivity studies on atypical natural opioid hexapeptides
processed from proenkephalin (PENK) precursor polypeptides. Comp
Biochem Physiol B Biochem Mol Biol. 174:29–35. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ng LL, Sandhu JK, Narayan H, Quinn PA,
Squire IB, Davies JE, Bergmann A, Maisel A and Jones DJ:
Proenkephalin and prognosis after acute myocardial infarction. J Am
Coll Cardiol. 63:280–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Treskatsch S, Feldheiser A, Shaqura M,
Dehe L, Habazettl H, Röpke TK, Shakibaei M, Schäfer M, Spies CD and
Mousa SA: Cellular localization and adaptive changes of the cardiac
delta opioid receptor system in an experimental model of heart
failure in rats. Heart Vessels. 31:241–250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Siong Chan DC, Cao TH and Ng LL:
Proenkephalin in heart failure. Heart Fail Clin. 14:1–11. 2018.
View Article : Google Scholar : PubMed/NCBI
|