Introduction
Phosphoglycerate mutase 1 (PGAM1) is an important
glycolytic enzyme that catalyzes the conversion of
3-phosphoglycerate into 2-phosphoglycerate in the glycolytic
pathway (1,2). PGAM1 is normally expressed in the
brain, liver and kidney tissues (3,4).
PGAM1 overexpression has previously been observed in multiple human
cancer types, including breast carcinoma, lung cancer,
hepatocellular carcinoma, oral squamous cellular carcinoma and
urothelial bladder cancer (5–9).
Furthermore, PGAM1 has been reported to be associated with the
migration, proliferation and apoptosis of tumor cells (10–13).
PGAM1 has also been reported to be associated with
autoimmune central nervous system diseases (14). The current group reported that
PGAM1 is involved in spermatogenic dysfunction and affected cell
proliferation, apoptosis and migration (15). However, the effect and mechanism of
PGAM1 knockdown on spermatogenic cells remains unclear. In the
current study, the aim was to investigate the association between
PGAM1 expression and busulfan-induced hypospermatogenesis and the
effect of PGAM1 knockdown on spermatogenic cell apoptosis. Firstly,
PGAM1 expression was detected in a mouse model of busulfan-induced
hypospermatogenesis by western blotting, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
immunohistochemistry (IHC). Then, spermatogenic cell apoptosis was
assessed by terminal deoxynucleotidyl-transferase-mediated dUTP
nick end labeling (TUNEL) assay. Finally, the effect and potential
mechanism of PGAM1 downregulation on spermatogenic cells was
explored.
Materials and methods
Animal model construction
A total of 10 C57 male mice weighing 18–21 g were
purchased and raised in Guangzhou Medical University Animal Center
(Guangzhou, China). All mice were housed under standard conditions
(21±2°C, 40% humidity and 12-h light/dark cycle), and had free
access to water and food. The procedures were performed as
described previously (15,16). Briefly, mouse models of
hypospermatogenesis (N=5) were established by a single
intraperitoneal injection of busulfan (30 mg/kg). Normal controls
(N=5) didn't receive any treatment. Following 2 weeks, fresh testis
tissues were collected and stored in liquid nitrogen or fixed with
1.25% Bouin solution for 6 h at room temperature. The current study
was approved by the Guangzhou Medical University Institutional
Animal Care and Use Committee.
Hematoxylin and eosin (HE)
staining
Testis tissues were fixed with Bouin's solution for
6 h at room temperature and embedded in paraffin. Sections
(2-µm-thick) were heated at 65°C for 4 h, deparaffinized with
dimethylbenzene, rehydrated in gradient ethanol series and stained
with HE for 5 min at room temperature. Under the light microscope,
reduced production of spermatozoid in seminiferous tubules was
easily observed in mouse models of hypospermatogenesis by counting
the number of sperm cells.
RT-qPCR
Total RNA was extracted from testis tissues and
spermatogenic cells using RNA TRIzol reagent (Takara Bio, Inc.,
Otsu, Japan) and synthesized using the PrimeScript™ RT Reagent kit
(Takara Bio, Inc.) at 37°C for 15 min. qPCR analysis was performed
using the SYBR Green RT-PCR kit (Takara Bio, Inc.) according to the
manufacturer's protocol. The primers of PGAM1 and GAPDH were
purchased from Shanghai GenePharma Co., Ltd. (Shanghai, China). The
following primers were used: PGAM1, forward (F)
5′-ATGCTAAGCCATGACCAGTGAG-3′ and reverse (R)
5′-ATCACCACGCAGGTTACATTCG-3′; GAPDH, F 5′-AGGTCGGTGTGAACGGATTTG-3′
and R 5′-TGTAGACCATGTAGTTGAGGTCA-3′. Cycling conditions were 95°C
for 5 min, followed by 40 cycles at 95°C for 40 sec, 60°C for 30
sec and 72°C for 30 sec. Experiments were repeated three times for
each reaction. The 2−ΔΔCq method was used to determine
the relative mRNA expression levels (16).
Western blot analysis
Total protein lysates were extracted from testis
tissues and spermatogenic cells using cell lysis buffer (cat. no.
P0013; Beyotime Institute of Biotechnology, Shanghai, China)
containing a protease inhibitor cocktail (cat. no. P1006; Beyotime
Institute of Biotechnology) and 100 nM phenylmethanesufonyl
fluoride. Protein concentration was determined by bicinchoninic
acid assay. Subsequently, 50 µg protein was separated by 10%
SDS-PAGE and transferred onto a polyvinylidene difluoride membrane
(EMD Millipore, Billerica, MA, USA). The membranes were dipped into
PBS with 0.1% Tween 2000 (BestBio, Shanghai, China) solution and
blocked with 5% skimmed milk at room temperature for 2 h, and then
incubated with rabbit monoclonal PGAM1 antibody (cat. no. ab129191;
1:1,000; Abcam, Cambridge, MA, USA), rabbit polyclonal P53 antibody
(cat. no. ab16033; 1:2,000; Abcam), rabbit polyclonal Caspase 3
antibody (cat. no. ab13847; 1:500; Abcam), rabbit polyclonal
Caspase 6 antibody (cat. no. ab231349; 1:500; Abcam), rabbit
polyclonal Caspase 9 antibody (cat. no. ab52298; 1:500; Abcam) and
monoclonal tubulin antibody (cat. no. ab6160; 1:2,000; Abcam),
respectively. Finally, polyvinylidene difluoride membranes were
incubated with a rabbit anti-goat horseradish peroxidase-conjugated
secondary antibody (cat. no. ZB-2306; 1:1,000; OriGene
Technologies, Inc., Beijing, China) at room temperature for 30 min.
Signals were detected using an EasyBlot ECL kit (cat. no.
C506668-0050; Sangon Biotech Co., Ltd., Shanghai, China) and an
enhanced chemiluminescence detecter (ProteinSimple; Bio-Techne,
Minneapolis, MN, USA).
IHC
Testis tissues were fixed with Bouin's solution for
6 h at room temperature and embedded in paraffin. Sections
(2-µm-thick) were deparaffinized with dimethylbenzene and
rehydrated with gradient ethanol. Antigen retrieval was performed
with 0.01 mol/l sodium citrate buffer (pH 7.0). The sections were
incubated with rabbit monoclonal PGAM1 antibody (cat. no. ab129191;
1:150; Abcam) for 2 h, followed by incubation with 50 µl
horseradish peroxidase-labeled goat anti-mouse/rabbit IgG polymer
(cat. no. PV-6000; OriGene Technologies, Inc.) for 30 min at room
temperature. Finally, sections were reacted with diaminobenzidine
at room temperature for 1–3 min and stained with hematoxylin for 3
min at room temperature. Negative controls were performed by
replacing the primary antibody with PBS. Sections with positive
expression of PGAM1 were treated as positive controls.
TUNEL assay
The procedures were performed as described
previously (17). Testis tissues
were fixed with Bouin's solution for 12 h at room temperature and
embedded in paraffin. Sections (3-µm-thick) were deparaffinized
with dimethylbenzene and rehydrated with gradient ethanol. Then,
sections were incubated with 1% proteinase K (20 mg/ml) at 37°C for
30 min. Following washing with PBS, sections were incubated with
TUNEL mix (45 µl Equilibration Buffer, 1.0 µl biotin-11-dUTP, 4.0
µl TdT enzyme and 50 µl reaction buffer) at 37°C for 60 min.
Finally, sections were stained with DAPI for 5 min at room
temperature and mounted in 50% glycerin diluted in water. A total
of 10 randomly chosen microscopic fields were analyzed under a
fluorescence microscope (magnification, ×40). The nuclei of
apoptotic cells were marked as red under fluorescence microscopy at
570 nm.
Cell culture and transfection
GC-1 spg cells (a mouse spermatogonia cell line) and
GC2 cells (a mouse spermatocyte cell line) were purchased from the
American Type Culture Collection (Manassas, VA, USA). Cells were
cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) at 37°C
in a 5% CO2 incubator. Cells at 40–60% confluence were
transfected with PGAM1 short hairpin (sh)RNA (sense,
5′-ATGCTAAGCCATGACCAGTGAG-3′ and anti-sense,
5′-ATCACCACGCAGGTTACATTCG-3′) lentivirus and recombinant PGAM1
lentivirus, respectively. The negative control was transfected with
scrambled shRNA lentivirus (sense, 5′-UUCUCCGAACGUGUCACGU-3′ and
anti-sense, 5′-AAGAGGCUUGCACAUGCA-3′).
Apoptosis analysis
At 48 h following transfection, 1×106 GC1
and GC2 cells were collected and washed with PBS. Then, cells were
incubated with propidium iodide (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and Annexin V-FITC (BestBio) at room
temperature for 15 min. Cell apoptosis was analyzed using a BD
FACSCalibur™ flow cytometer and BD CellQuest™ software version 5.1
(BD Biosciences, Franklin Lakes, NJ, USA).
Statistical analysis
All experiments were repeated three times and the
data are presented as the mean ± standard deviation. All data were
analyzed using SPSS 19.0 software (IBM Corp., Armonk, NY, USA).
Independent samples t test was used to compare the difference
between two groups. All tests were two-sided and P<0.05 was
considered to indicate a statistically significant difference.
Results
PGAM1 downregulation is associated
with busulfan-induced hypospermatogenesis and spermatogenic cell
apoptosis
To investigate the association between PGAM1
expression and hypospermatogenesis, the expression of PGAM1 was
detected in mouse models of busulfan-induced hypospermatogenesis by
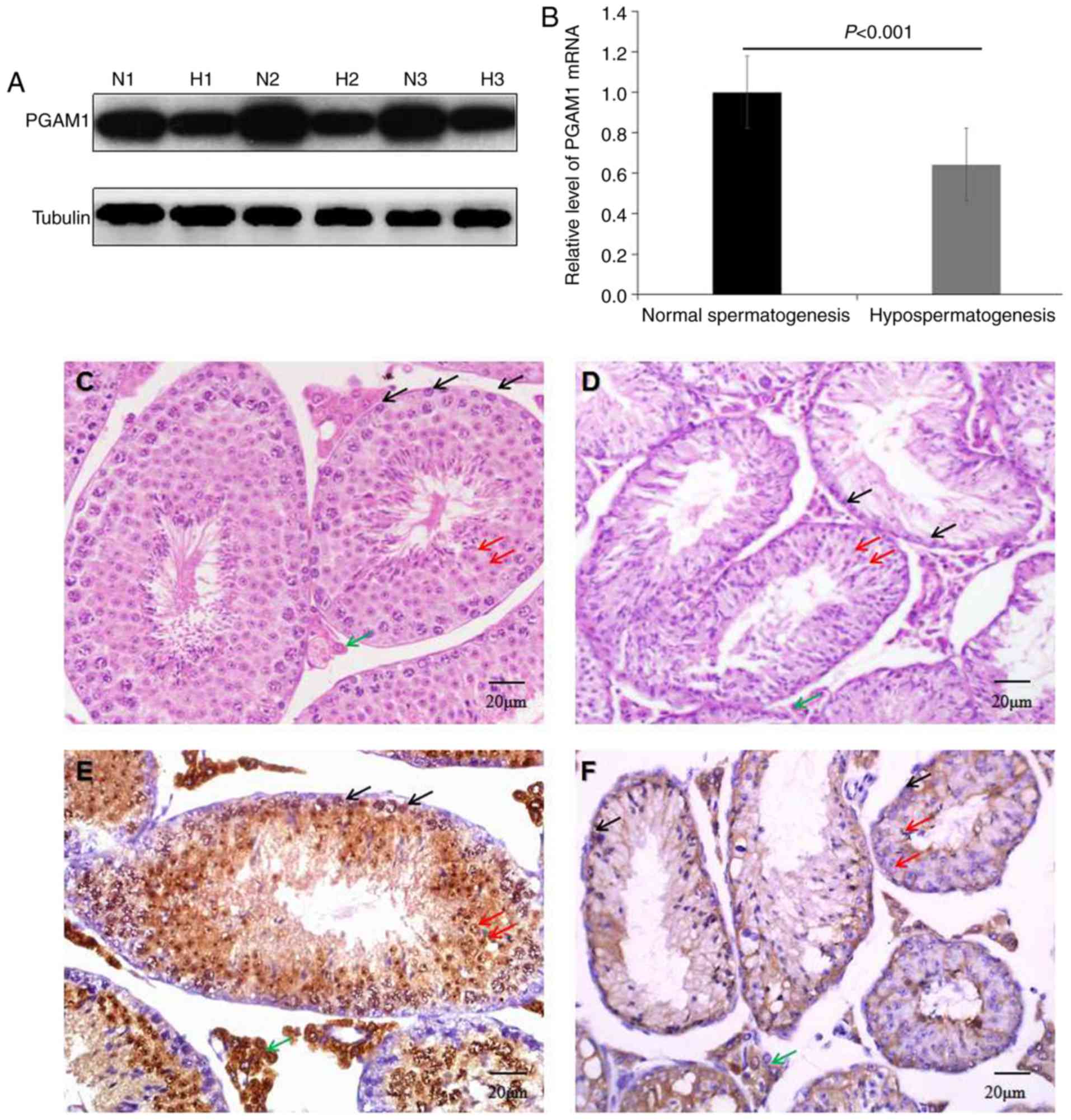
western blotting, RT-qPCR and IHC (Fig. 1). Western blotting (Fig. 1A) and RT-qPCR (Fig. 1B) results revealed that PGAM1
expression levels were significantly decreased compared with normal
controls (P<0.001). HE staining demonstrated that the number of
spermatogonia, spermatocytes and spermatozoid in seminiferous
tubules was decreased in mouse models of busulfan-induced
hypospermatogenesis compared with the controls (Fig. 1C and D). IHC results demonstrated
that positive PGAM1 expression, visible as brown staining, was
present in spermatogonia, spermatocytes and Leydig cells in the
testis tissues with normal spermatogenesis (Fig. 1E), while expression was hardly
detected in mouse models of busulfan-induced hypospermatogenesis
(Fig. 1F). These data indicated
that PGAM1 downregulation was associated with busulfan-induced
hypospermatogenesis.
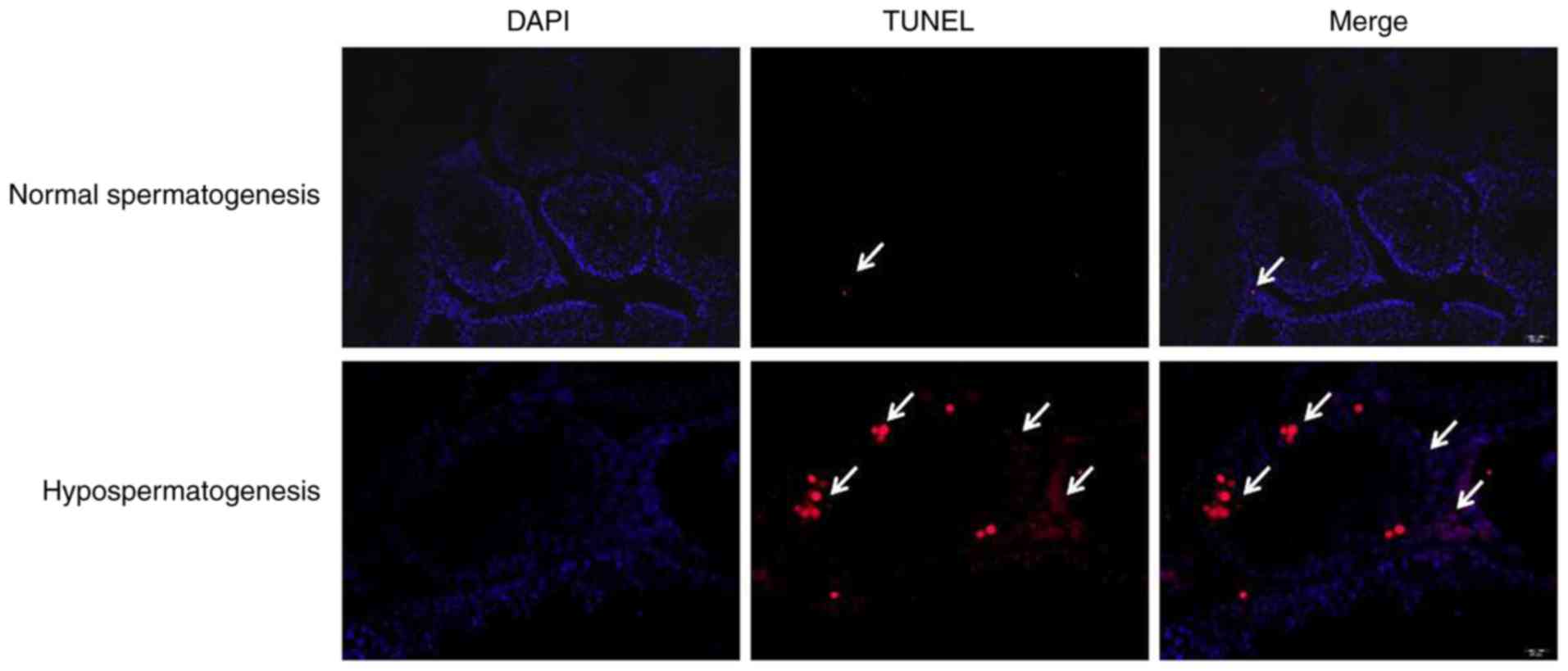
To investigate the association between PGAM1
expression and spermatogenic cell apoptosis in a mouse model of
busulfan-induced hypospermatogenesis, a TUNEL assay was performed.
As presented in Fig. 2, the
apoptosis rate of spermatogenic cells was accelerated in mouse
models of hypospermatogenesis compared with normal spermatogenesis.
These data indicated that PGAM1 downregulation may be associated
with spermatogenic cell apoptosis in mouse models of
busulfan-induced hypospermatogenesis.
PGAM1 knockdown promotes spermatogenic
cell apoptosis by regulating the P53/Caspase 3/Caspase 6/Caspase 9
pathway
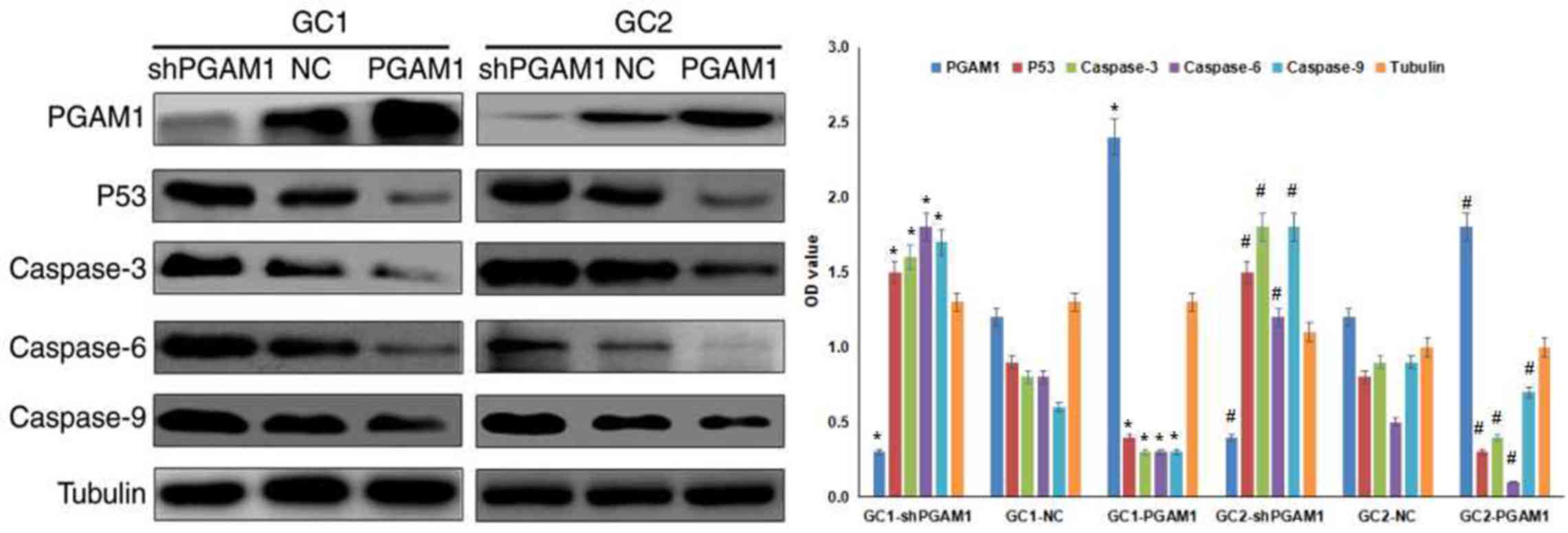
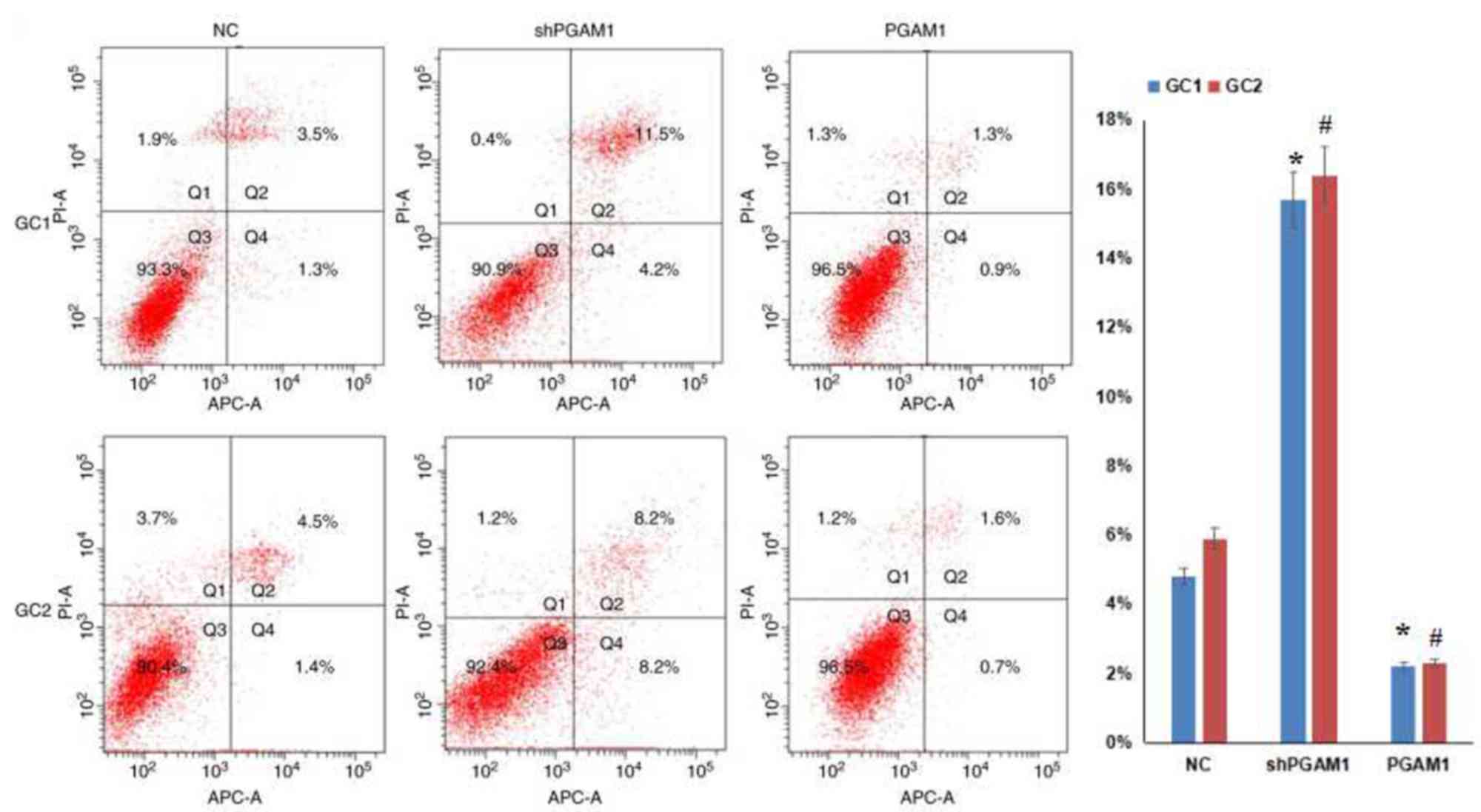
To further investigate the association between PGAM1
expression and spermatogenic cell apoptosis in vitro, a flow
cytometry assay was performed. Firstly, spermatogenic cells were
infected with PGAM1 shRNA lentivirus or recombinant PGAM1
lentivirus. As presented in Fig.
3, western blotting results indicated that PGAM1 was
successfully knocked down and upregulated following transfection,
respectively. Then, cell apoptosis was measured by flow cytometry
(Fig. 4). The rate of apoptosis
was 15.7±2.3% in GC1 cells with PGAM1 downregulation, which was
significantly higher compared with the negative controls (4.8±0.9%;
P<0.001). The rate of apoptosis was 2.2±0.6% in GC1 cells with
PGAM1 upregulation, which was significantly reduced compared with
negative controls (4.8±0.9%; P<0.001). In addition, the rate of
apoptosis was 16.4±2.8% in GC2 cells with PGAM1 knockdown, which
was significantly greater compared with negative controls
(5.9±1.1%; P<0.001). When PGAM1 expression was upregulated in
GC2 cells, the rate of apoptosis was 2.3±0.5%, which was
significantly reduced compared with negative controls (5.9±1.1%;
P<0.001). These data demonstrated that PGAM1 knockdown
contributed to spermatogenic cell apoptosis.
To investigate the mechanism of PGAM1, the
expression levels of P53, Caspase 3, Caspase 6 and Caspase 9 were
detected by western blotting. As presented in Fig. 3, the expression levels of P53,
Caspase 3, Caspase 6 and Caspase 9 were significantly elevated
following PGAM1 knockdown. However, PGAM1 upregulation produced the
opposite effects. These data indicated that PGAM1 knockdown
activated the P53/Caspase 3/Caspase 6/Caspase 9 signaling pathway,
which may result in spermatogenic cell apoptosis.
Discussion
Hypospermatogenesis is prevalent in azoospermic
patients and is associated with male infertility. It is
characterized by a lower production of spermatozoids in
seminiferous tubules (18,19). It is well known that abnormal
apoptosis of spermatogenic cells is implicated in
hypospermatogenesis (20–23). PGAM1, an important glycolytic
enzyme, is reported to be associated with the migration,
proliferation and apoptosis of tumor cells (10–13).
Furthermore, PGAM1 is involved in spermatogenic dysfunction and
affects cell proliferation, apoptosis and migration (15). Therefore, it has been demonstrated
that PGAM1 may be associated with hypospermatogenesis and
spermatogenic cell apoptosis. However, the mechanism of PGAM1 in
cell apoptosis remains unclear.
In the current study, the association between PGAM1
expression and busulfan-induced hypospermatogenesis was
investigated. First, PGAM1 expression was detected in a mouse model
of busulfan-induced hypospermatogenesis by western blotting,
RT-qPCR and IHC. Western blotting and RT-qPCR results revealed that
PGAM1 expression was reduced in mice with busulfan-induced
hypospermatogenesis compared with those with normal
spermatogenesis. IHC results indicated that PGAM1 was weakly
expressed in the mouse model of hypospermatogenesis, which was
consistent with a previous report (15). These data supported the proposal
that PGAM1 downregulation was associated with busulfan-induced
hypospermatogenesis. To investigate the association between PGAM1
and hypospermatogenesis and spermatogenic cell apoptosis, a TUNEL
assay was performed. The results demonstrated that spermatogenic
cell apoptosis was accelerated indicating that PGAM1 downregulation
may be associated with spermatogenic cell apoptosis in a mouse
model of busulfan-induced hypospermatogenesis. To further validate
the association between PGAM1 expression and spermatogenic cell
apoptosis, PGAM1 expression was successfully knocked down and
upregulated in spermatogenic cells. Subsequently, flow cytometry
results demonstrated that PGAM1 downregulation promoted the
apoptosis of spermatogenic cells. Furthermore, apoptotic proteins,
including P53, Caspase 3, Caspase 6 and Caspase 9, were activated
when PGAM1 expression was knocked down. P53, an important apoptotic
protein, participates in cell cycle arrest, senescence and
apoptosis of spermatogenic cells (24–26).
The activation of p53 signaling may induce mitochondria-associated
apoptotic cell death and disrupt sperm production and fertility
(27,28). In addition, caspase-mediated
apoptosis is involved in normal spermatogenesis (17). Caspase 3 and Caspase 6 may be
activated by Caspase 9 to cause cell apoptosis (29,30).
Therefore, these data indicated that PGAM1 knockdown promoted
spermatogenic cell apoptosis by regulating the P53/Caspase
3/Caspase 6/Caspase 9 pathway.
In conclusion, the current data indicates that PGAM1
knockdown is associated with busulfan-induced hypospermatogenesis
and spermatogenic cell apoptosis. Furthermore, spermatogenic cell
apoptosis caused by PGAM1 knockdown is associated with the
P53/Caspase 3/Caspase 6/Caspase 9 pathway. However, the current
data are not sufficient to explain the detailed mechanism of PGAM1
in spermatogenic cell apoptosis. Therefore, further investigations
in this area are required.
Acknowledgements
Not applicable.
Funding
This study was supported by the Young Science
Foundation of Guangzhou Medical University (grant no. 2016A01), the
Young Cultivation Foundation of PLA Medical science and Technology
(grant no. 17QNP044) and the Medical Research Foundation of
Guangdong province (grant no. A2017275).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and SZ designed the experiments; YZ performed in
the experiments; SZ analyzed the data and wrote the paper.
Ethics approval and consent to
participate
The present study was approved by the Guangzhou
Medical University Institutional Animal Care and Use Committee.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jiang X, Sun Q, Li H, Li K and Ren X: The
role of phosphoglycerate mutase 1 in tumor aerobic glycolysis and
its potential therapeutic implications. Int J Cancer.
135:1991–1996. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jacobowitz DM, Jozwik C, Fukuda T and
Pollard HB: Immunohistochemical localization of phosphoglycerate
mutase in capillary endothelium of the brain and periphery.
Microvasc Res. 76:89–93. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Betrán E, Wang W, Jin L and Long M:
Evolution of the phosphoglycerate mutase processed gene in human
and chimpanzee revealing the origin of a new primate gene. Mol Biol
Evol. 19:654–663. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gao H, Yu B, Yan Y, Shen J, Zhao S, Zhu J,
Qin W and Gao Y: Correlation of expression levels of ANXA2, PGAM1,
and CALR with glioma grade and prognosis. J Neurosurg. 118:846–853.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bührens RI, Amelung JT, Reymond MA and
Beshay M: Protein expression in human non-small cell lung cancer: A
systematic database. Pathobiology. 76:277–285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Durany N, Joseph J, Jimenez OM, Climent F,
Fernández PL, Rivera F and Carreras J: Phosphoglycerate mutase,
2,3-bisphosphoglycerate phosphatase, creatine kinase and enolase
activity and isoenzymes in breast carcinoma. Br J Cancer. 82:20–27.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Turhani D, Krapfenbauer K, Thurnher D,
Langen H and Fountoulakis M: Identification of differentially
expressed, tumor-associated proteins in oral squamous cell
carcinoma by proteomic analysis. Electrophoresis. 27:1417–1423.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ren F, Wu H, Lei Y, Zhang H, Liu R, Zhao
Y, Chen X, Zeng D, Tong A, Chen L, et al: Quantitative proteomics
identification of phosphoglycerate mutase 1 as a novel therapeutic
target in hepatocellular carcinoma. Mol Cancer. 9:812010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Peng XC, Gong FM, Chen Y, Qiu M, Cheng K,
Tang J, Ge J, Chen N, Zeng H and Liu JY: Proteomics identification
of PGAM1 as a potential therapeutic target for urothelial bladder
cancer. J Proteomics. 132:85–92. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu Z, Gong J, Wang C, Wang Y, Song Y, Xu
W, Liu Z and Liu Y: The diagnostic value and functional roles of
phosphoglycerate mutase 1 in glioma. Oncol Rep. 36:2236–2244. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang D, Jin N, Sun W, Li X, Liu B, Xie Z,
Qu J, Xu J, Yang X, Su Y, et al: Phosphoglycerate mutase 1 promotes
cancer cell migration independent of its metabolic activity.
Oncogene. 36:2900–2909. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang D, Wu H, Zhang X, Ding X, Huang M,
Geng M, Li H and Xie Z: Phosphoglycerate mutase 1 predicts the poor
prognosis of oral squamous cell carcinoma and is associated with
cell migration. J Cancer. 8:1943–1951. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qu J, Sun W, Zhong J, Lv H, Zhu M, Xu J,
Jin N, Xie Z, Tan M, Lin SH, et al: Phosphoglycerate mutase 1
regulates dNTP pool and promotes homologous recombination repair in
cancer cells. J Cell Biol. 216:409–424. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kimura A, Sakurai T, Koumura A, Yamada M,
Hayashi Y, Tanaka Y, Hozumi I, Tanaka R, Takemura M, Seishima M and
Inuzuka T: High prevalence of autoantibodies against
phosphoglycerate mutase 1 in patients with autoimmune central
nervous system diseases. J Neuroimmunol. 219:105–108. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang S, Zhao Y, Lei B, Li C and Mao X:
PGAM1 is Involved in spermatogenic dysfunction and affects cell
proliferation, apoptosis, and migration. Reprod Sci. 22:1236–1242.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lei B, Zhou X, Lv D, Wan B, Wu H, Zhong L,
Shu F and Mao X: Apoptotic and nonapoptotic function of caspase 7
in spermatogenesis. Asian J Androl. 19:47–51. 2017.PubMed/NCBI
|
|
18
|
Robin G, Boitrelle F, Leroy X, Peers MC,
Marcelli F, Rigot JM and Mitchell V: Assessment of azoospermia and
histological evaluation of spermatogenesis. Ann Pathol. 30:182–195.
2010.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng YS, Lu CW, Lin TY, Lin PY and Lin
YM: Causes and clinical features of infertile men with
nonobstructive azoospermia and histopathologic diagnosis of
hypospermatogenesis. Urology. 105:62–68. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takagi S, Itoh N, Kimura M, Sasao T and
Tsukamoto T: Spermatogonial proliferation and apoptosis in
hypospermatogenesis associated with nonobstructive azoospermia.
Fertil Steril. 76:901–907. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jaiswal D, Trivedi S, Agrawal NK and Singh
K: Dysregulation of apoptotic pathway candidate genes and proteins
in infertile azoospermia patients. Fertil Steril. 104:736–743.e6.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hegazy R, Hegazy A, Ammar M and Salem E:
Immunohistochemical measurement and expression of Mcl-1 in
infertile testes. Front Med. 9:361–367. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han K, Seo HW, Oh Y, Kang I, Park C, Han
JH, Kim SH and Chae C: Pathogenesis of type 1 (European genotype)
porcine reproductive and respiratory syndrome virus in male gonads
of infected boar. Vet Res Commun. 37:155–162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Napoletano F, Gibert B, Yacobi-Sharon K,
Vincent S, Favrot C, Mehlen P, Girard V, Teil M, Chatelain G,
Walter L, et al: p53-dependent programmed necrosis controls germ
cell homeostasis during spermatogenesis. PLoS Genet.
13:e10070242017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li T, Liu X, Jiang L, Manfredi J, Zha S
and Gu W: Loss of p53-mediated cell-cycle arrest, senescence and
apoptosis promotes genomic instability and premature aging.
Oncotarget. 7:11838–11849. 2016.PubMed/NCBI
|
|
26
|
Duan P, Hu C, Butler HJ, Quan C, Chen W,
Huang W, Tang S, Zhou W, Yuan M, Shi Y, et al: 4-Nonylphenol
induces disruption of spermatogenesis associated with oxidative
stress-related apoptosis by targeting p53-Bcl-2/Bax-Fas/FasL
signaling. Environ Toxicol. 32:739–753. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen D, Zheng W, Lin A, Uyhazi K, Zhao H
and Lin H: Pumilio 1 suppresses multiple activators of p53 to
safeguard spermatogenesis. Curr Biol. 22:420–425. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao Y, Tan Y, Dai J, Li B, Guo L, Cui J,
Wang G, Shi X, Zhang X, Mellen N, et al: Exacerbation of
diabetes-induced testicular apoptosis by zinc deficiency is most
likely associated with oxidative stress, p38 MAPK activation, and
p53 activation in mice. Toxicol Lett. 200:100–106. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Logue SE and Martin SJ: Caspase activation
cascades in apoptosis. Biochem Soc Trans. 36:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Das J, Ghosh J, Manna P and Sil PC:
Taurine protects rat testes against doxorubicin-induced oxidative
stress as well as p53, Fas and caspase 12-mediated apoptosis. Amino
Acids. 42:1839–1855. 2012. View Article : Google Scholar : PubMed/NCBI
|