Introduction
Kidney transplantation is the only effective
treatment for end-stage renal disease and the 10-year survival rate
following allografts is <50% (1,2).
Interstitial fibrosis and tubular atrophy are the principal
pathological features of chronic allograft nephropathy (CAN), which
is the primary cause of late renal allograft loss (3,4).
Epithelial-mesenchymal transition (EMT) has been demonstrated to
serve a crucial role in the process of interstitial fibrosis
(5).
CXC chemokine receptor 4 (CXCR4), the receptor of
stromal cell-derived factor 1 (SDF-1), has been demonstrated to be
expressed at lower levels in normal human kidneys (6), and the expression levels were
increased in a mouse model of renal ischemia (7). Activation of the SDF-1/CXCR4 pathway
has been demonstrated to contribute to the progression of CAN, and
inhibition of CXCR4 by AMD3100 attenuates the progression of renal
allograft fibrosis (8). Blockade
of SDF-1/CXCR4 by AMD3100 has been reported to serve a protective
role following kidney transplantation by mobilizing stem cells
(9–11), but the mechanism in allografts has
not been studied to the best of our knowledge. Studies have
demonstrated that SDF-1 is able to induce EMT and tissue fibrosis
(7,12,13),
including kidney fibrosis (14,15).
The Wnt signaling pathway has been reported to be involved in the
initiation and progression of chronic renal allograft damage in a
rat model (16). Previous studies
have also reported that the Wnt/β-catenin pathway serves an
important role in SDF-1-induced EMT in numerous types of tumors
(17,18). However, there are few studies that
have focused on the mechanisms of EMT in tubular epithelial cells
following kidney transplantation. To the best of our knowledge, the
present study is the first to investigate the mechanisms of SDF-1
and the Wnt pathway in kidney allograft fibrosis.
In the present study, EMT was demonstrated to be
ameliorated by AMD3100 in the rat CAN model. Variations in markers
and RNAs in the rat CAN model were analyzed and the results
revealed the association between SDF-1/CXCR4 and the Wnt pathway.
Subsequently, the involvement of the Wnt signaling pathway in
SDF-1-induced renal tubular EMT was investigated in vitro,
which may be a novel mechanism of renal allograft fibrosis.
Materials and methods
Animal models
The experimental animals included 10 male Fisher
(F344) rats as the donors and 10 male Lewis (LEW) rats as the
recipients. All animals were aged between 8–12 weeks and weighed
200–250 g. Animals were provided by Beijing Vital River Laboratory
Animal Technology Co., Ltd. (Beijing, China). The F344-LEW rat CAN
model was established as previously described (8). The rats were divided randomly into
the following two groups: The AMD3100 group and the CAN group. The
rats in the AMD3100 group were administered subcutaneous injections
of low-dose cyclosporine (1.5 mg/kg) every day in the first 10 days
post-transplantation and AMD3100 (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany; 1 mg/kg) at 0, 2, 4, 6, 8 and 10 days
post-transplantation, and the rats in the CAN group were
administered subcutaneous injections of low-dose cyclosporine (1.5
mg/kg/day) and saline at a dose equivalent to AMD3100 in the first
10 days following transplantation. At 12 weeks post-surgery, the
kidney allografts in both the AMD3100 and CAN groups were harvested
under anesthesia using 2% pentobarbital sodium (40 mg/kg), and the
rats were subsequently administered pentobarbital sodium (200
mg/kg). The animals were protected by following the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals (19). The animal
protocols were approved by the Medical Research Center, Beijing
Chaoyang Hospital, Capital Medical University (Beijing, China). The
housing conditions were as follows: Temperature, 22±1°C; relative
humidity, 55±5% and free access to water and normal diet under an
alternating 12:12-h light-dark cycle.
Histopathology and
immunohistochemistry
The kidneys were cut longitudinally along the
coronal plane and fixed in 10% neutral formalin solution at room
temperature for 24 h, followed by dehydration with alcohol,
embedding in paraffin, and sectioning into 2-µm thick slices.
Hematoxylin and eosin, and Masson's trichrome staining were
performed to assess the renal injury. For hematoxylin and eosin
staining, the sections were stained at room temperature with
hematoxylin for 5 min and eosin for 2 min. For Masson's trichrome
staining (all steps conducted at room temperature), the nuclei were
stained with Weigert's iron hematoxylin for 10 min, followed by
plasma membrane staining with acid fuchsin, Xylidine Ponceau, and
glacial acetic acid for 10 min, phosphomolybdic acid staining for 5
min, and fiber staining with methyl blue for 10 min. A total of two
experienced pathologists independently evaluated the results of the
histopathological analysis, and the images were acquired at ×400
magnification. The chronic allograft damage index (CADI) scores of
the kidney specimens in the two groups were determined. The CADI
scoring system covers the following six pathological aspects of
CAN: Renal interstitial inflammation, interstitial fibrosis,
tubular atrophy, basement membrane matrix thickening, glomerular
sclerosis and arterial intimal hyperplasia (20).
Immunohistochemistry was performed using the same
procedures of dehydration, embedding and sectioning. Endogenous
peroxidase was blocked with 3% H2O2 at room
temperature for 10 min, and then the specimens were heated in a
citrate buffer (pH 6.0) to 98°C for 15 min. The primary goat
anti-CXCR4 antibody (cat. no. ab1670; Abcam, Cambridge, UK;
1:1,000) was added in a dropwise manner to the sections, which were
incubated at 4°C overnight. Subsequently, the specimens were
incubated with horseradish peroxidase (HRP)-conjugated donkey
anti-goat immunoglobulin G (cat. no. A0181; Beyotime Institute of
Biotechnology, Haimen, China; 1:50) at 37°C for 30 min. A freshly
prepared 0.05% 3,3′-diaminobenzidine solution was added to each
specimen section. The color reaction was observed and controlled by
light microscopy (Olympus Corporation, Tokyo, Japan), and
terminated by washing with tap water. Images were acquired at ×400
magnification.
Microarrays and pathway analysis
Total RNA was extracted from the kidney tissues
using TRIzol® (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) reagent, and the purity and concentration
of the RNA was determined from the OD260/280 readings of
a spectrophotometer. The RNA integrity was determined by capillary
electrophoresis using the RNA 6000 Nano Lab-on-a-Chip kit and a
Bioanalyzer 2100 (both Agilent Technologies, Inc., Santa Clara, CA,
USA). Only RNA extracts with RNA integrity values of >6 were
subjected to further analysis. Higher yields of cDNA were labeled
with a fluorescent dye (Cy5 and Cy3-dCTP) using the CapitalBio cRNA
Amplification and Labeling kit (CapitalBio, Beijing, China). The
labeled cRNAs from long noncoding (lnc)RNAs and mRNAs were purified
and hybridized to the Agilent Rat lncRNA + mRNA Array V1.0 (Agilent
Technologies, Inc.). Images were captured by the Agilent microarray
scanner, gridded and then analyzed using Agilent Feature Extraction
software, version 10.10 (Agilent Technologies, Inc.). The raw data
were summarized and normalized using GeneSpring software V12.0
(Agilent Technologies, Inc.).
The threshold values of ≥2 absolute fold change (FC)
and a Benjamini-Hochberg corrected P-value of ≤0.05 were used. The
data were log2 transformed and median-centered using the Adjust
Data function of Cluster 3.0 software (http://bonsai.hgc.jp/~mdehoon/software/cluster/index.html).
These genes were classified according to the Gene Ontology (GO)
analysis provided by the Molecular Annotation System 3.0
(http://bioinfo.capitalbio.com/mas3/).
Signaling pathway analysis of these genes was performed with the
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database
(http://www.kegg.jp/kegg/pathway.html).
Cell culture
The rat renal tubular epithelial cell line NRK-52E
was purchased from American Type Culture Collection (Manassas, VA,
USA) and cultured in Dulbecco's modified Eagle's medium (DMEM)
containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.) and 1% penicillin/streptomycin (Lablead Biotech Co., Ltd.,
Beijing, China) at 37°C with 5% CO2. The NRK-52E cells
with optimal morphology were digested and seeded in a 60-mm dish at
a concentration of ~1×105 cells/dish.
Treatment in vitro
Once the cells had attached, the medium was changed
to DMEM without serum. After 24 h, the cells were divided into four
groups and treated as follows: SDF-1 group, 100 ng/ml SDF-1 (cat.
no. ab79959; Abcam); SDF-1 + AMD3100 group, 100 ng/ml SDF-1 and 5
µg/ml AMD3100 (Sigma-Aldrich; Merck KGaA); SDF-1 + DKK-1 group, 100
ng/ml SDF-1 and 200 ng/ml DKK-1 (R&D Systems, Inc.,
Minneapolis, MN, USA); and control group, PBS at a dose equivalent
to the SDF-1, AMD3100 and DKK-1 in the other groups.
Following incubation at 37°C for 24 h, the cellular
total protein and nuclear protein were extracted for western
blotting (WB) and the total RNA was extracted for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
WB
The cells were divided into two groups; one group
was lysed in radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology) containing phenylmethylsulfonyl
fluoride and the other group was lysed using a Nuclear and
Cytoplasmic Protein Extraction kit (Beyotime Institute of
Biotechnology). The lysed cells were centrifuged at 14,000 × g for
20 min at 4°C. The protein concentration was measured using a
bicinchoninic acid assay kit (Beyotime Institute of Biotechnology).
The proteins were mixed with loading buffer and boiled for 10 min,
and subjected to 10% SDS-PAGE, with 30 µg protein loaded per lane.
The separated proteins were transferred to polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA). Following blocking
of the nonspecific background in 5% milk at room temperature for 1
h, the membranes were incubated overnight at 4°C with the following
primary antibodies: Rabbit anti-α-tubulin (cat. no. 2125; Cell
Signaling Technology, Inc., Danvers, MA, USA; 1:2,000); rabbit
anti-α-SMA (cat. no. ab5694; Abcam; 1:4,000); mouse anti-E-cadherin
(cat. no. 14472; Cell Signaling Technology, Inc.; 1:1,000); rabbit
anti-CXCR4 (cat. no. ab124824; Abcam; 1:2,000); rabbit anti-lamin
B1 (cat. no. 13435; Cell Signaling Technology, Inc.; 1:1,000);
rabbit anti-CXXC finger protein 5 (CXXC5; cat. no. 84546; Cell
Signaling Technology, Inc.; 1:1,000) and rabbit anti-β-catenin
(cat. no. 8480; Cell Signaling Technology, Inc.; 1:1,000).
Subsequently, the membranes were washed with TBS with Tween-20 and
incubated with the HRP-conjugated secondary antibodies (cat. no.
TA130003 and cat. no. TA140003; OriGene Technologies, Inc.,
Beijing, China; 1:1,000) at room temperature for 1 h. Immobilon
Western Chemiluminescent Horseradish Peroxidase Substrate (EMD
Millipore) was used to detect positive immune reactions. α-Tubulin
was used as the reference for total protein and lamin B1 was used
as the reference for nuclear protein. Densitometric analysis was
performed using Image Lab 3.0 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
RT-qPCR
Total RNA was extracted from the cells using
TRIzol® reagent and reverse-transcribed into cDNA using
a SuperScript IV First-Strand Synthesis System (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocols.
The RT-qPCR reactions were performed on the ABI 7500 Real-Time PCR
System (Applied Biosystems; Thermo Fisher Scientific, Inc.) using a
SuperScript III Platinum SYBR Green One-Step RT-qPCR Kit
(Invitrogen; Thermo Fisher Scientific, Inc.); the primers used are
listed in Table I. The target mRNA
levels were normalized against GAPDH mRNA standards and calculated
using the 2−ΔΔCq method (21).
 | Table I.Primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers for reverse
transcription-quantitative polymerase chain reaction.
| Target gene | Primer name | Oligonucleotide
sequence (5′-3′) |
|---|
| GAPDH | GAPDH-F |
GGCAAGTTCAACGGCACAG |
|
| GAPDH-R |
CGCCAGTAGACTCCACGACAT |
| α-SMA | α-SMA-F |
AGCTGCTCCAGCTATGTGTG |
|
| α-SMA-R |
TCCCAGTTGGTGATGATGCC |
| E-cadherin | E-cadherin-F |
GTGCCACCACCAAAGATA |
|
| E-cadherin-R |
GGCTGAGACAACCCTAAT |
| CXXC5 | CXXC5-F |
GCAGTGCAGCAGTTGTAGGA |
|
| CXXC5-R |
GACGGAAGCATCACCTTCTC |
| LOC100912353 | LOC100912353-F |
CGGGAACCTAGGAATGACAA |
|
| LOC100912353-R |
CAACGTTCTTGGTCCTCCAT |
| LOC102551030 | LOC102551030-F |
AAGCCGGTGTGAAGATCAAC |
|
| LOC102551030-R |
TCCTCGGGAATCACAGAAAC |
| LOC102556393 | LOC102556393-F |
ATGTAGGTTTGCCCAAGCAC |
|
| LOC102556393-R |
AACTGCAGGACAGGCATCTAA |
Wound healing assay
The NRK-52E cells were cultured under standard
conditions to 80–90% confluence, and were pretreated with DMEM
without serum at 37°C for 24 h. Subsequently, the cells were
treated with 100 mg/ml SDF-1 with or without 5 µg/ml AMD3100 and
200 ng/ml DKK-1 during the wound healing assay. Cell migration was
assessed by measuring the movement of cells into the acellular area
created by a sterile insert at 0, 12 and 24 h following scratching
(magnification, ×400).
Statistical analysis
Statistical analysis and data processing were
performed using SPSS version 20 (IBM Corp., Armonk, NY, USA). The
normally distributed measurement data are expressed as the mean ±
standard deviation of experiments that were replicated three times.
The independent samples t-test was used to compare two groups when
the data were normally distributed and exhibited homogeneity of
variance. Comparison between multiple groups was conducted using
one-way analysis of variance followed by a least significance
difference test. If the data exhibited a skewed distribution or
failed to present homogeneity of variance, the Mann-Whitney U test
was used for the comparison of two groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
SDF-1/CXCR4 axis in the rat CAN
model
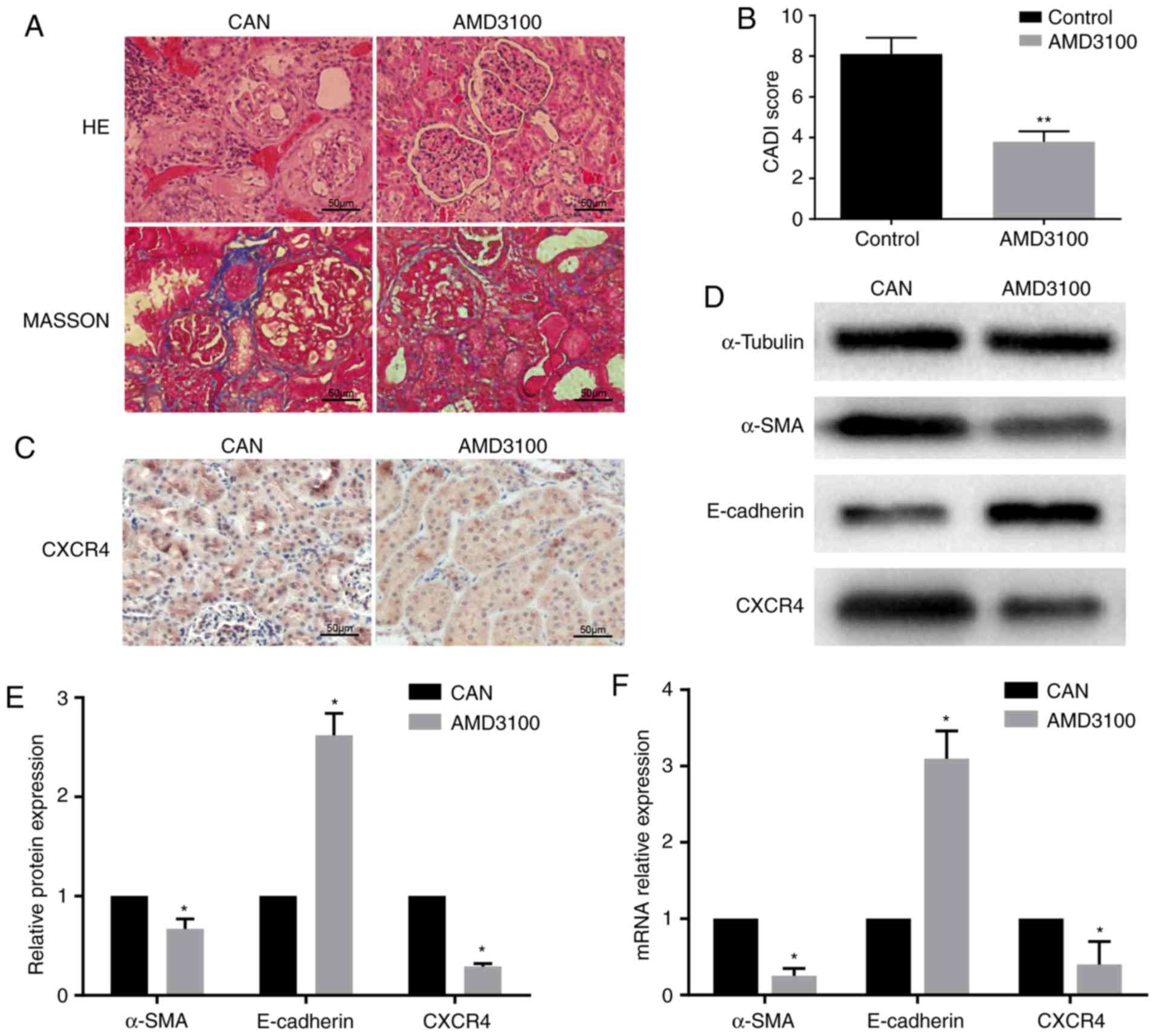
The histopathological changes and CADI scores
confirmed CAN in the rat kidney grafts at 12 weeks
post-transplantation. The AMD3100 group exhibited milder tubular
atrophy, inflammatory cell infiltration and interstitial fibrosis,
and also had a lower CADI score compared with the CAN group
(Fig. 1A and B). Subsequently, the
expression levels of a number of fibrosis-associated markers were
detected. Using immunohistochemistry, CXCR4 was observed in the
tubules in the CAN group, but it was expressed at lower levels in
the AMD3100 group (Fig. 1C).
RT-qPCR demonstrated that the expression of α-SMA was downregulated
and that of E-cadherin was upregulated in the AMD3100 group
compared with the CAN group, which was confirmed at the protein
level by WB (Fig. 1D-F). The
expression of CXCR4 was downregulated following administration of
AMD3100 (Fig. 1D and F). These
findings indicated the protective effect of AMD3100 in rat CAN by
inhibiting the SDF-1/CXCR4 axis, and the SDF-1/CXCR4 axis may serve
an important role in CAN.
Microarray and pathway analysis
In order to analyze alterations in the expression of
mRNAs and lncRNAs in the rat CAN model, the kidney allograft
tissues of the AMD3100 group and the CAN group were analyzed using
an mRNA + lncRNA microarray. The results of the microarray were
uploaded to the Gene Expression Omnibus database (GSE114088).
Clustering analysis revealed that 506 mRNAs and 404 lncRNAs were
significantly differentially expressed between the two groups,
including 81 mRNAs and 140 lncRNAs with an FC value >2.0
(Fig. 2A and B). KEGG pathway
analysis revealed that one of the top ranked pathways was the Wnt
signaling pathway (Fig. 2C), which
has been widely reported to be an important mechanism in EMT and
fibrosis (22,23). The results of the microarray also
predicted the association of a number of lncRNAs and mRNAs. CXXC5
mRNA was demonstrated to be negatively correlated with the lncRNAs
LOC100912353, LOC102551030 and LOC102556393, which, it may be
considered, may be a probable target for regulating renal allograft
fibrosis (Table II). The coding
potential of these lncRNAs is low, with the PhyloCSF (24) scores <-100 (Table II).
| Table II.Relevant lncRNAs. |
Table II.
Relevant lncRNAs.
| lncRNA gene
name | Chromosome | Strand | Start | End | GI | Length (bp) | NCBI RefSeq | NONCODE | PhyloCSF |
|---|
| LOC100912353 | 3 | + | 1732192 | 1733260 | 672046462 |
718 | XR_145884.3 | NONRATT018415 | −115.53 |
| LOC102551030 | 2 | + | 3404984 | 3412416 | 672042805 | 3190 | XR_591163.1 | – | −999.16 |
| LOC102556393 | X | + | 4623698 | 4626807 | 672087756 | 1937 | XR_597614.1 | – | −580.58 |
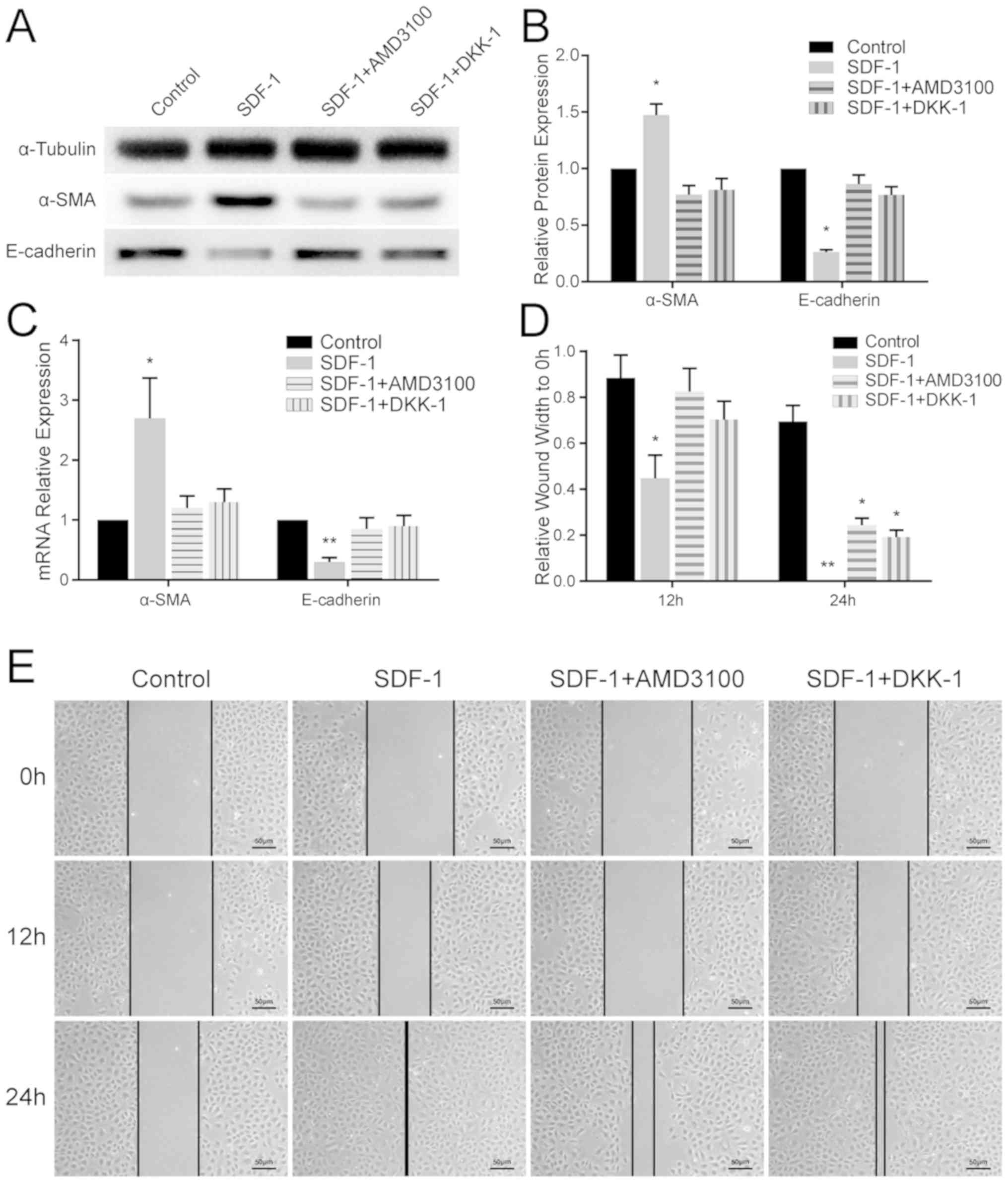
EMT is induced by SDF-1 and inhibited
by AMD3100 and DKK-1 in vitro
Variation in the expression levels of EMT-associated
markers in the different groups of NRK-52E cells was quantitated
using RT-qPCR and WB. In the SDF-1 group, α-SMA was upregulated and
E-cadherin was significantly downregulated (P<0.05) compared
with the other groups (Fig. 3A-C).
These results confirmed that EMT was induced by SDF-1 in renal
tubular epithelial cells, and this was antagonized by AMD3100 and
DKK-1.
Wound healing assay
The results of the wound healing assay revealed that
SDF-1 promoted cell migration across the wound edge into the
scratch area at 12 and 24 h compared with the other groups. In the
SDF-1 + AMD3100 group and SDF-1 + DKK-1 group, extensive cell
migration was not observed until 24 h post-treatment (Fig. 3D and E).
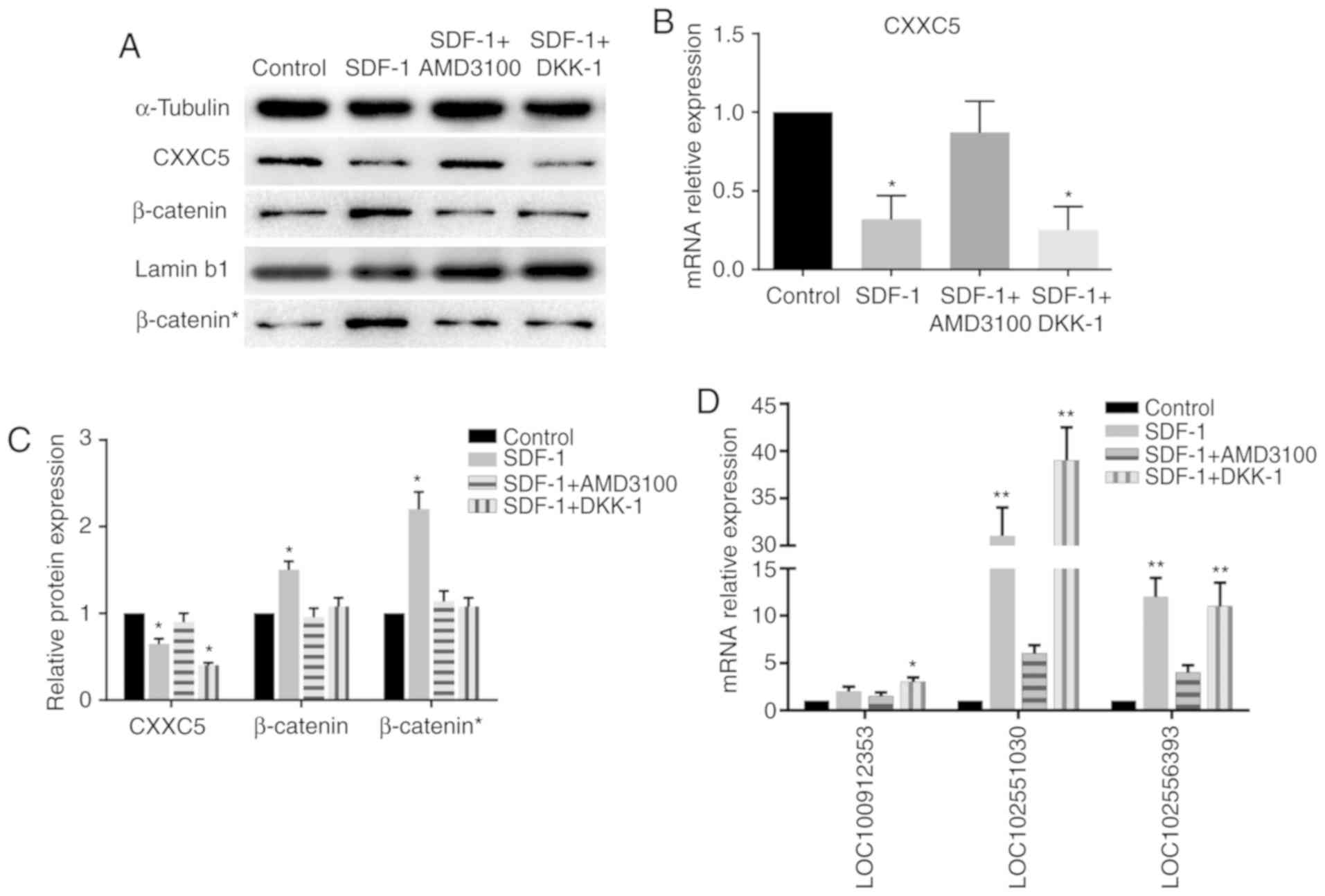
Markers of the Wnt/β-catenin
pathway
The protein expression levels of β-catenin in the
nucleus were increased significantly in the SDF-1 group, while the
increase in the levels of total β-catenin was not as significant
(Fig. 4), indicating the nuclear
accumulation of β-catenin. The WB and RT-qPCR results demonstrated
a reduction in the levels of CXXC5 expression in the SDF-1 group
compared with the control group; however, expression in the SDF-1 +
AMD3100 group was not significantly different to that in the
control group. Notably, CXXC5 was also downregulated in the SDF-1 +
DKK-1 group (Fig. 4A-C). These
results suggested that CXXC5 is involved in EMT induced by the
SDF-1/CXCR4 axis, and CXXC5 may be a potential upstream regulator
of the WNT/β-catenin pathway.
Involvement of lncRNAs
In order to detect the expression of the
aforementioned lncRNAs, RT-qPCR was performed. Two of these lncRNAs
were demonstrated to be significantly upregulated in the SDF-1
group and the SDF-1 + DKK-1 group, but not the SDF-1 + AMD3100
group, compared with the control group (Fig. 4D).
Discussion
The SDF-1/CXCR4 axis serves crucial roles in
promoting proliferation and metastasis (25). A previous study reported the
protective effect of AMD3100 in kidney injuries (26). A number of studies have
demonstrated that inhibition of the SDF-1/CXCR4 axis with AMD3100
leads to prolonged allograft survival, which is due to an influx of
host stem cells that results in a modulated host immune response
(9,10). A previous study revealed that the
progression of allograft fibrosis in a rat CAN model is attenuated
by AMD3100 (8). However, the
effect and mechanisms of SDF-1/CXCR4 on fibrosis in renal
allografts have rarely been reported.
In the present study, rats were treated with AMD3100
and it was demonstrated that the blockade of SDF-1/CXCR4 may
mediate the development of CAN. The results of the mRNA + lncRNA
microarray revealed that activation of the Wnt pathway may serve an
important role in the fibrosis of the CAN model. The in
vitro results demonstrated that SDF-1 promotes EMT in renal
tubular epithelial cells with the involvement of the Wnt signaling
pathway.
SDF-1/CXCR4 has been reported to serve an important
role in the progression of EMT in different types of cells
(13,17,27).
AMD3100 is a synthetic blocker that inhibits the binding of SDF-1
to CXCR4. The Wnt pathway has been widely reported to be associated
with fibrosis in different types of tissues (16,28).
β-catenin is the most potent member of the downstream Wnt pathway.
Activation of the Wnt pathway triggers intracellular signaling
cascades by recruiting segment polarity protein dishevelled homolog
DVL-1 (DVL-1) to the glycogen synthase kinase 3β complex, which
protects β-catenin from proteasomal degradation (29). Subsequently, β-catenin accumulates
in the cytoplasm and translocates into the nucleus, where it
stimulates the expression of numerous genes that are involved in
EMT (30). Hu et al
(17) demonstrated that
SDF-1/CXCR4 and the Wnt/β-catenin pathway have a synergistic effect
in the EMT of colorectal cancer cells via downregulation of
E-cadherin. E-cadherin, an exclusively expressed epithelial marker,
may bind to the cytoplasmic domain of β-catenin and prevent its
nuclear accumulation (31). In the
present study, EMT was observed in renal tubular epithelial cells
treated with SDF-1 through upregulation of α-SMA and downregulation
of E-cadherin, and it was inhibited by AMD3100. Hu et al
(17) also demonstrated that DKK-1
abolishes EMT by suppressing activation of the Wnt pathway. In the
present study, EMT was also inhibited by DKK-1, and the Wnt pathway
was demonstrated to be inactivated. Therefore, SDF-1 induces EMT in
renal tubular epithelial cells in vitro, and the Wnt pathway
may be one of the mechanisms involved.
The microarray in the present study revealed the
association between CXXC5 mRNA and three lncRNAs. Previous studies
have reported that CXXC5 is a negative regulator of the
Wnt/β-catenin pathway (32,33)
and that it interacts with DVL-1 (34–36).
In the present study, a negative association between CXXC5 and the
Wnt pathway was also observed in SDF-1-induced EMT. Notably, CXXC5
was downregulated in the cells treated with SDF-1 and DKK-1, and
the three lncRNAs were upregulated. These data suggested that CXXC5
and the three lncRNAs may be downstream of SDF-1/CXCR4 and regulate
activation of the Wnt pathway. Therefore, the present study may
provide novel targets for investigating the detailed mechanisms of
CAN and kidney fibrosis.
It is notable that the results of the present study
do not entirely explain kidney allograft fibrosis, and the exact
function of CXXC5 remains unknown. However, this study offers a
novel insight into the progression of CAN and may also aid tumor
research.
In conclusion, SDF-1 induces EMT by activating the
Wnt/β-catenin pathway in NRK-52E cells, and this involves CXXC5 and
three lncRNAs, which may be a novel mechanism and therapeutic
target in CAN.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81670679).
Availability of data and materials
The microarray data are available in the GEO
database (GSE114088), and other data are included in this published
article.
Authors' contributions
HT and YX contributed equally to this work,
performed the microarray, WB and RT-qPCR, and were major
contributors in writing the manuscript. ZZ performed the kidney
transplantation in the rats. SZ performed the histological
examination in the kidneys. WD and WJ assisted with analyzing the
experimental results. XH provided the idea of this study. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The animal protocols were approved by the Medical
Research Center, Beijing Chaoyang Hospital, Capital Medical
University (Beijing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Lamb KE, Lodhi S and Meier-Kriesche HU:
Long-term renal allograft survival in the United States: A critical
reappraisal. Am J Transplant. 11:450–462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gatault P, Bertrand D, Büchler M, Colosio
C, Hurault de Ligny B, Weestel PF, Rerolle JP, Thierry A, Sayegh J,
Moulin B, et al: Eight-year results of the Spiesser study, a
randomized trial comparing de novo sirolimus and cyclosporine in
renal transplantation. Transpl Int. 29:41–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Solez K, Colvin RB, Racusen LC, Sis B,
Halloran PF, Birk PE, Campbell PM, Cascalho M, Collins AB, Demetris
AJ, et al: Banff '05 Meeting Report: Differential diagnosis of
chronic allograft injury and elimination of chronic allograft
nephropathy (‘CAN’). Am J Transplant. 7:518–526. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hara S: Banff 2013 update: Pearls and
pitfalls in transplant renal pathology. Nephrology (Carlton). 20
(Suppl):S2–S8. 2015. View Article : Google Scholar
|
|
5
|
Mannon RB: Therapeutic targets in the
treatment of allograft fibrosis. Am J Transplant. 6:867–875. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Takabatake Y, Sugiyama T, Koharah,
Matsusaka T, Kurihara H, Koni PA, Nagasawa Y, Hamano T, Matsui I,
Kawada N, et al: The CXCL12 (SDF-1)/CXCR4 axis is essential for the
development of renal vasculature. J Am Soc Nephrol. 20:1714–1723.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ge G, Zhang H, Li R and Liu H: The
function of SDF-1-CXCR4 axis in SP cells-mediated protective role
for renal ischemia/reperfusion injury by SHH/GLI1-ABCG2 pathway.
Shock. 47:251–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu Y, Zhang Q, Xue W, Zeng S, Zhang Z,
Zhang X and Hu X: CXC chemokine receptor 4 (CXCR4) antagonist, a
novel pathway to prevent chronic allograft nephropathy. Ann
Transplant. 21:728–734. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu X, Okabayashi T, Cameron AM, Wang Y,
Hisada M, Li J, Raccusen LC, Zheng Q, Montgomery RA, Williams GM
and Sun Z: Chimeric allografts induced by short-term treatment with
stem cell-mobilizing agents result in long-term kidney transplant
survival without immunosuppression: A study in rats. Am J
Transplant. 16:2055–2065. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cameron AM, Wesson RN, Ahmadi AR, Singer
AL, Hu X, Okabayashi T, Wang Y, Shigoka M, Fu Y, Gao W, et al:
Chimeric allografts induced by short-term treatment with stem cell
mobilizing agents result in long-term kidney transplant survival
without immunosuppression: II, study in miniature swine. Am J
Transplant. 16:2066–2076. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zou XF, Gu JH, Cui ZL, Lu YW and Gu C: CXC
chemokine receptor type 4 antagonism ameliorated allograft fibrosis
in rat kidney transplant model. Exp Clin Transplant. 15:448–452.
2017.PubMed/NCBI
|
|
12
|
Aversa I, Zolea F, Ieranò C, Bulotta S,
Trotta AM, Faniello MC, De Marco C, Malanga D, Biamonte F,
Viglietto G, et al: Epithelial-to-mesenchymal transition in
FHC-silenced cells: The role of CXCR4/CXCL12 axis. J Exp Clin
Cancer Res. 36:1042017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He G, Ma M, Yang W, Wang H, Zhang Y and
Gao MQ: SDF-1 in mammary fibroblasts of bovine with mastitis
induces EMT and inflammatory response of epithelial cells. Int J
Biol Sci. 13:604–614. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu CH, Song JS, Chang KH, Jan JJ, Chen CT,
Chou MC, Yeh KC, Wong YC, Tseng CT, Wu SH, et al: Stem cell
mobilizers targeting chemokine receptor CXCR4: Renoprotective
application in acute kidney injury. J Med Chem. 58:2315–2325. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan A, Lee Y, Choi U, Moeckel G and
Karihaloo A: Chemokine receptor Cxcr4 contributes to kidney
fibrosis via multiple effectors. Am J Physiol Renal Physiol.
308:F459–F472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
von Toerne C, Schmidt C, Adams J, Kiss E,
Bedke J, Porubsky S, Gretz N, Lindenmeyer MT, Cohen CD, Gröne HJ
and Nelson PJ: Wnt pathway regulation in chronic renal allograft
damage. Am J Transplant. 9:2223–2239. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu TH, Yao Y, Yu S, Han LL, Wang WJ, Guo
H, Tian T, Ruan ZP, Kang XM, Wang J, et al: SDF-1/CXCR4 promotes
epithelial-mesenchymal transition and progression of colorectal
cancer by activation of the Wnt/beta-catenin signaling pathway.
Cancer Lett. 354:417–426. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shan S, Lv Q, Zhao Y, Liu C, Sun Y, Xi K,
Xiao J and Li C: Wnt/beta-catenin pathway is required for
epithelial to mesenchymal transition in CXCL12 over expressed
breast cancer cells. Int J Clin Exp Pathol. 8:12357–12367.
2015.PubMed/NCBI
|
|
19
|
Council NR, . Guide for the care and use
of laboratory animals. Eighth. The National Academies Press;
Washington, DC: 2011
|
|
20
|
Kahu J, Kyllonen L, Raisanen-Sokolowski A
and Salmela K: Donor risk score and baseline biopsy CADI value
predict kidney graft outcome. Clin Transplant. 25:E276–E283. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi J, Li F, Luo M, Wei J and Liu X:
Distinct roles of Wnt/beta-catenin signaling in the pathogenesis of
chronic obstructive pulmonary disease and idiopathic pulmonary
fibrosis. Mediators Inflamm. 2017:35205812017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sanchez-Duffhues G, Garcia de Vinuesa A
and Ten Dijke P: Endothelial-to-mesenchymal transition in
cardiovascular diseases: Developmental signaling pathways gone
awry. Dev Dyn. 247:492–508. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lin MF, Jungreis I and Kellis M: PhyloCSF:
A comparative genomics method to distinguish protein coding and
non-coding regions. Bioinformatics. 27:i275–i282. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun X, Cheng G, Hao M, Zheng J, Zhou X,
Zhang J, Taichman RS, Pienta KJ and Wang J: CXCL12/CXCR4/CXCR7
chemokine axis and cancer progression. Cancer Metastasis Rev.
29:709–722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zuk A, Gershenovich M, Ivanova Y,
MacFarland RT, Fricker SP and Ledbetter S: CXCR(4)antagonism as a
therapeutic approach to prevent acute kidney injury. Am J Physiol
Renal Physiol. 307:F783–F797. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xia R, Xu G, Huang Y, Sheng X, Xu X and Lu
H: Hesperidin suppresses the migration and invasion of non-small
cell lung cancer cells by inhibiting the SDF-1/CXCR-4 pathway. Life
Sci. 201:111–120. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Taoh, Yang JJ, Shi KH and Li J: Wnt
signaling pathway in cardiac fibrosis: New insights and directions.
Metabolism. 65:30–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Luo K, Gu X, Liu J, Zeng G, Peng L, Huang
H, Jiang M, Yang P, Li M, Yang Y, et al: Inhibition of disheveled-2
resensitizes cisplatin-resistant lung cancer cells through
down-regulating Wnt/beta-catenin signaling. Exp Cell Res.
347:105–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gilles C, Polette M, Mestdagt M,
Nawrocki-Raby B, Ruggeri P, Birembaut P and Foidart JM:
Transactivation of vimentin by beta-catenin in human breast cancer
cells. Cancer Res. 63:2658–2664. 2003.PubMed/NCBI
|
|
31
|
Wong SHM, Fang CM, Chuah LH, Leong CO and
Ngai SC: E-cadherin: Its dysregulation in carcinogenesis and
clinical implications. Crit Rev Oncol Hematol. 121:11–22. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Andersson T, Sodersten E, Duckworth JK,
Cascante A, Fritz N, Sacchetti P, Cervenka I, Bryja V and Hermanson
O: CXXC5 is a novel BMP4-regulated modulator of Wnt signaling in
neural stem cells. J Biol Chem. 284:3672–3681. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim HY, Yoon JY, Yun JH, Cho KW, Lee SH,
Rhee YM, Jung HS, Lim HJ, Lee H, Choi J, et al: CXXC5 is a
negative-feedback regulator of the Wnt/beta-catenin pathway
involved in osteoblast differentiation. Cell Death Differ.
22:912–920. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee SH, Kim MY, Kim HY, Lee YM, Kim H, Nam
KA, Roh MR, Min Do S, Chung KY and Choi KY: The dishevelled-binding
protein CXXC5 negatively regulates cutaneous wound healing. J Exp
Med. 212:1061–1080. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee SH, Seo SH, Lee DH, Pi LQ, Lee WS and
Choi KY: Targeting of CXXC5 by a competing peptide stimulates hair
regrowth and wound-induced hair neogenesis. J Invest Dermatol.
137:2260–2269. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma S, Choi J, Jin X, Kim HY, Yun JH, Lee
W, Choi KY and No KT: Discovery of a small-molecule inhibitor of
Dvl-CXXC5 interaction by computational approaches. J Comput Aided
Mol Des. 32:643–655. 2018. View Article : Google Scholar : PubMed/NCBI
|