Introduction
Isoflurane is one of the common inhalational
anesthetics applied in the clinic (1). The influence of isoflurane on infant
neural development has attracted great attention from doctors and
patients. Research has indicated that isoflurane may affect the
development of the central nervous system (2). In addition, it may cause
post-developmental cognitive decline in infants and young animals
(2). Additionally, it may damage
the activity and function of hippocampal neural stem cells within
newborn rats. As a result, isoflurane may promote neuronal
apoptosis, affecting neural systemic development, leading to
individual cognitive dysfunction (3). However, the precise molecular
mechanism by which isoflurane acts on neural stem cells and affects
neural development remains unclear.
The oxidative stress theory serves a vital role in
the mechanism of neurotoxicity (4), and has attracted extensive attention
in recent years (4). Oxidative
stress refers to cytotoxic effects generated by reactive oxygen
species (ROS), including superoxide anion free radical
(O-2), hydroxyl radical (OH) and hydrogen peroxide
(H2O2) (4).
These ROS are produced as by-products using oxygen molecules during
normal and abnormal metabolic processes. Oxidative stress serves an
important role in foreign compound-induced neurotoxic effects. In
addition, it may inhibit the activities of brain catalase (CAT),
superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px).
Finally, it leads to decreased brain antioxidant function (5).
Oxidative stress is the stress damage state induced
by the imbalance between cellular oxidation and antioxidants
(6). Oxidative stress may
stimulate cells to produce a large amount of active oxygen radicals
(6). This can directly injure the
neurons and lead to cytomembrane lipid peroxidation and increased
membrane permeability (7).
Finally, it may lead to neuronal swelling, degeneration and loss of
function (7). Oxidative stress is
involved in the early initiation of the neuron apoptotic signaling
pathway (8). In addition, it
serves a vital role in the genesis and development of
neurodegenerative disease (8).
Therefore, understanding and research on oxidative stress is of
great value in terms of theory and application.
In recent years, the role of the oxidative stress
response in the genesis and development of diabetes has attracted
attention (9). The close
association between these two has been extensively verified
(9). The nuclear erythroid-2
related factor 2 (Nrf2)/antioxidant response element (ARE)
signaling pathway is the most important endogenous antioxidative
stress pathway in the human body. Its association with diabetes is
of great interest (10). Nrf2
belongs to the leucine zipper activating transcription factor
family and is a vital nuclear transcription factor regulating
antioxidative stress (11).
Additionally, it serves a central regulatory role in cellular
anti-oxidative process (9).
It has been verified that over 200 endogenous genes
are regulated by the Nrf2/ARE signaling pathway (10). They include heme oxygenase-1
(HO-1), SOD, CAT, quinone reductase 1, glutathione-s-transferase
(GST), glutamate-cysteine ligase and epoxide hydrolase (11). The expression products of these
activated genes serve important roles in numerous processes. They
include antioxidative stress, the anti-inflammatory response,
antiapoptosis, neurotoxicity, immunoregulation and detoxification
processes (11). In addition, they
can act against the progression of numerous pathological processes.
Regulating the activity of such pathway exerts multiple actions in
anti-atherosclerosis, antitumor, anti-aging, as well as heart,
liver, microvascular and neural protection. Zhang et al
(12) demonstrated that Tanshinone
IIA protects against 6-hydroxydopamine-induced neurotoxicity via
the miR-153-mediated Nrf2/ARE signaling pathway. The present study
aimed to investigate the function of microRNA (miR/miRNA)-153
against isoflurane-induced neurotoxicity and the underlying
mechanism.
Materials and methods
Mouse model
Wild-type C57BL/6J male mice (n=12; aged 5–6 weeks
old; weight, 18–20 g) were purchased from Laboratory Animal Center
of Suzhou University (Suzhou, China), and housed in a controlled
environment (20–22°C; 55–60% humidity, 12-h light/dark on a
reversed light cycle, free access to food and water) and randomly
assigned to the isoflurane-anesthesia group (n=6) or the control
group (n=6). In the control group, mice received an intraperitoneal
injection of 100 µl normal saline. In the isoflurane-anesthesia
group, mice received 1.4% isoflurane and 100% oxygen for 2 h using
identical flow rates (2 l/min, Ohmeda; GE Healthcare, Chicago, IL,
USA) as described in the literature (13). After 2 h, mice from both groups
were sacrificed and employed for further analysis. The present
study was approved by the ethics committee of the Changzhou No. 2
People's Hospital Affiliated to Nanjing Medical University
(Chanzhou, China).
Hematoxylin and eosin staining
Hippocampal tissues were isolated and then washed
with PBS and then fixed using 4% paraformaldehyde for 24 h at room
temperature. Following this tissues were cut and embedded in
paraffin (5 µM). Sections were then stained using 0.1% hematoxylin
and eosin stain for 10 min at room temperature, and subsequently
visualized using an epifluorescence microscope (magnification,
×100; Zeiss Axiovert 200M; Carl Zeiss AG, Oberkochen, Germany) and
analyzed using Image Lab 3.0 software (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Cell culture and transient
transfection
Pc12 cells were purchased from the Cell Bank of Type
Culture Collection of Chinese Academy of Sciences (Shanghai, China)
and cultured in high-glucose Dulbecco's modified Eagle's medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% heat-inactivated fetal calf serum (Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin, 100 µg/ml
streptomycin and 2 mM L-glutamine at 37°C in a 5% CO2.
miRNA-153 (5′-UUGCAUAGUCACAAAAGUGAUC-3′), anti-miRNA-153
(5′-UUGCAUAGUCACAAAAGUGAUC-3′) and negative mimics
(5′-CCCCCCCCCCCCCCCCCCCCCC-3′) were purchased from Sangon Biotech
Co., Ltd. (Shanghai, China). miRNA-153 (100 ng), anti-miRNA-153
(100 ng) and negative mimics (100 ng) were transfected into Pc12
cells (1×106 cells/ml) using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.). Following
transfection at 4 h, the old medium was removed and cells were
treated with 2% isoflurane plus 21% O2 and 5%
CO2 for 6 h as described previously (13). A total of 4 h post-transfection,
the Nrf2 agonist dimethyl fumarate (2.5 µM) was incubated with the
cells for 44 h at 37°C, and the cells were treated with 2%
isoflurane, 21% O2 and 5% CO2 for 6 h at
37°C.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis of cellular miRNA and
mRNA levels
Total RNA was extracted from tissue samples and cell
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Total RNA was reverse-transcribed into cDNA using a TaqMan MicroRNA
Reverse Transcription kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) at 37°C for 60 min and 85°C for 1 min. RT-qPCR
was conducted using a Maxima SYBR Green PCR kit (Thermo Fisher
Scientific Inc.). The following thermocycling conditions were used
for qPCR: 95°C for 10 min; followed by 40 cycles at 95°C for 15
sec, 60°C for 30 sec and 72°C for 30 sec. The 2−ΔΔCq
method was used for quantification (14). The following primers were used:
miRNA-153 forward,
5′-TCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGATCACTTT-3′ and reverse,
5′-ACACTCCAGCTGGGTTGCATAGTCACAAAAGT-3′; U6 forward,
5′-CGCTTCACGAATTTGCGTGTCAT-3′ and reverse,
5′-GCTTCGGCAGCACATATACTAAAAT-3′.
ELISA assay
Extracts from harvested cells were subjected to
western blot analyses using radioimmunoprecipitation assay (RIPA)
buffer (Beyotime Institute of Biotechnology, Haimen, China) and
protein content was measured using a Bicinchoninic Acid (BCA)
protein assay (Beyotime Institute of Biotechnology). A total of 10
µg proteins were used to measure SOD (cat. no. A001-1-1), CAT (cat.
no. A007-1-1), MPO (cat. no. A044) and malondialdehyde (MDA; cat.
no. A003-1) levels using ELISA kits according to the manufacturer's
protocol (Nanjing Jiancheng Bioengineering Institute, Nanjing,
China).
MTT assay
Following isoflurane-induced neurotoxicity at 24, 48
and 72 h, cells were stained with 20 µl volume of 5 mg/ml MTT dye
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and incubated for 4
h at 37°C. Subsequently, 150 µl dimethylsulfoxide (Invitrogen;
Thermo Fisher Scientific, Inc.) was added to each cell and agitated
for 20 min at 37°C. The optical density was determined at 490 nm
using a Versamax microplate reader (Molecular Devices, LLC,
Sunnyvale, CA, USA).
Flow cytometric detection of
apoptosis
Following isoflurane-induced neurotoxicity at 48 h,
cell were washed twice with ice-cold PBS prior to being harvested
and stained with 5 µl Annexin V-fluorescein isothiocyanate and 5 µl
propidium iodide (cat. no. 550911; BD Biosciences, Franklin Lakes,
NJ, USA) for 15 min per treatment at room temperature in darkness.
Apoptosis was analyzed using a flow cytometer (BD C6 flow
cytometer; BD Biosciences) and FlowJo software (version 7.6.1;
FlowJo, LLC, Ashland, OR, USA).
DAPI assay
Cells were washed with PBS and fixed with 4%
paraformaldehyde for 15 min at room temperature. Cells were blocked
with 5% bovine serum albumin (Beyotime Institute of Biotechnology)
in PBS for 1 h at room temperature and then incubated with DAPI (5
mg/ml; Beyotime Institute of Biotechnology) for 30 min in the dark
at room temperature. Cells were washed with PBS and subsequently
visualized using an epifluorescence microscope (magnification,
×100; Zeiss Axiovert 200M; Carl Zeiss AG).
Western blotting
Extracts from harvested cells were subjected to
western blot analyses using RIPA buffer; protein content was
measured using a BCA assay. A total of 30–50 µg protein were
separated and concentrated using an 8–12% SDS-PAGE gel, and then
electrotransferred onto polyvinylidene fluoride (PVDF) membranes
(Merck KGaA). PVDF membranes were blocked with 5%-non-milk in TBS
with 0.1% Tween 20 for 1 h at 37°C and incubated with B-cell
lymphoma-2 (Bcl-2)-associated X (Bax; cat. no. sc-20067; 1:500;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA), Nrf2 (cat. no.
sc-722; 1:500; Santa Cruz Biotechnology, Inc.), HO-1 (cat. no.
sc-10789; 1:500; Santa Cruz Biotechnology, Inc.) and GAPDH (cat.
no. 5174; 1:5,000; Cell Signaling Technology, Inc., Danvers, MA,
USA) at 4°C overnight. PVDF membranes were incubated with an
anti-rabbit IgG (H+L) secondary antibody (cat. no. 14708; 1:5,000;
Cell Signaling Technology, Inc.) then detected by using enzymatic
chemiluminescence kit (Beyotime Institute of Biotechnology) and
analyzed using Image Lab 3.0 software (Bio-Rad Laboratories,
Inc.).
Determination of Caspase-3/9
activities
The activity levels of Caspase-3/9 proteins were
analyzed using 10 µg protein obtained from extracted cells and
Caspase-3/9 activities kits (cat nos. C1115 and C1157,
respectively; Beyotime Institute of Biotechnology). Optical
densities were determined at 405 nm using a Versamax microplate
reader (Molecular Devices, LLC, Sunnyvale, CA, USA).
Statistical analysis
Data were expressed as the mean ± standard deviation
using SPSS 19.0 (IBM Corp., Armonk, NY, USA). The Student's t-test
was used to analyze the differences between two groups; one-way
analysis of variance followed by Tukey's post hoc test was used to
analyze the differences between multiple groups. P<0.05 was
considered to indicate a statistically significant difference. All
experiments were repeated in triplicate.
Results
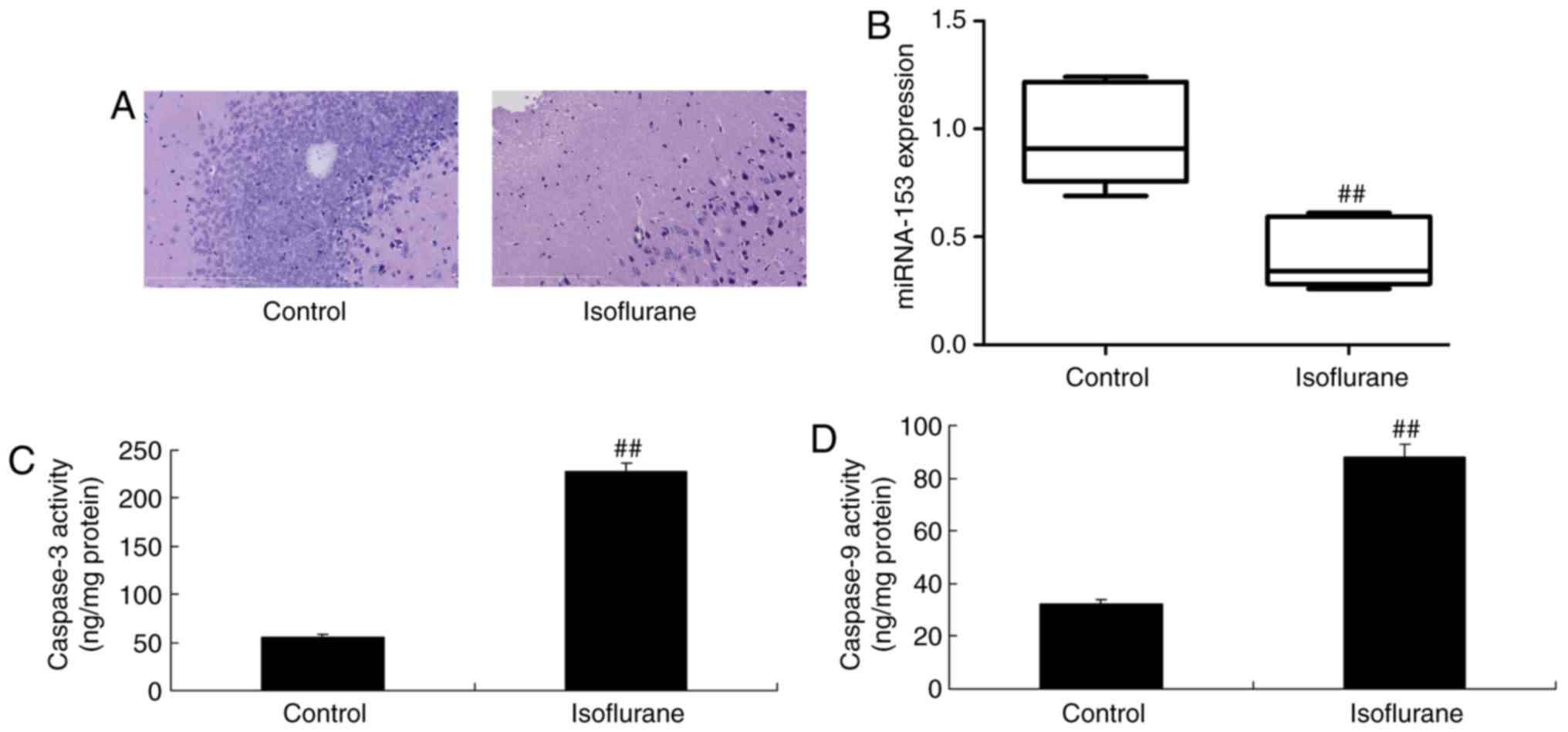
miRNA-153 expression
Hematoxylin and eosin staining of hippocampal
sections indicated the inhibition of neurocytes in
isoflurane-induced mice, compared with in the control group
(Fig. 1A). As present in Fig. 1B, qPCR demonstrated that the
miRNA-153 expression was significantly downregulated within
isoflurane-induced mice, compared with in the control group
(P<0.01). Additionally, significant increases in caspase-3/9
activities were observed within isoflurane-induced mice, compared
with in the control group (P<0.01; Fig. 1C and D). These results demonstrated
that miRNA-153 may participate in isoflurane-induced neurocyte
apoptosis.
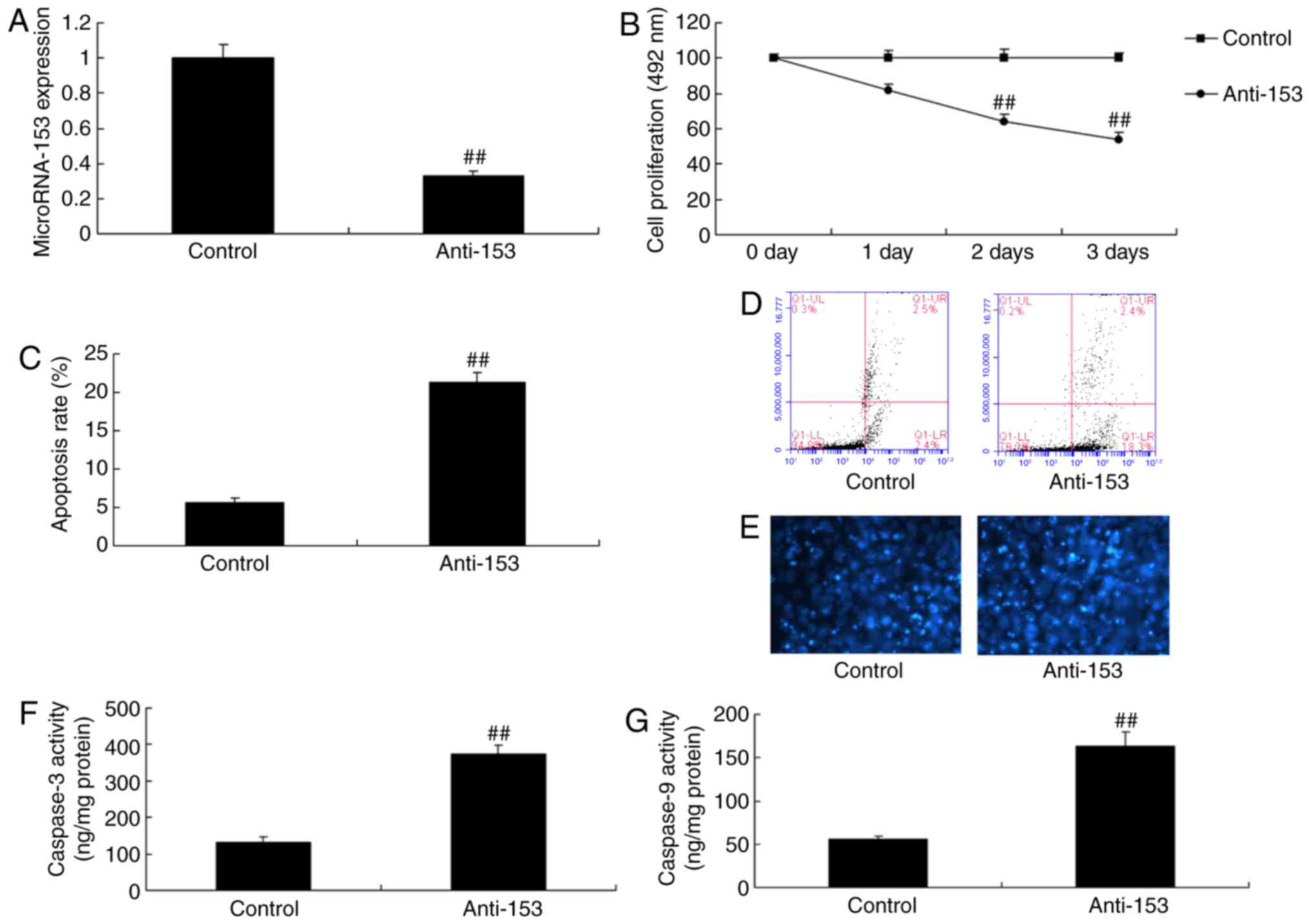
Downregulation of miRNA-153 induces
neurocyte apoptosis in vitro
miRNA-153 expression was reduced in vitro
using anti-miRNA-153 expression mimics. As presented in Fig. 2A, a significant inhibition of
miRNA-153 expression within the in vitro model was observed,
compared with in the control group (P<0.01). Subsequently, the
downregulation of miRNA-153 induced neurocyte apoptosis and reduced
cell growth in vitro, compared with in the control group
(P<0.01; Fig. 2B-D).
Downregulation of miRNA-153 significantly increased DAPI and
caspase-3 and caspase-9 activity in vitro, compared with in
the control group (Fig. 2E-G).
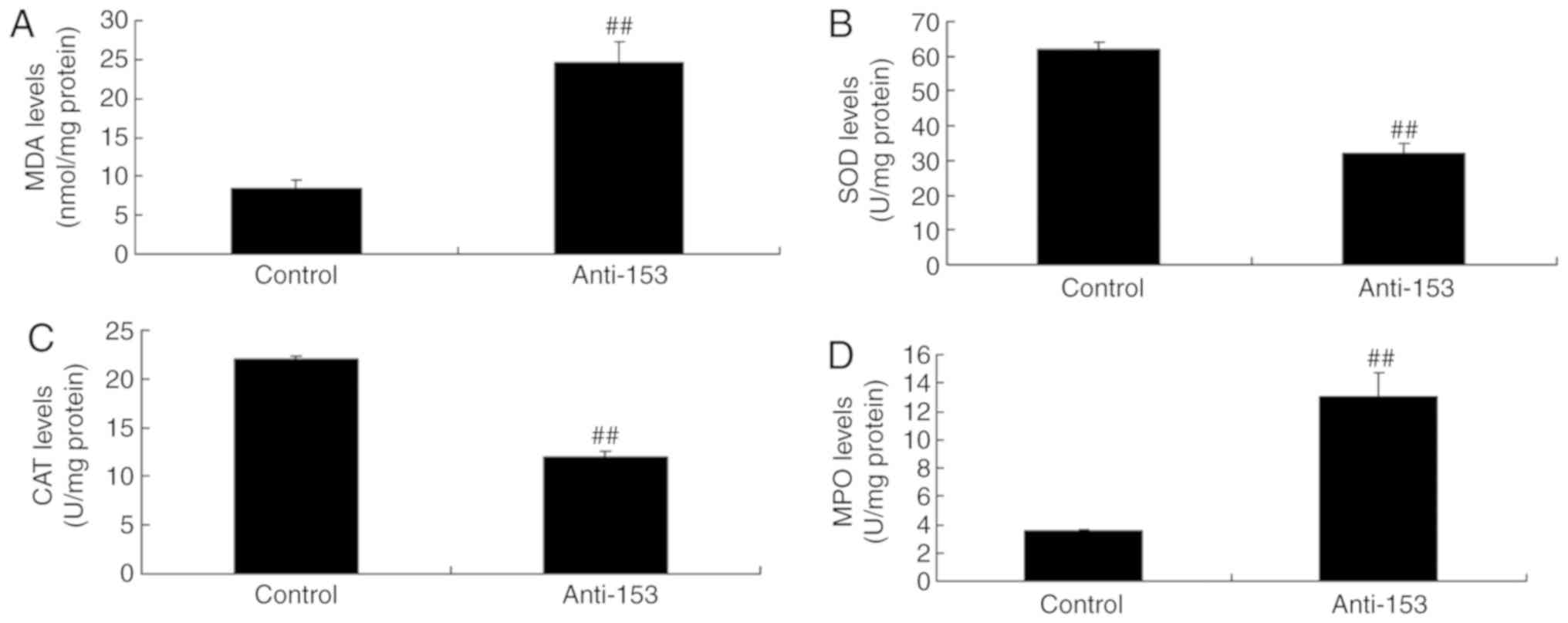
Downregulation of miRNA-153 induces
oxidative stress in vitro
Downregulation of miRNA-153 induced neurocyte
apoptosis in vitro; the effects of miRNA-153 on oxidative
stress were investigated in an in vitro model. As presented
in Fig. 3, the downregulation of
miRNA-153 significantly reduced SOD and CAT expression levels, and
induced MDA and MPO expression levels in an in vitro model
of isoflurane-induced neurotoxicity, compared with in the control
group (P<0.01).
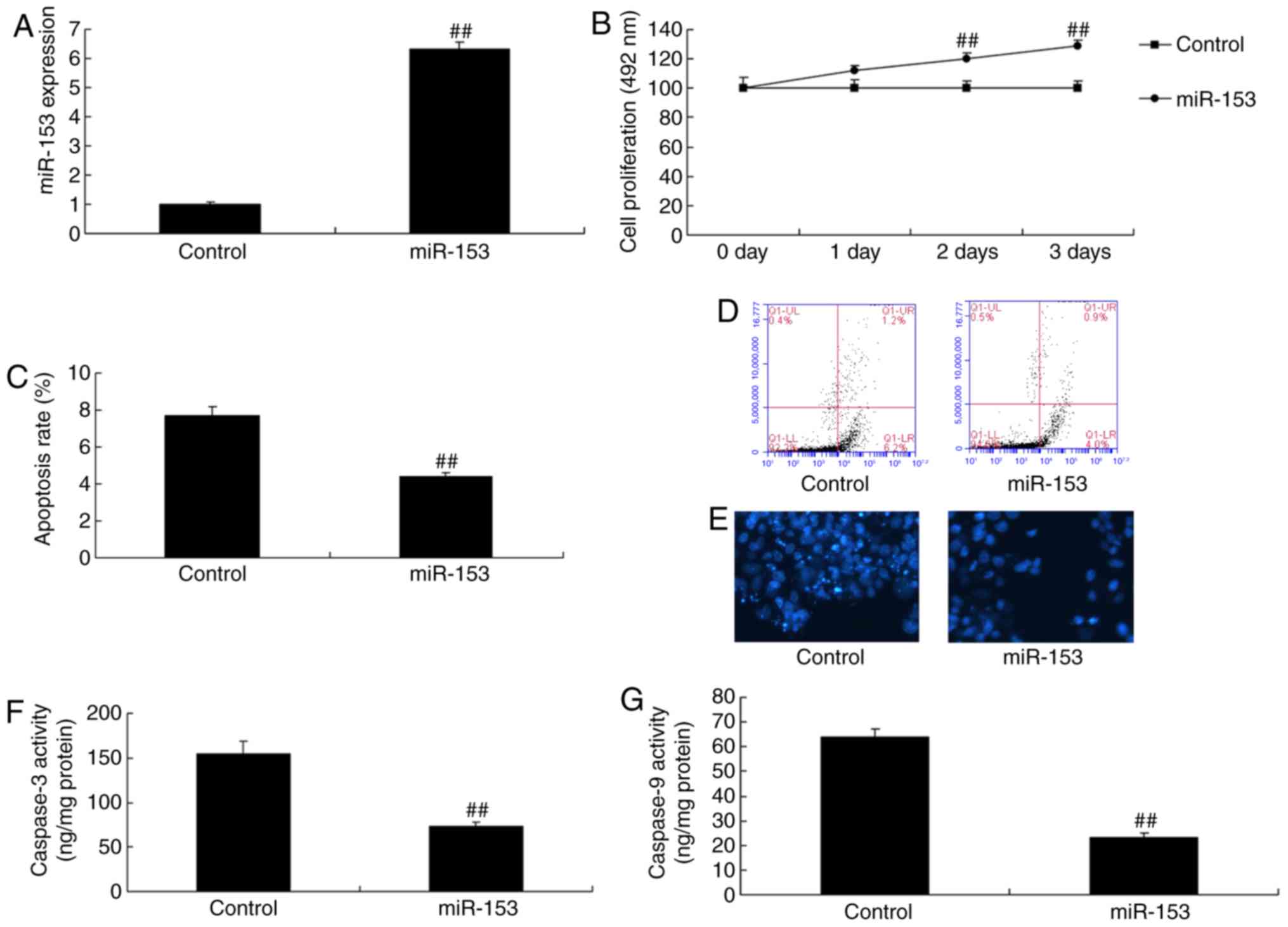
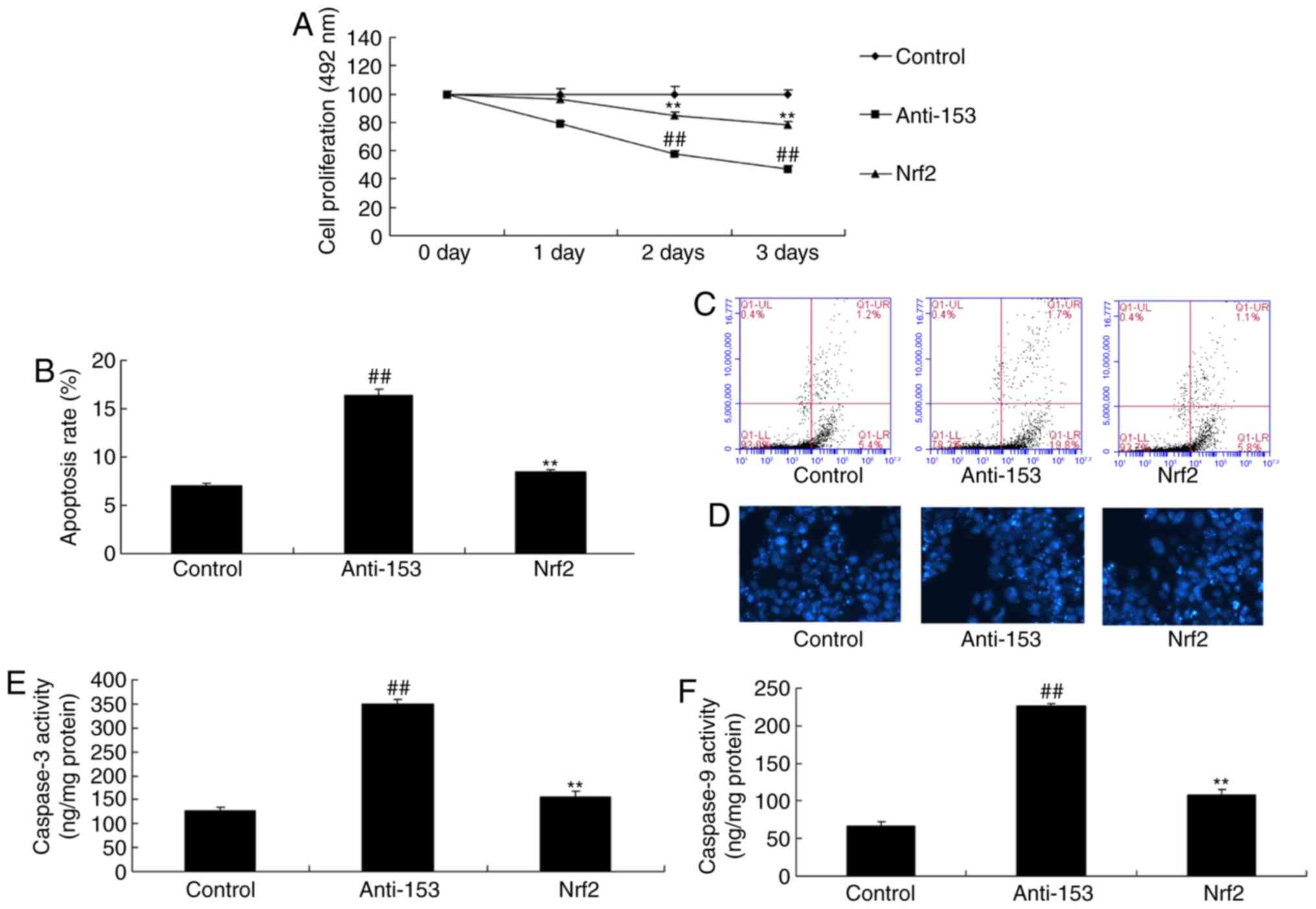
Overexpression of miRNA-153 reduces
neurocyte apoptosis in the in vitro model
miRNA-153 mimics significantly increased the
expression of miRNA-153 in vitro, compared with in the
control group (P<0.01; Fig.
4A). Overexpression of miRNA-153 significantly reduced
neurocyte apoptosis and promoted cell growth in the in vitro
model, compared with in the control group (P<0.01; Fig. 4B-E). Overexpression of miRNA-153
significantly suppressed caspase-3 and caspase-9 activity in an
in vitro model of isoflurane-induced neurotoxicity, compared
with in the control group (P<0.01; Fig. 4F and G).
Overexpression of miRNA-153 reduces
oxidative stress in an in vitro model
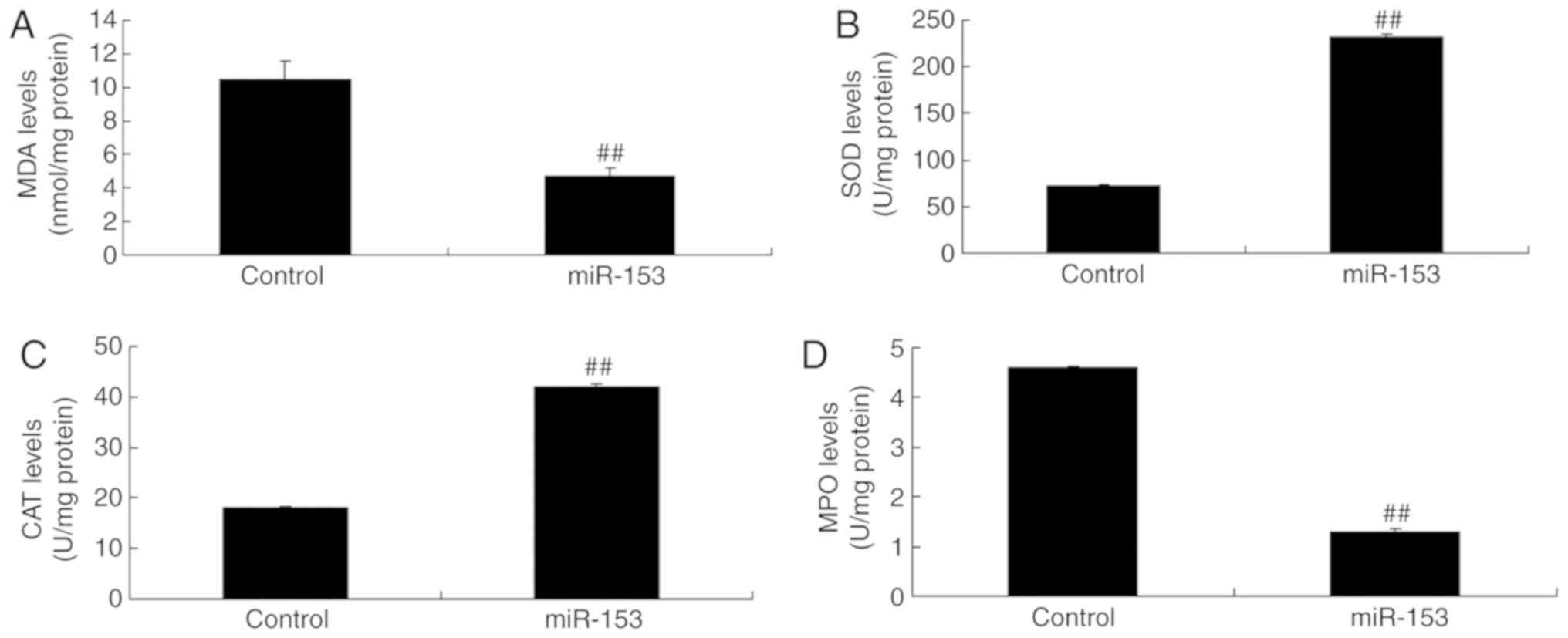
Overexpression of miRNA-153 increased SOD and CAT
expression levels, and reduced MDA and MPO expression levels in an
in vitro model of isoflurane-induced neurotoxicity, compared
with in the control group (Fig.
5). These results demonstrated that miRNA-153 regulated
oxidative stress to induce apoptosis in an in vitro model of
isoflurane-induced neurotoxicity.
Regulation of miRNA-153 affects
Nrf2/ARE signaling pathway in an in vitro model
In order to investigate the mechanisms of miRNA-153
on oxidative stress in vitro, the alterations of Nrf2/ARE
signaling pathway were analyzed. As presented in Fig 6A-D, the downregulation of miRNA-153
suppressed Nrf2/HO-1 protein expression and induced Bax protein
expression in an in vitro model of isoflurane-induced
neurotoxicity, compared with in the control group. Overexpression
of miRNA-153 induced Nrf2/HO-1 protein expression and significantly
suppressed Bax protein expression in an in vitro model of
isoflurane-induced neurotoxicity, compared with in the control
group (P<0.01; Fig. 6E-H).
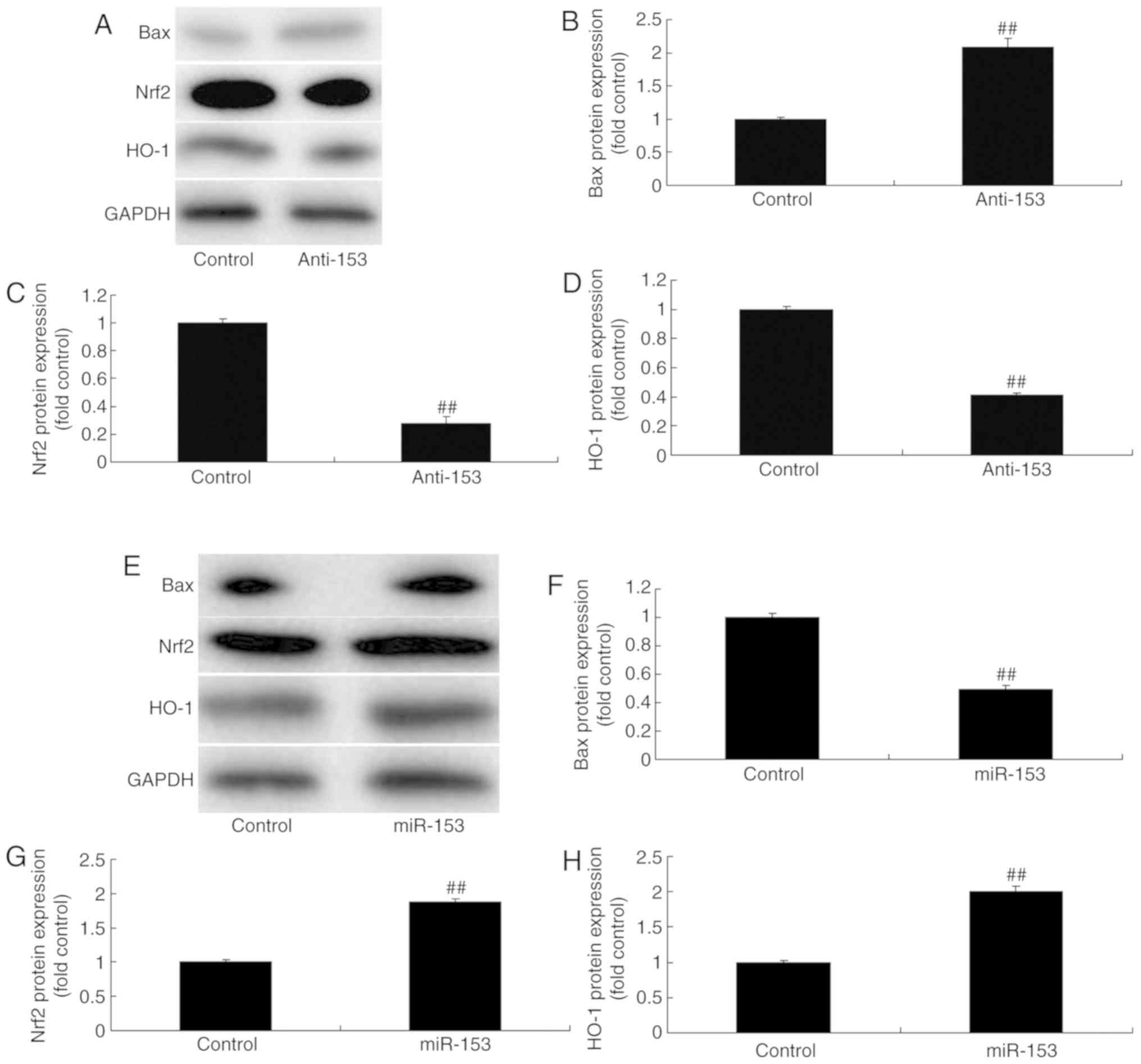 | Figure 6.Regulation of miRNA-153 affects the
Nrf2/HO-1 signaling pathway in an in vitro model. (A) Bax,
Nrf2 and ARE expression levels were detected by western blotting
and statistical analysis of (B) Bax, (C) Nrf2 and (D) HO-1
expression in the control and the miRNA-153-downregulated group.
(E) Bax, Nrf2 and ARE expression levels detected by western
blotting and of (F) Bax, (G) Nrf2 and (H) ARE expression in the
control and the miRNA-153-downregulated group. Data are expressed
as the mean ± standard deviation (n=3). ##P<0.01 vs.
the control group. Anti-153, miRNA-153 downregulation group; ARE,
antioxidant response element; Bax, B-cell lymphoma-2-associated X;
miR-153, miRNA-153 overexpression group; miR, microRNA; Nrf2,
nuclear erythroid-2 related factor 2; HO-1, heme oxygenase-1. |
Nrf2 agonist induces the Nrf2/ARE
signaling pathway and reduces oxidative stress to induce neurocyte
apoptosis in vitro following anti-miRNA-153 transfection
The role of the Nrf2/ARE signaling pathway on the
effects of anti-miRNA-153 were further investigated in an in
vitro model of isoflurane-induced neurotoxicity. The Nrf2
agonist significantly induced Nrf2 and HO-1 protein expression, and
suppressed Bax protein expression in cells transfected with
anti-miRNA-153, compared with the anti-miRNA-153 group (Fig. 7A-D). The Nrf2 agonist increased SOD
and CAT expression levels, and reduced MDA and MPO expression
levels an in vitro model of isoflurane-induced neurotoxicity
following anti-miRNA-153 transfection, compared with the
anti-miRNA-153 group (Fig. 7E-H).
Compared with in the anti-miRNA-153 group, the Nrf2 agonist
inhibited the effects of anti-miRNA-153 on the promotion of
neurocyte apoptosis, inhibition of cell growth and induction of
caspase-3/9 activity in an in vitro model of
isoflurane-induced neurotoxicity following anti-miRNA-153, compared
with in the anti-miRNA-153 group (Fig.
8).
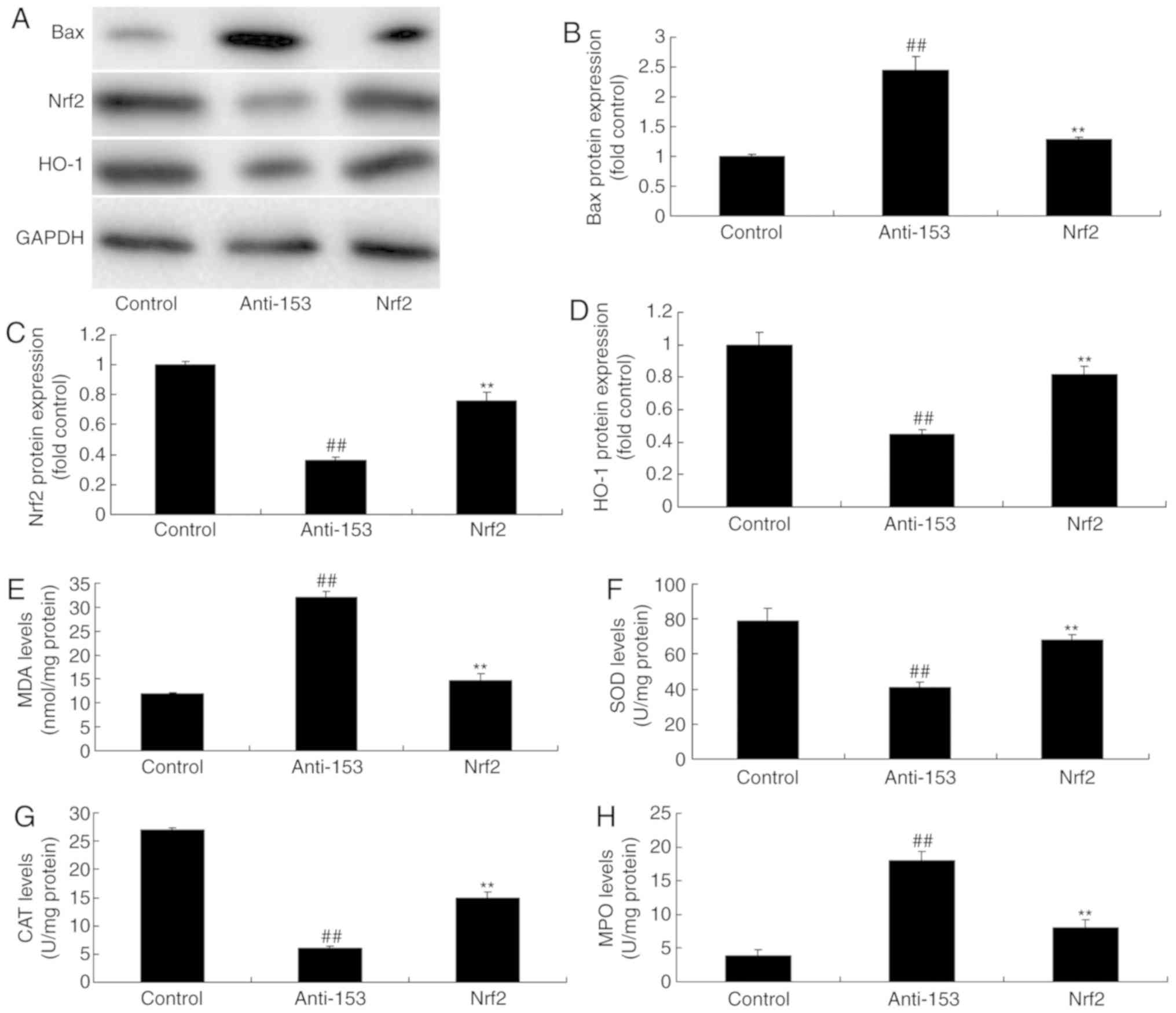 | Figure 7.Nrf2 agonist induces Nrf2/HO-1
signaling pathway and reduces oxidative stress in an in
vitro model following anti-miRNA-153 treatment. (A) Bax, Nrf2
and HO-1 expression levels detected by western blotting and
statistical analysis of (B) Bax, (C) Nrf2 and (D) HO-1 expression.
(E) MDA, (F) SOD, (G) CAT and (H) MPO expression levels. Data are
expressed as the mean ± standard deviation (n=3).
##P<0.01 vs. the control group, **P<0.01 compared
with anti-153 group. Anti-153, miRNA-153-downregulated group; ARE,
antioxidant response element; Bax, B-cell lymphoma-2-associated X;
CAT, catalase; MDA, malondialdehyde; miR, microRNA; MPO,
myeloperoxidase; Nrf2, nuclear erythroid-2 related factor 2; Nrf2
agonist + miRNA-153-downregulated group; SOD, superoxide dismutase;
HO-1, heme oxygenase-1. |
Discussion
Research has indicated that acute inhalation
anesthetic exposure to isoflurane may result in hippocampal neuron
apoptosis in developmental rats (3). Furthermore, it may also lead to
reduced learning and memory ability (15); however, evidence has been
contradictory (15). Nonetheless,
studies indicate that anesthetics exposure in postnatal animals
during the sensitive neural developmental period will lead to
neuron histopathological injury (15). Additionally, apoptosis has been
observed in all parts of the brain, which may result in subsequent
decreases in learning and memory ability (15).
Numerous processes are involved in the development
of the mammalian nervous system (16), including neurogenesis,
differentiation, migration, synaptogenesis, myelinogenesis and
neuron apoptosis (16). Apoptosis
can remove the excessive neurons during development, but is rarely
observed under normal conditions (16). Apoptosis can be achieved via the
endogenous and exogenous pathways and induces the activation of
caspases. Activation of the endogenous pathway includes
downregulation of Bcl-2 family proteins expression and enhanced
mitochondrial permeability. Cytochrome c outflow, as well as
the activation of caspase 9 and caspase 3 are also involved
(17). Activation of the exogenous
pathway includes the formation of the apoptosis induction signal
peptide complex, as well as the activation of caspase 8 and caspase
3 (17). Therefore, caspase 3 is
the common factor of endogenous and exogenous apoptosis pathways.
In the present study the downregulation of miRNA-153 induced
neurocyte apoptosis and reduced cell growth in an in vitro
model of isoflurane-induced neurotoxicity via Bax-caspase-3/9
activity. Zou et al (18)
demonstrated that miR-153 regulates apoptosis and autophagy of
cardiomyocytes by targeting Mcl-1.
Free radicals serve a critical role in cerebral
ischemia, cerebral injury, Parkinson's disease, amyotrophic lateral
sclerosis, Down's syndrome and Alzheimer's disease (19). The nervous system is richer in
lipid composition compared with other tissues (20) and has high oxygen consumption
(20). Therefore, it is sensitive
to free radical injury. Excessive oxidative injury may affect
subsequent life processes (20).
SOD, CAT and GSH-Px are the important enzymes in the antioxidant
system (4). Their activities may
directly reflect the levels of mitochondrial antioxidant system
function (20). Excessive
manganese inhibition can weaken the action of SOD on superoxide
anion. This may result in the accumulation of free radicals in the
body (20). Additionally,
catecholamine reactions, including excessive free radical catalysis
or accelerated dopamine may increase free radical formation and may
accelerate lipid peroxidation and induce neuron degeneration and
necrosis (21).
Nrf2 is an important antioxidative stress
transcription factor; HO-1 is a vital antioxidant enzyme against
endogenous and exogenous stimulation and can protect cells
(10). Nrf2 will translocate into
the nucleus following activation. In this manner, Nrf2 may bind
with the ARE. Therefore, it may initiate HO-1 expression and a
series of antioxidation mechanisms (22,23).
In the present study, the overexpression of miRNA-153 induced the
Nrf2/ARE signaling pathway within a model of isoflurane-induced
neurotoxicity, compared with in the control group. Wang et
al (24) demonstrated that
miRNA-153 regulates NRF2 expression, decreases apoptosis and
increases the colony formation ability of breast epithelial
cells.
Nrf2 is an important transcription factor regulating
the antioxidative stress response; Kelch-like ECH-associated
protein1 (Keap1) is its specific receptor (25). Under normal conditions, Keap1 and
Nrf2 will form a complex in the cytoplasm to inhibit Nrf2 activity.
Under oxidative stress, Keap1 will dissociate with Nrf2 (25). Subsequently, Nrf2 may bind with ARE
following nuclear translocation (26). This will initiate the expression of
ARE-regulated phase II detoxifying enzymes and antioxidase genes.
Finally, it will enhance cellular resistance to oxidative stress
and nucleophilic compounds. ARE-regulated antioxidant genes include
HO-1 and GST, which can protect the body from ROS. Investigation
has verified that a Nrf2 knockout mouse exhibited aggravated
cerebral ischemic injury. In addition, activating the
Keap1/Nrf2/ARE signaling pathway and promoting HO-1 expression may
alleviate neurotoxicity (23).
Zhang et al (12) suggested
that Tanshinone IIA may protect dopaminergic neurons against
6-hydroxydopamine-induced neurotoxicity via the miR-153/Nrf2/ARE
signaling pathway. The present study reported that the Nrf2 agonist
dimethyl fumarate (2.5 µM), induced the Nrf2/ARE signaling pathway
and reduced oxidative stress to induce neurocyte apoptosis in
vitro following anti-miRNA-153. Ji et al (27) indicated that the inhibition of
microRNA-153 may protect neurons against ischemia/reperfusion
injury via Nrf2/HO-1 signaling in an oxygen-glucose deprivation and
reoxygenation cellular model.
In conclusion, the results of the present study
suggested that the function of miRNA-153 against neurotoxicity and
apoptosis via the activation of Nrf2/ARE cytoprotection. This may
be associated with the upregulation of miRNA-153/Nrf2/ARE in
isoflurane-induced neurotoxicity and may provide a promising
therapeutic target for neurotoxicity following anesthesia.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZZ designed the present study; DS, ZW, SB and GF
performed the experiments; ZZ and DS analyzed the data; ZZ wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Changzhou No. 2 People's Hospital Affiliated to
Nanjing Medical University (Chanzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang B, Tian M, Zhen Y, Yue Y, Sherman J,
Zheng H, Li S, Tanzi RE, Marcantonio ER and Xie Z: The effects of
isoflurane and desflurane on cognitive function in humans. Anesth
Analg. 114:410–415. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lemkuil BP, Head BP, Pearn ML, Patel HH,
Drummond JC and Patel PM: Isoflurane neurotoxicity is mediated by
p75NTR-RhoA activation and actin depolymerization. Anesthesiology.
114:49–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen L, Zhang B, Shan S and Zhao X:
Neuroprotective effects of vitexin against isoflurane-induced
neurotoxicity by targeting the TRPV1 and NR2B signaling pathways.
Mol Med Rep. 14:5607–5613. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li C, Hou L, Chen D, Lin F, Chang T, Li M,
Zhang L, Niu X, Wang H, Fu S and Zheng J: Hydrogen-rich saline
attenuates isoflurane-induced caspase-3 activation and cognitive
impairment via inhibition of isoflurane-induced oxidative stress,
mitochondrial dysfunction, and reduction in ATP levels. Am J Transl
Res. 9:1162–1172. 2017.PubMed/NCBI
|
|
5
|
Sun J, Chen XL, Zheng JY, Zhou JW and Ma
ZL: Astragaloside IV protects new born rats from anesthesia-induced
apoptosis in the developing brain. Exp Ther Med. 12:1829–1835.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Abdul-Aziz A, MacEwan DJ, Bowles KM and
Rushworth SA: Oxidative stress responses and NRF2 in human
leukaemia. Oxid Med Cell Longev. 2015:4546592015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Song Zh, Tong G, Xiao K, Jiao le F, Ke Yl
and Hu Ch: L-cysteine protects intestinal integrity, attenuates
intestinal inflammation and oxidant stress, and modulates NF-κB and
Nrf2 pathways in weaned piglets after LPS challenge. Innate Immun.
22:152–161. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartolini D and Galli F: The functional
interactome of GSTP: A regulatory biomolecular network at the
interface with the Nrf2 adaption response to oxidative stress. J
Chromatogr B Analyt Technol Biomed Life Sci. 1019:29–44. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen C, Cheng W, Yu P, Wang L, Zhou L,
Zeng L and Yang Q: Resveratrol pretreatment attenuates injury and
promotes proliferation of neural stem cells following
oxygen-glucose deprivation/reoxygenation by upregulating the
expression of Nrf2, HO-1 and NQO1 in vitro. Mol Med Rep.
14:3646–3654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang LJ, Zhang SM, Li CW, Tang JY, Che FY
and Lu YC: Roles of the Nrf2/HO-1 pathway in the anti-oxidative
stress response to ischemia-reperfusion brain injury in rats. Eur
Rev Med Pharmacol Sci. 21:1532–1540. 2017.PubMed/NCBI
|
|
11
|
Sun J, Wei X, Lu Y, Cui M, Li F, Lu J, Liu
Y and Zhang X: Glutaredoxin 1 (GRX1) inhibits oxidative stress and
apoptosis of chondrocytes by regulating CREB/HO-1 in
osteoarthritis. Mol Immunol. 90:211–218. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang XS, Ha S, Wang XL, Shi YL, Duan SS
and Li ZA: Tanshinone IIA protects dopaminergic neurons against
6-hydroxydopamine-induced neurotoxicity through
miR-153/NF-E2-related factor 2/antioxidant response element
signaling pathway. Neuroscience. 303:489–502. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang W, Zhang J, Li Y, Yang X, He Y, Li T,
Ren F, Zhang J and Lin R: Divalproex sodium enhances the
anti-leukemic effects of imatinib in chronic myeloid leukemia cells
partly through SIRT1. Cancer Lett. 356:791–799. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qian C, Jin J, Chen J, Li J, Yu X, Mo H
and Chen G: SIRT1 activation by resveratrol reduces brain edema and
neuronal apoptosis in an experimental rat subarachnoid hemorrhage
model. Mol Med Rep. 16:9627–9635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li G, Xue Q, Luo Y, Hu X and Yu B: S6
inhibition contributes to isoflurane neurotoxicity in the
developing brain. Toxicol Lett. 233:102–113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu Z, Dong Y, Wu X, Zhang J, McAuliffe S,
Pan C, Zhang Y, Ichinose F, Yue Y and Xie Z: The potential dual
effects of anesthetic isoflurane on Aβ-induced apoptosis. Curr
Alzheimer Res. 8:741–752. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pan C, Xu Z, Dong Y, Zhang Y, Zhang J,
McAuliffe S, Yue Y, Li T and Xie Z: The potential dual effects of
anesthetic isoflurane on hypoxia-induced caspase-3 activation and
increases in β-site amyloid precursor protein-cleaving enzyme
levels. Anesth Analg. 113:145–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zou Y, Liu W, Zhang J and Xiang D: miR-153
regulates apoptosis and autophagy of cardiomyocytes by targeting
Mcl-1. Mol Med Rep. 14:1033–1039. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bi J, Zhang H, Lu J and Lei W: Nobiletin
ameliorates isoflurane-induced cognitive impairment via
antioxidant, anti-inflammatory and anti-apoptotic effects in aging
rats. Mol Med Rep. 14:5408–5414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cheng Y, Mitchell-Flack MJ, Wang A and
Levy RJ: Carbon monoxide modulates cytochrome oxidase activity and
oxidative stress in the developing murine brain during isoflurane
exposure. Free Radic Biol Med. 86:191–199. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liang C, Zhu H, Xu Y, Huang L, Ma C, Deng
W, Liu Y and Qin C: MicroRNA-153 negatively regulates the
expression of amyloid precursor protein and amyloid precursor-like
protein 2. Brain Res. 1455:103–113. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Li H, Huang H, Liu S, Mao X, Wang
S, Wong SS, Xia Z and Irwin MG: Cardioprotection from emulsified
isoflurane postconditioning is lost in rats with
streptozotocin-induced diabetes due to the impairment of
Brg1/Nrf2/STAT3 signalling. Clin Sci (Lond). 130:801–812. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Piras S, Furfaro AL, Brondolo L,
Passalacqua M, Marinari UM, Pronzato MA and Nitti M:
Differentiation impairs Bach1 dependent HO-1 activation and
increases sensitivity to oxidative stress in SH-SY5Y neuroblastoma
cells. Sci Rep. 7:75682017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang B, Teng Y and Liu Q: MicroRNA-153
regulates NRF2 expression and is associated with breast
carcinogenesis. Clin Lab. 62:39–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun X, Yang Y, Shi J, Wang C, Yu Z and
Zhang H: NOX4- and Nrf2-mediated oxidative stress induced by silver
nanoparticles in vascular endothelial cells. J Appl Toxicol.
37:1428–1437. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang C, Wang HJ, Bao QC, Wang L, Guo TK,
Chen WL, Xu LL, Zhou HS, Bian JL, Yang YR, et al: NRF2 promotes
breast cancer cell proliferation and metastasis by increasing
RhoA/ROCK pathway signal transduction. Oncotarget. 7:73593–73606.
2016.PubMed/NCBI
|
|
27
|
Ji Q, Gao J, Zheng Y, Liu X, Zhou Q, Shi
C, Yao M and Chen X: Inhibition of microRNA-153 protects neurons
against ischemia/reperfusion injury in an oxygen-glucose
deprivation and reoxygenation cellular model by regulating
Nrf2/HO-1 signaling. J Biochem Mol Toxicol. 31:2017.doi:
10.1002/jbt.21905. View Article : Google Scholar : PubMed/NCBI
|