Introduction
Cancer is the second most common disease that
threatens human health and mortality globally (1). Among women, breast cancer (BC) has
the highest incidence of all malignant tumors (2). Triple-negative BC (TNBC) is a subtype
of BC, which accounts for 10–20% of cases of BC globally. TNBC is
characterized by negative expression of estrogen receptor,
progesterone receptor and human epidermal growth factor receptor 2,
and is considered an independent clinicopathological type of cancer
that is characterized by strong invasiveness (3–5).
Metastasis is a critical biological hallmark of malignant tumors
and is the leading cause of mortality among patients with TNBC
(6–8); however, the specific process and
underlying mechanism remains unclear. Therefore, further study of
the mechanism underlying TNBC metastasis and exploration of its
effective prevention are of great significance in terms of
improving the survival rate and quality of life for patients.
MicroRNAs (miRNAs/miRs) are non-coding
single-stranded RNA molecules, which consist of ~22 nucleotides
(9). The role of miRNAs is
fulfilled by regulating the expression of target genes, which is
regulated by complementary binding of the miRNA to the
3′-untranslated region (3′UTR) of the target gene mRNA (10,11).
miRNAs may act as oncogenes or tumor suppressor genes in the
development of cancer (12–15).
Numerous studies have reported that the abnormal expression of
miRNAs is closely associated with the development of various tumor
types (13–18), and miRNAs are expected to be a
novel molecular target for the diagnosis and treatment of cancer.
The abnormal expression of miR-214 contributes to the formation of
various human tumors, including ovarian cancer, colorectal cancer,
gastric cancer and BC (19–22).
Furthermore, miR-214 is closely associated with regulation of the
development of tumor cells, including cell growth, apoptosis and
metastasis (21,23–25);
however, at present, the role and target of miR-214 in TNBC is not
fully understood.
The phosphoinositide 3-kinase (PI3K)/protein kinase
B (Akt)/mammalian target of rapamycin (mTOR) signal transduction
pathway is one of the most important signaling pathways in cells.
This pathway serves a vital role in regulating cell proliferation,
apoptosis, differentiation and metabolism by affecting the
activation state of several downstream effector molecules (26–28).
In recent years, abnormal activation of the PI3K/Akt/mTOR pathway
has been detected in numerous human malignant tumors, and
activation of this pathway may be a critical factor leading to the
proliferation and metastasis of tumor cells (29–31).
In the present study, the target gene of miR-214 in
TNBC was analyzed using the microRNA.org
website. In addition, the effects of miR-214 on the growth and
metastasis of TNBC cells, and the associated pathways were
analyzed.
Materials and methods
Tissue collection
Between June 2016 and November 2017, 37 TNBC, 37
non-TNBC BC and 37 normal tissues were collected from patients with
TNBC (38–55 years), patients with n-TNBC (35–53 years) and normal
control individuals (all female, 36–52 years) who were diagnosed
and subjected to mastectomy at the Second Affiliated Hospital of
Kunming Medical University (Kunming, China). All patients provided
written informed consent and allowed their tissues to be used for
research purposes. The present study was approved by the Ethics
Committee of the Second Affiliated Hospital of Kunming Medical
University.
Cell culture
The MDA-MB-231 human TNBC cell line was purchased
from Cobioer Biosciences Co., Ltd. (Nanjing, China). The cells were
cultured in RPMI-1640 medium (Huayueyang Biotechnology Co., Ltd.,
Beijing, China) supplemented with 10% fetal bovine serum (FBS;
Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.,
Hangzhou, China), and a mixture of penicillin (100 µg/ml) and
streptomycin (100 U/ml; Beijing Solarbio Science & Technology
Co., Ltd., Beijing, China) in a 95% humidified incubator (Sanyo
Electric Co., Ltd.; Panasonic Corporation, Kadoma, Japan)
containing 5% CO2 at 37°C.
Cell transfection
miR-214 mimics, miR-214 inhibitor and miRNA negative
control (NC) were purchased from Vigene Biosciences (Rockville, MD,
USA). The miRNA sequences were as follows: miR-214 mimics,
3′-UGACGGACAGACACGGACGACA-5′; miRNA inhibitor,
3′-ACUGCCUGUCUGUGCCUGCUGU-5′; and miRNA NC,
5′-UCUACUCUUUCUAGGAGGUUGUGA-3′. Cell transfection with miR-214
mimics (100 pmol), inhibitor (100 pmol) or miRNA NC (100 pmol) was
performed when the confluence of MDA-MB-231 cells reached ~60%.
Transfection was conducted using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at
37°C for 24 h. Reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis was performed to confirm successful
transfection.
Luciferase reporter assay
Putative targets of miR-214 were predicted using
miRanda version 3.3a (microRNA.org).
Luciferase activity was determined using the
Dual-Luciferase® Reporter Assay (Promega Corporation,
Madison, WI, USA). Briefly, 293 cells (Cobioer Biosciences Co.,
Ltd.) were co-transfected with miR-214 mimics or miRNA NC and
α1-AT-3′UTR or α1-AT-3′UTR mutant (mut) plasmids (400 ng) using
Lipofectamine® 2000 at 37°C for 24 h; a QuikChange
Site-Directed Mutagenesis Kit (Agilent Technologies, Inc., Santa
Clara, CA, USA) was used to obtain α1-AT-3′UTR mutant. The cells
were lysed using 1X passive lysis buffer at room temperature for 15
min and the suspension was transferred to the black enzyme plate.
LARII (100 µl) and Stop&Glo® reagent (100 µl) were
then added to the plates at room temperature, and luciferase
activity normalized to Renilla luciferase was immediately
measured using a GloMax® Discover Multimode Microplate
Reader (GM3000; Promega Corporation).
RT-qPCR analysis
Total RNA was extracted from cells and tissues using
RNA extraction kit (Takara Biotechnology Co., Ltd., Dalian, China).
Subsequently, 1 µg RNA was used to synthesize cDNA using the
SuperScript® VILO™ cDNA synthesis kit (Thermo Fisher
Scientific, Inc.). The RT reaction conditions were as follows: 30°C
for 10 min, 42°C for 30 min and 95°C for 5 min. cDNA was then
amplified using Fast SYBR® Green Master Mix (Thermo
Fisher Scientific, Inc.). The qPCR reaction conditions were as
follows: 95°C for 10 sec, followed by 40 cycles at 95°C for 5 sec
and 60°C for 30 sec, and a final step at 75°C for 1 min. Primer
sequences are listed in Table I.
U6 and β-actin were used as internal controls. mRNA expression
levels were quantified using the 2−ΔΔCq method (32).
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Primer name | Sequence
(5′-3′) | Product size
(bp) |
|---|
|
miR-214-forward |
ATAGAATTCTTTCTCCCTTTCCCCTTACTCTCC | 235 |
|
miR-214-reverse |
CCAGGATCCTTTCATAGGCACCACTCACTTTAC |
|
| α1-AT-forward |
TCAAGGACACCGAGGAAGAG | 190 |
| α1-AT-reverse |
AGGTGCTGTAGTTTCCCCTC |
|
|
E-cadherin-forward |
TTTGAAGATTGCACCGGTCG | 180 |
|
E-cadherin-reverse |
CAGCGTGACTTTGGTGGAAA |
|
| TIMP2-forward |
AGCACCACCCAGAAGAAGAG | 175 |
| TIMP2-reverse |
TGATGCAGGCGAAGAACTTG |
|
| MTA1-forward |
CTACGACCCACAGCAGAAGA | 180 |
| MTA1-reverse |
TGGTCGATCTGCTTGTCTGT |
|
| MMP2-forward |
TGGCTACACACCTGATCTGG | 184 |
| MMP2-reverse |
GAGTCCGTCCTTACCGTCAA |
|
| U6-forward |
ACACCAAGCAGTCCGAAGAG | 220 |
| U6-reverse |
ACAAAATTTCTCACGCCGGT |
|
|
β-actin-forward |
GGGAAATCGTGCGTGACATT | 219 |
|
β-actin-reverse |
AGGTAGTTTCGTGGATGCCA |
|
Western blot analysis
Total proteins were lysed from cells and tissues
using radioimmunoprecipitation assay buffer (Beijing Solarbio
Science & Technology Co., Ltd.), and protein concentrations
were determined using bicinchoninic acid protein assay (Thermo
Fisher Scientific, Inc.). Proteins (1 µg/µl; 10 µg/lane) were
separated by 12% SDS-PAGE and were transferred onto polyvinylidene
fluoride membranes (Hangzhou RENO Membrane Technology, Co., Ltd.,
Hangzhou, China, www.renomem.com). Subsequently, 5% non-fat milk was
used to block the membranes at room temperature for 1.5 h. The
membranes were then hybridized to anti-α1-AT (cat. no. ab179443,
1:1,000; Abcam, Cambridge, MA, USA), anti-E-cadherin (cat. no.
ab15148, 1:800; Abcam), anti-tissue inhibitor of
metalloproteinases-2 (TIMP2; cat. no. ab180630, 1:1,000; Abcam),
anti-metastatic tumour antigen 1 (MTA1; cat. no. ab751, 1:1,000;
Abcam), anti-matrix metalloproteinase-2 (MMP2; cat. no. ab37150,
1:1,200; Abcam), anti-phosphorylated (p)-PI3K (cat. no. ab182651,
1:1,000; Abcam), anti-PI3K (cat. no. MAB2686, 1:600; R&D
Systems, Inc.), anti-p-Akt (cat. no. MAB887, 1:800; R&D
Systems, Inc.), anti-Akt (cat. no. MAB2055, 1:800; R&D Systems,
Inc.), anti-p-mTOR (cat. no. ab109268, 1:1,000; Abcam), anti-mTOR
(cat. no. ab32028, 1:1,000; Abcam) and anti-β-actin (cat. no.
ab8227, 1:600; Abcam) at 4°C for 24 h. Subsequently, the membranes
were hybridized to the corresponding secondary antibodies (Abcam):
Horseradish peroxidase (HRP)-conjugated donkey anti-mouse
immunoglobulin (Ig)G H&L (cat. no. ab6820, 1:7,000),
HRP-conjugated rabbit anti-mouse IgG H&L (cat. no. ab6728,
1:7,000) and HRP-conjugated goat anti-rabbit IgG H&L (cat. no.
ab6721, 1:7,000) at 37°C for 60 min. The blots were detected by
enhanced chemiluminescence detection reagent (Shanghai Yeasen
Biotechnology Co., Ltd., Shanghai, China). VisionWorks®
LS Image Acquisition and Analysis software version 7.0 (UVP, LLC,
Phoenix, AZ, USA) was used to quantify band intensities.
Cell Counting kit-8 (CCK-8) assay
Cell viability was assessed by CCK-8 (Dalian Meilun
Biotechnology Co., Ltd., Dalian, China). Briefly, cells were seeded
in a 96-well plate (3×103 cells/well) at 37°C for 24 h.
Subsequently, cells were exposed to PBS (control), miRNA NC or
miR-214 inhibitor at 37°C for 6, 12 and 24 h. CCK-8 reagent (10 µl)
was then added to the cells, which were incubated for a further 4 h
at 37°C. The optical density value was then assessed at 450 nm
using a microplate reader (SMR16.1; Uscn Life Sciences, Inc.,
Wuhan, China).
Wound-healing assay
Cells were seeded in a 6-well plate
(6×104 cells/well) and were cultured for 24 h. The cells
were then exposed to PBS (control), or were transfected with miRNA
NC or miR-214 inhibitor. Subsequently, the cells were scratched
using a 200-µl pipette tip (Thermo Fisher Scientific, Inc.).
Following culturing at 37°C for 24 h, the cells were observed under
a DSX100 optical microscope (Olympus Corporation, Tokyo,
Japan).
Transwell assay
Matrigel (200 µg/ml, 100 µl; BD Biosciences,
Franklin Lakes, NJ, USA) was added to the upper Transwell chamber
at room temperature until it solidified, after which, cells were
added to the upper chamber. The medium supplemented with 15% FBS
was added to the lower chamber. The transfected cells were
centrifuged at 5,000 × g for 20 min at 4°C, and the cell suspension
(2×105 cells/ml) was cultured in the upper chamber at
37°C for 24 h. Crystal violet (0.1%; Shanghai Gefan Biotechnology
Co., Ltd., Shanghai, China) was used to stain the cells for 20 min.
Finally, cells were observed under a DSX100 optical microscope
(Olympus Corporation).
Statistical analysis
All data are presented as the mean ± standard error
of mean, as determined by Excel 2010 (Microsoft Corporation,
Redmond, WA, USA). Data were analyzed using SPSS software (version
13.0; SPSS, Inc., Chicago, IL, USA). The differences among groups
were determined by one-way analysis of variance, followed by
Dunnett's test. Each experiment was performed in triplicate and
repeated three times. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression levels of miR-214 and α1-AT
among patients with BC
RT-qPCR was used to analyze the expression levels of
miR-214 and α1-AT in normal, n-TNBC and TNBC tissues. In n-TNBC and
TNBC tissues, the expression levels of miR-214 were higher and the
mRNA expression levels of α1-AT were lower than in normal tissue
(P<0.01; Fig. 1A and 1B). In addition, according to miRanda,
α1-AT was predicted to possess an miR-214-binding site (Fig. 1C). Subsequently, it was revealed
that the luciferase activity of α1-AT-3′UTR was reduced in cells
transfected with miR-214 mimics; however, miR-214 mimics did not
affect the luciferase activity of α1-AT-3′UTR mut (Fig. 1D). Furthermore, the expression
levels of α1-AT were decreased following upregulation of miR-214
via transfection with miR-214 mimics (Fig. 1E-G). These results suggested that
α1-AT may be a target of miR-214.
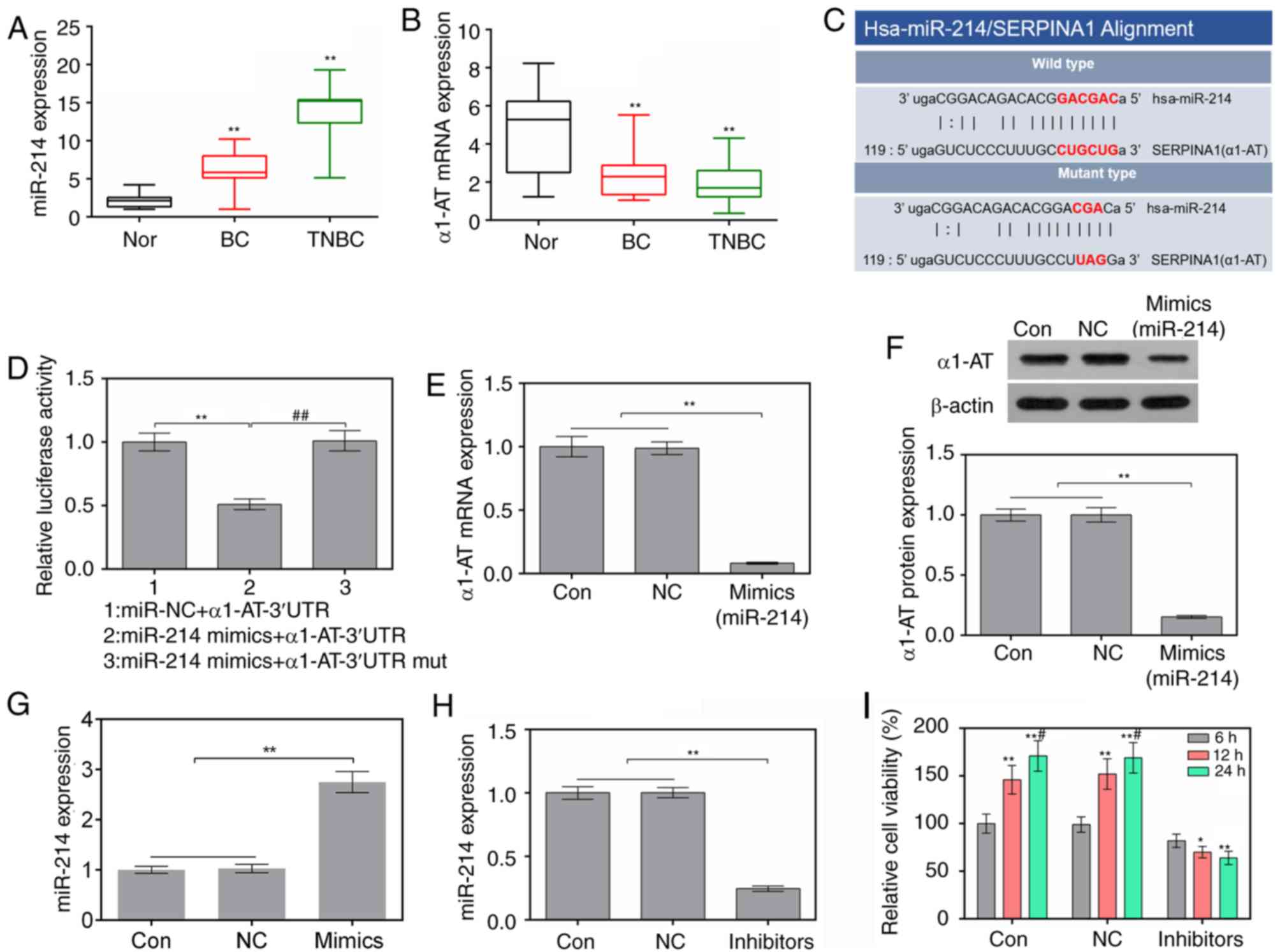 | Figure 1.Suppression of miR-214 attenuates the
viability of MDA-MB-231 cells. (A and B) Expression levels of
Hsa-miR-214 and α1-AT in normal tissues, TBNC tissues and BC
tissues, as determined by RT-qPCR. **P<0.01 vs. Normal tissues
(C) Binding sites of Hsa-miR-214 and α1-AT, as predicted by
microRNA.org. (D) miR-214 mimics and α1-AT-3′UTR
or α1-AT-3′UTR mut plasmids were co-transfected into 293 cells
using Lipofectamine® 2000. Luciferase activity was
measured by luciferase reporter assay. **P<0.01 vs. miR-NC +
α1-AT-3′UTR group; ##P<0.01 vs. miR-214 mimics +
α1-AT-3′UTR group. (E) mRNA expression levels of α1-AT in cells
were assessed using RT-qPCR. **P<0.01 vs. control and NC cells.
(F) Protein expression levels of α1-AT in cells were evaluated by
western blotting. **P<0.01 vs. control and NC cells. (G) mRNA
expression levels of miR-214 in MDA-MB-231 cells following
transfection with miR-214 mimics were assessed using RT-qPCR.
**P<0.01 vs. control and NC cells. (H) Expression levels of
miR-214 in MDA-MB-231 cells following transfection with miR-214
inhibitors were assessed by RT-qPCR. **P<0.01 vs. control and NC
cells. (I) Cell viability was determined using Cell Counting Kit-8.
*P<0.05, **P<0.01 vs. 6 h; #P<0.05 vs. 12 h.
3′UTR, 3′-untranslated region; α1-AT, α1-AT, α1-antitrypsin; BC,
breast cancer; miR-214, microRNA-214; mut, mutant; NC, negative
control; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction; TNBC, triple-negative BC. |
Suppression of miR-214 attenuates the
viability of MDA-MB-231 cells
In order to examine the effects of a miR-214
inhibitor on MDA-MB-231 cells, the expression levels of miR-214 and
cell viability were detected by RT-qPCR and CCK-8 assay,
respectively. The results of RT-qPCR revealed that the expression
levels of miR-214 were significantly decreased in cells transfected
with miR-214 inhibitor (Fig. 1H).
In addition, the miR-214 inhibitor markedly suppressed cell
viability at 12 and 24 h (Fig.
1I).
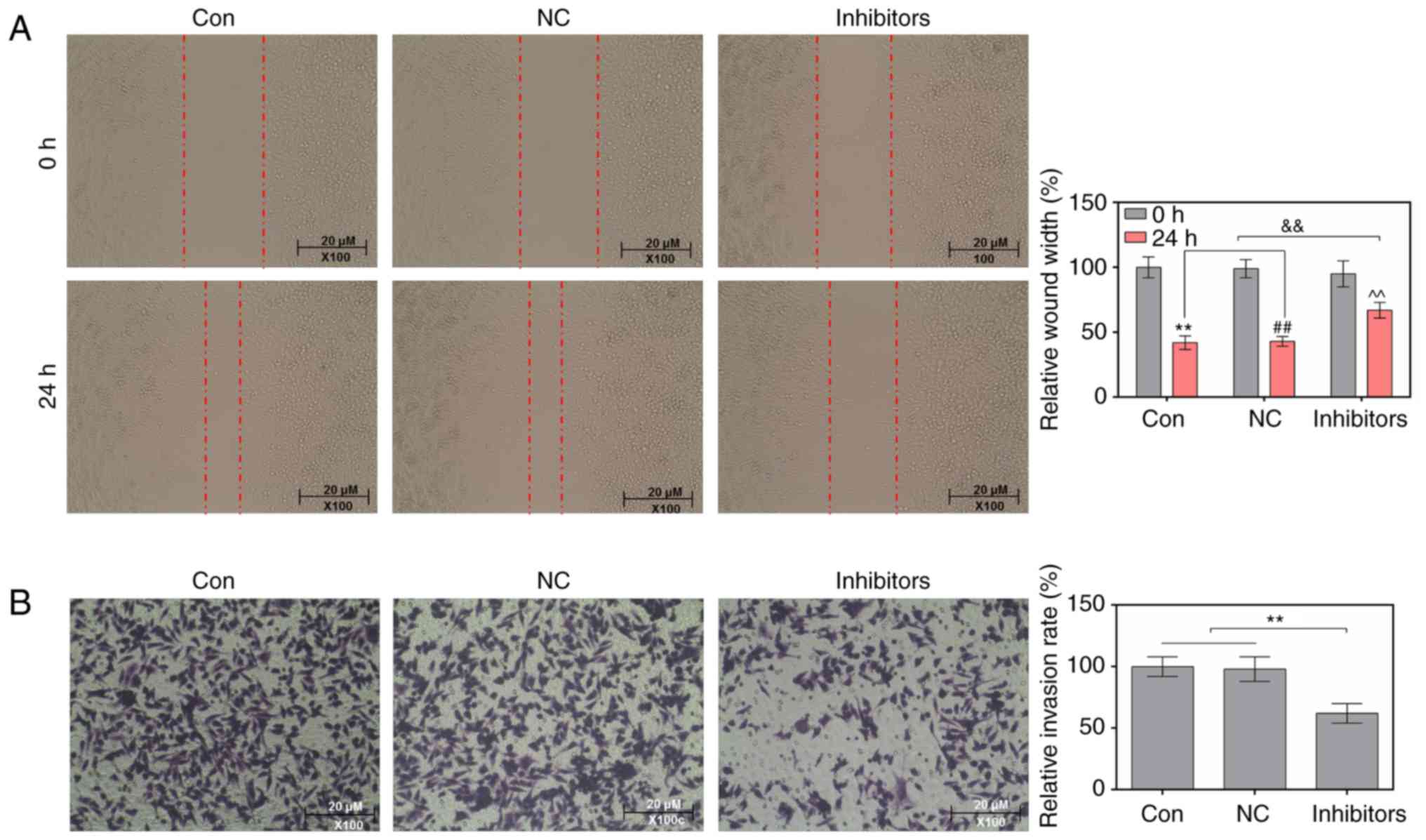
Suppression of miR-214 reduces the
migratory and invasive abilities of MDA-MB-231 cells
Wound-healing and Transwell assays were conducted to
study the effects of a miR-214 inhibitor on the migratory and
invasive abilities of MDA-MB-231 cells. As the wound-healing assay
results indicated, in cells transfected with a miR-214 inhibitor,
the relative wound width was markedly increased (Fig. 2A). The results of the Transwell
assay indicated that the miR-214 inhibitor significantly decreased
the relative invasion rate of cells (Fig. 2B).
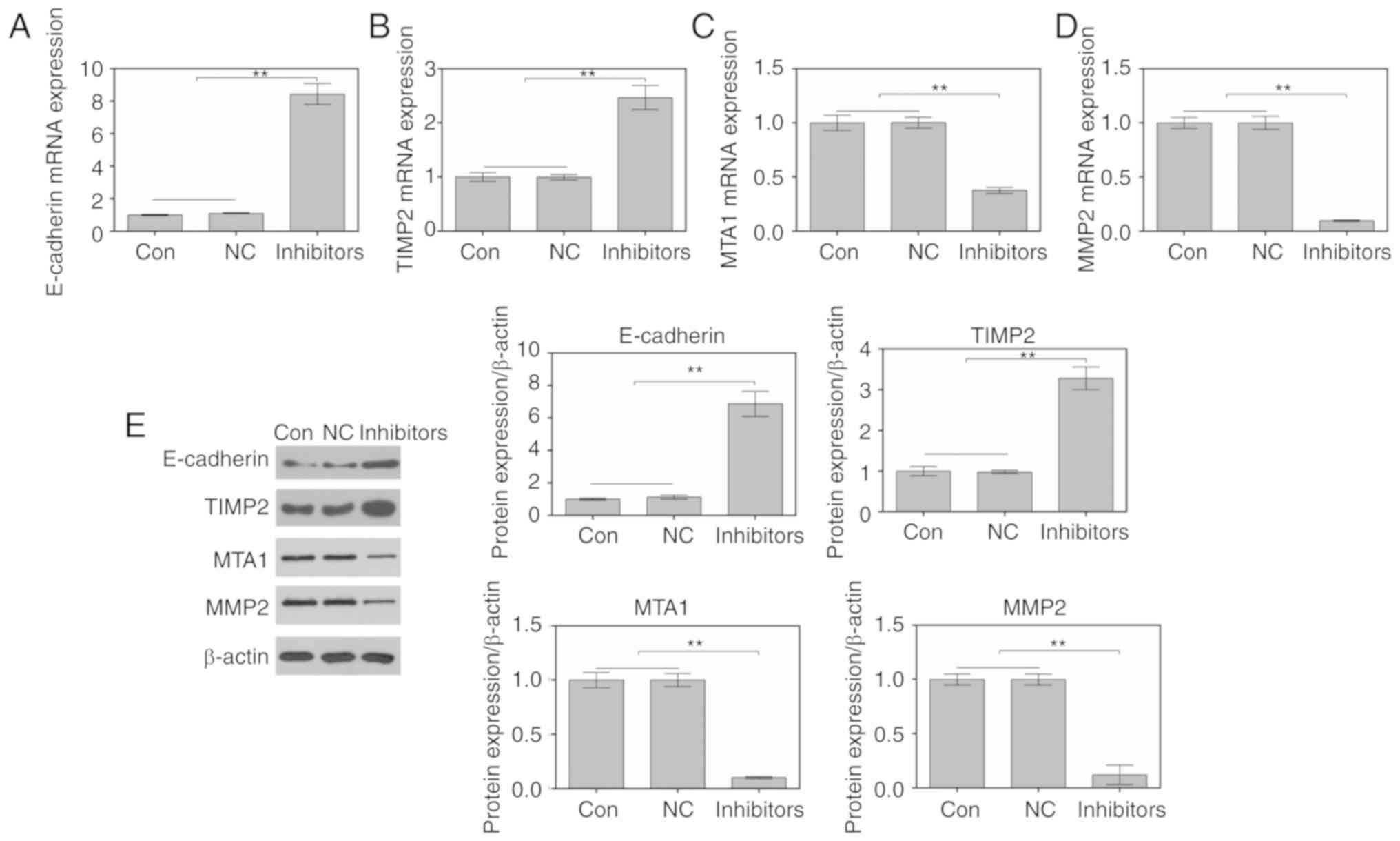
Suppression of miR-214 regulates
migration-associated factors in MDA-MB-231 cells
RT-qPCR and western blotting were performed to study
the effects of miR-214 inhibition on the expression of
migration-associated factors. As shown in Fig. 3A-D, the mRNA expression levels of
E-cadherin and TIMP2 were increased, whereas the mRNA expression
levels of MTA1 and MMP2 were decreased in cells transfected with a
miR-214 inhibitor. In addition, alterations in the protein
expression levels of E-cadherin, TIMP2, MTA1 and MMP2 were
consistent with alterations in the mRNA expression levels (Fig. 3E).
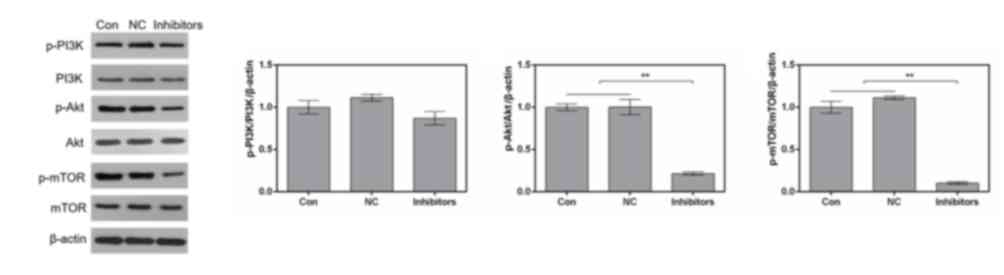
Suppression of miR-214 inhibits the
PI3K/Akt/mTOR signaling pathway in MDA-MB-231 cells
In order to investigate the effects of the miR-214
inhibitor on signaling in MDA-MB-231 cells, the PI3K/Akt/mTOR
signaling pathway was examined by western blotting. The results
revealed that transfection with the miR-214 inhibitor markedly
downregulated phosphorylation of PI3K, Akt and mTOR. However, the
expression levels of total PI3K, Akt and mTOR remained stable in
the various groups (Fig. 4). The
proportions of p-Akt/Akt and p-mTOR/mTOR were significantly reduced
by the miR-214 inhibitor (Fig.
4).
Discussion
Increasing evidence has demonstrated that miR-214 is
aberrantly expressed and its function is altered in various types
of tumor; in particular, low miR-214 expression has been detected
in liver cancer and ovarian carcinoma (21,33),
whereas high miR-214 expression has been observed in BC, gastric
cancer and pancreatic cancer (20,24,34).
Notably, Kalniete et al (20) detected high expression levels of
miR-214 in patients with TNBC. Similar to this previous study, the
present data demonstrated that miR-214 was highly expressed in TNBC
tissues. α1-AT is a member of the serine protease inhibitor
superfamily, which is dysregulated in lung cancer, prostate cancer
and BC (35,36). Previous studies have reported that
α1-AT may act as a tumor suppressor in BC cells (37,38).
The present data revealed that the expression levels of α1-AT were
decreased in TNBC tissues. According to microRNA.org, α1-AT was identified as a target gene of
miR-214. In addition, when MDA-MB-231 cells were transfected with
miR-214 mimics, the expression levels of α1-AT were downregulated.
Therefore, these findings suggested that α1-AT may be a target gene
of miR-214, and that miR-214 may exert its effect on TNBC through
targeting α1-AT.
Cell proliferation, invasion and migration are known
to be essential processes associated with tumor development. Wang
et al (39) revealed that
miR-214 increases the invasive capacity of BC cells. Zhang et
al (40) also reported that
miR-214 silencing decreases the proliferation of nasopharyngeal
carcinoma cells. Furthermore, Xin et al (41) demonstrated that miR-214 facilitates
the metastasis of gastric cancer cells. The present study explored
the effects of miR-214 on the viability, invasion and migration of
TNBC cells. The results demonstrated that inhibition of miR-214
significantly decreased the viability, migration and invasion of
MDA-MB-231 cells.
E-cadherin is an intercellular adhesion molecule,
and downregulation of E-cadherin decreases intercellular adhesion
(42). The main function of MMPs
is to degrade and reshape the dynamic balance of the extracellular
matrix. A previous study reported that when tumor cell metastasis
is inhibited, the levels of E-cadherin are upregulated and the
levels of MMPs are downregulated (43). TIMPs are tissue inhibitors of the
MMP family, and MTAs are also a class of critical tumor
metastasis-associated genes. It has been reported that tumor
necrosis factor α suppresses radiation-induced cell metastasis in
neuroblastoma via increasing TIMP2 and E-cadherin levels, and
decreasing MTA-2 and MMP levels (44). Therefore, this study examined the
effects of miR-214 on the migration/invasion-associated factors
using RT-qPCR and western blotting. It was revealed that miR-214
silencing decreased cell migration and invasion via upregulating
E-cadherin and TIMP2, and downregulating MTA1 and MMP2.
Previous studies have demonstrated that miR-214 has
a role in regulating the PI3K/Akt/mTOR pathway under various
pathological conditions (25,45,46).
Liu et al (25) reported
that miR-214 controls the PI3K/Akt pathway via targeting
phosphatase and tensin homolog in oophoroma. Zhao et al
(46) also indicated that miR-214
expedited osteoclastogenesis via regulating PI3K/Akt signaling. Li
et al (45) demonstrated
that miR-214 mediates the PI3K/Akt/mTOR signaling pathway in rat
muscle atrophy. Therefore, it was hypothesized that miR-214 may
mediate the PI3K/Akt/mTOR pathway in TNBC. As expected, knockdown
of miR-214 suppressed the PI3K/Akt/mTOR pathway in MDA-MB-231
cells.
In conclusion, this study demonstrated that miR-214
was highly expressed in TNBC tissues, and that α1-AT was a target
gene for miR-214. Silencing of miR-214 markedly suppressed the
proliferation, migration/invasion of MDA-MB-231 cells by
upregulating E-cadherin and TIMP2, and downregulating MTA1 and
MMP2. Furthermore, miR-214 silencing inhibited the PI3K/Akt/mTOR
pathway. These findings indicated that miR-214 targeting α1-AT may
be a potential mechanism underlying TNBC development.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JM made substantial contributions towards the design
of the study. YT collected breast cancer and normal tissues. YiL
performed gene expression experiments in tissues. YM performed
target prediction for miR-214. YaL performed cell transfections. LD
performed viability, migration and invasion assays. SL performed
RT-qPCR assays. ZZ performed western blotting. YZ analyzed the data
and drafted the manuscript. All authors read and approved of the
final version of the manuscript.
Ethics approval and consent to
participate
All patients provided written informed consent and
allowed their tissues to be used for research purposes. The present
study was approved by the Ethics Committee of the Second Affiliated
Hospital of Kunming Medical University.
Patient consent for publication
All participants provided written informed consent
prior to the present study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wu J, Goyal L, Nipp R, Wo J, Qadan M and
Uppot RN: The Tipping Point: Key oncologic imaging findings
resulting in critical changes in the management of malignant tumors
of the gastrointestinal tract. Curr Probl Diagn Radiol. 48:61–74.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dunne M, Dou YN, Drake DM, Spence T,
Gontijo SML, Wells PG and Allen C: Hyperthermia-mediated drug
delivery induces biological effects at the tumor and molecular
levels that improve cisplatin efficacy in triple negative breast
cancer. J Control Release. 282:35–45. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park IH, Kong SY, Kwon Y, Kim MK, Sim SH,
Joo J and Lee KS: Phase I/II clinical trial of everolimus combined
with gemcitabine/cisplatin for metastatic triple-negative breast
cancer. J Cancer. 9:1145–1151. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zenzola V, Cabezas-Quintario MA, Arguelles
M, Perez-Fernandez E, Izarzugaza Y, Correa A and Garcia-Foncillas
J: Prognostic value of Ki-67 according to age in patients with
triple-negative breast cancer. Clin Transl Oncol. 20:1448–1454.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Farah M, Nagarajan P, Torres-Cabala CA,
Curry JL, Amaria RN, Wargo J, Tawbi H, Ivan D, Prieto VG, Tetzlaff
MT and Aung PP: Metastatic melanoma with balloon/histiocytoid
cytomorphology after treatment with immunotherapy: A histologic
mimic and diagnostic pitfall. J Cutan Pathol. 45:545–549. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hejduk B, Bobek-Billewicz B, Rutkowski T,
Hebda A, Zawadzka A and Jurkowski MK: Application of Intravoxel
Incoherent Motion (IVIM) model for differentiation between
metastatic and non-metastatic head and neck lymph nodes. Pol J
Radiol. 82:506–510. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma J, Yang Y, Huo D, Wang Z, Zhai X, Chen
J, Sun H, An W, Jie J and Yang P: LincRNA-RoR/miR-145 promote
invasion and metastasis in Triple-negative breast cancer via
targeting MUC1. Biochem Biophys Res Commun. 500:614–620. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kumar MS, Lu J, Mercer KL, Golub TR and
Jacks T: Impaired microRNA processing enhances cellular
transformation and tumorigenesis. Nat Genet. 39:673–677. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abdellatif M: Differential expression of
microRNAs in different disease states. Circ Res. 110:638–650. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pillai RS: MicroRNA function: Multiple
mechanisms for a tiny RNA? RNA. 11:1753–1761. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo Y, Jiang Y, Sang M and Xu C:
Down-regulation of miR-373 increases the radiosensitivity of lung
cancer cells by targeting TIMP2. Int J Biochem Cell Biol.
99:203–210. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li X, Zhao Z, Li M, Liu M, Bahena A, Zhang
Y, Zhang Y, Nambiar C and Liu G: Sulforaphane promotes apoptosis,
and inhibits proliferation and self-renewal of nasopharyngeal
cancer cells by targeting STAT signal through miRNA-124-3p. Biomed
Pharmacother. 103:473–481. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu X, He B, Xu T, Pan Y, Hu X, Chen X and
Wang S: MiR-490-3p functions as a tumor suppressor by inhibiting
oncogene VDAC1 expression in colorectal cancer. J Cancer.
9:1218–1230. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Polakovicova I, Jerez S, Wichmann IA,
Sandoval-Borquez A, Carrasco-Veliz N and Corvalan AH: Role of
microRNAs and exosomes in helicobacter pylori and epstein-barr
virus associated gastric cancers. Front Microbiol. 9:6362018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shan X, Zhang H, Zhang L, Zhou X, Wang T,
Zhang J, Shu Y, Zhu W, Wen W and Liu P: Identification of four
plasma microRNAs as potential biomarkers in the diagnosis of male
lung squamous cell carcinoma patients in China Med 7. 2370–2381.
2018.PubMed/NCBI
|
|
17
|
Wang Y, Bao W, Liu Y, Wang S, Xu S, Li X,
Li Y and Wu S: miR-98-5p contributes to cisplatin resistance in
epithelial ovarian cancer by suppressing miR-152 biogenesis via
targeting Dicer1. Cell Death Dis. 9:4472018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang T, Liu W, Meng W, Zhao H, Yang Q, Gu
SJ, Xiao CC, Jia CC and Fu BS: Downregulation of miR-542-3p
promotes cancer metastasis through activating TGF-β/Smad signaling
in hepatocellular carcinoma. Onco Targets Ther. 11:1929–1939. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu JL, He GY, Lan XL, Zeng ZC, Guan J,
Ding Y, Qian XL, Liao WT, Ding YQ and Liang L: Inhibition of
ATG12-mediated autophagy by miR-214 enhances radiosensitivity in
colorectal cancer. Oncogenesis. 7:162018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kalniete D, Nakazawa-Miklasevica M,
Strumfa I, Abolins A, Irmejs A, Gardovskis J and Miklasevics E:
High expression of miR-214 is associated with a worse
disease-specific survival of the triple-negative breast cancer
patients. Hered Cancer Clin Pract. 13:72015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Lin J, Zhai S, Sun C, Xu C, Zhou H
and Liu H: MicroRNA-214 suppresses ovarian cancer by targeting
β-Catenin. Cell Physiol Biochem. 45:1654–1662. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X, Zhang H, Bai M, Ning T, Ge S, Deng
T, Liu R, Zhang L, Ying G and Ba Y: Exosomes serve as nanoparticles
to deliver anti-miR-214 to reverse chemoresistance to cisplatin in
gastric cancer. Mol Ther. 26:774–783. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen X, Wang YW, Zhu WJ, Li Y, Liu L, Yin
G and Gao P: A four-microRNA signature predicts lymph node
metastasis and prognosis in breast cancer. Hum Pathol. 76:122–132.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li HL, Liang S, Cui JH and Han GY:
Targeting of GSK-3β by miR-214 to facilitate gastric cancer cell
proliferation and decrease of cell apoptosis. Eur Rev Med Pharmacol
Sci. 22:127–134. 2018.PubMed/NCBI
|
|
25
|
Liu J, Chen W, Zhang H, Liu T and Zhao L:
miR-214 targets the PTEN-mediated PI3K/Akt signaling pathway and
regulates cell proliferation and apoptosis in ovarian cancer. Oncol
Lett. 14:5711–5718. 2017.PubMed/NCBI
|
|
26
|
Costa RLB, Han HS and Gradishar WJ:
Targeting the PI3K/AKT/mTOR pathway in triple-negative breast
cancer: A review. Breast Cancer Res Treat. 169:397–406. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ramakrishnan V and Kumar S: PI3K/AKT/mTOR
pathway in multiple myeloma: From basic biology to clinical
promise. Leuk Lymphoma. 1-11:2018.
|
|
28
|
Simioni C, Martelli AM, Zauli G, Vitale M,
McCubrey JA, Capitani S and Neri LM: Targeting the
phosphatidylinositol 3-kinase/Akt/mechanistic target of rapamycin
signaling pathway in B-lineage acute lymphoblastic leukemia: An
update. J Cell Physiol. 233:6440–6454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Du L, Li X, Zhen L, Chen W, Mu L, Zhang Y
and Song A: Everolimus inhibits breast cancer cell growth through
PI3K/AKT/mTOR signaling pathway. Mol Med Rep. 17:7163–7169.
2018.PubMed/NCBI
|
|
30
|
Yeh YH, Hsiao HF, Yeh YC, Chen TW and Li
TK: Inflammatory interferon activates HIF-1α-mediated
epithelial-to-mesenchymal transition via PI3K/AKT/mTOR pathway. J
Exp Clin Cancer Res. 37:702018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang H, Xu HL, Wang YC, Lu ZY, Yu XF and
Sui DY: 20(S)-Protopanaxadiol-induced apoptosis in MCF-7 breast
cancer cell line through the inhibition of PI3K/AKT/mTOR signaling
pathway. Int J Mol Sci. 19(pii): E10532018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Method. 25:402–408. 2001.
View Article : Google Scholar
|
|
33
|
Fan Y, Qian X and Zhang C: U/G SNP
rs111904020 in 3′UTR of STAT3 regulated by miR-214 promotes
hepatocellular carcinoma development in Chinese population. Tumour
Biol. 37:14629–14635. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kuninty PR, Bojmar L, Tjomsland V, Larsson
M, Storm G, Ostman A, Sandstrom P and Prakash J: MicroRNA-199a and
−214 as potential therapeutic targets in pancreatic stellate cells
in pancreatic tumor. Oncotarget. 7:16396–16408. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
El-Akawi ZJ, Al-Hindawi FK and Bashir NA:
Alpha-1 antitrypsin (alpha1-AT) plasma levels in lung, prostate and
breast cancer patients. Neuro Endocrinol Lett. 29:482–484.
2008.PubMed/NCBI
|
|
36
|
Hamrita B, Chahed K, Trimeche M, Guillier
CL, Hammann P, Chaieb A, Korbi S and Chouchane L: Proteomics-based
identification of alpha1-antitrypsin and haptoglobin precursors as
novel serum markers in infiltrating ductal breast carcinomas. Clin
Chim Acta. 404:111–118. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
García-Orad A, Arizti P, Durán L, Urcelay
B and De Pancorbo MM: Alpha-1-antitrypsin phenotypes among breast
cancer patients in the basque population. Hum Hered. 44:203–208.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lópezárias E, Aguilarlemarroy A, Felipe
JL, Morganvillela G, Mariscalramírez I, Martínezvelázquez M,
Alvarez AH, Gutiérrezortega A and Hernándezgutiérrez R: Alpha
1-antitrypsin: A novel tumor-associated antigen identified in
patients with early-stage breast cancer. Electrophoresis.
33:2130–2137. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang F, Lv P, Liu X, Zhu M and Qiu X:
microRNA-214 enhances the invasion ability of breast cancer cells
by targeting p53. Int J Mol Med. 35:1395–1402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang ZC, Li YY, Wang HY, Fu S, Wang XP,
Zeng MS, Zeng YX and Shao JY: Knockdown of miR-214 promotes
apoptosis and inhibits cell proliferation in nasopharyngeal
carcinoma. PLoS One. 9:e861492014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xin R, Bai F, Feng Y, Jiu M, Liu X, Bai F,
Nie Y and Fan D: MicroRNA-214 promotes peritoneal metastasis
through regulating PTEN negatively in gastric cancer. Clin Res
Hepatol Gastroenterol. 40:748–754. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pieters T and van Roy F: Role of cell-cell
adhesion complexes in embryonic stem cell biology. J Cell Sci.
127:2603–2613. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Che F, Xie X, Wang L, Su Q, Jia F, Ye Y,
Zang L, Wang J, Li H, Quan Y, et al: B7-H6 expression is induced by
lipopolysaccharide and facilitates cancer invasion and metastasis
in human gliomas. Int Immunopharmacol. 59:318–327. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Aravindan S, Natarajan M, Herman TS and
Aravindan N: Radiation-induced TNFalpha cross signaling-dependent
nuclear import of NFκB favors metastasis in neuroblastoma. Clin Exp
Metastasis. 30:807–817. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li G, Li QS, Li WB, Wei J, Chang WK, Chen
Z, Qiao HY, Jia YW, Tian JH and Liang BS: miRNA targeted signaling
pathway in the early stage of denervated fast and slow muscle
atrophy. Neural Regen Res. 11:1293–1303. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao C, Sun W, Zhang P, Ling S, Li Y, Zhao
D, Peng J, Wang A, Li Q, Song J, et al: miR-214 promotes
osteoclastogenesis by targeting Pten/PI3k/Akt pathway. RNA Biol.
12:343–353. 2015. View Article : Google Scholar : PubMed/NCBI
|