Introduction
Neural stem cells (NSCs) are undifferentiated
self-renewing progenitor cells that are able to differentiate into
neurons, astrocytes or oligodendrocytes (1). Previous studies have identified NSC
transplantation as a promising regenerative therapy for a number of
central nervous system disorders, including spinal cord injuries,
neurodegenerative diseases, traumatic brain injury and ischemic
brain injury (2,3). However, this potentially powerful
technology is underdeveloped for clinical application at present,
and additional studies are required to explore the mechanisms that
regulate NSC proliferation and differentiation.
MicroRNAs (miRNAs) are small noncoding RNAs,
measuring 21–22 nucleotides in length, that regulate gene
expression by binding to partially complementary sequences in the
3′untranslated region (UTR) of one or more target mRNAs (4). miRNAs have been demonstrated to serve
crucial roles in numerous cell processes, including proliferation,
differentiation and apoptosis (5–7).
Consistent with this, miRNAs are abundant in the central nervous
system, where they have critical roles in neurogenesis and neuronal
physiology (8,9). In addition, previous data have
indicated that miRNAs are involved in regulating stem cell
proliferation and differentiation by controlling the expression of
a number of stem cell regulators (10–12).
Based on a pilot miRNA microarray study, it was
hypothesized that miR-329-3p may be expressed at decreased levels
in NSCs compared with terminally differentiated neurons. However,
the expression levels and role of this miRNA in NSC biology remain
unclear. The present study identified that miR-329-3p expression
was upregulated during NSC differentiation, while that of
transcription factor E2F1 (E2F1), which was confirmed to be a
miR-329-3p target gene, was decreased. Downregulation of E2F1
inhibited the proliferation of NSCs. Transfection of NSCs with
miR-329-3p mimic inhibited E2F1 expression and cell proliferation,
which was reversed by the restoration of E2F1 expression. These
results indicated that miR-329-3p may serve an important role in
regulating NSC proliferation by targeting E2F1 mRNA. The results of
the present study may contribute to the development of regenerative
therapies based on NSC transplantation.
Materials and methods
Cell culture and transfection
The NSCs used in the present study were derived from
the hippocampus and the subventricular zone of the brain of
Sprague-Dawley rats at 14.5 days of gestation, and were purchased
from Cyagen Biosciences, Inc. (Guangzhou, China; RASNF-01001). The
cells were cultured in Dulbecco's modified Eagle's medium/F12
medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA) with
2% B27 supplement without vitamin A (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 20 ng/ml basic fibroblast
growth factor (PeproTech, Inc., Rocky Hill, NJ, USA), 20 ng/ml
epidermal growth factor (PeproTech, Inc.), and 2 mM L-glutamine
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Cells were
maintained at 37°C in a 5% CO2 humidified atmosphere and
the culture medium was changed every 2–3 days. The cells were
allowed to proliferate to form neurospheres.
NSCs were transfected with miR-329-3p-specific or
negative control (NC) mimics, E2F1-targeting or NC small
interfering RNAs (siRNAs) (50 nmol/l) (GenePharma Co., Ltd.,
Suzhou, China), and E2F1 overexpression or control vectors (Generay
Biotech Co., Ltd., Shanghai, China) using Lipofectamine®
3000 (Invitrogen; Thermo Fisher Scientific, Inc.), in accordance
with the manufacturer's protocol. Subsequent experiments took place
24 h after transfection. The mimic and siRNA sequences are
presented in Table I. The
transfection efficiency was evaluated by reverse transcription
quantitative polymerase chain reaction (RT-qPCR) as described
subsequently.
 | Table I.Sequences of mimics, siRNAs and
primers used in the present study. |
Table I.
Sequences of mimics, siRNAs and
primers used in the present study.
| A, Mimic and siRNA
sequences |
|---|
|
|---|
| Name | Direction | Sequences |
|---|
| miR-329-3p
mimic | Sense |
5′-AACACACCCAGCUAACCUUUUU-3′ |
|
| Antisense |
5′-AAAGGUUAGCUGGGUGUGUUUU-3′ |
| E2F1 siRNA | Sense |
5′-GCAGAAACGACGCAUCUAUTT-3′ |
|
| Antisense |
5′-AUAGAUGCGUCGUUUCUGCTT-3′ |
|
| B, Reverse
transcription-quantitative polymerase chain reaction
primers |
|
| Name |
Direction |
Sequences |
|
| miR-329-3p | Forward |
5′-GCGCGAACACACCCAGCTAACCTTTTT-3′ |
| U6 | Forward |
5′-CTCGCTTCGGCAGCACA-3′ |
|
| Reverse |
5′-AACGCTTCACGAATTTGCGT-3′ |
| E2F1 | Forward |
5′-GAAGAAGACCGGTTGTCACC-3′ |
|
| Reverse |
5′-GAAATCCAGAGGGGTCAAGTC −3′ |
| GAPDH | Forward |
5′-ATTCCATGGCACCGTCAAGGCTGA-3′ |
|
| Reverse |
5′-TTCTCCATGGTGGTGAAGACGCCA-3′ |
Cell differentiation
NSCs were cultured for 24 h in culture dishes which
had been coated with extracellular matrix proteins (Matrigel;
Corning Incorporated, Corning, NY, USA) at 37°C overnight, and
neuronal differentiation was then induced by changing the medium to
neurobasal medium supplemented with 2% B27 (both from Gibco; Thermo
Fisher Scientific, Inc.) and 2 mM l-glutamine (Sigma-Aldrich; Merck
KGaA).
GeneChip microarray assay
miRNAs were isolated from NSCs and differentiated
neurons with the miRNeasy kit (Qiagen GmbH, Hilden, Germany), as
described by Zou et al (13). GeneChip microarray assays were
performed by CapitalBio Technology (Beijing, China), also as
previously described (13). The
expression of key miRNAs was further verified by RT-qPCR.
RT-qPCR
Total RNA was isolated from NSCs using
TRIzol® reagent (Takara Biotechnology Co., Ltd., Dalian,
China), in accordance with the manufacturer's protocol. For
miR-329-3p detection, miRNAs were reverse transcribed using the
miRcute Plus miRNA First-Strand cDNA Synthesis kit (Tiangen Biotech
Co., Ltd., Beijing, China) and amplified by qPCR using a miRcute
Plus miRNA SYBR Green qPCR Detection kit (Tiangen Biotech Co.,
Ltd.). The reverse transcription of the miRNAs consisted of 60 min
at 42°C and 3 min at 95°C. The PCR cycle conditions included an
initial denaturation step at 95°C for 15 min, followed by 40 cycles
of 94°C for 20 sec and 60°C for 34 sec. For mRNAs, RNA was reverse
transcribed using a PrimeScript™ RT Reagent kit (Takara
Biotechnology Co., Ltd.) and amplified using SYBR Premix Ex Taq™ II
(Takara Biotechnology Co., Ltd.), in accordance with the
manufacturer's protocol. The reverse transcription process
consisted of 15 min at 37°C and 5 sec at 85°C, followed by storage
at 4°C. The qPCR cycle conditions included an initial denaturation
step at 95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec
and 60°C for 34 sec. RT-qPCR was performed using an ABI 7500 system
(Thermo Fisher Scientific, Inc.). GAPDH and U6 snRNA were used as
internal controls for the detection of mRNA and miRNA. The
2−ΔΔCq method was used to calculate relative gene
expression (14). Primer sequences
were synthesized by Sangon Biotech Co., Ltd., (Shanghai, China),
with the exception of a universal reverse primer for miR-329-3p
(Tiangen Biotech Co., Ltd.). The primer sequences are listed in
Table I.
Western blot analysis
Western blot analysis was performed in accordance
with standard methods. Cells were lysed on ice for 30 min in lysis
buffer containing phenylmethylsulfonyl fluoride (Beyotime Institute
of Biotechnology, Haimen, China) and centrifuged at 12,000 × g for
20 min at 4°C. The supernatants were collected and protein was
quantified using a BCA Protein Assay kit (Wanleibio, Co., Ltd.,
Shanghai, China). A total of 30 µg/lane total protein were
separated by 12% SDS-PAGE and electrotransferred to polyvinylidene
fluoride membranes. The membranes were blocked by incubation in
2.5% (wt/vol) skimmed milk powder in TBS containing 0.1% Tween-20
(TBST) for 2 h at room temperature, washed with TBST 3 times, and
then incubated at 4°C overnight with rabbit anti-rat polyclonal
E2F1 antibody (1:500 dilution; cat. no. WL02394; Wanleibio Co.,
Ltd) or rabbit anti-rat polyclonal β-actin antibody (1:1,000
dilution; cat. no. WL0002c; Wanleibio Co., Ltd) diluted in TBST.
The membranes were washed again in TBST and incubated with
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:5,000 dilution; cat. no. AS014; ABclonal Biotech Co.,
Ltd., Woburn, MA, USA) for 2 h at room temperature. Finally, the
membranes were washed 3 times for 10 min each and the protein bands
were visualized using an enhanced chemiluminescence detection
system. Densitometric analysis was performed using Image-Pro Plus
6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Cell proliferation assay
A Cell Counting Kit-8 assay (CCK-8; Beyotime
Institute of Biotechnology) and a 5-ethynyl-2′-deoxyuridine (EdU)
incorporation assay (Guangzhou RiboBio Co., Ltd., Guangzhou, China)
were used to measure NSCs proliferation, in accordance with the
manufacturers' protocols. For the CCK-8 assay, NSCs were plated at
5×103/100 µl/well (n=5/condition) in 96-well plates for
24 h and then transfected with the mimics or siRNAs using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Following incubation for 0, 24, 48 or 72 h at
37°C, 10 µl CCK-8 reagent was added to each well and the plates
were incubated at 37°C for an additional 2 h. The absorbance at 450
nm was measured using a microplate reader (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). For the EdU incorporation assay, the
cells were plated at 1×104 cells/well in 96-well plates,
transfected with the mimics or siRNAs for 48 h using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.), and incubated in culture medium containing 50 µM
EdU for 4 h. The cells were washed with PBS for 5 min, fixed in 4%
paraformaldehyde for 30 min, incubated in 50 µl 2 mg/ml glycine
solution for 5 min, washed with PBS for 5 min, permeabilized with
100 µl 0.5% Triton X-100/PBS for 10 min, washed again with PBS, and
then incubated in 100 µl of 1X Apollo® staining solution
(Guangzhou RiboBio Co., Ltd.) for 30 min. The cells were washed 3
times for 10 min each with 0.5% Triton X-100/PBS, incubated with
100 µl 1X Hoechst 33342 for 30 min, and then washed twice with PBS.
All of these procedures were performed at room temperature.
Finally, the cells were observed under a fluorescence microscope
(Olympus Corporation, Tokyo, Japan; magnification, ×400).
Prediction of miRNA target genes
Potential targets for miR-329-3p were predicted
using the bioinformatics databases TargetScan (http://www.targetscan.org/) and miRDB (http://mirdb.org/index.html) (15,16).
Dual-luciferase reporter assay
The wild-type (wt) or mutated (mt) E2F1 3′-UTR
sequence was cloned into the pmirGLO Luciferase reporter vector
(Generay Biotech Co., Ltd.) and transfected into 293 cells
(American Type Culture Collection, Manassas, VA, USA). Cells were
seeded at 1×105 cells per well in 24-well plates and
co-transfected at 37°C with 60 nM miR-329-3p or control mimic
together with 1 µg wt or mt E2F1 3′-UTR vectors using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 24 h from transfection, the cells were
given fresh medium and cultured for an extra 24 h. Subsequently,
luciferase activity was measured using the Dual-Luciferase Reporter
reagent (Promega Corporation, Madison, WI, USA), in accordance with
the manufacturer's protocol. Relative luciferase activity was
calculated as follows: (Firefly luciferase activity/Renilla
luciferase activity) ×100%.
Statistical analysis
GraphPad Prism 7.0 software (GraphPad Software,
Inc., La Jolla, CA, USA) was used for all analyses. Data are
presented as the mean ± standard deviation. Differences between
groups were analyzed using unpaired Student's t-test and one- or
two-way analysis of variance, followed by a Dunnett's or Sidak's
multiple comparisons test, as appropriate. P<0.05 was considered
to indicate a statistically significant difference.
Results
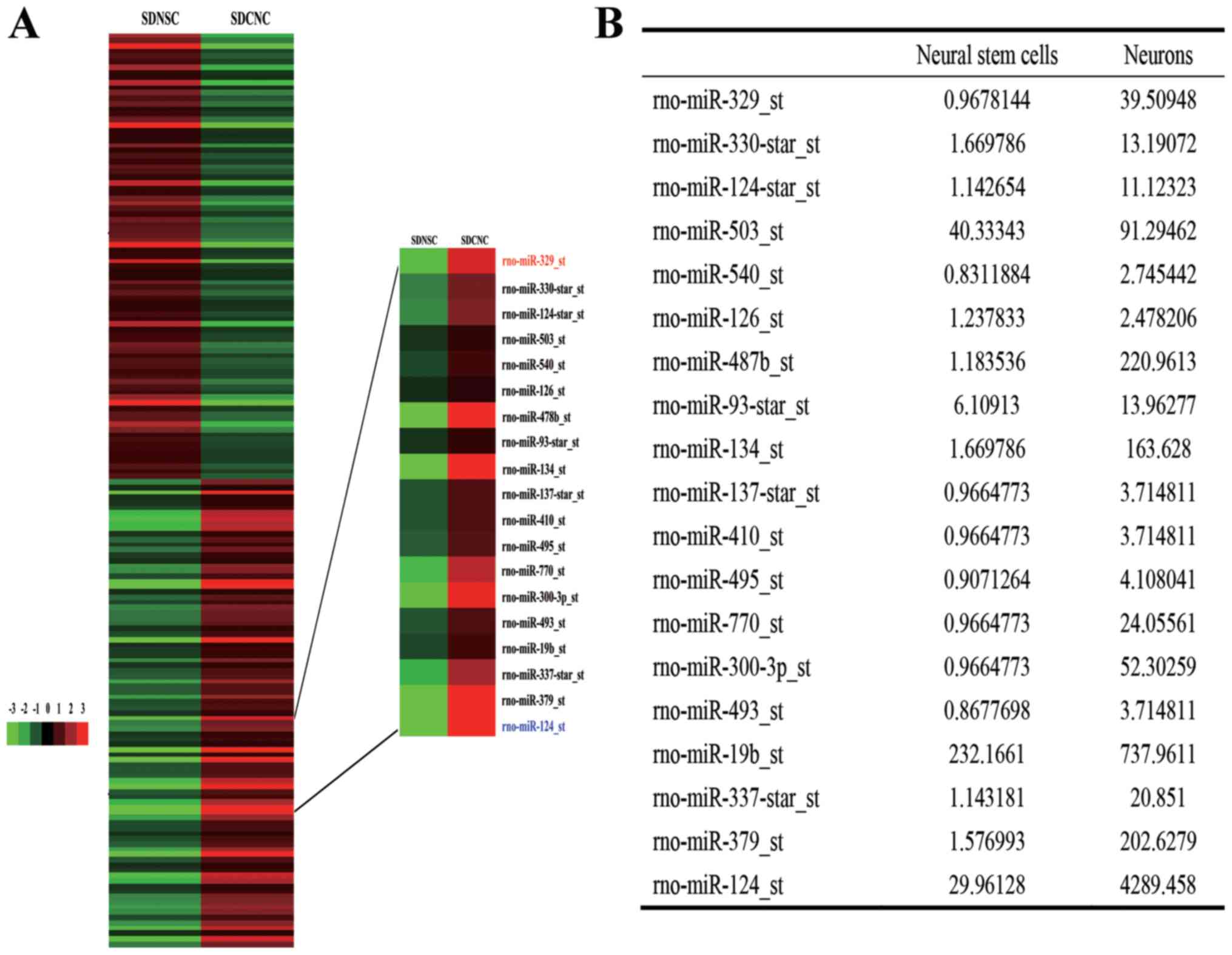
Differential expression of miRNAs in
NSCs and neurons
Analysis of the GeneChip data indicated that
miR-329-3p was expressed at increased levels in differentiated rat
cortical neurons compared with in the starting population of NSCs
(Fig. 1A and B).
miR-329-3p is upregulated and E2F1 is
downregulated during NSC differentiation
To validate the results of the GeneChip microarray
assay, RT-qPCR analysis of miR-329-3p levels in NSCs on day 0 and
after 1, 3, and 5 days of culture in medium that promotes neuronal
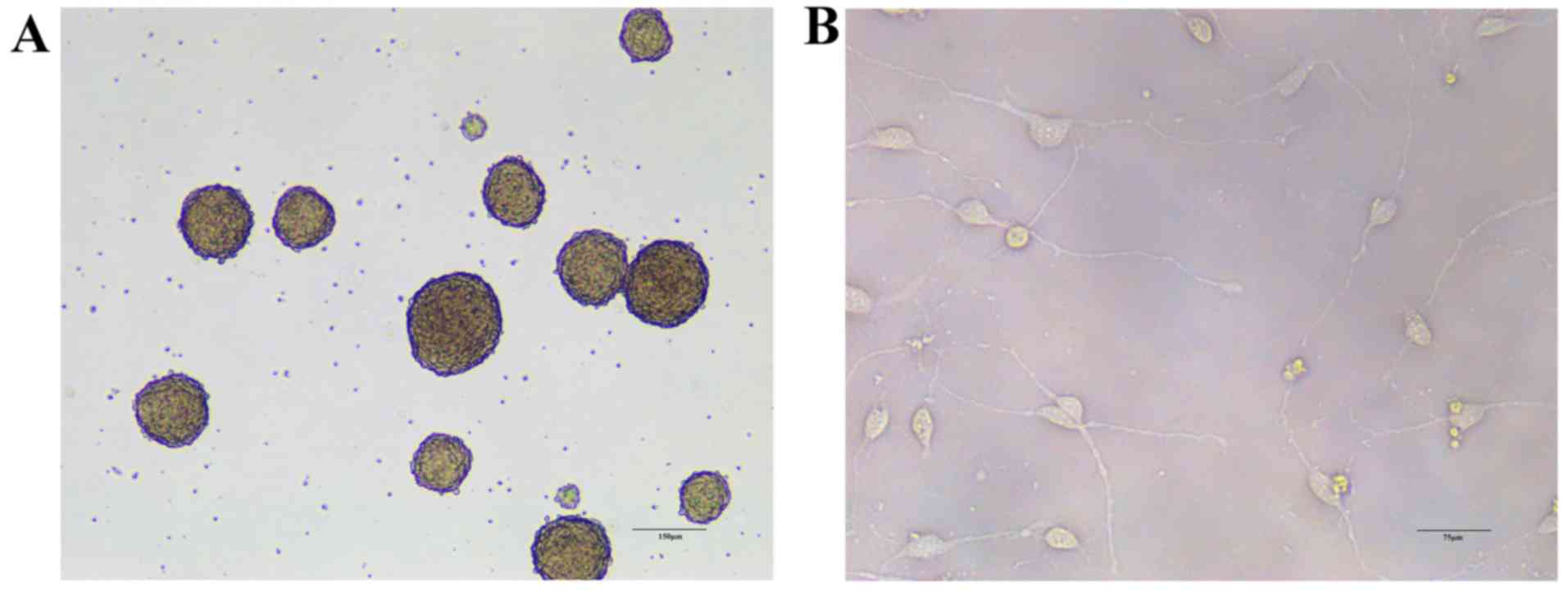
differentiation was performed. The morphology of NSCs and cells
differentiated from NSCs is presented in Fig. 2. The results demonstrated that
miR-329-3p expression was gradually upregulated during the
differentiation process (Fig. 3A).
The E2F1 expression was also measured in differentiating NSCs at
the mRNA and protein levels using RT-qPCR and western blot
analysis, respectively. Expression of E2F1 mRNA (Fig. 3B) and protein (Fig. 3C and D) was identified to decrease
over the course of neuronal differentiation, which contrasted with
the increase in miR-329-3p expression observed.
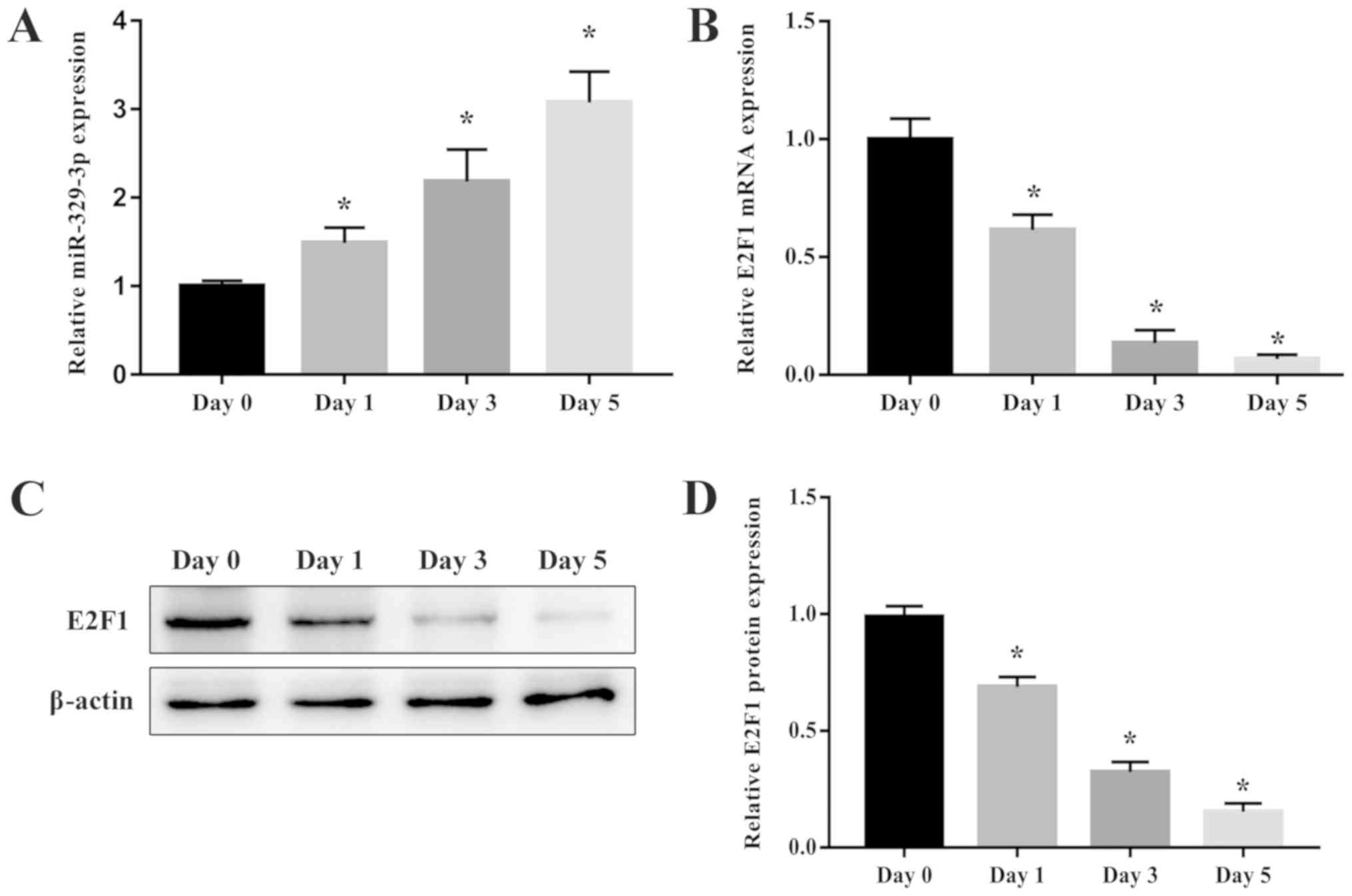 | Figure 3.Expression of miR-329-3p and E2F1
during NSC differentiation. (A) RT-qPCR analysis of miR-329-3p on
days 0, 1, 3, and 5 after induction of neuronal differentiation.
(B) RT-qPCR analysis of E2F1 mRNA expression and (C) western blot
analysis of E2F1 protein expression on days 0, 1, 3, and 5 after
induction of neuronal differentiation. GAPDH and β-actin were
probed as internal controls for (B) and (C) respectively. (D)
Quantification of protein expression. U6 was measured as an
internal control for miR-329-3p. Data are presented as the mean ±
standard deviation. *P<0.05 vs. day 0. miR, microRNA; E2F1,
transcription factor E2F1; NSC, neural stem cell; RT-qPCR, reverse
transcription quantitative polymerase chain reaction. |
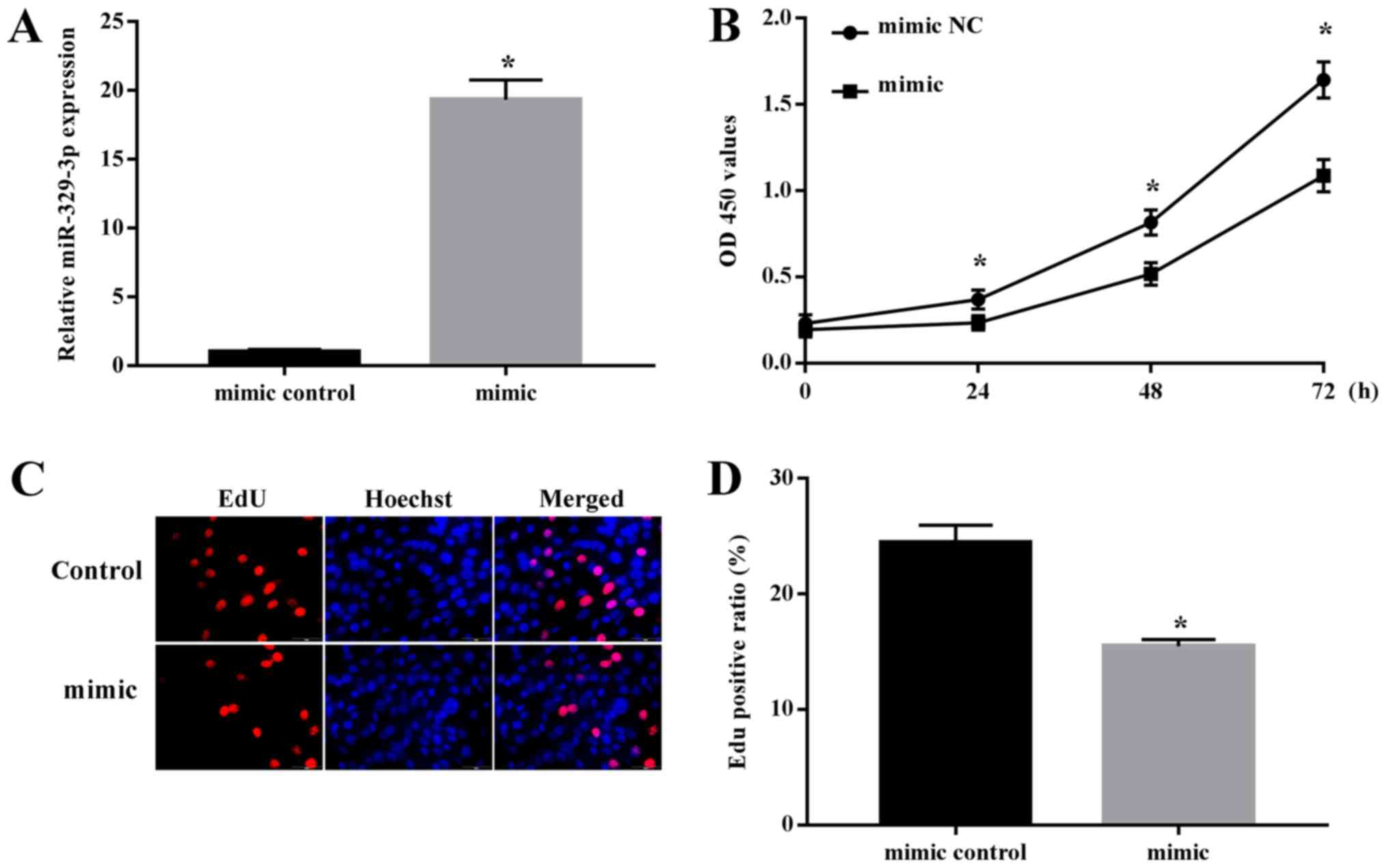
Upregulation of miR-329-3p suppresses
NSC proliferation
Previous data have suggested that the
differentiation of NSCs is accompanied by a concomitant decrease in
their ability to proliferate (17). As miR-329-3p levels gradually
increased during NSC differentiation, we hypothesized that this
miRNA may be involved in suppressing NSC proliferation. To examine
this, NSCs were transfected with miR-329-3p mimics or NC mimic and
cell proliferation was examined. RT-qPCR analysis confirmed that
miR-329-3p expression was significantly increased in NSCs
transfected with the miR-329-3p mimic compared with in those with
the NC mimic (Fig. 4A). Cell
proliferation was evaluated by the colorimetric CCK-8 assay and the
fluorescent EdU incorporation assay. Each of these assays indicated
that proliferation was decreased in cells transfected with the
miR-329-3p mimic compared with the level with the control mimic
(Fig. 4B-D), demonstrating that
miR-329-3p overexpression represses NSC proliferation.
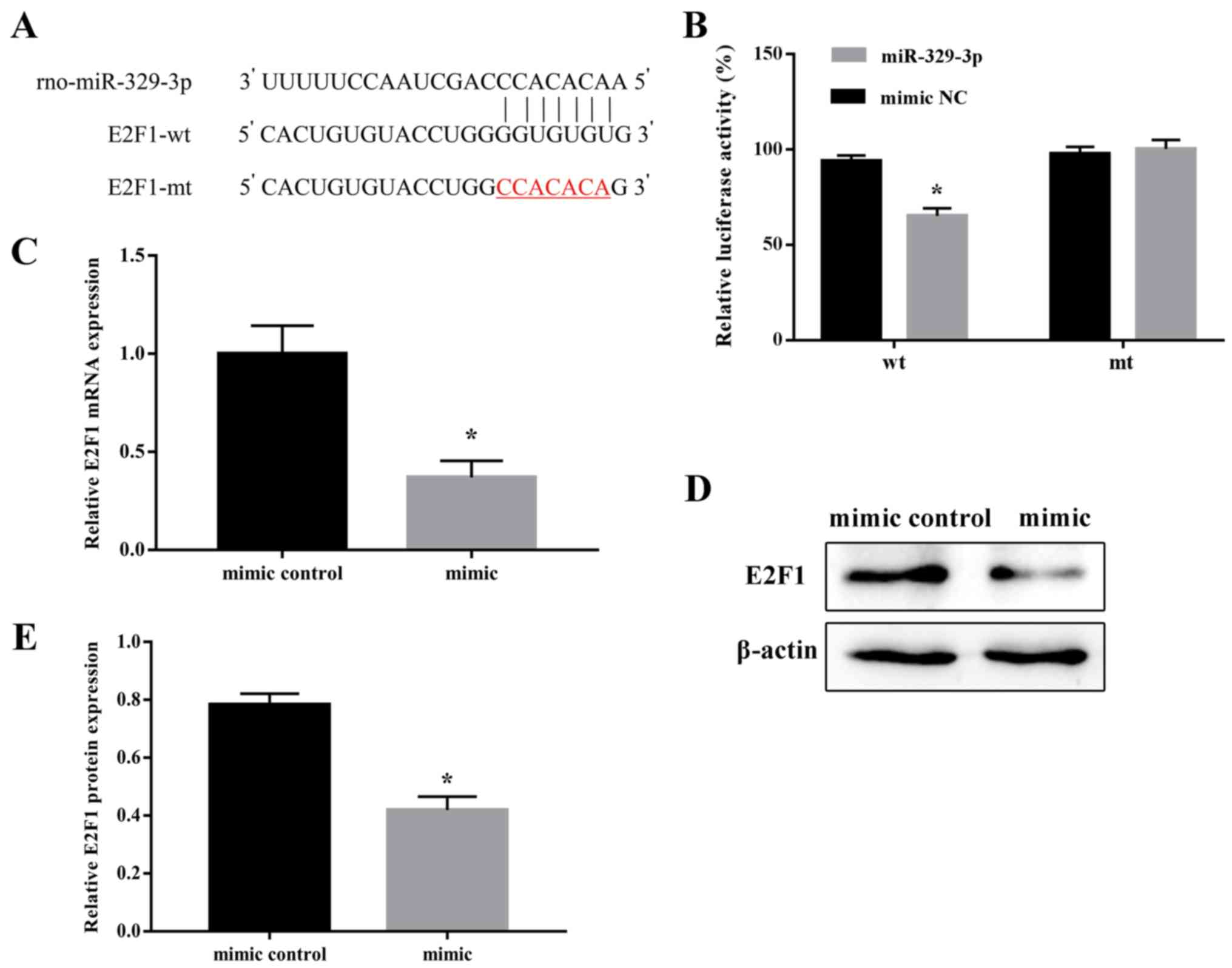
E2F1 is an miR-329-3p target gene
Screening of the bioinformatic databases TargetScan
(http://www.targetscan.org/) and miRDB
(http://mirdb.org/index.html) identified
a potential miR-329-3p-binding sequence in the 3′-UTR of E2F1
(Fig. 5A). To determine whether
E2F1 was a direct target of miR-329-3p, a dual-reporter assay was
performed, in which luciferase expression was driven by the wt or
mt form of the E2F1 3′-UTR sequence. For this, 293 cells were
co-transfected with the wt or mt E2F1 3′-UTR reporter vector
together with an miR-329-3p or NC mimic, and luciferase activity
was measured after 48 h. It was identified that the miR-329-3p
mimic significantly inhibited luciferase expression driven by the
wt, but not the mt, 3′-UTR sequence (Fig. 5B). These results confirmed that
miR-329-3p directly regulated E2F1 expression. To additionally
define the targeting association between miR-329-3p and E2F1, the
expression of E2F1 was examined following transfection of the
miR-329-3p mimic in NSCs. As expected, transfection of the
miR-329-3p mimic decreased the expression of E2F1 mRNA and protein
in NSCs (Fig. 5C-E).
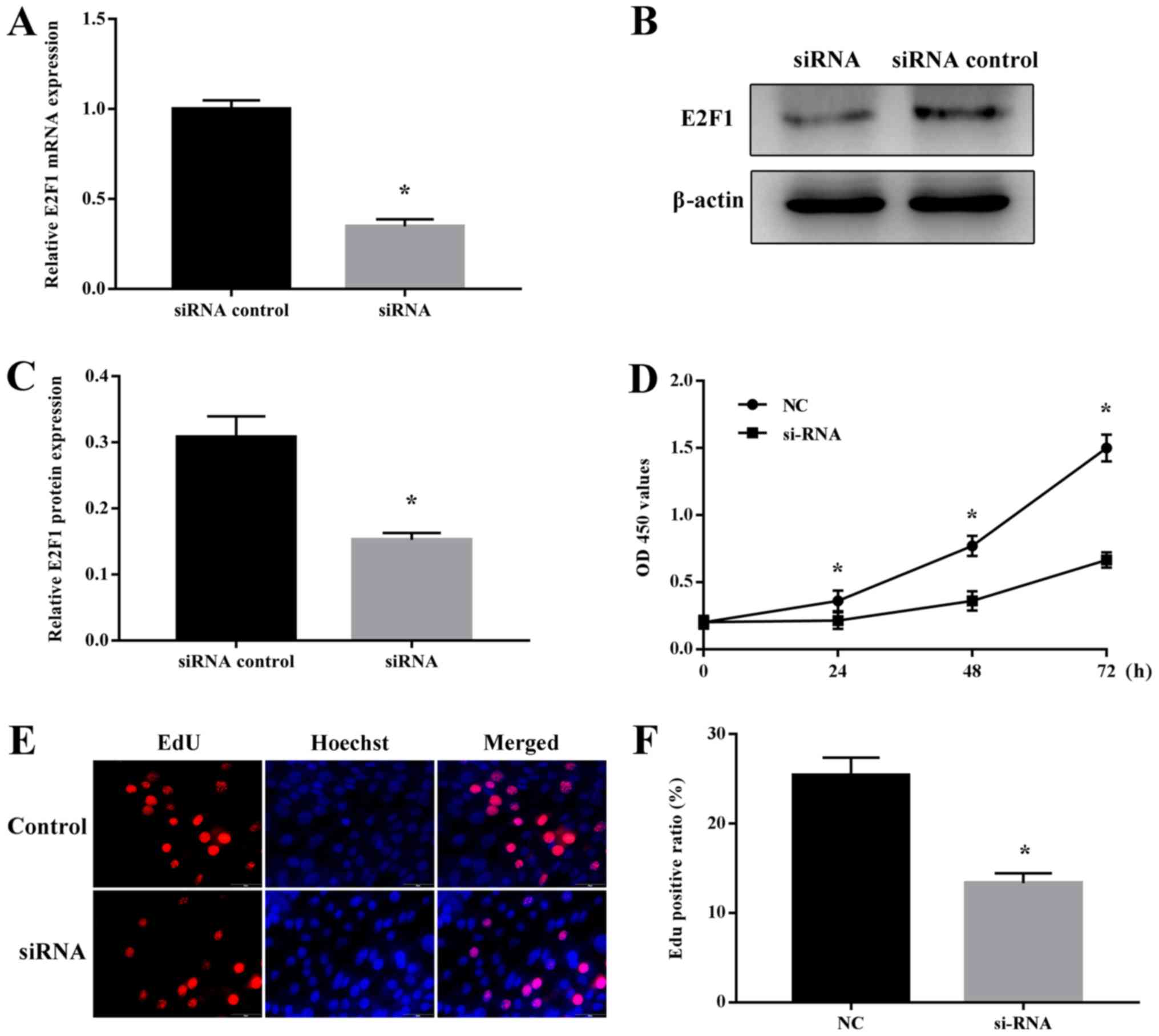
E2F1 regulates NSC proliferation
Several studies have demonstrated that E2F1 is
involved in the maintenance of stem cells (17–19).
To investigate the potential role of E2F1 in NSC proliferation,
cells were first transfected with E2F1-specific or NC siRNA, and it
was confirmed that E2F1 mRNA and protein expression was
specifically decreased by the E2F1-targeting siRNA (Fig. 6A-C). Notably, analysis of cell
proliferation using the CCK-8 and EdU assays revealed that E2F1
siRNA specifically inhibited the proliferation of NSCs (Fig. 6D-F). Therefore, E2F1 appeared to
serve a vital role in regulating NSC proliferation.
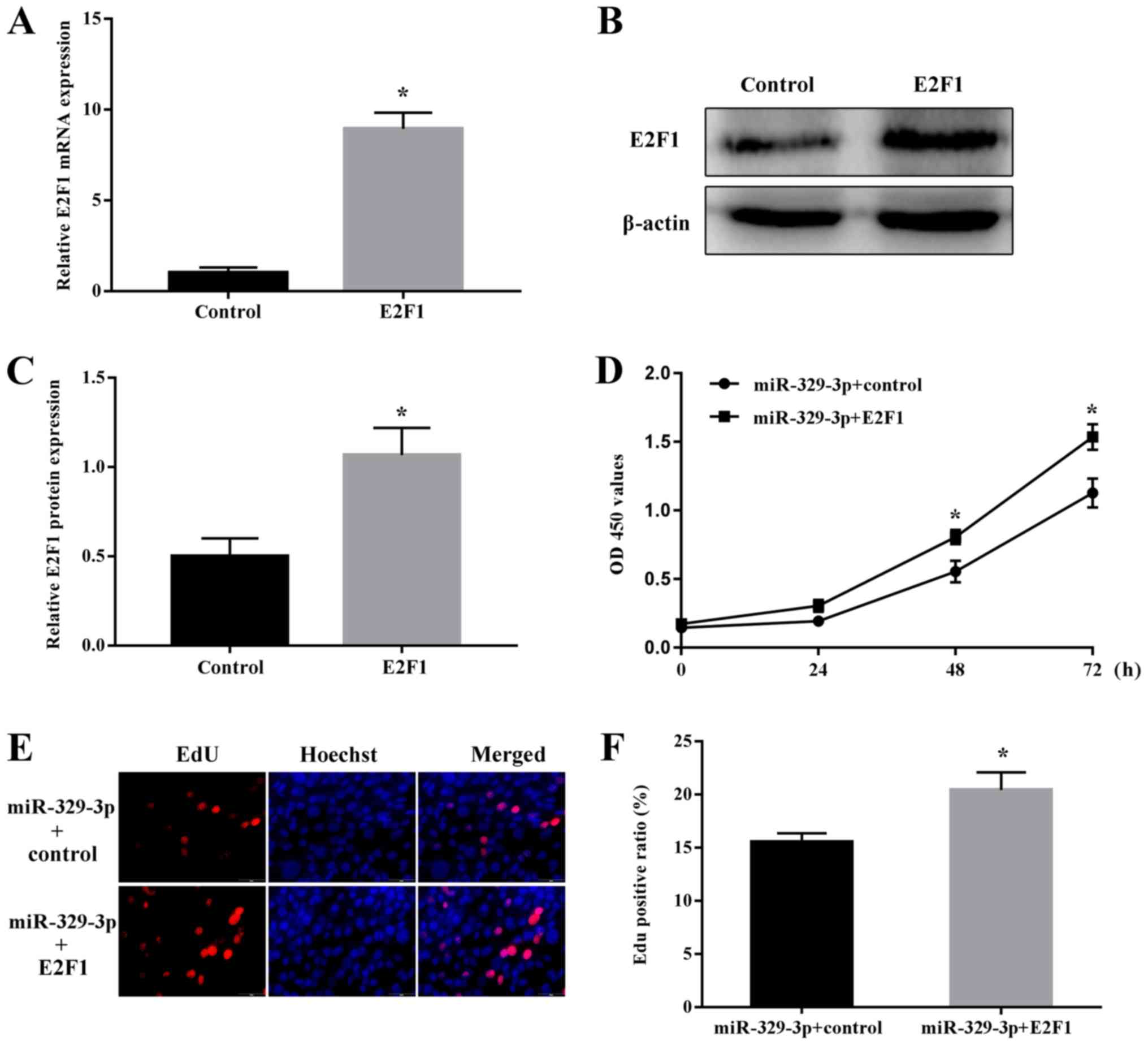
miR-329-3p regulates NSC proliferation
by targeting E2F1
Finally, the functional effects of
miR-329-3p-mediated regulation of E2F1 in NSCs were examined. Cells
were co-transfected with miR-329-3p together with the E2F1
overexpression vector (omitting the 3′-UTR sequence targeted by
miR-329-3p) or E2F1 vector control. Overexpression of E2F1 was
confirmed at the mRNA (Fig. 7A)
and protein levels (Fig. 7B and
C). Notably, ectopic expression of E2F1 reversed the decrease
in NSC proliferation induced by transfection with the miR-329-3p
mimic, as measured using the CCK-8 (Fig. 7D) and EdU incorporation assays
(Fig. 7E and F).
Discussion
NSCs have the capacity to proliferate, self-renew
and differentiate into neurons, astrocytes or oligodendrocytes
(20–22). The proliferation and
differentiation of NSCs are modulated by multiple regulators,
including miRNAs (17,23). For example, NSC proliferation and
differentiation are regulated by miR-125b, miR-378, miR-506-3p and
miR-1297 via modulation of Nestin, nuclear receptor subfamily 2
group E member 1, transcription factor E2-alpha and transcription
factor HES-1 expression, respectively (11,24–26).
It has also been demonstrated that multiple miRNAs interact
cooperatively to regulate cellular processes. For example, miR-124,
miR-128 and miR-137 were demonstrated to orchestrate biological
roles in NSCs by controlling the expression of the common target
gene Sp1 transcription factor (27). In addition, the abnormal expression
of miRNAs has been demonstrated to be associated with neurological
diseases including neuroblastoma and glioma (28,29).
In the GeneChip analysis in the present study, it
was identified that miR-329-3p was upregulated in differentiated
neurons compared with the level in NSCs. Previous studies have
suggested that miR-329-3p serves a key role in the proliferation of
various types of cancer (28–30).
For example, Yang et al (28) demonstrated that miR-329 inhibited
the growth and invasion of neuroblastoma cells by regulating
lysine-specific histone demethylase 1A expression. Xiao et
al (29) also demonstrated
that miR-329 expression was downregulated in glioma cells and that
its overexpression inhibited cell proliferation. In addition, Kang
et al (30) indicated that
downregulation of miR-329 promoted the development of human breast
cancer. However, the role of miR-329-3p in NSC biology remains
unclear. Therefore, miR-329-3p was selected for examination in the
present study. As the proliferative capacity of NSCs has been
demonstrated to decrease during differentiation (17), the present study initially examined
the expression of miR-329-3p during the differentiation of NSCs,
and it was identified that its expression gradually increased
during this process, which was consistent with the data from the
GeneChip analysis. Then, miR-329-3p was overexpressed in NSCs, and
it was demonstrated that this overexpression suppressed NSC
proliferation.
E2F1 is a member of the E2F family of transcription
factors and serves crucial roles in cell proliferation (17). E2F1 has been identified to be
closely associated with the growth of a variety of tumors (31–33).
For example, Lv et al (31)
demonstrated that E2F1 was involved in the growth and invasion of
hepatoma cells caused by aflatoxin B1. In addition, Sun et
al (32) revealed that the
expression of E2F1 was increased in lung cancer tissues and
asserted that E2F1 may be considered as a novel biomarker for the
prognosis of lung cancer. Zhang et al (33) also indicated that E2F1 promoted the
expression of lncRNA RGMB antisense RNA 1 (RGMB-AS1) by binding to
its promoter region, and that upregulated lncRNA RGMB-AS1 promoted
the proliferation, migration and invasion of papillary thyroid
carcinoma. It has also been suggested that E2F1 increased cell
proliferation by promoting transition through the G1/S checkpoint
of the cell cycle (34,35). Exiting the cell cycle is a crucial
step in cell differentiation (36), and E2F1 serves a key role in this
process (37,38). In concordance with this, NSC
differentiation is known to be accompanied by a decrease in
proliferation (17). In the
present study, it was identified that E2F1 expression was decreased
during NSC differentiation. The data demonstrating that the
downregulation of E2F1 inhibited the proliferation of NSCs was also
consistent with the upregulation of miR-329-3p. As the
overexpression of E2F1 restored the proliferative capacity of
miR-329-3p-overexpressing NSCs, the results from the present study
confirmed that miR-329-3p modulated NSC proliferation by repressing
E2F1 expression.
In conclusion, the present study revealed a novel
role for miR-329-3p in regulating NSC proliferation, in part
through the inhibition of E2F1 expression. These data provide
insight into the miRNA-mRNA regulatory network that controls NSC
proliferation, and suggest a new avenue of study into the functions
of miR-329-3p and E2F1 in the nervous system. In addition, the
results may contribute to the development of novel therapeutic
strategies based on NSC transplantation.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Plan Project of Liaoning Province (grant no.
201602857).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DL and GT designed the experiments. DL, YS, XD and
YH performed the experiments, and analyzed and collected the data.
DL wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CCK-8
|
Cell Counting Kit-8
|
|
E2F1
|
transcription factor E2F1
|
|
EdU
|
5-ethynyl-2′-deoxyuridine
|
|
miRNA/miR
|
microRNA
|
|
mt
|
mutated
|
|
NSC
|
neural stem cell
|
|
RT-qPCR
|
reverse-transcription quantitative
polymerase chain reaction
|
|
UTR
|
untranslated region
|
|
wt
|
wild-type
|
References
|
1
|
Liu S, Yin F, Zhang J, Wicha MS, Chang AE,
Fan W, Chen L, Fan M and Li Q: Regulatory roles of miRNA in the
human neural stem cell transformation to glioma stem cells. J Cell
Biochem. 115:1368–1380. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Makri G, Lavdas AA, Katsimpardi L,
Charneau P, Thomaidou D and Matsas R: Transplantation of embryonic
neural stem/precursor cells overexpressing BM88/Cend1 enhances the
generation of neuronal cells in the injured mouse cortex. Stem
Cells. 28:127–139. 2010.PubMed/NCBI
|
|
3
|
Mosher KI, Andres RH, Fukuhara T, Bieri G,
Hasegawa-Moriyama M, He Y, Guzman R and Wyss-Coray T: Neural
progenitor cells regulate microglia functions and activity. Nat
Neurosci. 15:1485–1487. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Le MT, Xie H, Zhou B, Chia PH, Rizk P, Um
M, Udolph G, Yang H, Lim B and Lodish HF: MicroRNA-125b promotes
neuronal differentiation in human cells by repressing multiple
targets. Mol Cell Biol. 29:5290–5305. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu X, Li Z, Shen J, Wu WK, Liang J, Weng X
and Qiu G: MicroRNA-10b promotes nucleus pulposus cell
proliferation through RhoC-Akt pathway by targeting HOXD10 in
intervetebral disc degeneration. PLoS One. 8:e830802013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu X, Li Z, Yu J, Chan MT and Wu WK:
MicroRNAs predict and modulate responses to chemotherapy in
colorectal cancer. Cell Prolif. 48:503–510. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Santra M, Chopp M, Santra S, Nallani A,
Vyas S, Zhang ZG and Morris DC: Thymosin beta 4 up-regulates
miR-200a expression and induces differentiation and survival of rat
brain progenitor cells. J Neurochem. 136:118–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan Z, Lu M, Qiao C, Zhou Y, Ding JH and
Hu G: MicroRNA-7 enhances subventricular zone neurogenesis by
inhibiting NLRP3/Caspase-1 axis in adult neural stem cells. Mol
Neurobiol. 53:7057–7069. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang JF, Shi LL, Zhang L, Zhao ZH, Liang
F, Xu X, Zhao LY, Yang PB, Zhang JS and Tian YF: MicroRNA-25
negatively regulates cerebral ischemia/reperfusion injury-induced
cell apoptosis through Fas/FasL pathway. J Mol Neurosci.
58:507–516. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bian S, Hong J, Li Q, Schebelle L, Pollock
A, Knauss JL, Garg V and Sun T: MicroRNA cluster miR-17-92
regulates neural stem cell expansion and transition to intermediate
progenitors in the developing mouse neocortex. Cell Rep.
3:1398–1406. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cui Y, Xiao Z, Han J, Sun J, Ding W, Zhao
Y, Chen B, Li X and Dai J: MiR-125b orchestrates cell
proliferation, differentiation and migration in neural
stem/progenitor cells by targeting nestin. BMC Neurosci.
13:1162012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Garg N, Po A, Miele E, Campese AF, Begalli
F, Silvano M, Infante P, Capalbo C, De Smaele E, Canettieri G, et
al: microRNA-17-92 cluster is a direct Nanog target and controls
neural stem cell through Trp53inp1. EMBO J. 32:2819–2832. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zou D, Chen Y, Han Y, Lv C and Tu G:
Overexpression of microRNA-124 promotes the neuronal
differentiation of bone marrow-derived mesenchymal stem cells.
Neural Regen Res. 9:1241–1248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wong N and Wang X: miRDB: An online
resource for microRNA target prediction and functional annotations.
Nucleic Acids Res. 43:D146–152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
ELife. 4:2015. View Article : Google Scholar
|
|
17
|
Palm T, Hemmer K, Winter J, Fricke IB,
Tarbashevich K, Sadeghi Shakib F, Rudolph IM, Hillje AL, De Luca P,
Bahnassawy L, et al: A systemic transcriptome analysis reveals the
regulation of neural stem cell maintenance by an E2F1-miRNA
feedback loop. Nucleic Acids Res. 41:3699–3712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pulikkan JA, Dengler V, Peramangalam PS,
Peer Zada AA, Müller-Tidow C, Bohlander SK, Tenen DG and Behre G:
Cell-cycle regulator E2F1 and microRNA-223 comprise an
autoregulatory negative feedback loop in acute myeloid leukemia.
Blood. 115:1768–1778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Han BW, Feng DD, Li ZG, Luo XQ, Zhang H,
Li XJ, Zhang XJ, Zheng LL, Zeng CW, Lin KY, et al: A set of miRNAs
that involve in the pathways of drug resistance and leukemic
stem-cell differentiation is associated with the risk of relapse
and glucocorticoid response in childhood ALL. Hum Mol Genet.
20:4903–4915. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao C, Deng W and Gage FH: Mechanisms and
functional implications of adult neurogenesis. Cell. 132:645–660.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brett JO, Renault VM, Rafalski VA, Webb AE
and Brunet A: The microRNA cluster miR-106b~25 regulates adult
neural stem/progenitor cell proliferation and neuronal
differentiation. Aging (Albany NY). 3:108–124. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi Y, Sun G, Zhao C and Stewart R: Neural
stem cell self-renewal. Crit Rev Oncol Hematol. 65:43–53. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rybak A, Fuchs H, Smirnova L, Brandt C,
Pohl EE, Nitsch R and Wulczyn FG: A feedback loop comprising lin-28
and let-7 controls pre-let-7 maturation during neural stem-cell
commitment. Nat Cell Biol. 10:987–993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang Y, Liu X and Wang Y: MicroRNA-378
regulates neural stem cell proliferation and differentiation in
vitro by modulating Tailless expression. Biochem Biophys Res
Commun. 466:214–220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Jiaqi C, Zhaoying C and Huimin C:
MicroRNA-506-3p regulates neural stem cell proliferation and
differentiation through targeting TCF3. Gene. 593:193–200. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng J, Yi D, Shi X and Shi H: miR-1297
regulates neural stem cell differentiation and viability through
controlling Hes1 expression. Cell Prolif. 50:2017. View Article : Google Scholar :
|
|
27
|
Santos MC, Tegge AN, Correa BR, Mahesula
S, Kohnke LQ, Qiao M, Ferreira MA, Kokovay E and Penalva LO:
miR-124, −128, and −137 orchestrate neural differentiation by
acting on overlapping gene sets containing a highly connected
transcription factor network. Stem Cells. 34:220–232. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang H, Li Q, Zhao W, Yuan D, Zhao H and
Zhou Y: miR-329 suppresses the growth and motility of neuroblastoma
by targeting KDM1A. FEBS Lett. 588:192–197. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiao B, Tan L, He B, Liu Z and Xu R:
MiRNA-329 targeting E2F1 inhibits cell proliferation in glioma
cells. J Transl Med. 11:1722013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kang H, Kim C, Lee H, Rho JG, Seo JW, Nam
JW, Song WK, Nam SW, Kim W and Lee EK: Downregulation of
microRNA-362-3p and microRNA-329 promotes tumor progression in
human breast cancer. Cell Death Differ. 23:484–495. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lv J, Yu YQ, Li SQ, Luo L and Wang Q:
Aflatoxin B1 promotes cell growth and invasion in hepatocellular
carcinoma HepG2 cells through H19 and E2F1. Asian Pac J Cancer
Prev. 15:2565–2570. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun CC, Zhou Q, Hu W, Li SJ, Zhang F, Chen
ZL, Li G, Bi ZY, Bi YY, Gong FY, et al: Transcriptional E2F1/2/5/8
as potential targets and transcriptional E2F3/6/7 as new biomarkers
for the prognosis of human lung carcinoma. Aging (Albany NY).
10:973–987. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Z, Li SY and Zhang LB: LncRNA
RGMB-AS1 is activated by E2F1 and promotes cell proliferation and
invasion in papillary thyroid carcinoma. Eur Rev Med Pharmacol Sci.
22:1979–1986. 2018.PubMed/NCBI
|
|
34
|
DeGregori J, Kowalik T and Nevins JR:
Cellular targets for activation by the E2F1 transcription factor
include DNA synthesis- and G1/S-regulatory genes. Mol Cell Biol.
15:4215–4224. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Inoshita S, Terada Y, Nakashima O,
Kuwahara M, Sasaki S and Marumo F: Regulation of the G1/S
transition phase in mesangial cells by E2F1. Kidney Int.
56:1238–1241. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma Q, Hu QS, Xu RJ, Zhen XC and Wang GH:
Protease Omi facilitates neurite outgrowth in mouse neuroblastoma
N2a cells by cleaving transcription factor E2F1. Acta Pharmacol
Sin. 36:966–975. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McClellan KA and Slack RS: Novel functions
for cell cycle genes in nervous system development. Cell Cycle.
5:1506–1513. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tsume M, Kimura-Yoshida C, Mochida K,
Shibukawa Y, Amazaki S, Wada Y, Hiramatsu R, Shimokawa K and Matsuo
I: Brd2 is required for cell cycle exit and neuronal
differentiation through the E2F1 pathway in mouse neuroepithelial
cells. Biochem Biophys Res Commun. 425:762–768. 2012. View Article : Google Scholar : PubMed/NCBI
|