Introduction
Intrauterine adhesion (IUA) is an acquired uterine
condition characterised by the formation of scar tissue inside the
uterine cavity, which, in many cases, results in adherence to the
opposing endometrium (1,2). Risk factors including age,
myomectomy, obesity, delivery, pelvic infections and genital
tuberculosis can increase the morbidity of IUA (1), and IUA can cause amenorrhea, abnormal
uterine bleeding, sterility, consecutive spontaneous abortions,
menstruation and abnormal placentation (3,4).
Over the past two decades, the increased use of curettage and/or
dilation has led to an increased prevalence of IUA. Furthermore,
the recurrence of adhesion remains high, and curing IUA is
challenging in patients with moderate to severe IUA (5).
Vaginal secretions and vaginal epithelial cells
provide a rich source of nutrients that support bacterial growth
(6), and the vaginal microbiota is
made up of an extensive and varied spectrum of pathogenic and
non-pathogenic organisms (7). The
bacterial population present in the lower genital tract of females
plays a key role in maternal and neonatal health. The normal
vaginal microbiota in healthy women should be dominated by
Lactobacillus species, with abnormal microbiota
characterised by a low number of lactobacilli and a high number of
anaerobic bacteria, such as Gardnerella vaginalis,
Prevotella and Mobiluncus (8,9).
Moreover, previous studies have indicated that vaginal
dysbacteriosis is strongly related to postpartum endometritis,
preterm delivery, pelvic inflammatory diseases, spontaneous
abortion and the delivery of low birth weight infants (10–12).
Based on our knowledge, the pathological changes of
IUA are bound to influence the physiology and metabolites in the
uterus, which will cause side effects in the adjacent vaginal
tissue and influence the vaginal microbial diversity. Therefore,
whether IUA could also disrupt the microbial composition in the
vagina, and if the presence of certain bacteria in the vaginal
tract could affect the pathogenic condition of IUA was
investigated. To answer these questions, 80 women with and without
IUA were studied. Specifically, the microbial diversity between
healthy women and women with moderate IUA were compared using
high-throughput sequencing.
Patients and methods
Ethics statement
This study was approved by the Institutional Review
Boards of The Second Affiliated Hospital of Nanchang University
(Nanchang, China). Patients provided written informed consent for
sample collection.
Study groups and sampling
The trial enrolled 50 women aged between 16 and 33
years who were newly diagnosed with IUA during hysteroscopy
examination between November 2017 and June 2018 at The Second
Affiliated Hospital of Nanchang University, based on the revised
criteria of the American Fertility Society, [intrauterine adhesion
vaginal secretion (IAVS) group]. Patients had not used vaginal
medications, received cervical treatment or performed douching
within the previous 7 days, and had not engaged in sexual activity
within the previous 2 days. Thirty healthy women (15–35 years old;
mean age, 29.46 years) were recruited as the control group between
November 2017 and June 2018 at The Second Affiliated Hospital of
Nanchang University [healthy vaginal secretion (HVS) group].
Participants had no diagnosed endocrine or autoimmune disorders,
cancer, severe pelvic adhesion, hysteromyoma, endometriosis,
adenomyosis or acute inflammation. Self-administered vaginal swabs
were used to collect vaginal specimens, which were immediately
stored at −80°C for DNA extraction (Table I).
 | Table I.Baseline patient demographics and
characteristics. |
Table I.
Baseline patient demographics and
characteristics.
| Variable | HVS group
(n=30) | IAVS group
(n=50) |
|---|
| Percentage of total
enrollment, no. (%) | 30 (37.50) | 50 (62.50) |
| Age [years, mean
(SD)] | 29.46±2.19 | 28.38±1.27 |
| Age at first sexual
intercourse (years) |
|
|
|
<18 | 5 (16.67) | 10 (20.00) |
|
18-22 | 13 (43.33) | 19 (38.00) |
|
>22 | 12 (40.00) | 21 (42.00) |
| No. of
abortions/complete curettage of uterine cavity |
|
|
| 1 | 9 (30.00) | 15 (30.00) |
| 2 | 13 (43.33) | 23 (46.00) |
| 3 | 8 (26.27) | 12 (24.00) |
| No. of
deliveries |
|
|
| 0 | 10 (33.33) | 16 (32.00) |
| 1 | 12 (40.00) | 22 (44.00) |
|
>2 | 8 (26.27) | 12 (24.00) |
| Degree of uterine
adhesion, no. (%) |
|
|
|
0-grade | 30 (100) | 0 (0) |
|
Low-grade | 0 (0) | 18 (36.00) |
|
Middle-grade | 0 (0) | 27 (54.00) |
|
High-grade | 0 (0) | 5 (10.00) |
Extraction of genomic DNA and
high-throughput sequencing
For the extraction of bacterial DNA from vaginal
samples, the combination of genomic DNA kits (Tiangen Biotech Co.,
Ltd., Beijing, China) and the bead beating method were used
(13), and the concentration and
quality of purified DNA was determined via a spectrophotometer at
230 nm (A230) and 260 nm (A260; NanoDrop; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). The V4 region of the 16S ribosomal (r)DNA
genes in each sample was amplified using SYBR Green Master mix
(Qiagen GmbH, Hilden, Germany) and 515F/806R primers (515F,
5′-GTGCCAGCMGCCGCGGTAA-3′; 806R, 5′-GGACTACVSGGGTATCTAAT-3′). PCR
was conducted as follows: 98°C for 2 min, followed by 30 cycles of
98°C for 15 sec, 55°C for 30 sec and 72°C for 30 sec. PCR products
were sequenced with an IlluminaHiSeq 2000 platform (GenBank
accession no. SRP155123; Illumina, Inc., San Diego, CA, USA)
(14).
Bioinformatics and multivariate
statistical analysis
Paired-end reads from the original DNA fragments
were processed using Cutadapt (version 1.9.1, http://cutadapt.readthedocs.io/en/stable/) and UCHIME
Algorithm (http://www.drive5.com/usearch/manual/uchime_algo.html)
(15). Sequence analysis was
subsequently performed using the UPARSE software package (version
7.0.100, http://drive5.com/uparse), and sequences
with ≥97% similarity were assigned to the same operational
taxonomic units (OTU). Then, QIIME software (version 1.9.1,
http://qiime.org/) was used to analyse the α-diversity
(within samples, indexes of observed-OTUs, Chao1, Shannon, Simpson,
abundance-based coverage estimator metric, good's-coverage) and the
β-diversity [among samples, principal component analysis, principal
coordinates analysis (PCoA) and nonmetric multidimensional scaling]
(16,17). The cluster analysis was preceded by
determination of the weighted UniFrac distance using the QIIME
software package (version 1.8.0) (18), and partial least squares
discriminate analysis (PLS-DA) was preceded by the use of SIMCA-P
software (version 11.5; Umetrics; Sartorius Stedim Biotech, Malmö,
Sweden), and differently abundant taxa identifications were
compared using linear discriminant analysis effect size (LEfSe)
analysis (Galaxy; http://huttenhower.sph.harvard.edu/galaxy/) (19). The statistical significance was set
at P<0.05 for correction of multiple comparisons.
Results
Baseline characteristics of
participants
Between November 2017 and June 2018, 80 women were
recruited to either the IAVS group (50 patients with IUA) or HVS
group (30 healthy women), and the baseline characteristics of
patients in the two groups were similar (Table I).
All participants were thoroughly informed about
their conditions, and the IAVS and HVS groups were well balanced
with no marked differences. The age, age at first sexual
intercourse, number of abortions, complete curettage of the uterine
cavity, number of deliveries and degree of uterine adhesion are
summarised in Table I. The vaginal
samples of women with no IUA and those with mid-grade IUA were used
for high-throughput sequencing.
Sequencing coverage
To compare the microbial diversity between HVS and
IAVS groups, 16S rRNA amplicon sequencing analysis was applied to
sequence the V4 hypervariable region of bacteria. Data were
obtained by filtering the raw data, and sequences with >97%
similarity were cultured to the same OTU. In total, 3,051,883
filtered clean reads (76,297.08 reads/sample) and 12,266 OTUs were
obtained from all samples, with an average of 306.65 OTUs per group
(Table II).
| Table II.Number of raw reads, clean reads,
observed species and effective in groups HVS and IAVS by
high-throughput sequencing. |
Table II.
Number of raw reads, clean reads,
observed species and effective in groups HVS and IAVS by
high-throughput sequencing.
| Sample name | Raw reads | Clean reads | Observed
species | Effective (%) |
|---|
| HVS1 | 82299 | 80306 | 529 | 97.58 |
| HVS2 | 91596 | 86891 | 900 | 94.86 |
| HVS3 | 77376 | 76554 | 164 | 98.94 |
| HVS4 | 55907 | 55314 | 264 | 98.94 |
| HVS5 | 86040 | 80337 | 370 | 93.37 |
| HVS6 | 72413 | 70253 | 458 | 97.02 |
| HVS7 | 84510 | 80429 | 163 | 95.17 |
| HVS8 | 84881 | 80177 | 252 | 94.46 |
| HVS9 | 83311 | 80259 | 295 | 96.34 |
| HVS10 | 84538 | 80185 | 218 | 94.85 |
| HVS11 | 66322 | 64824 | 167 | 97.74 |
| HVS12 | 82364 | 80271 | 349 | 97.46 |
| HVS13 | 58278 | 57101 | 485 | 97.98 |
| HVS14 | 78482 | 77649 | 148 | 98.94 |
| HVS15 | 101853 | 98324 | 193 | 96.54 |
| HVS16 | 81362 | 80314 | 143 | 98.71 |
| HVS17 | 65697 | 63227 | 127 | 96.24 |
| HVS18 | 83160 | 80320 | 187 | 96.58 |
| HVS19 | 83135 | 80132 | 195 | 96.39 |
| HVS20 | 83759 | 80318 | 499 | 95.89 |
| IAVS1 | 80796 | 80112 | 156 | 99.15 |
| IAVS2 | 68475 | 64205 | 667 | 93.76 |
| IAVS3 | 85975 | 80132 | 230 | 93.2 |
| IAVS4 | 67704 | 65111 | 183 | 96.17 |
| IAVS5 | 84329 | 80170 | 271 | 95.07 |
| IAVS6 | 80902 | 80247 | 265 | 99.19 |
| IAVS7 | 84042 | 83435 | 283 | 99.28 |
| IAVS8 | 67329 | 66696 | 397 | 99.06 |
| IAVS9 | 84233 | 81691 | 191 | 96.98 |
| IAVS10 | 68959 | 64354 | 179 | 93.32 |
| IAVS11 | 78713 | 75827 | 228 | 96.33 |
| IAVS12 | 79026 | 75605 | 338 | 95.67 |
| IAVS13 | 81928 | 80129 | 296 | 97.8 |
| IAVS14 | 61256 | 57481 | 190 | 93.84 |
| IAVS15 | 71488 | 69808 | 379 | 97.65 |
| IAVS16 | 85869 | 80434 | 314 | 93.67 |
| IAVS17 | 84245 | 80046 | 244 | 95.02 |
| IAVS18 | 84923 | 80215 | 310 | 94.46 |
| IAVS19 | 97284 | 93566 | 781 | 96.18 |
| IAVS20 | 80790 | 79434 | 258 | 98.32 |
| Total | 3165549 | 3051883 | 12266 | / |
| Average | 79138.73 | 76297.08 | 306.65 | 96.45 |
α-Diversity of the microbial community
in the HVS and IAVS groups
As presented in Fig.
1, the results for the observed species, Shannon index and
Simpson index indicated that IUA had little effect on the
α-diversity of the vaginal microbial community between HVS and IAVS
groups, with 1,540 and 1,360 OTUs in the HVS and IAVS groups,
respectively. The percentage of common OTUs was 71.95%
(1,108/1,540) and 81.47% (1,108/1,360), respectively.
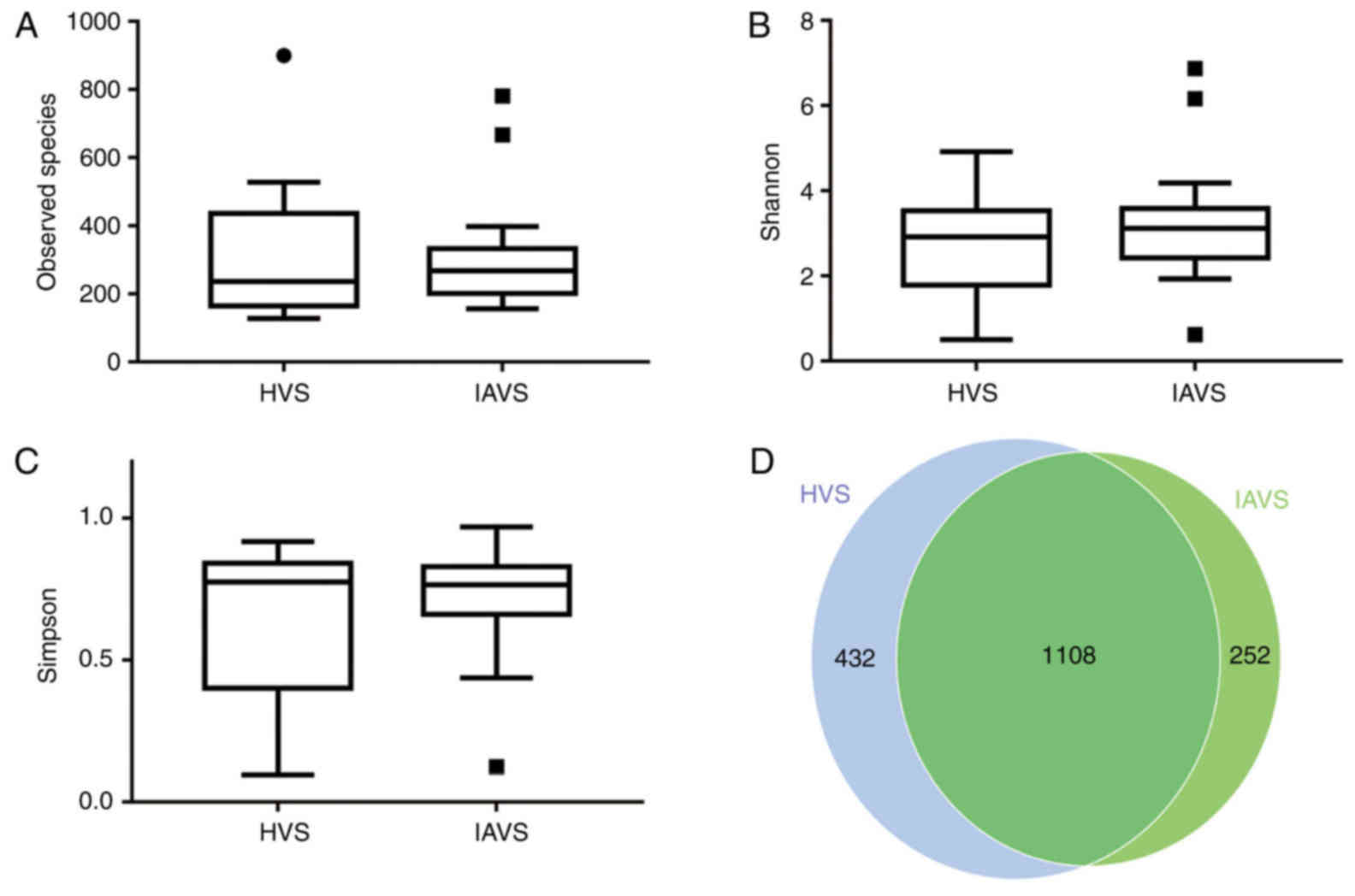 | Figure 1.Effects of intrauterine adhesion on
the α-diversity of the vaginal microbial community. α-Diversity
distances between the HVS and IAVS groups, revealing (A) the
observed species, (B) the Shannon index, (C) the Simpson index and
the (D) Scalar-Venn representation. The α-diversity distances did
not significantly differ between the HVS and IAVS groups, and the
Venn results indicated that there were 1,540 and 1,360 OTUs in the
HVS and IAVS groups, respectively, with a total OTU number of
1,108. Circles and squares represent outliers. HVS, healthy vaginal
secretion; IAVS, intrauterine adhesion patients' vaginal secretion;
OTU, operational taxonomic units. |
Composition of the microbial community
in the HVS and IAVS groups at the phylum level
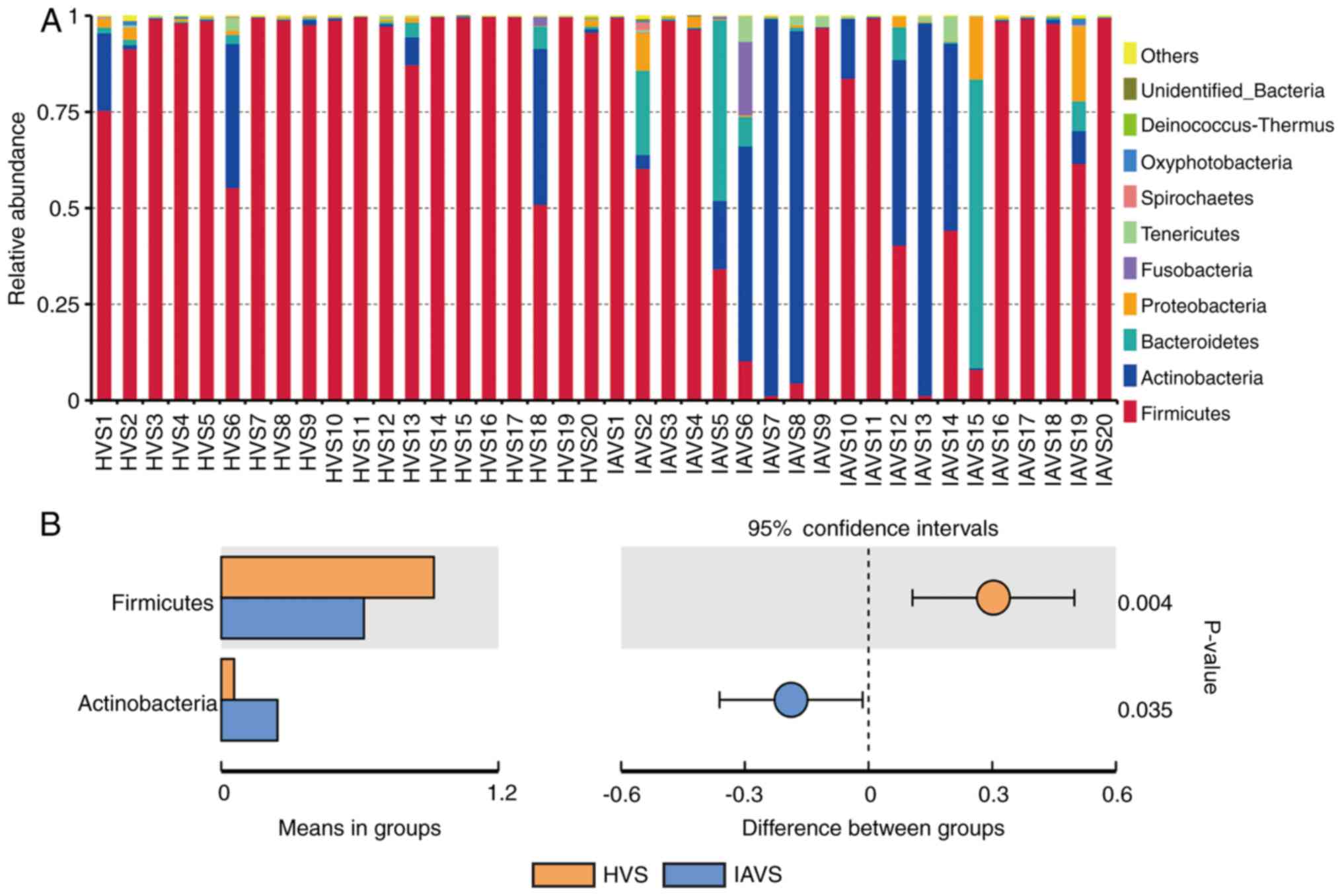
As presented in Fig.
2, data for the top 10 microorganism populations at the phylum
level were analysed. At the phylum level, Firmicutes,
Actinobacteria, Bacteroidetes and Proteobacteria constituted the
four most predominant phyla in the HVS (92.12, 5.58, 0.92 and
0.64%, respectively) and IAVS (61.84, 24.37, 8.64 and 2.74%,
respectively) groups, which accounted for 99.26 and 97.58% of the
total sequences in the HVS and IAVS groups, respectively. In the
HVS group, the percentage of Firmicutes was over 90% in most
samples except for HVS1, HVS6 and HVS18, while only few Firmicutes
were detected in samples IAVS5, IAVS6, IAVS7, IAVS8, IAVS13 and
IAVS15. In addition, a marked increase in Actinobacteria and
Bacteroidetes was observed in the IAVS group, particularly
Actinobacteria in samples IAVS6, IAVS7, IAVS8, IAVS12, IAVS13 and
IAVS14, and Bacteroidetes in samples IAVS5, IAVS6, IAVS12, IAVS15
and IAVS19. The statistical analysis also indicated that the HVS
group had a significantly higher percentage of Firmicutes and a
significantly lower percentage of Actinobacteria compared to the
IAVS group (P<0.05; Fig.
2B).
Composition of the microbial community
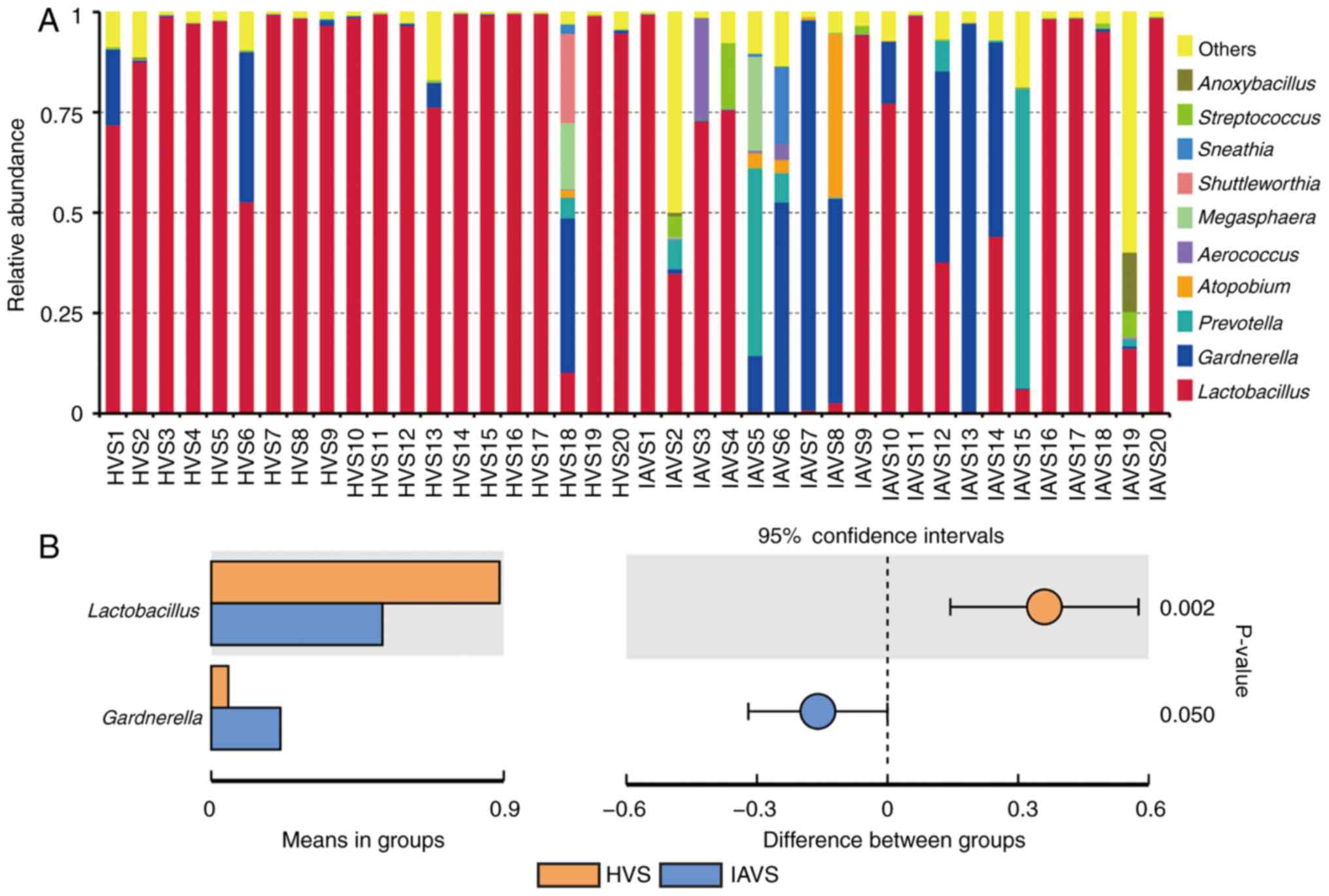
in the HVS and IAVS groups at the genus level
Similar to Fig. 2,
the top 10 genera were analysed, with Lactobacillus revealed
to account for >97% in most samples in the HVS group (Fig. 3A). For patients with IUA, the
uterine disorder markedly reduced the percentage of
Lactobacillus, but greatly enhanced the percentage of
Gardnerella and Prevotella. Statistical analysis
revealed a significant difference in Lactobacillus and
Gardnerella between the HVS and IAVS groups (P<0.05;
Fig. 3B). Notably, some samples
(IAVS9, IAVS11, IAVS16, IAVS17, IAVS18 and IAVS20) also possessed a
high percentage of Lactobacillus.
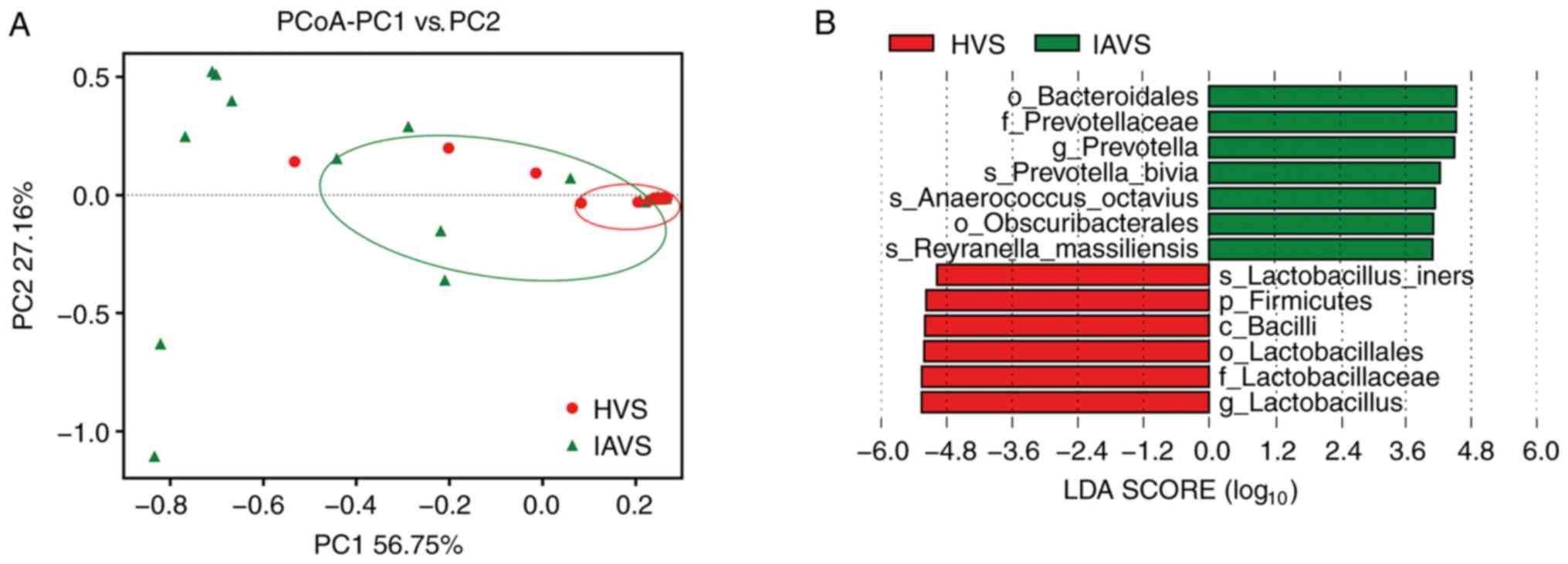
β-Diversity of the microbial community
in the HVS and IAVS groups
PCoA of the HVS and IAVS groups was carried out to
explore differences in the microbial diversity between groups. As
presented in Fig. 4, most samples
in the HVS group were clustered together on the right, while 50% of
samples (10/20) were scattered far away from the HVS group,
indicating that IUA greatly altered the vaginal microbial diversity
of IUV patients. In addition, the LEfSe analysis indicated that
Bacteroidales (order), Prevotellaceae (family), Prevotella
(genus), Prevotella bivia (species), Anaerococcus
octavius (species), Obscuribacterales (order) and Reyranella
massilliensis (species) were markedly higher in the IAVS group
(P<0.05), while Lactobacillus iners (species), Firmicutes
(phylum), Bacilli (class), Lactobacillales (order),
Lactobacillaceae (family) and Lactobacillus (genus) were
significantly higher in the HVS group (P<0.05).
Discussion
As one of the most common diseases of the
reproductive system in fertile women, the incidence of IUA is
leading to an increase in intrauterine surgeries (e.g.,
hysteromyomectomy, dilation and curettage) (20). Although transcervical resection of
the adhesion has been widely applied in the treatment of moderate
and severe IUA (5), the high
postoperative recurrence rates and low pregnancy rates presents a
great challenge for clinical management (21).
Although the uterus is considered a sterile tissue,
serious pathological changes of the uterine tissue of patients with
IUA have a great influence on the blood supply, inflammatory and
immune status and homeostasis of the uterine tissue (20). In addition, systemic disorders
undoubtedly influence the microbial composition of the vagina. To
date, many studies have been carried out to study the role of the
vaginal microbiota in cancer, vaginal infection, abortion,
sterility and menstrual disorders (9,22–27);
however, no studies have explored the interaction between IUA and
the vaginal microbiota.
In the present study, high-throughput sequencing was
used to evaluate the effect of IUA on vaginal microbial diversity.
A total of 80 fertile women (50 patients with IUA and 30 healthy
women) were recruited, and vaginal samples of 20 healthy women (HVS
group) and 20 mid-grade patients with IUA (IAVS group) were used
for microbial evaluation. A total of 12,266 OTUs were obtained from
all samples, and the average OTU number in each group was 306.65
(Table II). Although the Venn
results indicated that the OTU numbers in the HVS and IAVS groups
were 1,540 and 1,360, respectively, the findings for α-diversity of
the observed species, Shannon index and Simpson index indicated
that there was no significant difference between the groups.
Therefore, IUA did not alter the microbial species between healthy
women and those with IUA (Fig.
1).
When microbial communities were compared between the
HVS and IAVS groups at the phylum level, it was observed that
Firmicutes and Actinobacteria were the predominant phyla in the two
groups. Firmicutes was markedly higher in the HVS group than in the
IAVS group, while Actinobacteria was significantly lower in this
group (P<0.05; Fig. 2). The
Firmicutes usually have a Gram-positive cell wall structure, and
most Firmicutes can produce endospores to defend against
desiccation in extreme conditions. Thus, this phylum can be found
in various environments, and is known to play an important role in
beer, wine and cider spoilage (28). Moreover, Firmicutes constitute the
largest portion of the mouse and human gut microbiota involved in
energy resorption, and previous studies have confirmed that
Firmicutes are part of a normal, healthy placental microbiome
(29). Actinobacteria is another
phylum of Gram-positive bacteria, and they are of great economic
importance to humans due to their role in agriculture and forests,
specifically their contribution to soil systems; however, some
genera living in human faeces and vaginal sections have been
reported to be harmful for human health (11,23–25,30).
At the genus level, Lactobacillus was clearly
the dominant bacteria in the vaginal samples of healthy women, with
a marked reduction in their percentage in the IAVS group
accompanied by the overgrowth of pathogenic Gardnerella and
Prevotella genera (P<0.05; Fig. 3). As in the intestines, disruption
of the vaginal microbiota can lead to infection (31–33),
and previous studies have consistently indicated that vaginal
microbiota dominated by Lactobacillus was linked to good
vaginal health (31,34). Lactobacilli in the vagina could
protect the female urogenital tract against pathogen colonisation,
and these bacteria can protect the female genitourinary tract
against infections and help maintain a healthy genital system
(9,35). Therefore, the significant reduction
in Lactobacillus and overgrowth of Gardnerella and
Prevotella observed in patients with IUA would disrupt the
microbial homeostasis; however, 7 patients with IUA still possessed
a high number of Lactobacilli in their vaginal samples, indicating
that vaginal microbiota disorder only occurred in some patients
with IUA. Future studies will focus on the differences in prognosis
and recurrence between patients with IUA with high and low
percentages of Lactobacillus, initially in an animal model
and subsequently in volunteers. The β-diversity between HVS and
IAVS groups was also compared using PCoA analysis, and it was
revealed that ~50% of samples in the IAVS group (10/20) were
scattered far away from the HVS groups, indicating that IUA altered
the ratio of certain bacteria (Fig.
4).
In the present study, our group firstly explored the
interaction between IUA and the vaginal microbiota using
high-throughput sequencing technology, revealing that IUA
significantly reduced the percentage of Lactobacillus and
significantly increased Gardnerella and Prevotella in
~50% of patients with IUA, which may worsen the degree of IUA and
increase the risk of recurrence. Therefore, supplementation of
vaginal Lactobacillus during IUA treatment may help
accelerate recovery and reduce the recurrence of IUA. However, due
to the limited sample size of patients in the current study, a
larger number of patients is required to obtain a more confident
result.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81503364 and
31560264), the Excellent Youth Foundation of JiangXi Scientific
Committee (grant no. 20171BCB23028) and the Science and Technology
Plan of Jianxi Health Planning Committee (grant no. 20175526).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
TC, ZL and XD designed the experiments, analyzed the
data and wrote the manuscript. YK, YG, YR and CZ performed the
experiments. All authors discussed the results and commented on the
final manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the study are
appropriately investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Institutional Review
Boards of the Second Affiliated Hospital of Nanchang University.
Patient samples were obtained with written informed consent in
accordance with the Ethics Committee's requirements.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Salma U, Xue M, Sheikh SA, Guan X, Xu B,
Zhang A, Huang L and Xu D: Role of transforming growth factor-β1
and smads signaling pathway in intrauterine adhesion. Mediators
Inflamm. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chi Y, He P, Lei L, Lan Y, Hu J, Meng Y
and Hu L: Transdermal estrogen gel and oral aspirin combination
therapy improves fertility prognosis via the promotion of
endometrial receptivity in moderate to severe intrauterine
adhesion. Mol Med Rep. 17:6337–6344. 2018.PubMed/NCBI
|
|
3
|
Schenker JG: Etiology of and therapeutic
approach to synechia uteri. Eur J Obstet Gynecol Reprod Biol.
65:109–113. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Menzies D: Postoperative adhesions: Their
treatment and relevance in clinical practice. Ann R Coll Surg Engl.
75:147–153. 1993.PubMed/NCBI
|
|
5
|
Pabuccu R, Onalan G, Kaya C, Selam B,
Ceyhan T, Ornek T and Kuzudisli E: Efficiency and pregnancy outcome
of serial intrauterine device-guided hysteroscopic adhesiolysis of
intrauterine synechiae. Fertil Steril. 90:1973–1977. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nunn KL and Forney LJ: Unraveling the
dynamics of the human vaginal microbiome. Yale J Biol Med.
89:331–337. 2016.PubMed/NCBI
|
|
7
|
Chen H, Luo T, Chen T and Wang G: Seminal
bacterial composition in patients with obstructive and
non-obstructive azoospermia. Exp Ther Med. 15:2884–2890.
2018.PubMed/NCBI
|
|
8
|
van de Wijgert JHHM: The vaginal
microbiome and sexually transmitted infections are interlinked:
Consequences for treatment and prevention. PLoS Med.
14:e10024782017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Witkin SS and Linhares IM: Why do
lactobacilli dominate the human vaginal microbiota? BJOG.
124:606–611. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kenyon C, Colebunders R and Crucitti T:
The global epidemiology of bacterial vaginosis: A systematic
review. Am J Obstet Gynecol. 209:505–523. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bradshaw CS and Sobel JD: Current
treatment of bacterial vaginosis-limitations and need for
innovation. J Infect Dis. 214 (Suppl 1):S14–S20. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hay PE, Lamont RF, Taylor-Robinson D,
Morgan DJ, Ison C and Pearson J: Abnormal bacterial colonisation of
the genital tract and subsequent preterm delivery and late
miscarriage. BMJ. 308:295–298. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu X, Wu X, Qiu L, Wang D, Gan M, Chen X,
Wei H and Xu F: Analysis of the intestinal microbial community
structure of healthy and long-living elderly residents in Gaotian
Village of Liuyang City. Appl Microbiol Biotechnol. 99:9085–9095.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu J, Lian F, Zhao L, Zhao Y, Chen X,
Zhang X, Guo Y, Zhang C, Zhou Q, Xue Z, et al: Structural
modulation of gut microbiota during alleviation of type 2 diabetes
with a Chinese herbal formula. ISME J. 9:552–562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bolger A, Lohse M and Usadel B:
Trimmomatic: A flexible trimmer for Illumina sequence data.
Bioinformatics. 30:2114–2120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Edgar RC: UPARSE: Highly accurate OTU
sequences from microbial amplicon reads. Nat Methods. 10:996–998.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Magurran AE: Measuring Biological
Diversity. Wiley-Blackwell; Hoboken, NJ: 2004
|
|
18
|
Caporaso JG, Kuczynski J, Stombaugh J,
Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich
JK and Gordon JI: QIIME allows analysis of high-throughput
community sequencing data. Nat Methods. 7:335–336. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Afgan E, Baker D, Batut B, van den Beek M,
Bouvier D, Cech M, Chilton J, Clements D, Coraor N, Grüning BA, et
al: The Galaxy platform for accessible, reproducible and
collaborative biomedical analyses: 2018 update. Nucleic Acids Res.
46:W537–W544. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Johary J, Xue M, Zhu X, Xu D and Velu PP:
Efficacy of estrogen therapy in patients with intrauterine
adhesions: Systematic review. J Minim Invasive Gynecol. 21:44–54.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Deans R and Abbott J: Review of
intrauterine adhesions. J Minim Invasive Gynecol. 17:555–569. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Younes JA, Lievens E, Hummelen R, van der
Westen R, Reid G and Petrova MI: Women and their microbes: The
unexpected friendship. Trends Microbiol. 26:16–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nasioudis D, Linhares IM, Ledger WJ and
Witkin SS: Bacterial vaginosis: A critical analysis of current
knowledge. BJOG. 124:61–69. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moreno I, Codoñer FM, Vilella F, Valbuena
D, Martinez-Blanch JF, Jimenez-Almazán J, Alonso R, Alamá P, Remohí
J, Pellicer A, et al: Evidence that the endometrial microbiota has
an effect on implantation success or failure. Am J Obstet Gynecol.
215:684–703. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Champer M, Wong AM, Champer J, Brito IL,
Messer PW, Hou JY and Wright JD: The role of the vaginal microbiome
in gynecological cancer. BJOG. 125:309–315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jenmalm MC: The mother-offspring dyad:
Microbial transmission, immune interactions and allergy
development. J Intern Med. 282:484–495. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Blaser MJ and Dominguez-Bello MG: The
human microbiome before birth. Cell Host Microbe. 20:558–560. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wolf M, Müller T, Dandekar T and Pollack
JD: Phylogeny of Firmicutes with special reference to Mycoplasma
(Mollicutes) as inferred from phosphoglycerate kinase amino acid
sequence data. Int J Syst Evol Microbiol. 54:871–875. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mor G and Kwon JY: Trophoblast-microbiome
interaction: A new paradigm on immune regulation. Am J Obstet
Gynecol. 213 (4 Suppl):S131–S137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ningthoujam DS, Sanasam S, Tamreihao K and
Salam N: Antagonistic activities of local actinomycete isolates
against rice fungal pathogens. Afr J Microbiol Res. 3:737–742.
2009.
|
|
31
|
Humphries C: Microbiome: Detecting
diversity. Nature. 550:S12–S14. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Martin DH: The microbiota of the vagina
and its influence on women's health and disease. Am J Med Sci.
343:2–9. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Anukam KC, Osazuwa EO, Ahonkhai I and Reid
G: Lactobacillus vaginal microbiota of women attending a
reproductive health care service in Benin city, Nigeria. Sex Transm
Dis. 33:59–62. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tachedjian G, Aldunate M, Bradshaw CS and
Cone RA: The role of lactic acid production by probiotic
Lactobacillus species in vaginal health. Res Microbiol.
168:782–792. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Reid G: Probiotic agents to protect the
urogenital tract against infection. Am J Clin Nutr. 73:437S–443S.
2001. View Article : Google Scholar : PubMed/NCBI
|