Introduction
Immunosuppression mediated by regulatory T cells
(Tregs) is considered to be a key facilitator of tumor immune
evasion (1,2). Marked Treg infiltration into tumors
is generally associated with pathogenesis and a poor clinical
outcome (3). In addition to the
immunosuppressive function of Tregs, emerging evidence suggests a
link between angiogenesis and the accumulation of Tregs at tumor
sites (4,5).
Previous studies have shown that Tregs contribute to
tumor angiogenesis through indirect and direct mechanisms. Tregs
can promote angiogenesis indirectly by inhibiting effector
cell-derived angiostatic cytokines, including interferon γ (IFN-γ)
and C-X-C motif chemokine 10 (CXCL10) (6). Tregs that have been recruited to
hypoxic regions can also directly stimulate angiogenesis through
the production of vascular endothelial growth factor (VEGF)
(4). The association of Treg
infiltration with the overexpression of VEGF and increased
microvessel density in endometrial and breast cancer has provided
clinical clues for a link between Tregs and angiogenesis (6). VEGF promoted the proliferation of
Tregs, and VEGF/VEGF receptor (VEGFR) antibodies and inhibitors
decreased the number of Tregs in patients with colon cancer and in
mouse models (7). Sustained
angiogenesis is also critical for leukemogenesis (8). Our previous study demonstrated that
the expression of Helios in Tregs was involved in angiogenesis
in vitro (9).
Helios, a member of the Ikaros family, serves an
important role in the regulation of lymphoid cell proliferation and
differentiation (10). The
findings of previous studies have led to increased interest in
Helios, which may serve a critical role in controlling certain
aspects of Tregs, including their suppressive function,
differentiation and survival (10,11).
Our previous study confirmed that the increased proportion of
Helios+ Tregs in patients with pediatric acute
lymphoblastic leukemia (ALL) serves an important role in the
mechanism of oncogenesis, and may be involved in the regulation of
bone marrow angiogenesis in ALL (9). However, the mechanism requires
further clarification.
The present study aimed to investigate whether the
expression of Helios in Tregs influences leukemic angiogenesis
in vivo, and to examine the mechanism further. To this end,
a number of Helioshigh Tregs derived from umbilical cord
blood (UCB) were infused into nude mice with defective T cell
function, and leukemia was established in these mice. The findings
suggest the importance of Tregs expressing Helios for angiogenesis
in ALL in vivo.
Materials and methods
Cell culture
The Nalm-6 human pre-B ALL cell line was provided by
Professor Daoxin Ma (Qilu Hospital, Jinan, China). A total of five
normal pediatric peripheral blood (PB) samples were obtained from
consenting individuals (5 female, ages 5–13) with no history of
tumors, antecedent primary hematologic abnormalities or
immunodeficiency diseases who underwent orthopedic surgery at the
Qilu Hospital (Jinan, China) from May-July 2016. A total of five
UCB samples were donated by healthy mothers (ages 23–29) at the
Qilu Hospital from May-July 2016. Human umbilical vein endothelial
cells (HUVECs) were derived from cryopreserved cells in stem cell
and regenerative medicine research center (Shandong University,
Jinan, China). The Nalm-6 cells were cultured in complete RPMI-1640
medium (HyClone, GE Healthcare Life Sciences, Logan, UT, USA)
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The HUVECs were cultured in
M199 medium (Gibco; Thermo Fisher Scientific, Inc.) containing 15%
FBS and 40 ng/ml fibroblast growth factor (PeproTech, Inc., Rocky
Hill, NJ, USA). Monocytes from the UCB or PB were isolated from the
samples and cultured in complete RPMI-1640 medium. On day 1, 200
U/ml human recombinant interleukin-2 (rIL-2) (PeproTech, Inc.), 2.5
ng/ml transforming growth factor (TGF)-β (eBioscience; Thermo
Fisher Scientific, Inc.), 0.5 µg/ml anti-CD3e (OKT3; eBioscience;
Thermo Fisher Scientific, Inc.) and 1 µg/ml anti-CD28 (CD28.2;
eBioscience; Thermo Fisher Scientific, Inc.) were added to the
medium. In accordance with routine culturing, the cells were
passaged every 3 days with the addition of 100 U/ml rIL-2. During
the culture period, CD4+CD25+ Tregs and
CD4+CD25− T helper (Th) cells were isolated
using a CD4+CD25+ Treg Isolation kit
(Miltenyi Biotec GmbH, Bergisch Gladbach, Germany). The purity of
the Th and Treg cell populations was >90%, as determined by flow
cytometry using a Guava easyCyte 8HT flow cytometer (EMD Millipore,
Billerica, MA, USA). The cells were counted using a hemocytometer
on days 0, 7, 14, 21 and 28 of the culture period. All cells were
cultured at 37°C under a 5% CO2 atmosphere. All study
participants, including direct participants, blood donors and
parent/guardians of child participants, provided written informed
consent. The study was approved by the Research Ethics Committee of
Qilu Hospital.
Expression of Helios in UCB Tregs
To confirm the role of Helios in Tregs, the
expression of Helios was reduced in Treg cells on day 14 of
culture. A lentiviral vector was used to silence the expression of
Helios in the UCB-derived Tregs on day 14 of the culture period.
The lentiviral vectors, GV248-enhanced green fluorescent protein
(EGFP) short hairpin (sh) RNA-Helios and GV248-EGFP, were developed
by Shanghai GeneChem Co., Ltd. (Shanghai, China; Fig. S1A). The Helios sequence was
obtained from GeneBank (accession no. NM_016260). According to the
RNAi sequence design principle, multiple RNAi target sequences were
designed, and the optimal kinetic parameter target was selected to
enter the subsequent experimental procedure. The Helios-targeting
shRNA sequences (RNAi-sense
5′-CCGGGGAAGATTGTAAGGAACAACTCGAGTTGTTCCTTACAATCTTCCTTTTTG-3′;
RNAi-antisense,
5′-AATTCAAAAAGGAAGATTGTAAGGAACAACTCGAGTTGTTCCTTACAATCTTCC-3′) were
designed. These were inserted between the AgeI and EcoRI sites in
the GV248 vector (Shanghai GeneChem Co., Ltd.) by homologous
recombination using the T4 DNA ligase enzyme.
Cells in the logarithmic growth phase were digested
with trypsin 24 h prior to being transfected. The cells were
re-suspended at 5.0×106 cells/l5 ml in Dulbecco's
modified Eagle's medium containing 10% FBS, and maintained at 37°C
in an atmosphere of 5% CO2 for 24 h until the cells were
70–80% confluent. The cell culture medium was replaced with
serum-free medium 2 h prior to transfection. DNA was extracted
using the Plasmid Maxi kit (Qiagen China Co., Ltd., Shanghai,
China), according to the manufacturer's instructions. A solution
containing the prepared DNA (20 µg pGV-shRNA vector, 15 µg pHelper
vector 1.0 and 10 µg pHelper vector 2.0) was added to a sterile
centrifuge tube, and the final volume was brought to 1 ml. The
solution was incubated at room temperature for 15 min, and 293T
cells were transfected using GeneChem transfection reagent
(Shanghai GeneChem Co., Ltd.) according to the manufacturer's
instructions. The transfected cells were incubated in serum-free
medium for 6 h, the medium was subsequently replaced with medium
containing 10% serum, and the cells were cultured for an additional
48–72 h. The packaging 293T cell line was transfected with the
lentiviral vector.
The culture supernatants were collected when the
cells expressed high levels of green fluorescence and cell fusion
was observed (48–72 h). The virus in the supernatant was
concentrated to the target volume by centrifugation (4°C, 4,000 ×
g, 10 min). The viral concentrate was removed and aliquot into a
tube for long-term storage at −80°C. In order to determine the
viral titer, 10 µl viral stock was used to make 1 ml serial
dilutions (10−2−10−6), which were used to
infect 293T cells in a 96-well plate (4×104 cells).
Following 4 days in culture, EGFP expression in each well was
observed under a fluorescence microscope (magnification, ×200). The
number of cells expressing EGFP was counted and multiplied by the
dilution ratio to determine the viral titer [transducing units
(TU)/ml]. The viral supernatant was harvested 48 h after
transfection, and the lentiviral particle titer was determined
(5×108 TU/ml).
The Tregs were cultured in 25 cm2 culture
flasks at a density of 1×106 cells/ml. Following
overnight culture, the cells were infected for 12 h at 37°C with
the lentiviral vector. Subsequently, the cells were washed and
cultured in fresh medium for 3 days. The viral concentrates of
shRNA-Helios were diluted to infect the UCB Tregs at a multiplicity
of infection of 50. The transduction rate and expression of Helios
in the Tregs were confirmed by flow cytometry (Fig. S1B and C).
Establishment of murine models of
ALL
A total of 48 4-week-old female BALB/c nude mice
(14±1 g) were obtained from Beijing HFK Bioscience (Beijing,
China). The animals were housed in a specific pathogen-free
environment, with a constant temperature between 25°C and 27°C, in
a 12/12 h light/dark cycle. The mice were fed with a standard diet
and water ad libitum. On days 1 and 2, all mice received
cyclophosphamide (2 mg/mouse; Heng Rui Medicine Co., Ltd., Jiangsu,
China) by intraperitoneal injection. Animals in the blank group
received PBS only on day 3. The model group was injected with
Nalm-6 cells (3×106/mouse) through the tail vein on day
3. In the Helioslow group, Nalm-6 cells
(2.5×106/mouse) were injected first through the tail
vein on day 3, followed by UCB-derived Tregs
(0.6×106/mouse) with shRNA-Helios transduction. In the
Helioshigh group, Nalm-6 cells
(2.5×106/mouse) were injected first through the tail
vein on day 3, followed by UCB-derived Tregs on day 14 of the
culture period (0.6×106/mouse). Each group consisted of
12 mice. Mouse weights were determined once a week. In the
Helioshigh group, one mouse died on day 32 and another
on day 41. In the model group, one mouse died on day 41. All mice
were sacrificed on day 42 when the hind legs had become paralyzed
in the majority of the leukemia model animals. The liver, spleen
and kidneys were weighed to calculate the organ index, calculated
as the organ weight to total body weight ratio. Bone marrow smears
from the femur were examined in the Department of Pathology of Qilu
Hospital. The use of the animals was approved by the Animal Care
and Use Committee of Shandong University (Jinan, China).
Flow cytometry
The cells were stained with the following
fluorochrome-conjugated antibodies: Phycoerythrin (PE) anti-human
(h)CD19 (cat. no. SJ25C1, 12-0198, 0.06 µg/test; eBioscience;
Thermo Fisher Scientific, Inc.), fluorescein isothiocyanate
anti-hCD4 (cat. no OKT4, 11-0048, 0.25 µg/test; eBioscience; Thermo
Fisher Scientific, Inc.), PE anti-hCD25 (cat. no BC96, 302606, 5
µl/test; BioLegend, Inc., San Diego, CA, USA), PerCP/Cy5.5
anti-mouse (m)CD4 (cat. no GK1.5, 100434, 0.25 µg/106
cells; BioLegend, Inc.), and allophycocyanin anti-mCD25 (cat. no
3C7, 101910, 0.25 µg/106 cells; BioLegend, Inc.).
Allophycocyanin anti-human forkhead box P3 (hFoxP3; cat. no PCH101,
17-4776, 0.5 µg/test; eBioscience; Thermo Fisher Scientific, Inc.),
PE anti-hHelios (cat. no 22F6, 12-9883, 0.03 µg/test; eBioscience;
Thermo Fisher Scientific, Inc.) and Alexa Fluor® 488
anti-mFOXP3 (cat. no 150D, 320012, 5 µl/test; BioLegend, Inc.) were
detected using the FoxP3/Transcription Factor Staining Buffer set
(cat. no 00-5523; eBioscience; Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. Analyses were performed
using a Guava easyCyte 8HT flow cytometer (EMD Millipore), and the
data were analyzed using Guava software 6.1. Multi-color flow
cytometry with fluorescence minus one control was used to determine
background signal levels.
Carboxyfluorescein diacetate
succinimidyl ester (CFSE)-based proliferation and suppression
assay
The proliferation capacity of the Treg population
was detected as follows: Treg cells (1×105 cells/well)
were labeled with 5 µM CFSE and incubated for 48 h at 37°C in a
12-well plate. The proliferation of Treg cells was evaluated by
flow cytometry.
The suppressive capacity of the Treg population was
detected as follows: CD4+CD25− Th cells were
labeled with 5 µM CFSE and incubated for 48 h at 37°C. The Tregs
(1×105 cells/well) were cultured in RPMI-1640 medium
supplemented with 100 U/ml rIL-2 and CD3/CD28 beads (bead: Cell
ratio of 1:1; Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C.
These Tregs were then co-cultured with Th cells for 72 h at 37°C.
The proliferation of Th cells was evaluated by flow cytometry.
Modfit LT software (Version 3.1, Verity Software House, Inc.,
Topsham, ME, USA) was used for data analyses.
Western blotting
Western blotting was performed as described
previously (9). Antibodies against
VEGFA (cat. no. ab46154; 1:1,000; Abcam, Cambridge, MA, USA),
VEGFR2 (cat. no. ab11939; 1:500; Abcam), VEGFR2 (phosphoS473; cat.
no. ab5473; 1:800; Abcam), CD31 (cat. no. ab28364; 1:1,000; Abcam)
and β-actin (cat. no. 60008-1-Ig; 1:2,000; ProteinTech Group, Inc.)
were used. The proteins were detected using the ECL
Chemiluminescence Detection kit (EMD Millipore) and quantitated
with Image Studio Digits Ver 4.0 (LI-COR Biosciences, Lincoln, NE,
USA). The signal intensities of all target proteins were normalized
against β-actin signals.
In vitro vascular tube formation
assay
To verify the effect of Helios+ Tregs on
endothelial tube formation, the culture supernatant of transfected
Tregs was obtained by centrifugation at 800 × g for 10 minutes at
room temperature. There were four groups: Blank group (blank),
normal Tregs group on day 0 of the culture period (control), Tregs
with high expression of Helios on day 14 of the culture period
(Helioshigh Treg), and Tregs with low expression of
Helios (Helioslow Treg). Growth factor-reduced Matrigel
(BD Biosciences, San Jose, CA, USA) was allowed to polymerize in a
96-well plate at 37°C for at least 30 min. The HUVECs
(5×103 cells /well) were suspended in 100 µl medium
conditioned with culture supernatant from the normal Tregs
(Control), Helioslow Tregs or Helioshigh
Tregs. Following incubation for 6 h at 37°C, images of
capillary-like structures in the Matrigel were captured under an
Olympus IX71 inverted fluorescence microscope (Olympus Corporation,
Tokyo, Japan). Tube formation was analyzed by determining the total
tube length in each well using ImageJ software 1.43u (National
Institutes of Health, Bethesda, MA, USA). The tube length of the
entire culture well was measured. Fold changes of capillary length
with respect to the control were determined. The total tube length
of the blank group was set to one.
Immunohistochemical assessment of
microvessel density (MVD)
Paraffin-embedded and sliced bone marrow, liver and
spleen tissues (4-µm thick) were produced by Servicebio Company
(Wuhan, Hubei, China). The sections were prepared for microscopic
examination by routine immunohistochemical methods using an
antibody against CD31 (cat. no. ab28364, 1:50, Abcam). Low
magnification (×100) was used to identify the regions of highest
vascular density (hot spots), and three hot spots were selected to
count the number of microvessels under high magnification (×400). A
microvessel was considered as a brown-stained endothelial cell or
endothelial cell cluster, with or without lumen, that was clearly
separated from the adjacent tissue. The average value of these
three counts represented the MVD for that case.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from samples using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). First-strand cDNA was synthesized using 1 µg of total RNA in
a 20 µl of reverse transcription reaction mixture using the
ReverTra Ace qPCR RT Master Mix kit (Toboyo Co., Ltd., Osaka,
Japan) with primers. The temperature protocol was as follows: 37°C
15 min, 95°C 5 min and 4°C hold. qPCR analysis was performed on an
ABI Prism 7500 sequence detection system using Thunderbird SYBR
qPCR mix (Toboyo Co., Ltd.) in 20 µl of reaction mixture. The
thermocycling conditions were as follows: Initial denaturation at
95°C for 5 min, followed by 33 cycles of 95°C (15 sec) and 60°C (1
min). Data were analyzed using Sequence Detection Software 1.4
(Applied Biosystems; Thermo Fisher Scientific, Inc.). GAPDH was
used as an internal control. Values for the blank group were set to
one. RNA expression levels were quantified using the
2−ΔΔCq method and normalized to the expression levels of
GAPDH (12). The primer sequences
are presented in Table SI.
Measurement of chemokine CC-chemokine
ligand 22 (CCL22) by ELISA
Following the onset of ALL in mice, the mice were
sacrificed and plasma was obtained by centrifugation at 1,000 × g
for 10 minutes at room temperature for CCL22 protein detection. The
protein levels of CCL22 were measured using an ELISA kit (cat. no.
ab204525; Abcam) according to the manufacturer's protocol.
Statistical analysis
All data are presented as the mean ± standard
deviation of three separate experiments. Differences in the number
of PB-Tregs and UCB-Tregs, and the proportion of Helios+
cells following shRNA-Helios transduction were assessed using
Student's t-test. Other experiments were assessed by one-way
analysis of variance (ANOVA), which was used to compare multiple
groups. If the ANOVA result was significant, the Bonferroni test
was performed. Statistical analyses were performed using SPSS
Statistics 17.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Isolation, amplification and
identification of
CD4+CD25+FoxP3+Helios+
Tregs from fresh UCB
UCB is considered to be a universal source of Tregs,
the function of which is not influenced by HLA antigen expression.
The present study compared the proliferation and suppression
abilities of
CD4+CD25+FoxP3+Helios+
Tregs derived from UCB and pediatric PB during the culture period.
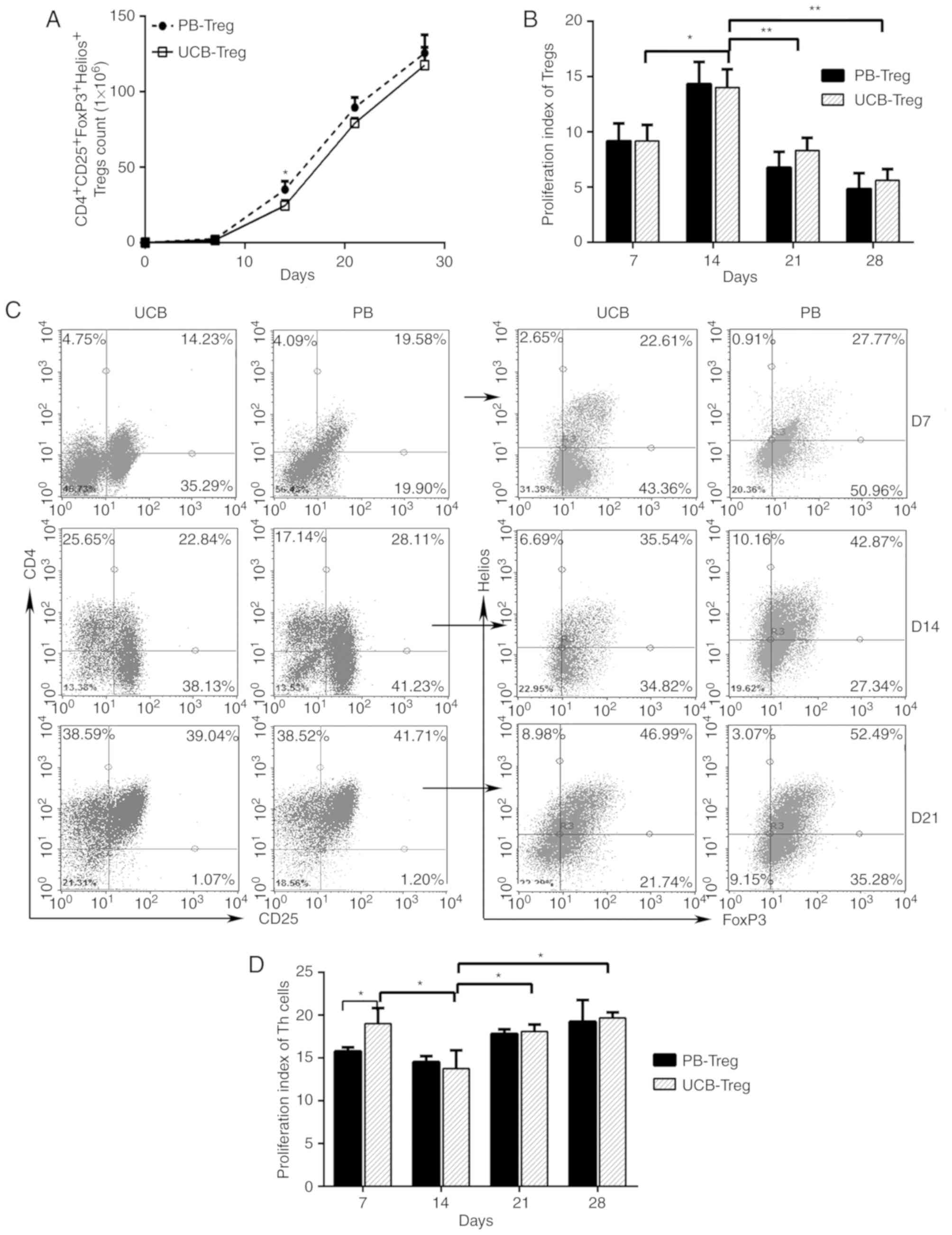
As shown in Fig. 1A, the number of
induced CD4+CD25+ Treg cells derived from the
UCB and PB increased steadily between days 0 and 28. A
statistically significant difference in the number of PB Treg cells
and UCB Treg cells was observed on day 14. However, no significant
differences in the numbers of these cells were observed on days 7,
21 and 28 of cultivation. On day 14, the proliferation capability
of the Tregs was significantly enhanced in the UCB and PB (Fig. 1B). The ratio of Helios+
FoxP3+ T cells to isolated
CD4+CD25+ T cells increased gradually between
days 7 and 21, and reached a peak of almost 50% on day 21 (Fig. 1C). After day 21, cell debris
significantly increased in the culture medium. The suppression
ability of UCB Tregs was significantly enhanced on day 14 (Fig. 1D). On days 14, 21 and 28, the
immunosuppressive ability of the UCB Treg cells did not differ
significantly from that of the PB Treg cells (Fig. 1D). Based on the high expression of
Helios and high immunosuppressive ability, Treg cells on day 14 of
the culture period were selected for the subsequent
experiments.
UCB-derived Helios+ Tregs
promote angiogenesis in vitro
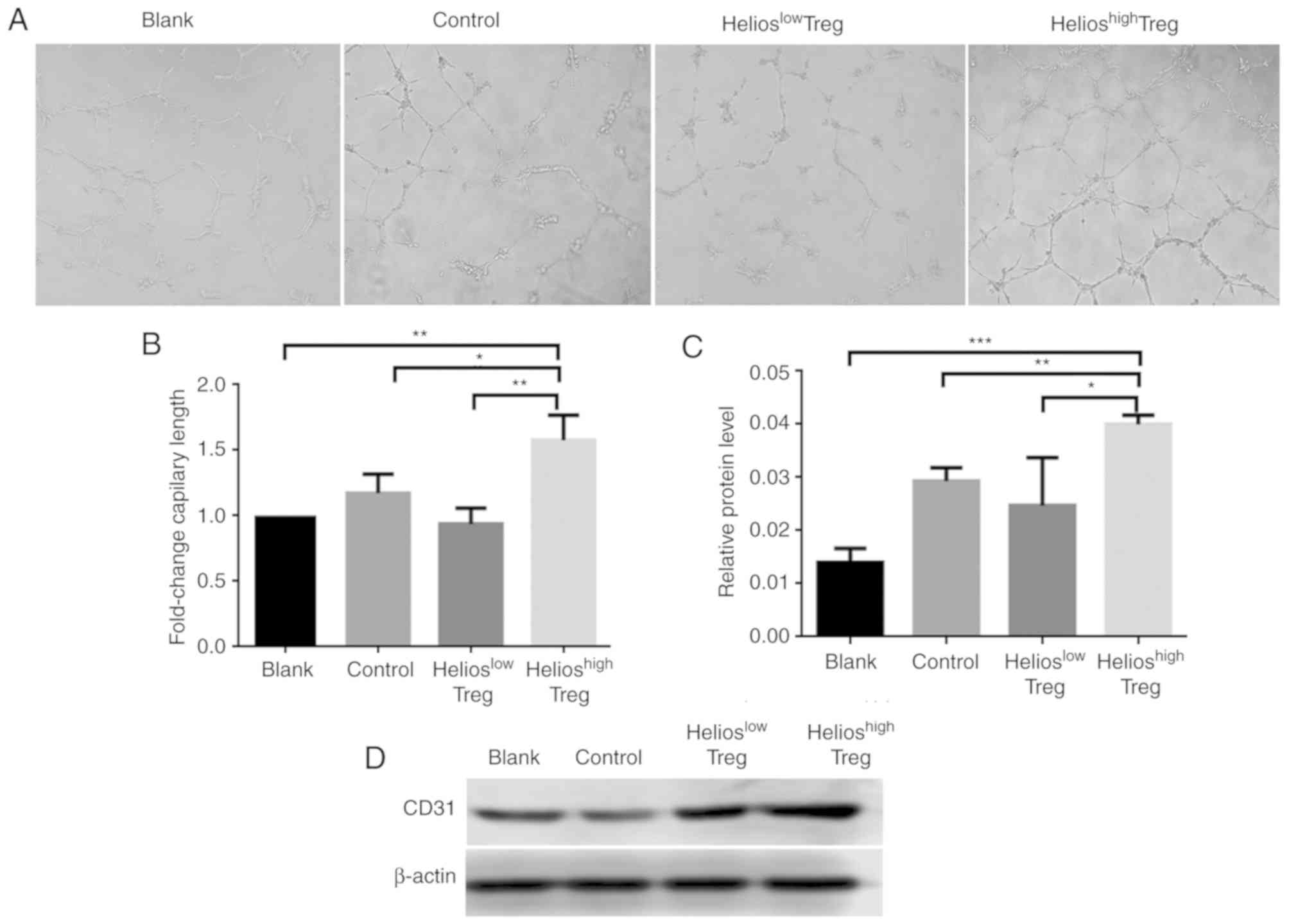
The effect of Helios+ Tregs on
angiogenesis in vitro was subsequently examined. The results
showed that, compared with the normal Tregs, the supernatant from
Helioshigh Tregs promoted angiogenesis (Fig. 2A and B). By contrast, inhibiting
the expression of Helios in UCB Treg cells via shRNA-Helios reduced
the angiogenic ability (Fig. 2A and
B).
For further verification, protein levels of CD31 in
HUVECs in the co-culture system were evaluated. Compared with the
control and blank groups, the HUVECs incubated in supernatants from
Helioshigh Tregs exhibited higher protein levels of
CD31. By contrast, when shRNA-Helios was present, the expression of
CD31 was decreased (Fig. 2C and
D).
UCB-derived Helios+ Tregs
increase the infiltration of leukemia cells into the bone
marrow
The weights of mice with ALL were lower after 5
weeks compared with those in the blank group; however, there were
no significant differences in weights among the three ALL groups
(Fig. S2A). The ALL mice
exhibited significant increases in the liver, spleen and kidney
indices, compared with those in the normal nude mice. However,
among the three ALL groups, no significant differences were
observed in the liver, spleen or kidney indices (Fig. S2B). At 1 week post-Tregs infusion,
neither human nor murine Tregs were detected in the PB of any
groups.
Subsequently, the present study performed a BM smear
to analyze the incidence of leukemia. Of the recipients, 90% (9/10)
developed extensive bone marrow lymphoblastic infiltration in the
Helioshigh group. This was compared with 83% (10/12) in
the Helioslow group, 64% (7/11) in the model group and
0% in the blank group (Fig. 3A).
Furthermore, human CD19+ cells in the BM were detected
by flow cytometry. Compared with the model group and
Helioslow group, the proportion of human
CD19+ cells in the BM was increased in the
Helioshigh group of ALL mice (Fig. 3B and C). Human CD19+
cells were also detected in the PB. The accumulation of Tregs
resulted in a higher number of transplanted primary leukemia cells,
however, no differences were found between the
Helioshigh and Helioslow groups (Fig. 3D).
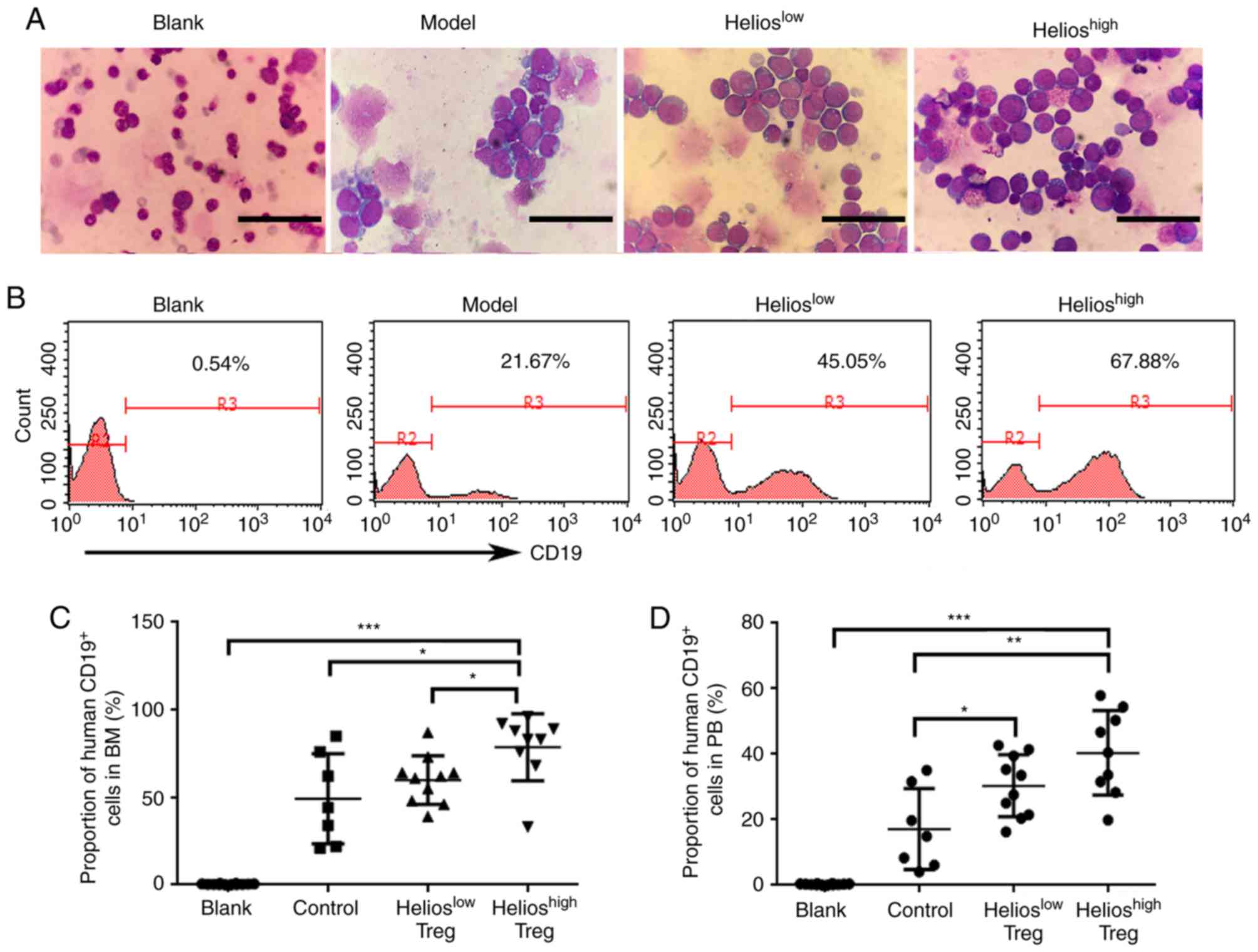 | Figure 3.Helios+ Tregs promote the
infiltration of lymphoblasts in mouse models of acute lymphoblastic
leukemia. (A) Proportions of lymphoblasts in the BM of the blank,
model, Helioslow and Helioshigh groups. Scale
bar=25 µm. (B) Proportions and (C) quantitative analyses of human
CD19+ cells in the BM of the blank, model,
Helioslow and Helioshigh groups, determined
by flow cytometry. (D) Quantitative analyses of human
CD19+ cells in the PB of the model, Helioslow
and Helioshigh groups. (***P<0.0005, **P<0.005,
*P<0.05). BM, bone marrow; PB, peripheral blood; Treg,
regulatory T cell. |
UCB-derived Helios+ Tregs
promote angiogenesis in ALL mice
The correlations between the accumulation of
Helios+ Tregs and biomarkers of angiogenesis, including
VEGF family expression and MVD, were examined in four groups. In
the bone marrow, the levels of VEGFA, VEGFR2 and phosphorylated
(p)-VEGFR2 were increased in the mice infused with UCB-derived
Tregs, and this was more marked in the Helioshigh group
(Fig. 4A). In the liver, spleen
and kidneys of the ALL model mice, the protein expression levels of
VEGFA, VEGFR1, VEGFR2 and p-VEGFR2 were significantly increased
compared with those in the normal mice; however, no statistically
significant differences in protein expression were observed among
the three ALL groups (P>0.05).
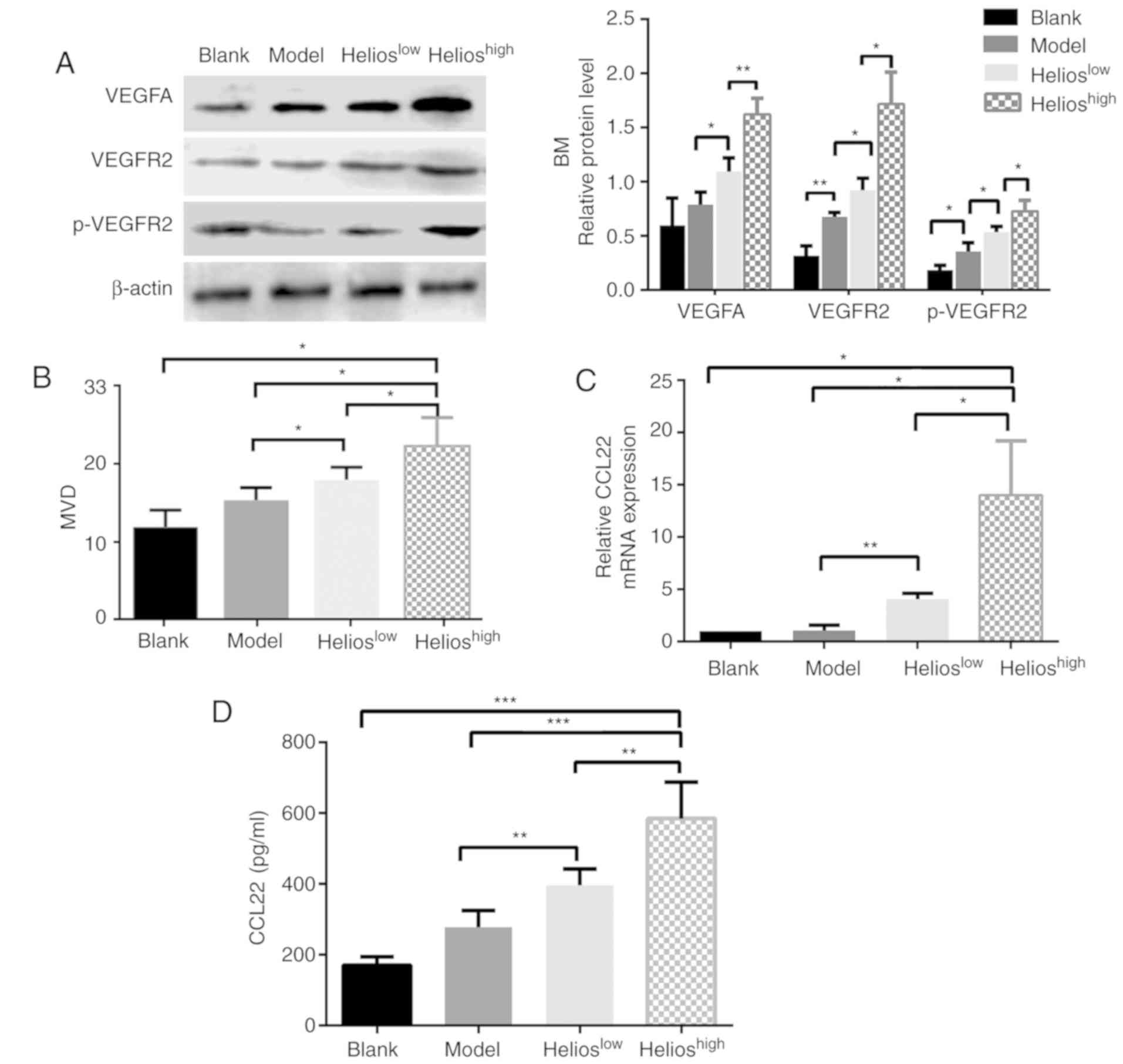 | Figure 4.Helios+ Tregs promote
angiogenesis through the VEGFA/VEGFR2 pathway in a mouse model of
acute lymphoblastic leukemia. (A) Western blot and quantitative
analyses of VEGFA, VEGFR2 and p-VEGFR2 proteins in the BM of the
blank, model, Helioshigh and Helioslow
groups. (B) Quantitative analyses of the CD31 MVD values of the BM
in the blank, model, Helioslow and Helioshigh
groups. (C) Quantitation of the mRNA expression of CCL22 in the BM
of the blank, model, Helioslow and Helioshigh
groups. (D) Quantitation of the protein levels of CCL22 in the
plasma of the blank, model, Helioslow and
Helioshigh groups. (***P<0.0005, **P<0.005,
*P<0.05). VEGF, vascular endothelial growth factor; VEGFR2, VEGF
receptor 2; p-VEGFR2, phosphorylated VEGFR2; MVD, microvascular
density; BM, bone marrow; CCL2, CC-chemokine ligand 22. |
The mean MVD was determined in the bone marrow,
liver and spleen. The mice in the Helioshigh group had
higher MVD in the bone marrow (Fig.
4B), although no differences were present between the
Helioshigh and Helioslow groups with respect
to the liver and spleen (P>0.05). The downstream genes regulated
by Helios+ Tregs were then identified. A higher
expression of Helios in Tregs significantly increased the mRNA
expression of CCL22 in the bone marrow (Fig. 4C), whereas the expression of other
factors, including C-X-C motif chemokine ligand 6 (CXCL6),
interleukin (IL)-17a, interferon-γ, TGF-β1, IL-10, CCL28 and
neuropilin 1, remained unchanged (data not shown). Furthermore, the
secretion of CCL22 protein in the plasma of mice was examined. The
results showed that a higher expression of Helios in Tregs
significantly increased the protein expression of CCL22 in the
plasma (Fig. 4D). When
shRNA-Helios inhibited the expression of Helios in Tregs, the
expression of CCL22 in the plasma decreased, compared with that in
the Helioshigh group (Fig.
4D).
Discussion
In the present study, an immunosuppressive internal
environment was established by infusing a number of
Helios+CD4+CD25+FoxP3+
Tregs into nude mice. The results confirmed that the infusion of
Helios+ Tregs increased the extent of leukemic cell
infiltration in the bone marrow, with Helios serving a key role in
angiogenesis. Consequently, Helios+ Tregs are closely
associated with the onset of leukemia, which is consistent with our
previous study in pediatric patients with ALL (9).
Previous studies have reported a positive
correlation between angiogenesis and tumor-infiltrating Tregs
(13,14). Our previous study found an increase
in the expression of Helios in ALL Tregs, and the capacity of
PB-derived Helios+ Tregs to ostensibly control the
process of neovascularization in vitro (9). The present study confirmed that the
overexpression of Helios in Tregs activated microvascular formation
in the bone marrow of ALL mice. Due to the short onset time of ALL
in mice, Treg cells may have mainly promoted leukemia cell
infiltration of the bone marrow, which is the site of leukemia, and
had minimal effect on liver and spleen infiltration. Therefore, the
pro-angiogenic effect of Treg cells was mainly reflected in the
bone marrow. Tregs can contribute to tumor angiogenesis through
indirect and direct mechanisms. The mass of Tregs in the tumor
microenvironment effectively restricts the Th 1 effect, which
decreases the secretion of anti-angiogenic factors and indirectly
promotes tumor angiogenesis (15).
By contrast, Tregs can directly synthesize and secrete certain
pro-angiogenic factors, including VEGF, neuropilin-l and apelin
(16–18).
VEGF promotes tumor angiogenesis through stimulating
the proliferation and survival of endothelial cells, and also by
increasing the permeability of vessels and recruiting vascular
precursor cells from the bone marrow (19). In the present study, the effects of
Helios+ Tregs on the microvasculature during ALL were
mediated by the VEGFA/VEGFR2 pathway. VEGFA has been the subject of
more investigations than other VEGF family members, and is a
critical regulator of angiogenesis. VEGFR2 is the main signaling
VEGFR in blood vascular endothelial cells (19,20).
The blockade of VEGFA with a specific antibody decreases the number
of Tregs, and inhibiting VEGFA/VEGFR-transduced signals counteracts
the induction of Tregs by malignanT cells (21). Sunitinib, an agent targeting
VEGFRs, has been reported to reduce the number of Tregs in
tumor-bearing mice and in patients with metastatic renal carcinoma
(22). Notably, the depletion of
CD25+ or CCR10+ cells has been shown to
eliminate Treg cells from the tumor microenvironment, and
significantly suppress the expression of VEGF and angiogenesis at
tumor sites (4). The present study
demonstrated that the high expression of Helios in Tregs is an
important factor in regulating bone marrow angiogenesis in ALL mice
via the VEGF pathway.
Helios is expressed at relatively high levels in
functional Tregs. Studies have shown that the overexpression of
Helios enhances the immunosuppressive function of normal Tregs on
Th cells (23). By contrast,
Helios-deficient Tregs within tumors acquire effector T cell
function and contribute to immune responses against cancer
(11,24). CD4+ invariant natural
killer T cells protect from graft-versus-host disease-associated
morbidity and mortality through an expansion of donor
Helios+ Tregs (25).
Helios not only influences the expression of FoxP3, but also acts
as a positive regulator of the TGF-β suppressor-effector function
(9). A previous study found that
tumor-infiltrating Tregs were mainly Helios+ activated
Tregs and significantly correlated with the concentration of CCL22
in ovarian tumor cell culture supernatants (26). The results of the present study
also showed that a high expression of Helios in Tregs induced
abnormal expression of the pro-angiogenic factor CCL22, which is
responsible for Treg recruitment at tumor sites, and thus promoted
angiogenesis further.
Chemokines have been shown to have pleiotropic
effects in promoting tumor invasion, migration and vascularization
(27). The upregulation of CCL28
caused by tumor hypoxia in ovarian cancer resulted in marked Treg
accumulation, increased levels of VEGF and significantly increased
blood vessel development (4).
Tumor environments contain high levels of CCL22, which is likely
derived from tumor cells and tumor macrophages (28). CCL22 can recruit Tregs through
CCR4, and Treg migration can be abrogated through the inhibition of
CCR4 in vitro (29). Human
T-lymphotropic virus type 1 induces and maintains a high frequency
of FoxP3+ T cells by inducing expression of chemokine
CCL22; the frequency is particularly high in patients with chronic
leukemia (30). In the present
study, a high expression of Helios in Treg cells stimulated
macrophages, dendritic cells or leukemia cells to secrete CCL22,
thereby recruiting Treg cells to the tumor site and then activating
the VEGF signaling pathway to promote angiogenesis in tumor
sites.
As a traditional and convenient source of cells for
hematopoietic stem cell transplantation, UCB T cells contain a
higher percentage of the naïve CD4+CD25+
subset compared with adult T cells (31). Therefore, the expansion of UCB
Tregs can overcome the cell number limitation that often prevents
their use (31,32). In the present study, UCB
Helios+ Tregs exhibited a similar proliferative capacity
to pediatric PB Tregs. Although allogeneic Tregs from donors may
offer improved therapeutic benefits, UCB Tregs have low
immunogenicity and a high safety margin (33,34).
Therefore, UCB may represent a safe and potent source for in
vitro Treg expansion.
In conclusion, the results of the present study
further clarify the mechanism of Tregs in the pathogenesis of ALL
and highlight the impact of the Treg expression of Helios in on
angiogenesis in ALL. These findings may assist in understanding the
mechanisms by which Treg cells are involved in the pathogenesis of
ALL, and may be useful for developing a molecular therapeutic
strategy for ALL by targeting Tregs expressing Helios.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by Shandong Province Major
Scientific Research Projects (grant nos. 2017GSF18155 and
2017GSF218015), the Shandong Province Natural Science Foundation
(grant no. ZR2018MH012), the Ji'nan Science and Technology
Development Foundation (grant no. 201704066) and the Shandong
Province Natural Science Fund (grant no. 2014ZRE27630).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XJ and DL designed the study and drafted the
manuscript; XL performed the cell culture and animal experiments,
and was a major contributor in writing the manuscript. QS performed
the in vitro vascular tube formation assay. XH performed
CFSE-based proliferation and suppression assay. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All study participants, including direct
participants, blood donors and parent/guardians of child
participants provided written informed consent. The study was
approved by the Research Ethics Committee of Qilu Hospital and the
use of the animals was approved by the Animal Care and Use
Committee of Shandong University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nishikawa H and Sakaguchi S: Regulatory T
cells in tumor immunity. Int J Cancer. 127:759–767. 2010.PubMed/NCBI
|
|
2
|
Takeuchi Y and Nishikawa H: Roles of
regulatory T cells in cancer immunity. Int Immunol. 28:401–409.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Duell J, Dittrich M, Bedke T, Mueller T,
Eisele F, Rosenwald A, Rasche L, Hartmann E, Dandekar T, Einsele H
and Topp MS: Frequency of regulatory T cells determines the outcome
of the T-cell-engaging antibody blinatumomab in patients with
B-precursor ALL. Leukemia. 31:2181–2190. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Facciabene A, Peng X, Hagemann IS, Balint
K, Barchetti A, Wang LP, Gimotty PA, Gilks CB, Lal P, Zhang L and
Coukos G: Tumour hypoxia promotes tolerance and angiogenesis via
CCL28 and T(reg) cells. Nature. 475:226–230. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weis SM and Cheresh DA: Tumor
angiogenesis: Molecular pathways and therapeutic targets. Nat Med.
17:1359–1370. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Facciabene A, Motz GT and Coukos G:
T-regulatory cells: key players in tumor immune escape and
angiogenesis. Cancer Res. 72:2162–2171. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang J, Yan J and Liu B: Targeting
VEGF/VEGFR to modulate antitumor immunity. Front Immunol.
9:9782018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aguayo A, Kantarjian H, Manshouri T, Gidel
C, Estey E, Thomas D, Koller C, Estrov Z, O'Brien S, Keating M, et
al: Angiogenesis in acute and chronic leukemias and myelodysplastic
syndromes. Blood. 96:2240–2245. 2000.PubMed/NCBI
|
|
9
|
Li X, Li D, Huang X, Zhou P, Shi Q, Zhang
B and Ju X: Helios expression in regulatory T cells promotes
immunosuppression, angiogenesis and the growth of leukemia cells in
pediatric acute lymphoblastic leukemia. Leukemia Res. 67:60–66.
2018. View Article : Google Scholar
|
|
10
|
Getnet D, Grosso JF, Goldberg MV, Harris
TJ, Yen HR, Bruno TC, Durham NM, Hipkiss EL, Pyle KJ, Wada S, et
al: A role for the transcription factor Helios in human
CD4(+)CD25(+) regulatory T cells. Mol Immunol. 47:1595–1600. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim HJ, Barnitz RA, Kreslavsky T, Brown
FD, Moffett H, Lemieux ME, Kaygusuz Y, Meissner T, Holderried TA,
Chan S, et al: Stable inhibitory activity of regulatory T cells
requires the transcription factor Helios. Science. 350:334–339.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
D'Alessio FR, Zhong Q, Jenkins J,
Moldobaeva A and Wagner EM: Lung angiogenesis requires CD4(+)
forkhead homeobox protein-3(+) regulatory T cells. Am J Respir Cell
Mol Biol. 52:603–610. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Woidacki K, Meyer N, Schumacher A,
Goldschmidt A, Maurer M and Zenclussen AC: Transfer of regulatory T
cells into abortion-prone mice promotes the expansion of uterine
masT cells and normalizes early pregnancy angiogenesis. Sci Rep.
5:139382015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Miyara M, Ito Y and Sakaguchi S: TREG-cell
therapies for autoimmune rheumatic diseases. Nat Rev Rheumatol.
10:543–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Terme M, Pernot S, Marcheteau E, Sandoval
F, Benhamouda N, Colussi O, Dubreuil O, Carpentier AF, Tartour E
and Taieb J: VEGFA-VEGFR pathway blockade inhibits tumor-induced
regulatory T-cell proliferation in colorectal cancer. Cancer Res.
73:539–549. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yadav M, Louvet C, Davini D, Gardner JM,
Martinez-Llordella M, Bailey-Bucktrout S, Anthony BA, Sverdrup FM,
Head R, Kuster DJ, et al: Neuropilin-1 distinguishes natural and
inducible regulatory T cells among regulatory T cell subsets in
vivo. J Exp Med. 209:1713–1722, S1711-1719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leung OM, Li J, Li X, Chan VW, Yang KY, Ku
M, Ji L, Sun H, Waldmann H, Tian XY, et al: Regulatory T cells
promote apelin-Mediated sprouting angiogenesis in Type 2 diabetes.
Cell Rep. 24:1610–1626. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sia D, Alsinet C, Newell P and Villanueva
A: VEGF signaling in cancer treatment. Curr Pharm Des.
20:2834–2842. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shibuya M: VEGF-VEGFR signals in health
and disease. Biomol Ther(Seoul). 22:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Terme M, Tartour E and Taieb J:
VEGFA/VEGFR2-targeted therapies prevent the VEGFA-induced
proliferation of regulatory T cells in cancer. Oncoimmunology.
2:e251562013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xin H, Zhang C, Herrmann A, Du Y, Figlin R
and Yu H: Sunitinib inhibition of Stat3 induces renal cell
carcinoma tumor cell apoptosis and reduces immunosuppressive cells.
Cancer Res. 69:2506–2513. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Syed Khaja AS, Toor SM, El Salhat H, Ali
BR and Elkord E: Intratumoral foxP3(+)helios(+) regulatory T cells
upregulating immunosuppressive molecules are expanded in human
colorectal cancer. Front Immunol. 8:6192017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakagawa H, Sido JM, Reyes EE, Kiers V,
Cantor H and Kim HJ: Instability of Helios-deficient Tregs is
associated with conversion to a T-effector phenotype and enhanced
antitumor immunity. Proc Natl Acad Sci U S A. 113:6248–6253. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schneidawind D, Pierini A, Alvarez M, Pan
Y, Baker J, Buechele C, Luong RH, Meyer EH and Negrin RS: CD4+
invariant natural killer T cells protect from murine GVHD lethality
through expansion of donor CD4+CD25+FoxP3+ regulatory T cells.
Blood. 124:3320–3328. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fialova A, Partlova S, Sojka L, Hromádková
H, Brtnický T, Fučíková J, Kocián P, Rob L, Bartůňková J and Spíšek
R: Dynamics of T-cell infiltration during the course of ovarian
cancer: The gradual shift from a Th17 effector cell response to a
predominant infiltration by regulatory T-cells. Int J Cancer.
132:1070–1079. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kumai T, Nagato T, Kobayashi H,
Komabayashi Y, Ueda S, Kishibe K, Ohkuri T, Takahara M, Celis E and
Harabuchi Y: CCL17 and CCL22/CCR4 signaling is a strong candidate
for novel targeted therapy against nasal natural killer/T-cell
lymphoma. Cancer Immunol Immunother. 64:697–705. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Niens M, Visser L, Nolte IM, van der
Steege G, Diepstra A, Cordano P, Jarrett RF Te Meerman GJ, Poppema
S and van den Berg A: Serum chemokine levels in Hodgkin lymphoma
patients: Highly increased levels of CCL17 and CCL22. Br J
Haematol. 140:527–536. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pere H, Montier Y, Bayry J,
Quintin-Colonna F, Merillon N, Dransart E, Badoual C, Gey A, Ravel
P, Marcheteau E, et al: A CCR4 antagonist combined with vaccines
induces antigen-specific CD8+ T cells and tumor immunity against
self antigens. Blood. 118:4853–4862. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bangham CR and Toulza F: Adult T cell
leukemia/lymphoma: FoxP3(+) cells and the cell-mediated immune
response to HTLV-1. Adv Cancer Res. 111:163–182. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brunstein CG, Miller JS, McKenna DH,
Hippen KL, DeFor TE, Sumstad D, Curtsinger J, Verneris MR,
MacMillan ML, Levine BL, et al: Umbilical cord blood-derived T
regulatory cells to prevent GVHD: Kinetics, toxicity profile, and
clinical effect. Blood. 127:1044–1051. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seay HR, Putnam AL, Cserny J, Posgai AL,
Rosenau EH, Wingard JR, Girard KF, Kraus M, Lares AP, Brown HL, et
al: Expansion of human tregs from cryopreserved umbilical cord
blood for GMP-compliant autologous adoptive cell transfer therapy.
Mol Ther Methods Clin Dev. 4:178–191. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Theil A, Tuve S, Oelschlagel U, Maiwald A,
Döhler D, Oßmann D, Zenkel A, Wilhelm C, Middeke JM, Shayegi N, et
al: Adoptive transfer of allogeneic regulatory T cells into
patients with chronic graft-versus-host disease. Cytotherapy.
17:473–486. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Brunstein CG, Miller JS, Cao Q, McKenna
DH, Hippen KL, Curtsinger J, Defor T, Levine BL, June CH,
Rubinstein P, et al: Infusion of ex vivo expanded T regulatory
cells in adults transplanted with umbilical cord blood: Safety
profile and detection kinetics. Blood. 117:1061–1070. 2011.
View Article : Google Scholar : PubMed/NCBI
|