Introduction
Ovarian cancer is one of the most common
gynecological malignancies. Ovarian cancer can be resistance to
chemotherapy, radiotherapy and targeted therapies (1,2).
Mutations of p53 and KRAS are common in ovarian cancer (3,4).
Understanding the molecular mechanism underlying ovarian cancer may
facilitate the development of novel therapies.
Cellular senescence induces cell cycle arrest
following cellular stress (5). A
previous study observed that senescence is an important
tumor-suppressive mechanism (6).
Furthermore, accumulating evidence has demonstrated that p53, p21
[encoded by the cyclin dependent kinase inhibitor (CDKN)1A gene],
p16 (encoded by CDKN2A) and retinoblastoma protein may have
principal roles in regulating senescence (7). Genetic mutations in the p53 gene or
downregulation of p53 caused by an increase in the expression level
of the p53 ubiquitin ligase MDM2 proto-oncogene (MDM2) were
identified as mechanisms that suppress senescence, and these
processes were observed to cause therapeutic resistance (8).
A previous study observed that senescence occurs in
ovarian cancer (9). However,
whether senescence promotes the progression of ovarian cancer
remains unclear (10,11). A number of previous studies have
demonstrated that chemotherapy drugs induce cellular senescence in
tumor cells (12,13). A recent study demonstrated that
ovarian cancer cells promote hepatocyte growth factor-dependent
senescence of peritoneal mesothelial cells, which may be involved
in the formation of a metastatic niche for ovarian cancer cells
within the peritoneal cavity (14,15).
Therefore, characterization of the mechanisms underlying senescence
in ovarian cancer may facilitate the development of novel
treatments.
Calbindin 1 (CALB1) is a member of the
calcium-binding protein superfamily that includes calmodulins and
troponin C (16). CALB1 contains
four active calcium-binding domains and two modified domains that
are unable to bind calcium (17).
CALB1 was demonstrated to regulate calcium influx following the
activation of glutamate receptors (18). Furthermore, genetic mutations in
CALB1 gene have been observed in patients with Huntington disease
(19). However, the role of CALB1
in cancer remains unknown.
In the present study, the expression pattern of
CALB1 in ovarian cancer was examined. Additionally, the mechanisms
associated with the role of CALB1 in the progression of this
malignancy were investigated.
Materials and methods
Cell culture and transfection
Ovarian cancer cell lines (OVCA429, OVCA433 and
OVCAR3) and normal ovarian epithelial cells (IOSE144) were
purchased from The Cell Bank of Shanghai Institutes for Biological
Science. Cells were maintained in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml streptomycin
(Thermo Fisher Scientific, Inc.) in an incubator with 5%
CO2 at 37°C.
In total, 106 cells were plated in each
dish 18 h before transfection. A total of 8 µg plasmid was
transfected into ovarian cancer cells using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacture's protocol. Cells were incubated with antibiotics (100
U/ml penicillin and 100 mg/ml streptomycin) for 3 days, and the
resistant cells were pooled and used for the subsequent
experiments.
Clinical samples
In total, 30 ovarian cancer samples and paired
non-cancerous tissues were collected from patients who underwent
surgery at The First People's Hospital of Jining (Jining, China)
between April 2009 and March 2015. No treatment was performed prior
to surgery. Written informed consent was obtained prior to the
surgery. The collected tissues were stored in liquid nitrogen. The
present study was approved by The Ethics Committee of The First
People's Hospital of Jining.
Western blot analysis
The proteins were extracted from tissues and cell
lines using RIPA lysis buffer (Cell Signaling Technology, Inc.),
the protein concentration was measured by bicinchoninic acid assay.
In total, 20 µg protein was loaded in each lane. Proteins were
separated by 8% SDS-PAGE (Sangon Biotech Co., Ltd.). Subsequently,
the proteins were transferred onto a PVDF membrane (EMD Millipore).
Following blocking with 5% BSA (Sangon Biotech Co., Ltd.) for 1 h
at room temperature, the membranes were incubated with primary
antibodies overnight at 4°C. The membranes were subsequently washed
with TBS-Tween-20 (0.5%) and incubated with the appropriate
horseradish peroxidase-conjugated secondary antibody (cat. no.
7074; Cell Signaling Technology, Inc.) for 1 h at room temperature.
The proteins were visualized using an enhanced chemiluminescence
kit (Pierce; Thermo Fisher Scientific, Inc.). Antibodies anti-CALB1
(1:3,000; cat. no. ab108404), p16 (1:3,000; cat. no. ab51243), p21
(1:3,000; cat. no. ab218311), p27 (also known as CDKN1B; 1:3,000;
cat. no. ab32034), Tubulin (1:6,000; cat. no. ab210797),
glutathione S-transferase (GST; 1:5,000; cat. no. ab111947), human
influenza hemagglutinin (HA)-tag (1:5,000; cat. no. ab9110) and
Flag (1:5,000; cat. no. ab49763) were purchased from Abcam.
Anti-GAPDH (1:5,000; cat. no. 5174), myc (1:1,000; cat. no. 2276),
β-actin (1:5,000; cat. no. 3700) and MDM2 (1:1,000; cat. no. 86934)
were purchased from Cell Signaling Technology, Inc.
Immunohistochemistry (IHC)
The 5-µm-thick sections were deparaffinized and
rehydrated using xylene (100%) and descending ethanol series
(75–100%). Each incubation was performed for 10 min at room
temperature. Subsequently, 0.3% H2O2 for 10
min at room temperature was used to block endogenous peroxidase
activity. Subsequently, the antigens were retrieved using sodium
citrate (pH 6.0, 0.01 M) at 98°C for 20 min). Non-specific binding
was blocked using 5% BSA (Sangon Biotech Co., Ltd.) at room
temperature for 1 h. The sections were stained with CALB1 primary
antibody (Abcam) at 4°C overnight and visualized with the
appropriate horseradish-conjugated secondary antibody using an
EnVision system (Agilent Technologies, Inc.) according to the
manufacturer's protocol. Subsequently, the slides were developed
with 3,3′-diaminobenzidine at room temperature for 4 min and
counterstained with hematoxylin at room temperature for 4 min. The
staining intensity and the protein expression levels were evaluated
using the Vectra2 system (PerkinElmer, Inc.). The staining was
assessed using the H-score as previously described (20), which was calculating by multiplying
the percentage of positive cells by the staining intensity of the
tumor cells (ranging between 0 and 3).
Senescence-associated β-galactosidase
(β-gal) staining
Cells at 80% confluence were washed twice with PBS
and fixed with 4% formaldehyde for 5 min at room temperature.
Subsequently, cells were incubated at 37°C with β-gal staining
solution (Beyotime Institute of Biotechnology) overnight according
to the manufacturer's protocol. Stained cells were visualized using
an inverted phase contrast light microscope (Olympus Corporation).
In total, >300 random cells were analyzed per sample, and
percentages of stained cells were calculated.
GST pull-down
The coding sequence of CALB1 was cloned into the
expression vector pGEX-4T-1 (Clontech Laboratories, Inc.). A PCR
was performed using cDNA derived from OVCA429 cells as template.
The primers used to clone CALB1 were as follows: Forward,
5′-ATGGCAGAATCCCACCTGCA-3′ and reverse,
5′-CTAGTTATCCCCAGCACAGAG-3′. The PCR was performed using KOD
polymerase (Takara Bio, Inc.). The thermocycling conditions of the
PCR were as follows: Initial denaturation at 95°C for 5 min,
followed by 35 cycles of 94°C for 30 sec, 56°C for 1 min, 72°C for
2 min, with a final extension at 72°C for 10 min. The fusion
protein GST-CALB1 was purified as previously described (21). The whole cell lysate of OVCA429
cells was extracted using a lysis buffer (containing 50 mM Tris-HCl
pH 7.5, 150 mM NaCl, 0.1% Tergitol-type NP-40 and a protease
inhibitor cocktail). The protein concentration was determined using
the bicinchoninic acid assay. Subsequently, 5 µg GST-CALB1 fusion
protein and 500 µg total protein from OVCA429 cells were incubated
overnight at 4°C. In total, 50 µl Glutathione Sepharose 4B beads
(GE Healthcare) were added to the samples and incubated at 4°C for
1 h to allow the binding between the beads and the GST fusion
protein. Following three washes with lysis buffer, the proteins
were eluted in Laemmli buffer and analyzed by SDS-PAGE as
aforementioned.
Immunoprecipitation assay
Cells were lysed with RIPA buffer (Cell Signaling
Technology, Inc.). Following centrifugation at 4°C for 20 min at
12,000 × g, the supernatant of the cell lysate was incubated with a
primary antibody overnight at 4°C. Subsequently, the supernatant
was incubated with 50 µl protein A beads (GE Healthcare) for 4 h at
4°C, and the beads were washed with RIPA buffer three times. The
immunoprecipitated proteins were mixed with Laemmli (Cell Signaling
Technology, Inc.) buffer and boiled for 5 min at 100°C prior to
western blot analysis as aforementioned.
Plasmids
The coding sequence of CALB1 was amplified by PCR
and inserted into the expression vector pcDNA3.1 (Clontech
Laboratories, Inc.) to construct the myc-tagged CALB1. The coding
sequence of MDM2 was amplified by PCR and inserted into the
expression vector pCMVTag2B (Clontech Laboratories, Inc.) to
construct the Flag-tagged MDM2. The coding sequence of p53 was
amplified by PCR and inserted into expression vector pCMV-HA
(Clontech Laboratories, Inc.) to construct the HA-tagged p53. The
cDNA was synthesized using RNA extracted from ovarian cancer cells
as aforementioned. The primer sequences for MDM2 and p53 were as
follows: MDM2 forward, 5′-ATGTGCAATACCTACATGTCTGTACC-3′, MDM2
reverse, 5′-CTAGGGGAAATAAGTTAGCACAA-3′; P53 forward,
5′-ATGGAGGAGCCGCAGTCAGATCCTAGCG-3′ and P53 reverse,
5′-TCAGTCTGAGTCAGGCCCTTCTGT-3′. The PCR was performed using KOD
polymerase (Takara Bio, Inc.). The thermocycling conditions of the
PCR were as follows: Initial denaturation at 95°C for 5 min,
followed by 35 cycles of 94°C for 30 sec, 56°C for 1 min and 72°C
for 2 min, with a final extension at 72°C for 10 min.
Knockdown of CALB1 expression
To knockdown CALB1, P21 and P27, lentiviral
particles (108 PFU) containing short hairpin
(sh)-control or sh-CALB1, sh-P21 and sh-P27 (Shanghai GeneChem Co.,
Ltd.) were used to infect cells at 60% confluence for 24 h, and
cells exhibiting stable knockdown were selected using DMEM
containing puromycin (2 µg/ml) for ≥1 week. The sequences for
sh-CALB1 were as follows: sh-CALB1 #1, 5′-AATCCCACCTGCAGTCATCCC-3′;
sh-CALB1 #2, 5′-AATATGATACTGACCACAGTG-3′. The sequences for the
additional sh-RNAs were as follows: sh-con,
5′-ACCACATCGCGTCTACACCTC-3′; sh-P21, 5′-AACCGGCTGGGGATGTCCGTC-3′;
sh-P27, 5′-AAGAGTTAACCCGGGACTTGG-3′. In the senescence assay, only
one sh-RNA for each gene was used. Only the sh-P21/P27 that was
used in the transfection efficiency experiment was used in the
senescence assay experiments.
MTT assay
Cells were plated in 96-well plates at a density of
1×103 cells/well. Cell proliferation was determined
using the MTT colorimetric assay (R&D Systems, Inc.) over 1
week. Every other day, cell proliferation was determined following
incubation with the MTT solution (50 µg/well) for 4 h. Following
the MTT incubation, the purple formazan crystals were dissolved
using DMSO (Sigma-Aldrich; Merck KGaA) and proliferation was
analyzed at a wavelength of 540 nm. All experiments were performed
in triplicate.
Soft agar colony formation assay
For the soft agar assay, 12-well plates were coated
with solidified DMEM-agarose [0.5% agarose (Sangon Biotech Co.,
Ltd.) in DMEM supplemented with 10% FBS]. Subsequently,
2×103 cells/well suspended in a DMEM-agarose liquid
solution (0.35% agarose in DMEM supplemented with 10% FBS) were
added in the pre-coated wells. Following 14 days of incubation, the
colonies were counted and measured using a light microscope
(magnification, 20×). All experiments were performed ≥3 times.
Gene Expression Profiling Interactive
Analysis tool (GEPIA) database analysis
GEPIA (http://gepia.cancer-pku.cn/) is a tool used to deliver
fast and customizable functionalities based on TCGA and GTEx
databases (22), including
differential expression analysis, profiling plotting, correlation
analysis, patient survival analysis and similar gene detection, as
previously described (19). In the
present study, GEPIA was used to investigate the association of
CALB1 with disease-free survival rates of patients with ovarian
cancer. Patients were grouped into high expression group and low
expression group according to the median value of gene
expression.
Statistical analysis
Data are presented as the mean ± SEM. Each
experiment was repeated three times. Two-tailed Student's t-test
was performed to compare data between two groups. One-way ANOVA
followed by Student-Newman-Keuls post hoc test was used to compare
multiple groups. Kaplan-Meier analysis and log-rank tests were used
for the survival analyses. Data were analyzed using the Prism
software 5.0 (GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression level of CALB1 is increased
in ovarian cancer
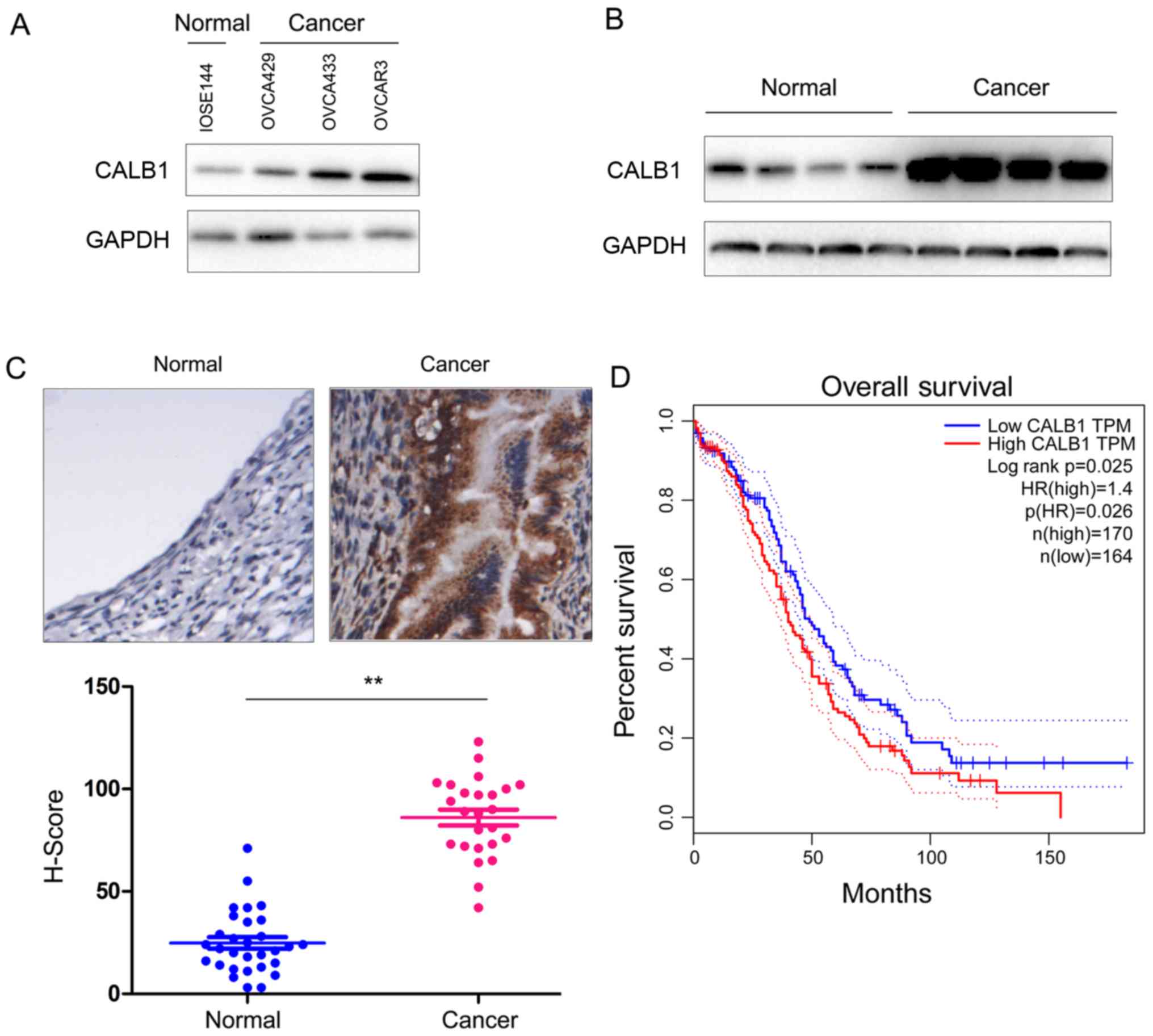
To examine the expression of CALB1 in ovarian
cancer, the protein expression level of CALB1 was assessed in
healthy and ovarian cancer cells. The present results demonstrated
that the protein expression level of CALB1 was increased in cancer
cells compared with the normal cell line (Fig. 1A). Subsequently, the protein
expression level of CALB1 was analyzed in ovarian cancer samples
and adjacent non-cancerous tissues using western blot analysis and
IHC. The present results suggested that the protein expression
level of CALB1 was increased in cancer tissues (Fig. 1B and C). Furthermore, the analysis
of the Gene Expression Profiling Interactive Analysis database
(gepia.cancer-pku.cn/detail.php?gene=CALB1) demonstrated that an
increased expression of CALB1 was associated with a reduced
survival rate (Fig. 1D). The
present results suggested that the protein expression level of
CALB1 is upregulated in ovarian cancer cell lines and tissues, and
its expression level may be associated with poor prognosis in
patients with ovarian cancer.
CALB1 promotes proliferation and
inhibits senescence of ovarian cancer cells
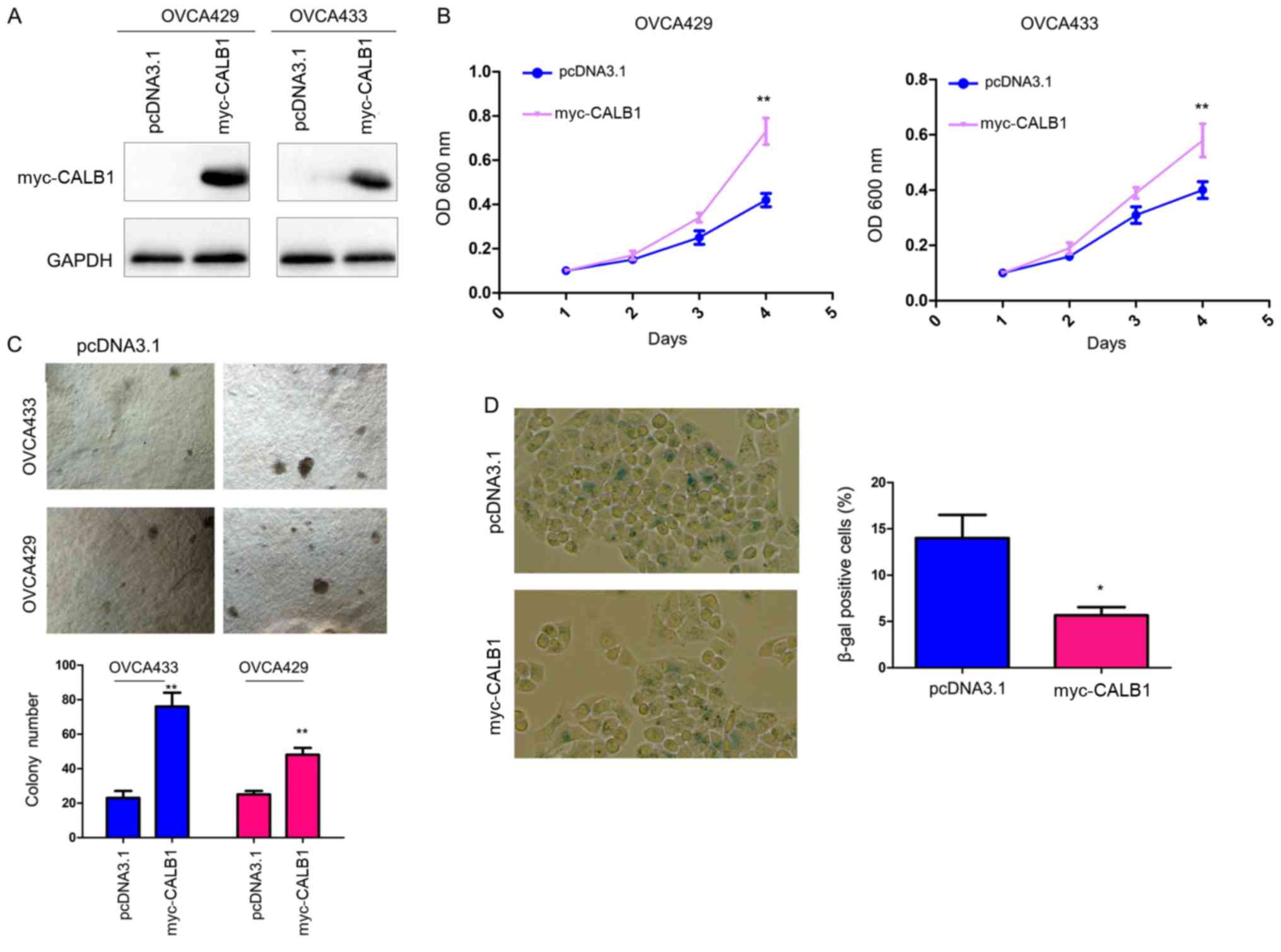
To examine the biological functions of CALB1 in
ovarian cancer, CALB1 was overexpressed in OVCA429 and OVCA433
cells (Fig. 2A). MTT and soft agar
assays were used to examine the effects of CALB1 overexpression on
the proliferation of ovarian cancer cells. Overexpression of CALB1
increased the proliferation of cancer cells in liquid medium after
4 days and the anchorage-independent cell proliferation in soft
agar, as assessed by MTT and colony formation assay, respectively
(Fig. 2B and C). Notably, the
β-gal staining results suggested that the number of control cells
positive for β-gal was higher than cells overexpressing CALB1
(Fig. 2D), suggesting that CALB1
may inhibit senescence in ovarian cancer cells.
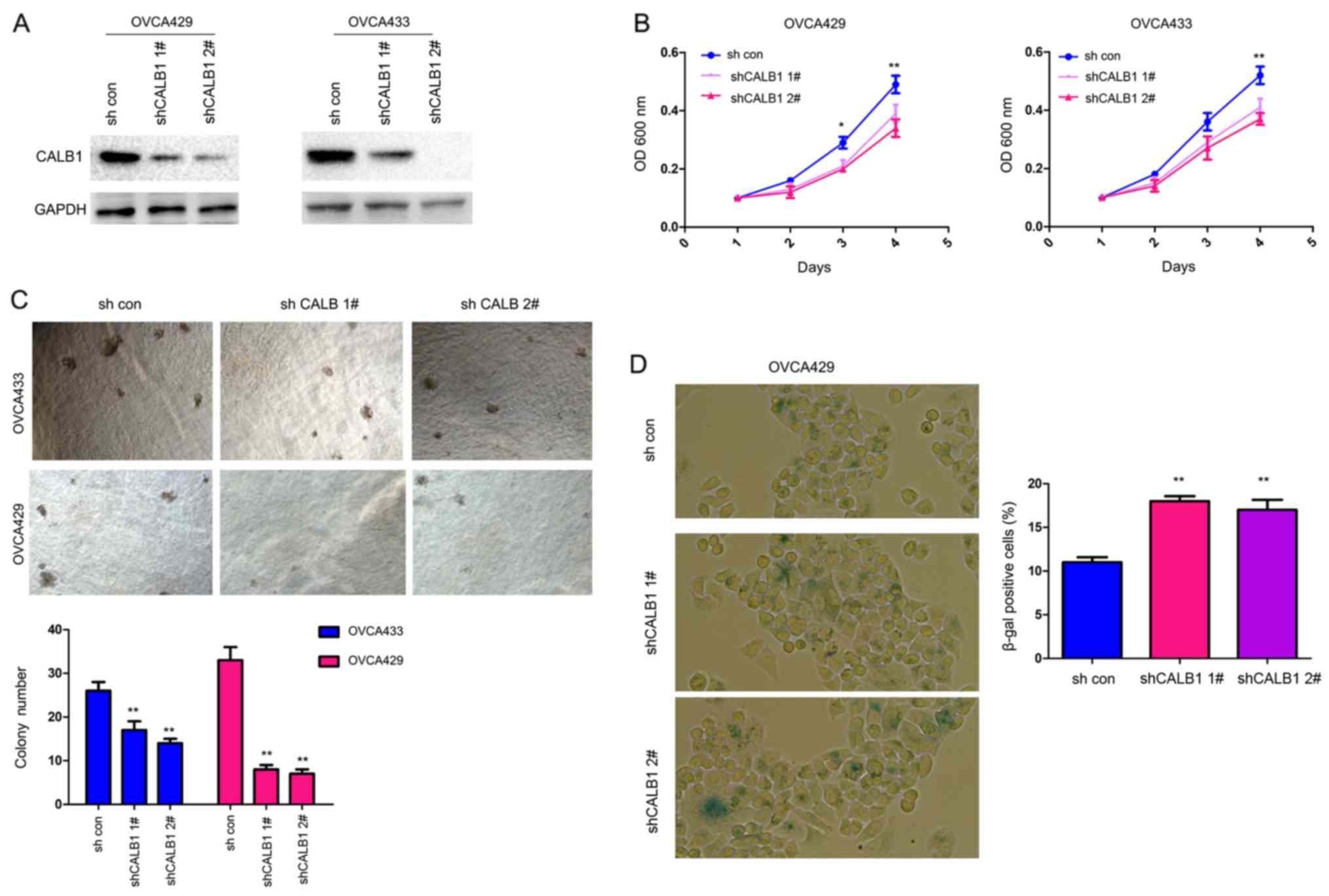
To further investigate the biological functions of
CALB1 in ovarian cancer cells, the expression of CALB1 was silenced
using two sh-RNA sequences targeting CALB1 in OVCA429 and OVCA433
cells (Fig. 3A). Knockdown of
CALB1 inhibited the proliferation of ovarian cancer cells in liquid
culture after 3 days in OVCA429 cells and after 4 days in both cell
types (Fig. 3B). Additionally,
knockdown of CALB1 decreased the colony formation of ovarian cancer
cells in soft agar (Fig. 3C).
Furthermore, following transfection with sh-CALB1, the number of
senescent cells increased, as assessed by β-gal staining (Fig. 3D). Collectively, the present data
suggested that CALB1 promoted proliferation and inhibited
senescence of ovarian cancer cells.
p21 and p27 are involved in the
sh-CALB1-induced senescence
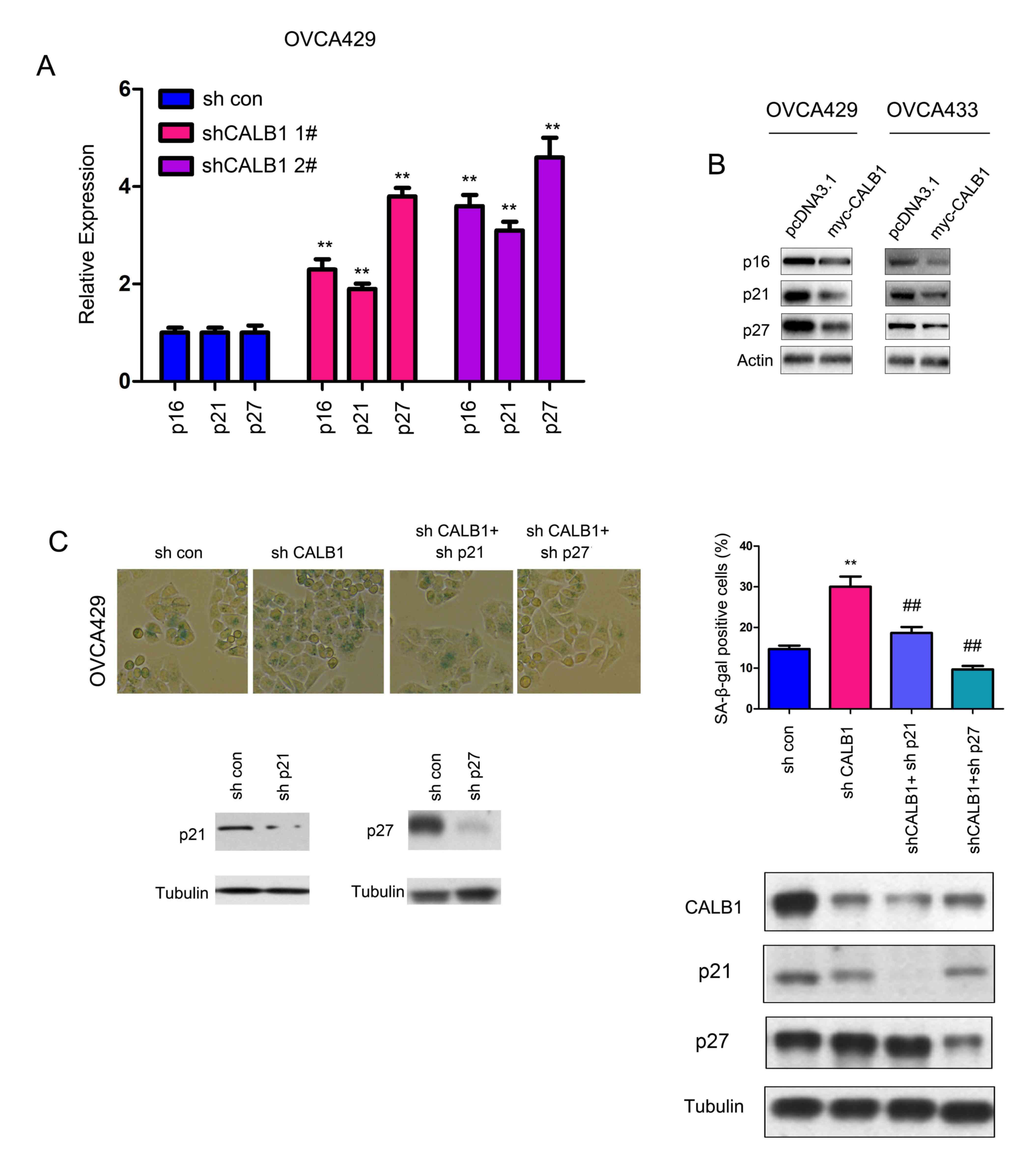
The protein expression levels of various regulators
of senescence were examined. In particular, the protein expression
levels of p16, p21 and p27 were assessed by western blot analysis.
Knocking down the expression of CALB1 significantly upregulated the
mRNA expression levels of p16, p21 and p27 (Fig. 4A). Conversely, overexpression of
CALB1 decreased the protein expression level of p16, p21 and p27
(Fig. 4B). Moreover, knocking down
the expression of p21 and p27 abolished the senescence induced by
sh-CALB1, as assessed by β-gal staining (Fig. 4C). The present data suggested that
p21 and p27 were responsible for the increase in senescence induced
by CALB1 knockdown.
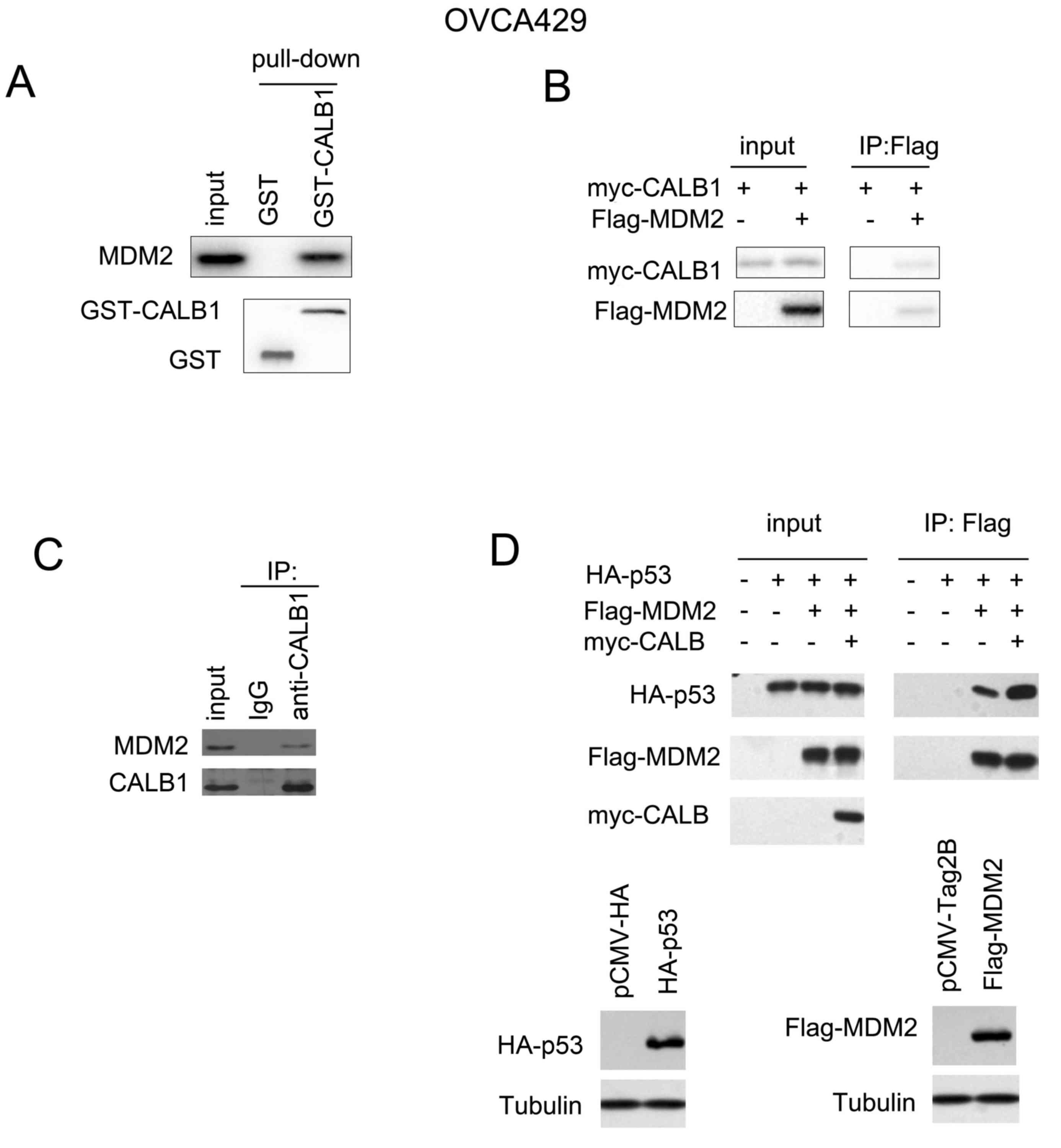
CALB1 interacts with MDM2, and
promotes the interaction between MDM2 and p53
Numerous previous studies have demonstrated that p21
and p27 are downstream target genes of p53 (23). Therefore, the interaction between
CALB1 and other members of the p53 signaling pathway was
investigated in the present study. In the GST pull down assay, the
fusion protein GST-CALB1 interacted with endogenously expressed
MDM2 (Fig. 5A), as confirmed by
the co-immunoprecipitation assay (Fig.
5B and C). Furthermore, CALB1 enhanced the interaction between
p53 and MDM2 (Fig. 5D), suggesting
that CALB1 may promote the degradation of p53 via MDM2.
Discussion
Numerous previous studies have demonstrated that
cellular senescence contributes to tumor suppression (24). However, the molecular mechanism
underlying this process remains unclear. In the present study,
CALB1 was identified to be upregulated in ovarian cancer.
Functional analysis suggested that CALB1 promoted the proliferation
and colony formation of ovarian cancer cells by inhibiting
senescence. The molecular experiments demonstrated the interaction
between MDM2 and CALB1, and CALB1 enhanced the interaction between
p53 and MDM2, thus potentially leading to the degradation of the
p53 tumor suppressor. In line with this hypothesis, CALB1 knockdown
upregulated the expression levels of p16, p21 and p27, and
overexpression of CALB1 decreased the protein expression level of
p16, p21 and p27. The present results suggested that CALB1 may act
as an oncogene in ovarian cancer.
The present study identified the oncogenic roles of
CALB1 in ovarian cancer. To the best of our knowledge, a few
previous studies have investigated the roles of CALB1 in
carcinogenesis (25). The present
study suggested that CALB1 may act as an oncogene in ovarian
cancer, and CALB1 may represent a novel therapeutic target to treat
ovarian cancer.
In the present study, knockdown of p21 and p27, two
target genes of p53 (26),
suppressed the senescence induced by CALB1 knockdown, which further
emphasized the roles of p53 and its target genes in the process of
senescence. It is established that ovarian cancer develops due to
accumulation of multiple genetic mutations (27). In addition to KRAS, other
regulators of senescence, such as p53 and p16, are frequently
mutated in human ovarian cancer (28).
In summary, the present study suggested that CALB1
may act as an oncogene in ovarian cancer by inhibiting senescence,
and suggested that CALB1 may be a novel therapeutic target for the
treatment of ovarian cancer. Further experiments using a large
number of clinical samples are required to investigate the
potential to use CALB1 as an ovarian cancer biomarker.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MP and ML designed the present study, and
interpreted and analyzed the data. YW and LC performed the
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved by The Ethics Committee of
The Seventh People's Hospital of Jinan, and written informed
consent was obtained from all patients.
Patient consent for publication
All patients within this study provided consent for
the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics, 2018. CA Cancer J Clin. 68:284–296.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mori R, Futamura M, Morimitsu K, Saigo C,
Miyazaki T and Yoshida K: The diagnosis of a metastatic breast
tumor from ovarian cancer by the succession of a p53 mutation: A
case report. World J Surg Oncol. 15:1172017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakayama N, Nakayama K, Yeasmin S,
Ishibashi M, Katagiri A, Iida K, Fukumoto M and Miyazaki K: KRAS or
BRAF mutation status is a useful predictor of sensitivity to MEK
inhibition in ovarian cancer. Br J Cancer. 99:2020–2028. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bennett MR and Clarke MC: Basic research:
Killing the old: Cell senescence in atherosclerosis. Nat Rev
Cardiol. 14:8–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Burgess DJ: Senescence: Double or quit?
Nat Rev Cancer. 11:3892011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mathon NF and Lloyd AC: Cell senescence
and cancer. Nat Rev Cancer. 1:203–213. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Burgess DJ: Senescence: Tumorigenesis
under surveillance. Nat Rev Cancer. 12:62011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Visser JA, Schipper I, Laven JS and
Themmen AP: Anti-Mullerian hormone: An ovarian reserve marker in
primary ovarian insufficiency. Nat Rev Endocrinol. 8:331–341. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nardella C, Clohessy JG, Alimonti A and
Pandolfi PP: Pro-senescence therapy for cancer treatment. Nat Rev
Cancer. 11:503–511. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Campisi J and d'Adda di Fagagna F:
Cellular senescence: When bad things happen to good cells. Nat Rev
Mol Cell Biol. 8:729–740. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Killock D: Chemotherapy: Life gained,
years lost? Nat Rev Clin Oncol. 11:3032014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Collado M and Serrano M: Senescence in
tumours: Evidence from mice and humans. Nat Rev Cancer. 10:51–57.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Mao Q, He J, She H, Zhang Z and Yin
C: Human umbilical cord mesenchymal stem cells improve the reserve
function of perimenopausal ovary via a paracrine mechanism. Stem
Cell Res Ther. 8:552017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mikuła-Pietrasik J, Uruski P, Pakuła M,
Maksin K, Szubert S, Woźniak A, Naumowicz E, Szpurek D, Tykarski A
and Książek K: Oxidative stress contributes to hepatocyte growth
factor-dependent pro-senescence activity of ovarian cancer cells.
Free Radic Biol Med. 110:270–279. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jung EM, Choi KC and Jeung EB: Expression
of calbindin-D28k is inversely correlated with proapototic gene
expression in hydrogen peroxide-induced cell death in endometrial
cancer cells. Int J Oncol. 38:1059–1066. 2011.PubMed/NCBI
|
|
17
|
Goffigan-Holmes J, Sanabria D, Diaz J,
Flock D and Chavez-Valdez R: Calbindin-1 expression in the
hippocampus following neonatal hypoxia-ischemia and therapeutic
hypothermia and deficits in spatial memory. Dev Neurosci. March
12–2019.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Parkash J, Chaudhry MA and Rhoten WB:
Calbindin-D28k and calcium sensing receptor cooperate in MCF-7
human breast cancer cells. Int J Oncol. 24:1111–1119.
2004.PubMed/NCBI
|
|
19
|
Massouh M, Wallman MJ, Pourcher E and
Parent A: The fate of the large striatal interneurons expressing
calretinin in Huntington's disease. Neurosci Res. 62:216–224. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cai Z, Qian ZY, Jiang H, Ma N, Li Z, Liu
LY, Ren XX, Shang YR, Wang JJ, Li JJ, et al: hPCL3s promotes
hepatocellular carcinoma metastasis by activating β-catenin
signaling. Cancer Res. 78:2536–2549. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Deng YZ, Yao F, Li JJ, Mao ZF, Hu PT, Long
LY, Li G, Ji XD, Shi S, Guan DX, et al: RACK1 suppresses gastric
tumorigenesis by stabilizing the β-catenin destruction complex.
Gastroenterology. 142:812–823.e15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levine AJ: The p53 tumor suppressor gene
and gene product. Princess Takamatsu Symp. 20:221–230.
1989.PubMed/NCBI
|
|
24
|
Ohtani N, Mann DJ and Hara E: Cellular
senescence: Its role in tumor suppression and aging. Cancer Sci.
100:792–797. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jin C, Lin T and Shan L: Downregulation of
Calbindin 1 by miR-454-3p suppresses cell proliferation in nonsmall
cell lung cancer in vitro. Cancer Biother Radiopharm. 34:119–127.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang F, von Knethen A and Brune B:
Modulation of nitric oxide-evoked apoptosis by the p53-downstream
target p21(WAF1/CIP1). J Leukoc Biol. 68:916–922. 2000.PubMed/NCBI
|
|
27
|
Labidi-Galy SI, Olivier T, Rodrigues M,
Ferraioli D, Derbel O, Bodmer A, Petignat P, Rak B, Chopin N,
Tredan O, et al: Location of mutation in BRCA2 gene and survival in
patients with ovarian cancer. Clin Cancer Res. 24:326–333. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schuyer M, van Staveren IL, Klijn JG, vd
Burg ME, Stoter G, Henzen-Logmans SC, Foekens JA and Berns EM:
Sporadic CDKN2 (MTS1/p16ink4) gene alterations in human ovarian
tumours. Br J Cancer. 74:1069–1073. 1996. View Article : Google Scholar : PubMed/NCBI
|