Introduction
Melanoma is an aggressive form of skin cancer, which
is formed by the malignant transformation of melanocytes located at
the basement of the epidermis, deriving from blemishes or pigmented
spots (1,2). It is one of the most lethal
malignancies globally, particularly in fair-skinned individuals,
accounting for 80% of skin cancer-related mortality (3). Although efforts have been made to
develop improved understanding of melanoma progression and more
effective treatments, they have met with limited success, mainly
due to its high metastatic potential and poor prognosis (4). Therefore, further investigation into
the molecular mechanisms underlying the development and progression
of melanoma is urgently required to design new therapeutic
strategies and improve clinical outcomes. Previously, it has been
found that the abnormal expression of genes, such as B-Raf and
Asparaginyl-TNRA synthetase, serves an important role in the
formation and metastasis of melanoma (5,6).
Targeted manipulation of the altered expression or regulatory
mechanisms of these genes may be a potential treatment strategy
(6).
MicroRNAs (miRNAs/miRs), a class of short,
endogenous, noncoding RNAs ranging between 19 and 25 nucleotides in
length, were found to participate in a variety of physiological and
pathological activities, including cell proliferation,
transformation and tumorigenesis (7). Furthermore, the function of miRNA in
gene regulation is exerted primarily via binding to the
untranslated region (UTR) of target genes, which leads to mRNA
degradation and/or reduced translation (8,9).
Generally, an individual miRNA can regulate the expression of
multiple target genes, and several miRNAs can synergistically act
on one target gene (10). At
present, several miRNAs have been demonstrated to contribute to the
aggressive phenotype of melanoma cells (11,12).
For example, enhanced expression of miR-612 suppressed the growth,
invasion and tumorigenesis of melanoma cells by targeting Espin
(13); miR-637 inhibited melanoma
cell proliferation by inactivating the AKT signaling pathway and
induced apoptosis through regulation of Bcl-2/Bax expression by
targeting phosphatidylinositol 3,4,5-triphosphate-dependent Rac
exchanger 2 protein (14); and
miR-143-3p inhibited the growth and invasiveness of melanoma cells
by targeting cyclooxygenase-2, and was inversely correlated with
malignant melanoma progression (15).
miR-21 is an example of an oncomiR that is
upregulated in various types of cancer (16). For instance, reduced expression of
miR-21 replicated the effects of celastrol on OVCAR3 cells and
inhibited the PI3K/phosphorylated-Akt-NF-κB signaling pathway in an
in vitro model of ovarian carcinoma (17). In addition, miR-21 has also been
reported to be involved in the formation of melanoma (18,19).
miR-21 was able to regulate the ERK/NF-κB signaling pathway to
affect the proliferation, migration and apoptosis of human melanoma
A375 cells by targeting protein sprout homolog 1, programmed cell
death protein 4 and PTEN (20,21).
Mitogen-activated protein kinase kinase 3 (MKK3), a member of the
dual-specificity MKK protein kinase group that belongs to the
mitogen-activated protein kinase (MAPK) signaling pathway, was
identified to be associated with tumor invasion and progression
(22). For example, the activation
of the MKK3/MKK6-p38 axis is closely related to the cell survival
of MDA-MB-231 breast cancer cells (23). In the present study, the expression
levels of MKK3 mRNA and protein, and miR-21 were evaluated in
biopsy samples from patients with melanoma, and the functional
effects of miR-21 and MKK3 on melanoma cell proliferation,
apoptosis, cell cycle, migration and invasion were examined, with
the interaction between miR-21 and MKK3 being further
investigated.
Materials and methods
Tissue specimens
Melanoma tissues and their corresponding matched
adjacent normal tissues, that were 5 cm away from the tumor tissue,
were collected from 15 cases of patients with melanoma who did not
receive cutaneous radiotherapy or chemotherapy prior to surgical
resection at the Department of Oncology, Qilu Hospital of Shandong
University (Qingdao, China) between January 2017 and January 2018.
The cohort included 8 males and 7 females, with a median age of 51
years (range, 39–78 years). The diagnosis of melanoma was performed
by two experienced pathologists on the basis of hematoxylin and
eosin staining of 5-µm sections conducted according to the
manufacturer's protocols (cat. no. C0105; Beyotime Institute of
Biotechnology) and immunohistochemistry staining performed as
previously described (23),
following fixation in 4% paraformaldehyde at room temperature for 2
h. Following resection, tumor tissues were flushed with 0.9%
saline, and regions with no necrosis and good vitality were
selected for experiments, while the normal tissues were similarly
collected. Nerves, blood vessels and connective tissues were
removed. Then, all tissues were rapidly snap-frozen in liquid
nitrogen and stored at −80°C until use.
The present study was performed in strict accordance
with the Declaration of Helsinki and approved by the Clinical
Management Committee of Hunan First People's Hospital (Changsha,
China). Written informed consent for research purposes was obtained
from all patients. The information of the patients is presented in
Table I.
 | Table I.Patient information. |
Table I.
Patient information.
| Specimen
number | Type of
melanoma | Tumor
metastasis |
|---|
| 1 | Primary
melanoma | Yes |
| 2 | Primary
melanoma | Yes |
| 3 | Primary
melanoma | No |
| 4 | Primary
melanoma | No |
| 5 | Primary
melanoma | Yes |
| 6 | Primary
melanoma | No |
| 7 | Primary
melanoma | No |
| 8 | Primary
melanoma | No |
| 9 | Primary
melanoma | Yes |
| 10 | Primary
melanoma | Yes |
| 11 | Primary
melanoma | No |
| 12 | Primary
melanoma | Yes |
| 13 | Primary
melanoma | Yes |
| 14 | Primary
melanoma | No |
| 15 | Primary
melanoma | No |
Cell lines and culture
The melanoma A2058 and HS294T cell lines, skin
fibroblast foreskin BJ and CCC-HSF-1 cell lines, epithelial HME1
cells from mammary glands and 293T cells were purchased from
American Type Culture Collection. 293T cells were cultured in DMEM
(Gibco; Thermo Fisher Scientific, Inc.), whereas all other cell
lines were cultured in RPMI-1640 medium (Thermo Fisher Scientific,
Inc.), respectively, which were both supplemented with 10% FBS
(Invitrogen; Thermo Fisher Scientific, Inc.), 2 mM L-glutamine, 100
U/ml penicillin and 100 µg/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) at a temperature of 37°C in a humidified
incubator with 95% O2 and 5% CO2. HME1 skin
cells were used as controls.
RNA extraction, cDNA synthesis and
reverse transcription-quantitative PCR (RT-qPCR) analysis
Total RNA was obtained from tissues and all cell
lines using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Then,
the isolated RNA was cleared of contaminating genomic DNA using
DNase treatment (Thermo Fisher Scientific, Inc.) and the RT
reaction was performed using a PrimeScript™ RT kit (Takara Bio,
Inc.) in a PCR thermocycling instrument (Applied Biosystems; Thermo
Fisher Scientific, Inc.) at 16°C for 30 min, 42°C for 30 min and
85°C for 5 min to obtain cDNA. Finally, the expression levels of
MKK3 and miR-21 were detected using PCR amplification with a
commercially available SYBR Green qPCR SuperMix kit (Invitrogen;
Thermo Fisher Scientific, Inc.) on an ABI PRISM® 7500
Sequence Detection System (Applied Biosystems; Thermo Fisher
Scientific, Inc.) at 95°C for 2 min, followed by 40 cycles of 95°C
for 15 sec, and 60°C for 32 sec (with data collection at the end of
the 60°C step at each cycle), followed by a melting curve. GAPDH
and U6 small nuclear RNA were used as the endogenous controls to
quantify mRNA and miRNA expression levels, respectively. The fold
changes of gene expression levels were calculated using the
2−∆∆Cq method (24).
The primers used are presented in Table II.
| Table II.Sequences of the primers applied in
the RT-quantitative PCR assays. |
Table II.
Sequences of the primers applied in
the RT-quantitative PCR assays.
| miR or gene | Primer sequences
(5′-3′) |
|---|
| miR-21 | F:
ACACTCCAGCTGGGTAGCTTATCAGACTGA |
|
| R:
TGGTGTCGTGGAGTCG |
| U6 | RT:
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAAAAATATGG |
|
| F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
| MKK3 | F:
TGCTCATGGACCTGGACATC |
|
| R:
GAGGGCTTCACATCTCTGTG |
| GAPDH | F:
CCTGGATACCGCAGCTAGGA |
|
| R:
GCGGCGCAATACGAATGCCCC |
Western blot analysis
Protein lysate preparation from human tissues and
cells was performed on ice using RIPA buffer (Beyotime Institute of
Biotechnology) comprising 150 mM NaCl, 50 mM Tris-HCl (pH 8.0),
0.5% sodium deoxycholate, 1% NP-40 and 0.1% SDS, and supplemented
with 1 mg/ml aprotinin, sodium 1 mM orthovanadate and 0.1 mg/ml
phenylmethylsulfonyl fluoride. Concentrations of protein were
assessed with a bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.). Then, the proteins (30 µg) were separated on a
12% SDS-PAGE gel and transferred to a 0.22-µm PVDF membrane (EMD
Millipore). After blocking the membranes in 5% skimmed milk in TBS
with 0.1% Tween-20 (TBST) for 2 h at room temperature, the PVDF
membranes were incubated in PBS containing dilutions of rabbit
anti-MKK3 (1:1,500; cat. no. ab5428; Abcam) and mouse anti-GAPDH
(1:2,000; cat. no. ab9484; Abcam) antibodies at 4°C overnight.
Subsequently, the PVDF membranes were washed with TBST buffer three
times, and incubated with PBS, including an anti-rabbit (cat. no.
BA1056) or anti-mouse (cat. no. BA1050) secondary antibody (Wuhan
Boster Biological Technology, Ltd.) conjugated to horseradish
peroxidase (1:10,000) for 1 h at room temperature with gentle
agitation. Following three washes in TBST, the membranes were
incubated with ECL substrates (Beyotime Institute of Biotechnology)
for 1 min, and the Amersham Imager 600 system (GE Healthcare Life
Sciences) or X-ray films (Kodak) were used to visualize the target
proteins. All results were normalized to the expression of GAPDH
protein using ImageJ version 1.51 software (National Institutes of
Health).
Transfection and dual-luciferase
reporter assay
All plasmids (pLC3) used were designed and
synthesized by Sangon Biotech Co., Ltd. using sequences purchased
from Guangzhou RiboBio Co., Ltd., including miR-21 mimics (cat. no.
miR10000076-1-5), miR-21 inhibitor (cat. no. miR20000076-1-5a), NC
inhibitor (cat. no. miR1N0000001-2-5), MKK3-overexpression plasmid
and MKK3-small interfering (si) RNA (cat. no. siB08711160442-1-5).
Empty pLC3 vector was used as a NC for all transfections except
inhibitor transfection. A2058 cells plated in 6-well plates at a
density of 2×105 cells/well were transfected with the
various plasmids (1 ng) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Subsequent experiments were performed 48 h
following transfection.
The binding sequences between MKK3 and miR-21 were
predicted using TargetScan version 7.2 software (25). For the dual-luciferase reporter
assay, 293T cells seeded into 24-well plates were transfected with
200 ng miR-21 mimics, miR-21 inhibitor or the corresponding control
vectors along with 50 ng psiCHECK™-2 containing wild-type (WT) or
mutant 3′-UTR of MKK3 mRNA inserted downstream of Renilla
luciferase (Promega Corporation). Following a 48-h incubation, the
cells were harvested and assessed for Renilla and firefly
luciferase signals using a dual-luciferase reporter gene detection
system, according to the manufacturer's protocol (cat. no. E2920;
Promega Corporation).
Cell proliferation evaluation
A2058 cells (1,000 cells/well) were treated with the
indicated conditions in 96-well plates. Cell proliferation was
determined using a Cell Counting Kit-8 (CCK-8) assay according to
the manufacturer's protocol. At 0, 1, 2 and 3 days, 90 µl fresh
culture media and 10 µl CCK-8 solution (cat. no. C0038; Beyotime
Institute of Biotechnology) were added to each sample.
Subsequently, transfected melanoma cells were incubated at 37°C for
2 h. Finally, the absorbance was measured at 450 nm using a
microplate reader (Bio-Rad Laboratories, Inc.).
Soft agar assay
Transfected A2058 cells (1,000 cells/well) were
treated with the indicated conditions in 6-well plates. After 48 h,
the cells were trypsinized, gently mixed with 0.45% agar medium
mixture and re-seeded on 6-well plates covered with a layer of 0.9%
agar in DMEM. After 2 weeks, the colonies were stained with 0.2%
crystal violet for 5 min at room temperature, and quantified by
colony counting.
Apoptosis and cell cycle
assessment
A2058 cells were transfected with control or miR-21
mimics, MKK3 expression vector or small interfering RNA in 6-well
plates. After 48 h of transfection, 2×105 cells were
harvested, centrifuged for 5 min at room temperature within 500 ×
g, and washed three times with cold PBS. For cell apoptosis
analysis, a cell apoptosis detection kit was used, according to the
manufacturer's protocol (cat. no. C1062; Beyotime Institute of
Biotechnology). Briefly, the harvested cells were suspended in 500
ml binding buffer and incubated at room temperature in the dark for
15 min after being labeled with 5 ml Annexin-V-FITC and 5 ml
propidium iodide (PI). For the cell cycle analysis, the cells were
first fixed with 70% ethanol at room temperature for 1 h, and then
incubated with RNase (50 µg/ml) and stained with PI for 30 min at
4°C in the dark. A flow cytometer and WinMDI version 2 software (BD
Biosciences) was used to evaluate apoptosis and the cell cycle. The
apoptosis rate was calculated as the percentage of cells in early +
late apoptosis.
Cell migration and cell invasion
assays
The migration and invasion of A2058 cells were
quantitatively measured using a Transwell chamber assay (Costar;
Corning, Inc.). The cells (5×104 cells) treated with
indicated conditions were seeded on the upper insert coated without
or with 2% Matrigel (Corning, Inc.) in 24-well plates for migration
or invasion assays, respectively. Subsequently, the lower chamber
was filled with 600 µl DMEM supplemented with 10% FBS as a
chemoattractant, followed by an incubation at 37°C with 5%
CO2 for 24 h. Next, the cells were permeated with 100%
methanol for 15 min at room temperature, stained with 0.1% crystal
violet for 20 min at room temperature, washed with PBS three times
and dried at 80°C for 30 min. Non-migrated or non-invaded cells on
the inside of the wells were removed with a cotton swab, and the
the cells were counted in images captured in five random fields
under a light microscope (magnification, ×200; Olympus
Corporation).
Statistical analysis
Statistical calculations were performed using IBM
SPSS software, version 23.0 (IBM Corp.). Linear regression analysis
of MKK3 and miR-21 expression in melanoma tissue was conducted
using Microsoft Excel 2010 (Microsoft Corporation). All cell
experiments were independently repeated at least three times and
all data are presented as the mean ± standard deviation. Protein
and mRNA expression values in individual patient tissues are
presented as the mean ± standard deviation of three replicates. Any
differential expression between two groups was determined using an
independent Student's t-test. Statistical comparisons among
multiple groups were performed using Tukey's one-way analysis of
variance. P<0.05 was considered to indicate a statistically
significant difference.
Results
Upregulation of MKK3 and
downregulation of miR-21 in melanoma
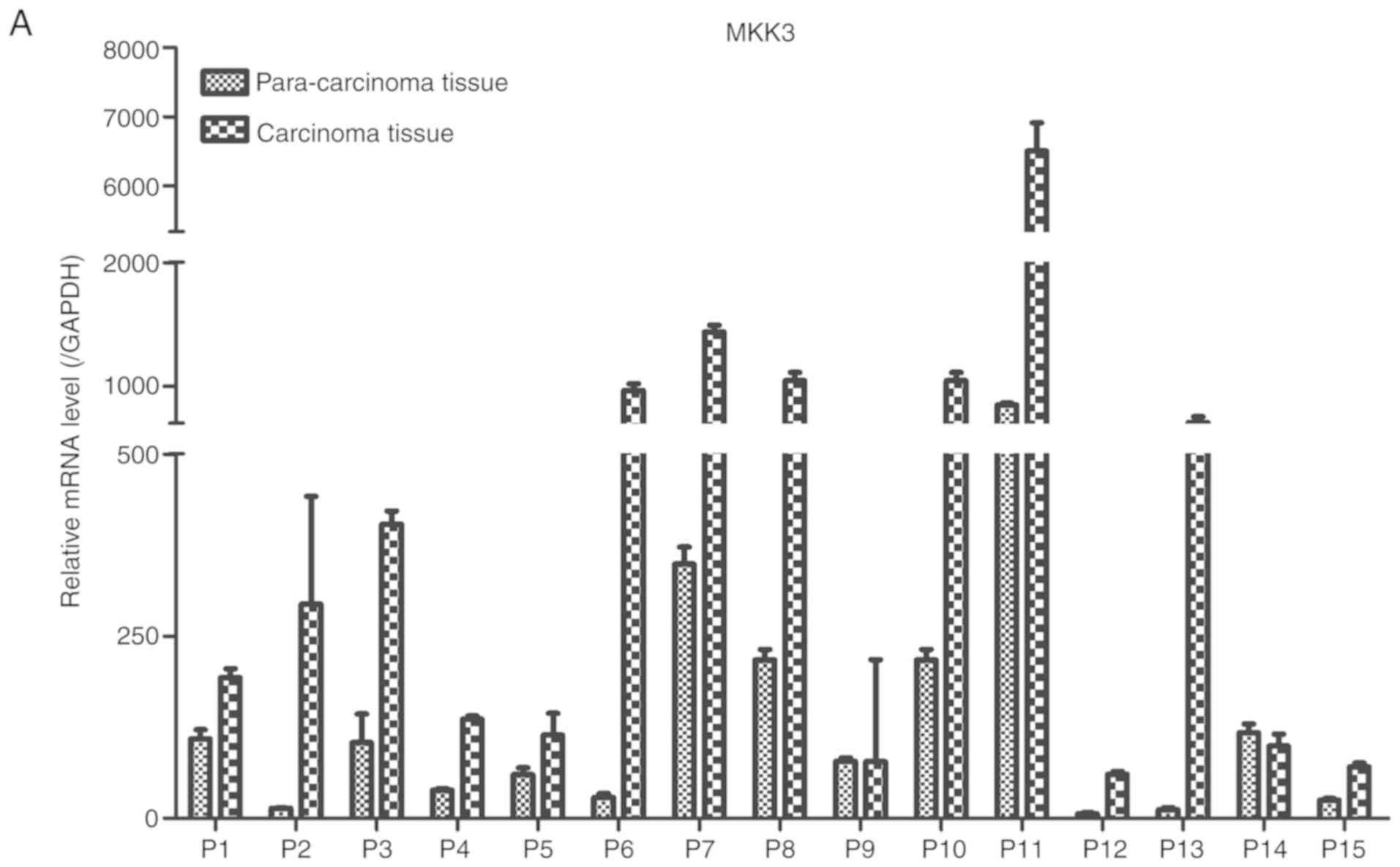
Expression of MKK3 and miR-21 in melanoma was
evaluated via RT-qPCR in 15 pairs of human primary melanoma
specimens and corresponding adjacent skin. The protein expression
levels of MKK3 in the tissues were also assessed using western blot
analysis. The results demonstrated that MKK3 mRNA was upregulated
and MKK3 protein was overexpressed in melanoma biopsy samples
compared with their corresponding adjacent normal tissue samples
(Fig. 1A and B). Conversely, the
miR-21 expression profile was reversed compared with the MKK3
expression in these tissues, which indicated that miR-21 expression
was lower in primary melanoma tissues compared with the matched
adjacent healthy tissues. The mRNA expression of miR-21 was also
determined (Fig. 1C). Therefore,
these findings indicated that MKK3 was upregulated, and miR-21 was
downregulated in melanoma.
MKK3 is significantly increased and
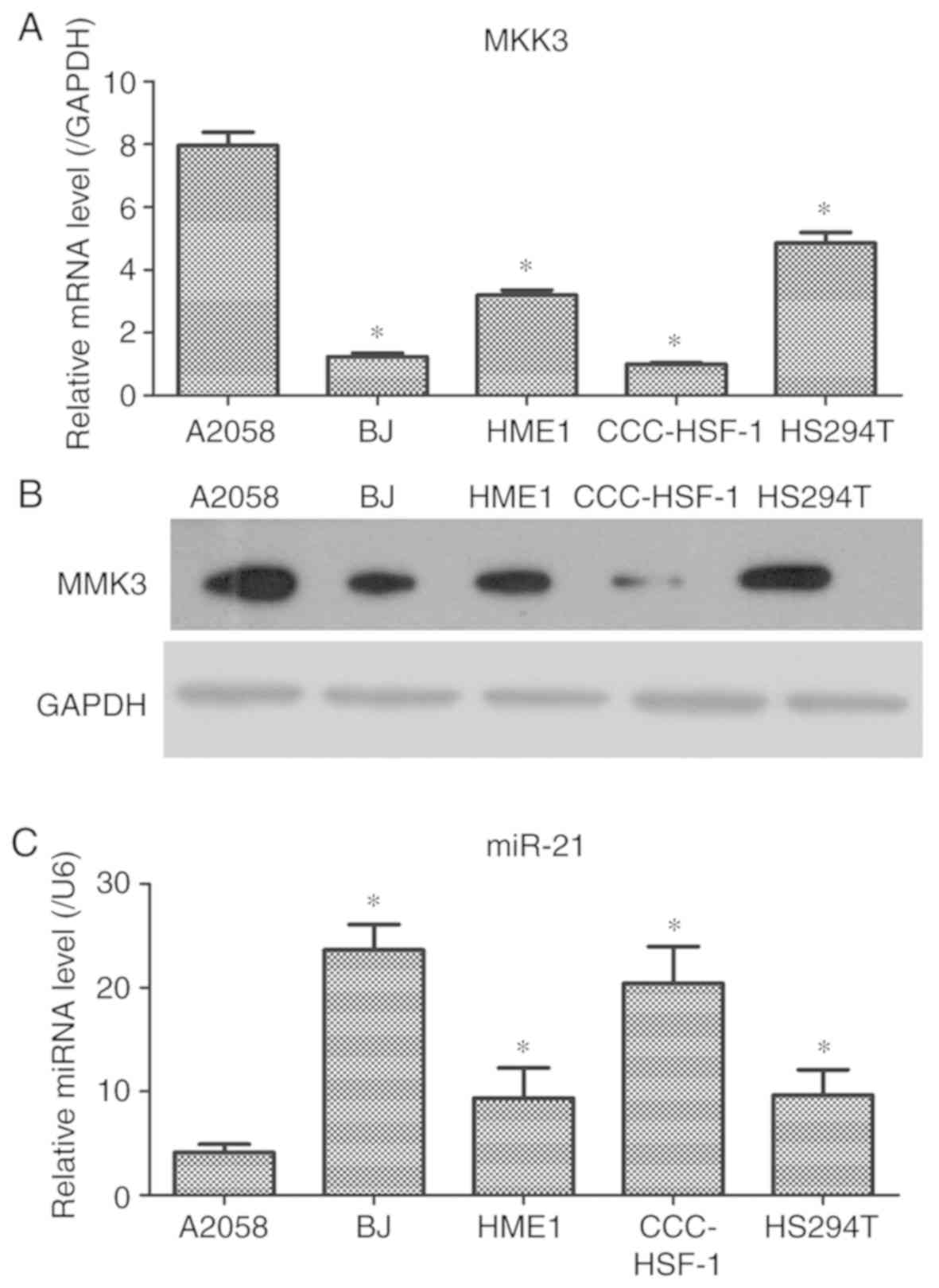
miR-21 is decreased in melanoma cells
The expression levels of MKK3 and miR-21 in melanoma
cells were further examined. The results demonstrated that the mRNA
and protein levels of MKK3 in A2058 cells were significantly
increased compared with HS294T melanoma cells and the control skin
cells (Fig. 2A and B). It was also
observed that miR-21 expression in A2058 cells was significantly
decreased compared with that in HS294T melanoma cells and the
control skin cells (Fig. 2C).
Therefore, these data indicated that MKK3 may be enhanced and
miR-21 may be downregulated in A2058 cells, which was similar to
the results from the clinical specimens; A2058 cells were selected
for the following experiments, as the cell line exhibited the
highest MMK3 and lowest miR-21 expression levels.
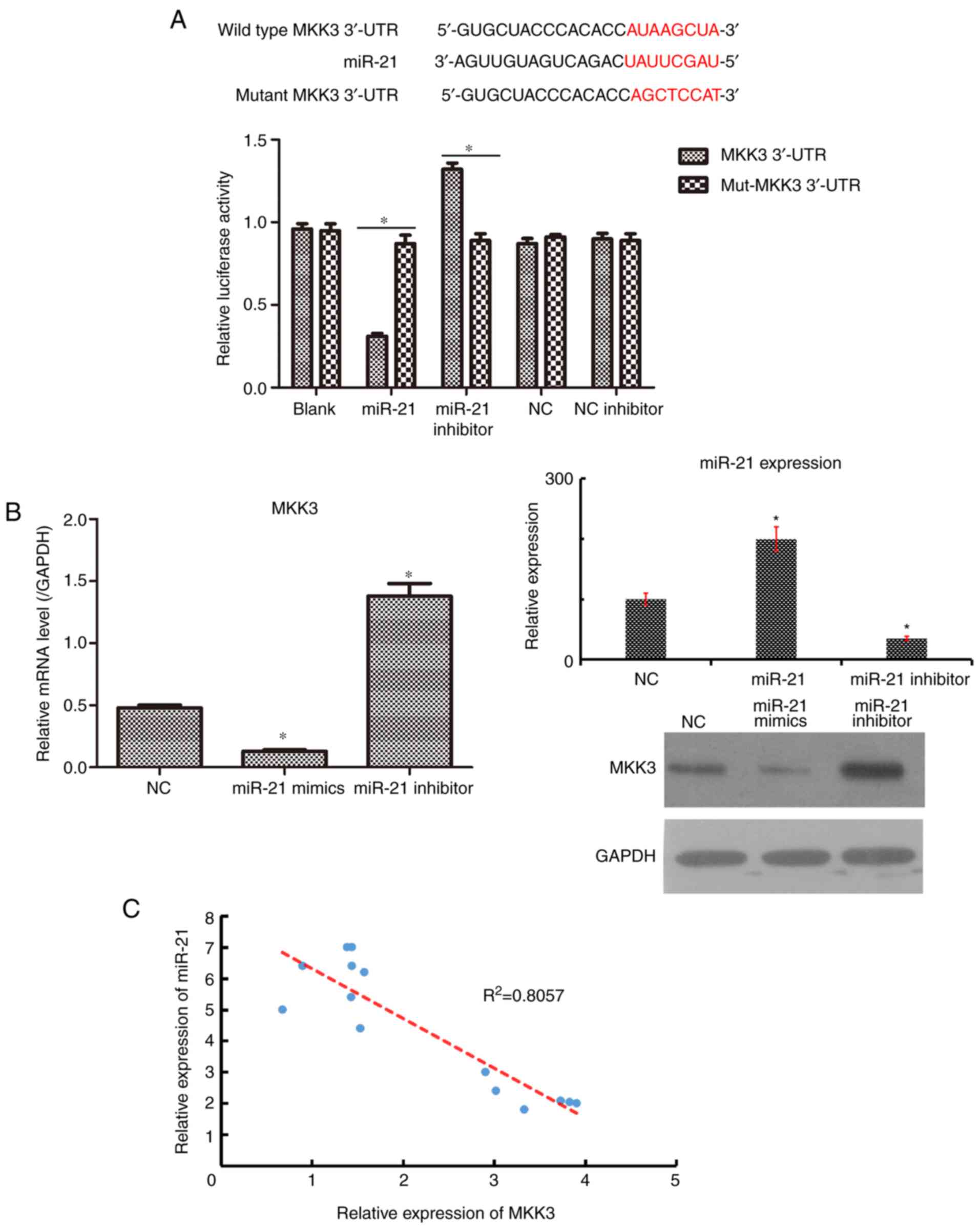
MKK3 is a novel target gene of
miR-21
Based on the inverse expression profiles of MKK3 and
miR-21, the underlying interaction between MKK3 and miR-21 required
investigation. First, TargetScan predicted that MKK3 was a
potential target gene of miR-21 (25). Cells were transfected with plasmids
containing the WT and mutant 3′-UTR of MKK3 with miR-21. A
luciferase reporter assay was conducted to confirm that miR-21
could directly target the 3′-UTR of MKK3. As presented in Fig. 3A, the forced expression of miR-21
decreased the luciferase activity in 293T cells with the WT plasmid
(P<0.05), although no significant change was observed in cells
transfected with the mutant plasmid. To further detect whether
miR-21 could affect MKK3 expression in melanoma cells, RT-qPCR and
western blot analyses were conducted to quantify MKK3 mRNA and
protein expression following transfection with miR-21 mimics and
miR-21 inhibitor in A2058 cells. It was observed that the
upregulation of miR-21 suppressed MKK3 expression at the mRNA and
protein levels, whereas the downregulation of miR-21 promoted MKK3
expression at the mRNA and protein levels, compared with NC plasmid
(Fig. 3B). In addition, the
expression of MKK3 in 15 melanoma tissues was negatively correlated
with miR-21 expression (Fig. 3C).
Thus, these results collectively suggested that MKK3 may be a
direct target of miR-21.
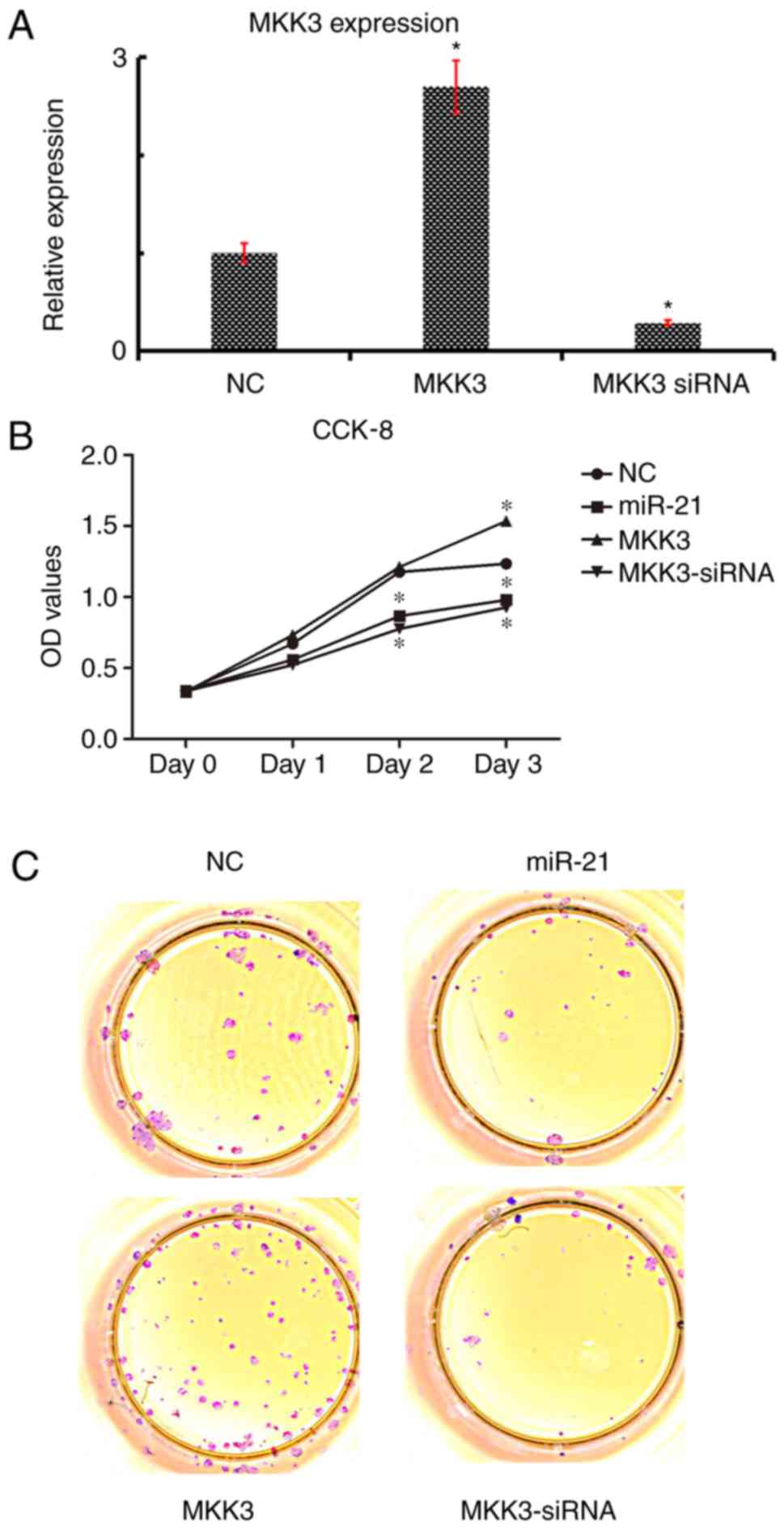
MKK3 mitigates miR-21 stimulation of
proliferation in melanoma cells
To explore the functions of MKK3 and miR-21 in A2058
cells, miR-21 was overexpressed by transfecting with miR-21 mimics,
and MKK3 was overexpressed by transfecting with an MKK3
overexpression vector. MKK3 expression was significantly increased
in A2058 cells following transfection with the MKK3 vector;
conversely, expression was significantly downregulated following
transfection with MKK3 siRNA (Fig.
4A) Initially, the roles of miR-21 and MKK3 in cell growth were
investigated; compared with the empty vector NC group, miR-21
overexpression and anti-MKK3 expression markedly inhibited the
growth of melanoma cells, as determined by CCK-8 and soft agar
assays (Fig. 4B and C).
Additionally, it was demonstrated that MKK3 overexpression
accelerated the growth of melanoma cells (Fig. 4B and C). Therefore, these data
suggested that miR-21 might restrain cell growth and MKK3 might
stimulate the cell growth of melanoma cells.
miR-21 activates melanoma cell
apoptosis and causes cell cycle arrest, but MKK3 plays an inverse
role
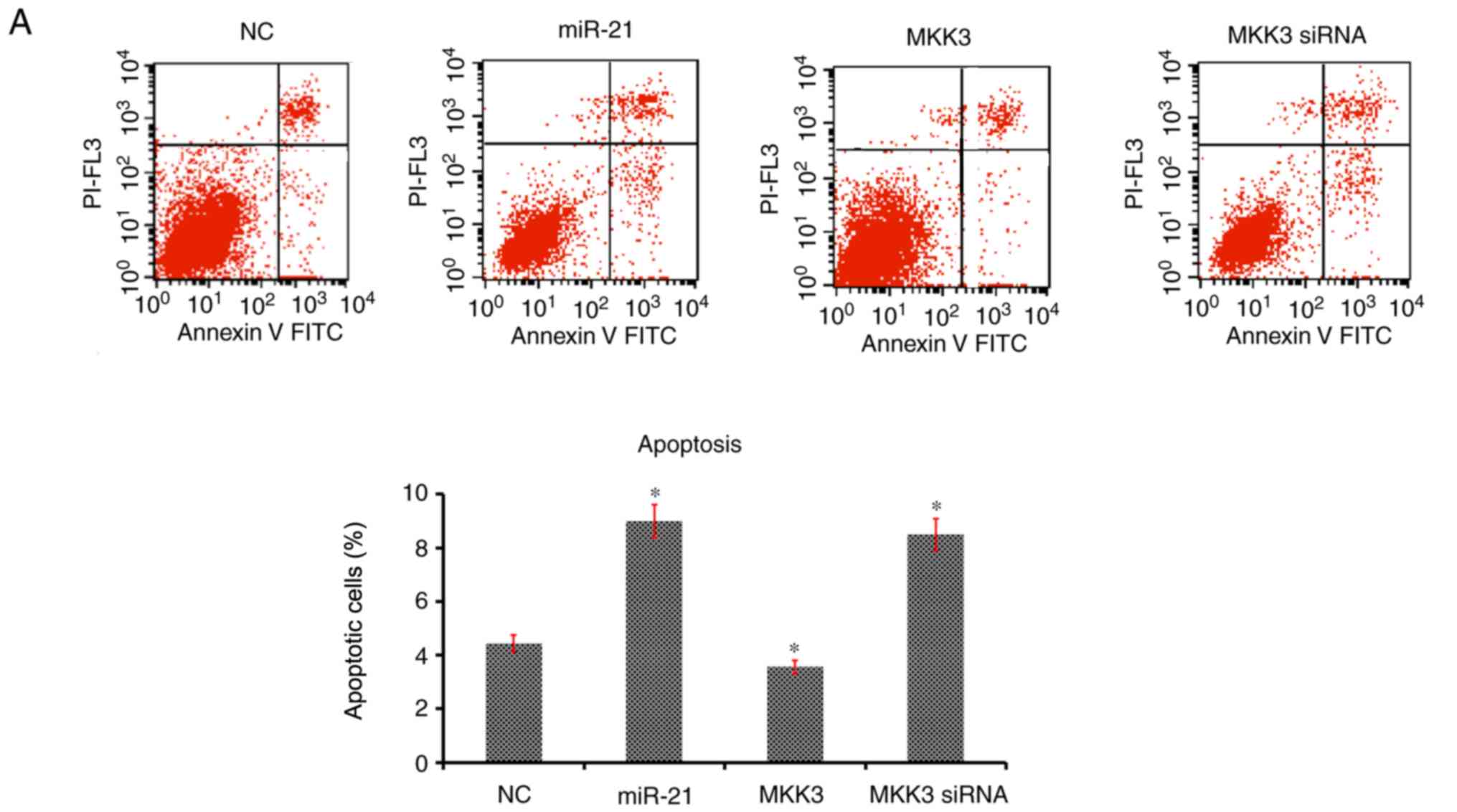
The effects of miR-21 and MKK3 on cell apoptosis and
cell cycle distribution were assessed. It was revealed that
enhanced expression of miR-21 and reduced expression of MKK3
significantly aggravated the percentage of cells undergoing
apoptosis and arrested the cell cycle at the G1 phase
compared with the NC (Fig. 5A and
B). Nevertheless, overexpression of MKK3 did not elevate the
cell apoptosis percentage or delay the cell cycle in G1
phase (Fig. 5A and B). Taken
together, these results suggested that miR-21 may stimulate
melanoma cell apoptosis and contributed to cell cycle arrest in
G1 phase, and MKK3 exerted an opposite effect on the
apoptosis and cell cycle of melanoma cells.
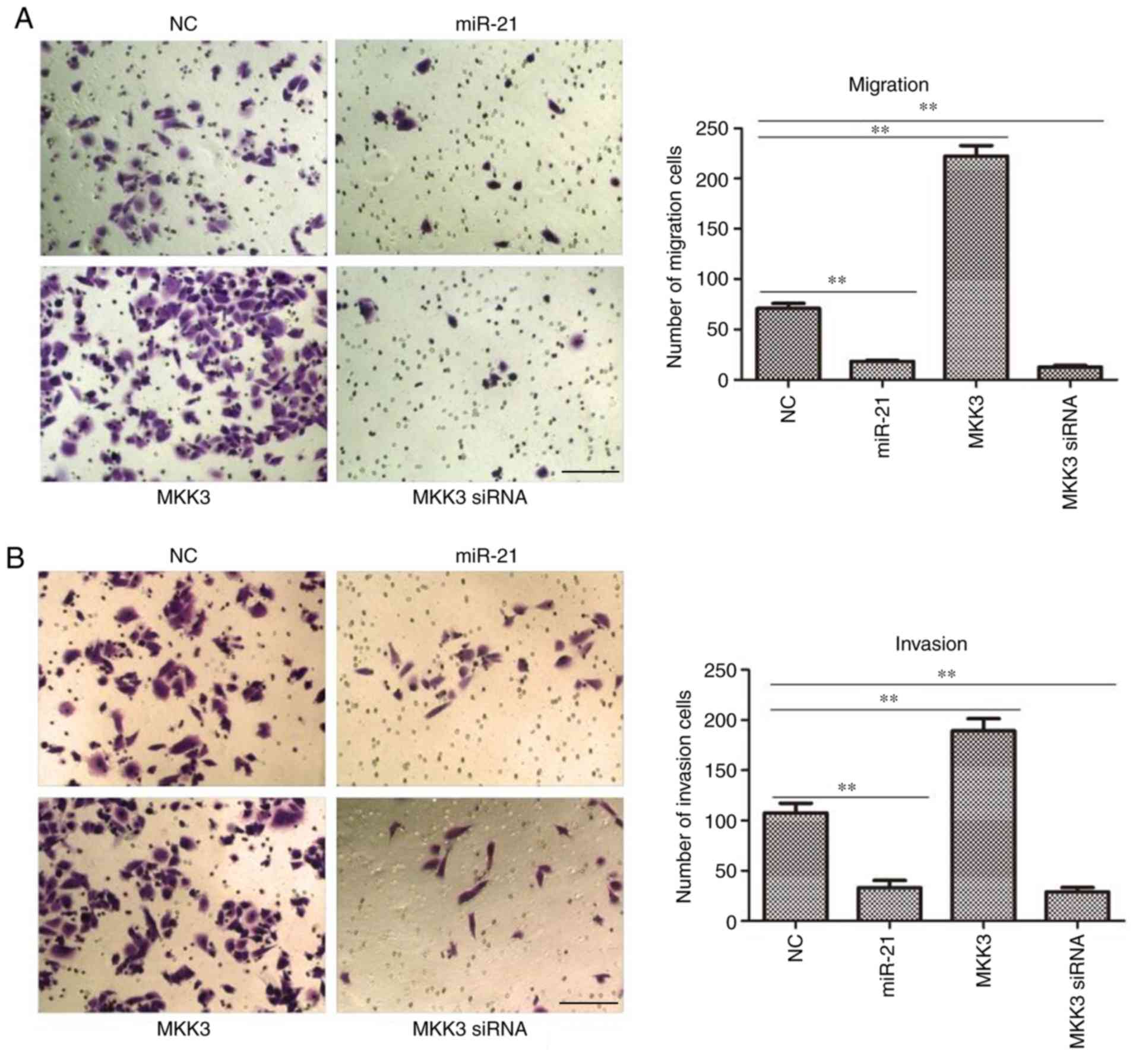
miR-21 expression suppresses, and MKK3
upregulation potentiates the migration and invasion of melanoma
cells
As it was observed that the expression of miR-21 and
MKK3 were closely associated with melanoma features, it was
hypothesized that miR-21 and MKK3 may serve an important role in
its capacity to metastasize. To test this hypothesis, a Transwell
chamber was used to quantify the changes in cell migration and
invasion. It was demonstrated that both miR-21 mimics- and
MKK3-siRNA-transfected A2058 cells exhibited significantly reduced
migratory and invasive abilities compared with NC cells (both
P<0.01; Fig. 6A and B).
Nevertheless, MKK3-overexpressing melanoma cells exhibited greater
migratory and invasive abilities than the NC cells (both P<0.01;
Fig. 6A and B). Taken together,
these in vitro findings concluded that miR-21 might inhibit
tumor cell migration and invasion, whereas MKK3 might induce tumor
cell migration and invasion.
Discussion
Melanoma is the most lethal tumor derived from
melanocytes found predominantly in the skin (2). A previous epidemiological study has
reported that the global incidence of melanoma has increased over
the past two decades (26). The
tumorigenesis and progression of melanoma is a complex and
multistep process that results from interactions between
chromosomal abnormalities, translocations, gene mutations,
epigenetic changes and environmental factors (6,27).
Patients who acquire melanoma at an early stage could be treated
with surgical management; however, patients at advanced stages
respond poorly to current treatments (28,29).
Although the understanding of the molecular mechanisms involved in
the pathogenesis of melanoma and the myriad of treatment studies
have rapidly advanced over the past two decades, effective early
diagnostic technologies and systemic therapeutic strategies of this
disease remain unsatisfactory, presenting a five-year survival rate
<15% (30). Therefore,
effective management of melanoma depends heavily on early diagnosis
and further research mainly focuses on the identification of new
high sensitivity and specificity biomarkers that are involved in
the initiation, promotion and progression of melanoma (12).
MKK3, a member of the dual specificity protein
kinase group that belongs to the MAPK kinase family, has a notable
role in the invasion and progression of gliomas and breast tumors;
it is a promising target for anticancer therapies (31). In the present study, it was
demonstrated that the mRNA and protein expression levels of MKK3
were increased in melanoma patients and cell lines. It was
additionally observed that the miR-21 expression was decreased in
these samples. A previous study suggested that miR-21, as an
oncogenic miRNA, could regulate cancer cell proliferation,
migration and apoptosis by suppressing the expression of tumor
suppressors (16). For example,
miR-21 can modulate malignant phenotypes, including proliferation,
anti-apoptosis, cell cycle progression and invasion of colorectal
cancer cells by downregulating PTEN protein expression (32). One possible explanation for this
discrepancy is that miR-21 serves its biological roles in a
tissue/developmental stage-specific context. Thus, these results
suggested that the abnormal expressions of MKK3 and miR-21 might
play an important role in the progression of melanoma.
Based on the opposing expression profiles of MKK3
and miR-21, it was further hypothesized that there was a target
relationship between MKK3 and miR-21. Subsequently, this hypothesis
was verified by a dual-luciferase reporter assay. In order to
investigate the influences of MKK3 and miR-21 on the phenotype of
melanoma cells, cell proliferation, apoptosis, the cell cycle,
migration and invasion were examined by transfecting melanoma cells
with miR-21 mimics, MKK3 mimics and MKK3-siRNA. Cell growth is
strictly regulated by cell proliferation and apoptosis, as well as
the cell cycle (33).
Dysregulation of cell proliferation, apoptosis and the cell cycle,
induced by miRNA changes, is often implicated in the occurrence and
development of cancer (34). In
the present study, upregulation of miR-21 inhibited cell growth,
and knockdown of MKK3 expression had similar effects to the miR-21
mimics group on cell growth in melanoma cells. Additionally,
accumulating evidence has suggested that metastasis is the final
stage of the deterioration process in tumor development for decades
(35,36). Furthermore, metastasis causes ~90%
of all cancer fatalities and mainly contains two vital steps,
namely migration and invasion (37). In the present study, the results
demonstrated that overexpression of miR-21 and reduced expression
of MKK3 markedly decreased the number of migratory and invasive
cells, but enhanced expression of MKK3 induced opposing effects on
the migration and invasion of melanoma cells.
In conclusion, in the present study, it was
demonstrated for the first time, to the best of our knowledge, that
MKK3 and miR-21 were abundantly expressed in patients with melanoma
and melanoma cell lines. Additionally, the abnormal expression of
these two demonstrated an inverse association, and it was further
confirmed using a dual-luciferase reporter assay that miR-21 could
directly target the 3′-UTR of MKK3. Furthermore, it was also
revealed that miR-21 could regulate cell biological behaviors,
including cell proliferation, colony formation, apoptosis, cell
cycle, cell migration and invasion, by negatively interacting with
MKK3. Collectively, these findings may provide novel insight for
melanoma diagnosis and prognosis, as well as therapeutic
strategy.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
MZ and CL conceived and designed the study,
collected the clinical data and drafted the manuscript. ZJ and XY
performed the cellular experiments. WW interpreted the data and
revised the manuscript.
Ethics approval and consent to
participate
The present study was performed in strict accordance
with the Helsinki Declaration and was approved by the Clinical
Management Committee of Hunan First People's Hospital (Changsha,
China). Written informed consent for research purposes was obtained
from all the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Owens B: Melanoma. Nature. 515
(Suppl):S1092014. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mohammadpour A, Derakhshan M, Darabi H,
Hedayat P and Momeni M: Melanoma: Where we are and where we go. J
Cell Physiol. 234:3307–3320. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Little EG and Eide MJ: Update on the
current state of melanoma incidence. Dermatol Clin. 30:355–361.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ko JS: The immunology of melanoma. Clin
Lab Med. 37:449–471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hayward NK, Wilmott JS, Waddell N,
Johansson PA, Field MA, Nones K, Patch AM, Kakavand H, Alexandrov
LB, Burke H, et al: Whole-genome landscapes of major melanoma
subtypes. Nature. 545:175–180. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong CW, Fan YS, Chan TL, Chan AS, Ho LC,
Ma TK, Yuen ST and Leung SY; Cancer Genome Project, : BRAF and NRAS
mutations are uncommon in melanomas arising in diverse internal
organs. J Clin Pathol. 58:640–644. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fuchs Wightman F, Giono LE, Fededa JP and
de la Mata M: Target RNAs strike back on MicroRNAs. Front Genet.
9:4352018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Treiber T, Treiber N and Meister G:
Regulation of microRNA biogenesis and its crosstalk with other
cellular pathways. Nat Rev Mol Cell Biol. 20:5–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mumford SL, Towler BP, Pashler AL,
Gilleard O, Martin Y and Newbury SF: Circulating MicroRNA
biomarkers in melanoma: Tools and challenges in personalised
medicine. Biomolecules. 8:E212018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mirzaei H, Gholamin S, Shahidsales S,
Sahebkar A, Jaafari MR, Mirzaei HR, Hassanian SM and Avan A:
MicroRNAs as potential diagnostic and prognostic biomarkers in
melanoma. Eur J Cancer. 53:25–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu Y, Zhang HL, Wang QY, Chen MJ and Liu
LB: Overexpression of microRNA-612 restrains the growth, invasion,
and tumorigenesis of melanoma cells by targeting espin. Mol Cells.
41:119–126. 2018.PubMed/NCBI
|
|
14
|
Zhang J, Liu WL, Zhang L, Ge R, He F, Gao
TY, Tian Q, Mu X, Chen LH, Chen W and Li X: MiR-637 suppresses
melanoma progression through directly targeting P-REX2a and
inhibiting PTEN/AKT signaling pathway. Cell Mol Biol
(Noisy-le-grand). 64:50–57. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Panza E, Ercolano G, De Cicco P, Armogida
C, Scognamiglio G, Botti G, Cirino G and Ianaro A: MicroRNA-143-3p
inhibits growth and invasiveness of melanoma cells by targeting
cyclooxygenase-2 and inversely correlates with malignant melanoma
progression. Biochem Pharmacol. 156:52–59. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pfeffer SR, Yang CH and Pfeffer LM: The
role of miR-21 in cancer. Drug Dev Res. 76:270–277. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang H, Li J, Li G and Wang S: Effects of
celastrol on enhancing apoptosis of ovarian cancer cells via the
downregulation of microRNA21 and the suppression of the
PI3K/Akt-NF-κB signaling pathway in an in vitro model of ovarian
carcinoma. Mol Med Rep. 14:5363–5368. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mu Z and Sun Q: Cantharidin inhibits
melanoma cell proliferation via the miR21mediated PTEN pathway. Mol
Med Rep. 18:4603–4610. 2018.PubMed/NCBI
|
|
19
|
Wandler A, Riber-Hansen R, Hager H,
Hamilton-Dutoit SJ, Schmidt H, Nielsen BS, Stougaard M and
Steiniche T: Quantification of microRNA-21 and microRNA-125b in
melanoma tissue. Melanoma Res. 27:417–428. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mao XH, Chen M, Wang Y, Cui PG, Liu SB and
Xu ZY: MicroRNA-21 regulates the ERK/NF-κB signaling pathway to
affect the proliferation, migration, and apoptosis of human
melanoma A375 cells by targeting SPRY1, PDCD4, and PTEN. Mol
Carcinog. 56:886–894. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang HL, Si LB, Zeng A, Long F, Qi Z,
Zhao R and Bai M: MicroRNA-21 antisense oligonucleotide improves
the sensitivity of human melanoma cells to cisplatin: An in vitro
study. J Cell Biochem. 119:3129–3141. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bossi G: MKK3 as oncotarget. Aging (Albany
NY). 8:1–2. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huth HW, Albarnaz JD, Torres AA, Bonjardim
CA and Ropert C: MEK2 controls the activation of MKK3/MKK6-p38 axis
involved in the MDA-MB-231 breast cancer cell survival: Correlation
with cyclin D1 expression. Cell Signal. 28:1283–1291. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Grimson A, Farh KK, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: Determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fattouh K, Ducroux E, Decullier E,
Kanitakis J, Morelon E, Boissonnat P, Sebbag L, Jullien D and
Euvrard S: Increasing incidence of melanoma after solid organ
transplantation: A retrospective epidemiological study. Transpl
Int. 30:1172–1180. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee JA: Current evidence about the causes
of malignant melanoma. Prog Clin Cancer. 6:151–161. 1975.PubMed/NCBI
|
|
28
|
Wikstrom JD, Lundeberg L, Frohm-Nilsson M
and Girnita A: Differences in cutaneous melanoma treatment and
patient satisfaction. PLoS One. 13:e02055172018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marcell Szasz A, Malm J, Rezeli M,
Sugihara Y, Betancourt LH, Rivas D, Gyorffy B and Marko-Varga G:
Challenging the heterogeneity of disease presentation in malignant
melanoma-impact on patient treatment. Cell Biol Toxicol. 39:1–14.
2019. View Article : Google Scholar
|
|
30
|
Rockberg J, Amelio JM, Taylor A, Jörgensen
L, Ragnhammar P and Hansson J: Epidemiology of cutaneous melanoma
in Sweden-Stage-specific survival and rate of recurrence. Int J
Cancer. 139:2722–2729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Baldari S, Ubertini V, Garufi A, D'Orazi G
and Bossi G: Targeting MKK3 as a novel anticancer strategy:
Molecular mechanisms and therapeutical implications. Cell Death
Dis. 6:e16212015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu Y, Song Y, Xiong Y, Wang X, Xu K, Han
B, Bai Y, Li L, Zhang Y and Zhou L: MicroRNA-21 (Mir-21) promotes
cell growth and invasion by repressing tumor suppressor PTEN in
colorectal cancer. Cell Physiol Biochem. 43:945–958. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lynch MP, Nawaz S and Gerschenson LE:
Evidence for soluble factors regulating cell death and cell
proliferation in primary cultures of rabbit endometrial cells grown
on collagen. Proc Natl Acad Sci USA. 83:4784–4788. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Broussard L, Howland A, Ryu S, Song K,
Norris D, Armstrong CA and Song PI: Melanoma cell death mechanisms.
Chonnam Med J. 54:135–142. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Miller FR: Immune mechanisms in the
sequential steps of metastasis. Crit Rev Oncog. 4:293–311.
1993.PubMed/NCBI
|
|
36
|
Scully OJ, Bay BH, Yip G and Yu Y: Breast
cancer metastasis. Cancer Genomics Proteomics. 9:311–320.
2012.PubMed/NCBI
|
|
37
|
Wu J, Zhang Y, Cheng R, Gong W, Ding T,
Zhai Q, Wang Y, Meng B and Sun B: Expression of
epithelial-mesenchymal transition regulatorsTWIST, SLUG and SNAIL
in follicular thyroid tumours may relate to widely invasive, poorly
differentiated and distant metastasis. Histopathology. 74:780–791.
2019. View Article : Google Scholar : PubMed/NCBI
|