Introduction
Neural stem/progenitor cells (NSPCs) are
self-renewing, multipotent cells that generate the neurons,
astrocytes and oligodendrocytes of the nervous system. They are
present not only in developing, but also in the adult central
nervous system (CNS) (1).
Neurogenesis occurs mainly in two areas: The subgranular zone of
the dentate gyrus and the anterior part of the subventricular zone
along the ventricle (2). NSPCs can
be activated in response to brain injury, contributing towards the
repair and motor functional recovery of the CNS (3). A number of factors regulate the
maintenance, activation, and fate choice of NSPCs, including
extracellular signaling molecules [basic fibroblast growth factor
(bFGF), epidermal growth factor (EGF), glutamate], intracellular
signal molecules (GLI family zinc finger 1, AKT, Notch1) and
microRNAs (miRNAs/miRs) (4).
miRNAs are small noncoding RNA molecules (~22
nucleotides) present in plants, animals and certain viruses, which
contribute to the post-transcriptional regulation of gene
expression by binding and silencing target RNAs, and thus regulate
cell behavior (5). Among them,
miR-29 is notable due to its regulatory role in cell proliferation,
differentiation and survival, not only in cancer cells but also in
stem/progenitors cells (6,7). Previous studies have indicated that
miR-29 may serve an important role in regulating the behavior of
stem cells (8,9); however, the effects of miR-29 on
NSPCs and the underlying intracellular signaling mechanisms remain
unknown.
In the present study, the effects of miR-29 on
proliferation in NSPCs were investigated. It was demonstrated that
overexpression of miR-29 promoted NSPC proliferation and inhibited
the expression of phosphatase and tensin homologue deleted on
chromosome 10 (PTEN) protein, potentially via a mechanism involving
the AKT signaling pathway. This research may provide novel insight
for the development of strategies for stem cell-mediated treatment
of CNS diseases.
Materials and methods
Rat NSPCs culture
Rat NSPCs were prepared from E14.5 Sprague-Dawley
rat embryos as previously described, with minor modifications
(10). A total of six pregnant
rats were used for this study, and five or six embryos obtained
from each pregnant rat were used for NSPCs culture. For NSPC
culture, the serum-free complete medium consisted of DMEM/F12
(1:1), 1% N-2 (cat. no. 17502048), 2% B27 (cat. no. 17504044), 20
ng/ml EGF and 10 ng/ml Bfgf (10).
HepG2 cells, obtained from ATCC (HB-8065), were cultured in DMEM
medium (Hyclone; GE Healthcare Life Sciences), supplemented with
10% fetal bovine serum (Invitrogen; Thermo Fisher Scientific,
Inc.). All the cells were incubated at 37°C and 5% CO2
in a humidity incubator (Sanyo Electric Co., Ltd.). After three to
five days, cells that multiplied in primary neurospheres (P0 cells)
with the a diameter of approximately 90–150 µm were dissociated
into single cells by trypsin (0.05%, Invitrogen; Thermo Fisher
Scientific, Inc.). All materials used for cell culture were
purchased from Thermo Fisher Scientific, Inc. For single-cell
adhesive culture, single NSPCs in serum-free complete medium were
allowed to attach onto poly-D-lysine-coated coverslips. For NSPCs
differentiation, single cells were cultured in differentiation
medium, which removing mitogens (bFGF and EGF) and contained 1%
fetal bovine serum. Following culture for 3 days, cell were fixed
by 4% paraformaldehyde at room temperature for 30 min and used for
immunostaining test.
Pregnant SD rats were purchased from the
Experimental Animal Center of Xi'an Jiaotong University Health
Science Center (certificate no. 22-9601018). All experimental
protocols were approved by the Animal Care and Use Regulation of
Xi'an Jiaotong University Health Science Center. All efforts were
made to minimize animals' suffering and reduce the number of
animals used to a minimum.
Transfection
Cells were plated in poly-D-lysine-coated 24-well or
6-well plates. Cells were 70–90% confluent at the time of
transfection, and transient transfections were performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols. The
miR-NC, miR-29 mimic and miR-29 inhibitor were purchased from
Shanghai GenePharma Co., Ltd. The target sequences were as follows:
miR-29 mimic: 5′-ACUGAUUUCUUUUGGUGUUCAG-3′; miR-29 inhibitor:
5′-ACTGATTTCAAATGGTGCT-3′; miR-NC: 5′-UUCUCCGAACGUGUCACGUTT-3′.
Before transfection, the culture medium was washed three times by
Opti-MEM (Invitrogen; Thermo Fisher Scientific, Inc.). Cells were
transfected with 100 nM microRNA for 6 h in the presence of
Lipofectamine® 2000 according to the manufacturer's
instructions (Invitrogen; Thermo Fisher Scientific, Inc.). Control
treatment involved addition of an equivalent volume of medium
without miRNA. Knockdown of mGluR4 expression was further evaluated
using reverse transcription (RT) PCR. Cells were cultured for 24
more hours before further treatments.
RT-quantitative PCR (RT-qPCR)
After transfection for 24 h, total RNA was isolated
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. RT was performed
using TaqMan® MicroRNA Reverse Transcription kit and
specific primers (Applied Biosystems; Thermo Fisher Scientific,
Inc.). All RT reactions were carried out in triplicates in
Mastercycler ep gradient S (Eppendorf). RT was carried out in an MJ
Research Thermal Cycler (Bio-Rad Laboratories) at 16°C for 30 min,
42°C for 30 min, and 85°C for 5 min. qPCR was performed using
single tube TaqMan® MicroRNA Assays and
TaqMan® 2X Universal polymerase chain reaction PCR
Master Mix, No AmpErase® UNG (Applied Biosystems; Thermo
Fisher Scientific, Inc.) in 10 µl reactions. All qPCR reactions
were performed in triple in iQ5 Real-Time PCR Detection System
(Bio-Rad Laboratories, Inc.). According to the manufacturer's
protocol. The primer pairs were synthesized by Takara Biotechnology
Co., Ltd. and were as follows: miR-29-forward:
5′-AGTGAATGAGGCCTTCGAGA-3′; miR-29-reverse:
5′-GCATCTGAGTCGCCACTGTA-3′; U6-forward: 5′-GCATCTGAGTCGCCACTGTA-3′;
U6-reverse: 5′-CGCTTCACGAATTTGCGTGTCAT-3′. The PCR cycles were
performed by initial denaturation at 95°C for 5 min, then 40 cycles
at 95°C for 10 sec followed by 60°C for 1 min. Data were analyzed
using the 2−ΔΔCq method (11).
Cell viability assay
Cell viability was evaluated as previously described
(12) using a Cell Counting Kit-8
(Roche Applied Science) according to the manufacturer's protocols.
For the miR-29 group, NSPCs were grown in 24-well plates, the other
groups were grown in 96-well plates at 5,000 cells/well for 24 h
before transfection. Cell viability was detected at the end of each
treatment (6, 12, 24, 48 and 72 h following transfection) using an
Epoch multi-microplate spectrophotometer (Bio-Tek Instruments,
Inc.).
Cell culture 5-bromo-2-deoxyuridine
(BrdU) incorporation assay
For the cell proliferation assay, 10 µM BrdU
(Sigma-Aldrich; Merck KGaA) solutions were added to NSPCs
(1×105 cells) plated on coverslips for 2 h. The cells
were then fixed with 4% paraformaldehyde for 30 min at room
temperature. The BrdU-labeled cells were visualized via
immunostaining as described below, and stained using propidium
iodide (10 µg/ml, Sigma-Aldrich; Merck KGaA) at 37°C for 30 min.
Immunoreactive cells were observed using a BX51 fluorescent
microscope equipped with a DP70 digital camera (Olympus
Corporation). Cells from 5 randomly selected fields on each
coverslip were counted at ×200 magnification. The experiments were
performed in triplicates and repeated independently at least three
times. The percentage of labeled cells was evaluated and normalized
by the PI staining nuclei. All of the counts were conducted on
Image-Pro Plus software (version 5.0; Media Cybernetics, Inc.).
Immunostaining
Immunostaining was performed as previously described
(13). Briefly, NSCs were fixed in
4% PFA for 30 min at room temperature and washed twice with PBS, at
the end of treatments. Cells were permeabilized in 0.3% Triton
X-100 and blocked for 1 h in 10% normal donkey serum. Then the
cells were incubated overnight using the primary antibodies at
appropriate dilution (Table I).
After thorough rinsing, the cells were detected for 2 h at room
temperature by appropriate Alexa Fluor 488 or 594 conjugated
secondary antibodies (Invitrogen; Thermo Fisher Scientific, Inc.).
Nuclei were counterstained using DAPI (1 µg/ml). Fluorescence
images were observed using a BX51 fluorescent microscope equipped
with a DP70 digital camera. For quantification, images were
imported into Image-Pro Plus software. Cells from 5 randomly
selected fields on each coverslip were counted at ×400
magnification. The experiments were performed in triplicates and
repeated independently at least three times.
 | Table I.Antibodies used during the study. |
Table I.
Antibodies used during the study.
| Antigen | Supplier | Antibody type | Catalogue
number | Dilution |
|---|
| Nestin | Merck KGaA | Mouse
monoclonal | MAB353 | 1:200 in PBS
(IF) |
| Tuj1 | Merck KGaA | Mouse
monoclonal | MAB1637 | 1:200 in PBS
(IF) |
| GFAP | Abcam | Rabbit
monoclonal | ab33922 | 1:200 in PBS
(IF) |
| BrdU | Abcam | Sheep
polyclonal | ab1893 | 1:200 in PBS
(IF) |
| Anti-Mouse IgG
(H+L) secondary | Donkey
polyclonal | Invitrogen | R37114 | 1:500 in PBS
(IF) |
| Antibody, Alexa
Fluor 488 | antibody |
|
|
|
| Anti-Rabbit IgG
(H+L) secondary | Donkey
polyclonal | Invitrogen | R37119 | 1:500 in PBS
(IF) |
| Antibody, Alexa
Fluor 594 | antibody |
|
|
|
| Anti-Sheep IgG
(H+L) Cross-Adsorbed | Donkey
polyclonal | Invitrogen | A-11016 | 1:500 in PBS
(IF) |
| Secondary
Antibody | antibody |
|
|
|
| PTEN | Cell Signaling | Rabbit
polyclonal | 9188 | 1,1000 in TBST
(WB) |
|
| Technology,
Inc. |
|
|
|
| p-AKT | Cell Signaling | Rabbit
polyclonal | 9271 | 1,1000 in TBST
(WB) |
|
| Technology,
Inc. |
|
|
|
| AKT | Cell Signaling | Rabbit
polyclonal | 9272 | 1,1000 in TBST
(WB) |
|
| Technology,
Inc. |
|
|
|
| β-actin | Sigma-Aldrich | Mouse
polyclonal | A1978 | 1,1000 in TBST
(WB) |
|
| (Merck KGaA) |
|
|
|
Cell cycle analysis
The cell cycle was analyzed using fluorescence
activated cell sorting as previously described (13). Briefly, NSPCs were fixed using
ice-cold 75% ethanol overnight at 4°C. After washing in PBS, the
cells were stained with 100 µg/ml Propidium iodide containing 100
µg/ml RNase A for 30 min at 37°C in dark. Samples were analyzed
using a flow cytometer (FACSCalibur; BD Biosciences), and DNA
contents and cell cycle distribution were determined using Modfit
LT software (version 3.2, BD Biosciences). The proliferation index
(PI) used to evaluate differences in the cell cycle distribution
was calculated as follows: PI=(S +
G2/M)/[G0/G1 + (S +
G2/M)].
Western blot analysis
Western blotting was performed as previously
described (13). Briefly, NSPCs
were grown in 6-well plates at 2×105 cells/well for 24
hours prior to the experiments. After transfection, NSPCs protein
were collected at the end of each treatment (5, 15, 30, 60 and 120
h). The effects of miR-29 on NSPCs were further confirmed by
blockade of PTEN using antagonist VO-OHpic trihydrate (VU-OH, 5 µM,
Abcam). Before transfection 24 h, 2 µM VO-OH were added in the
anti-miR-29 group medium and still contained VO-OH in the medium
after transfection. Because VO-OH were dissolved in DMSO, the same
volume of DMSO were added in the control group as negative group.
After 30 h, NSPCs protein were collected and used for western blot.
Protein concentration was determined using the BCA method (Pierce)
and lysates (20–40 µg/sample) were subjected to 12% SDS-PAGE, and
transferred onto PVDF membranes. Membranes were blocked for 1 h in
5% non-fat dry milk in Tris-HCl buffer followed by incubated in
primary antibodies (Table I)
overnight at 4°C. Membranes were then incubated in HRP-conjugated
secondary antibody for 1 h at room temperature. Following a
thorough rinse, Immunoreactive bands were detected using an
enhanced chemiluminescent (ECL) substrate (Pierce; Thermo Fisher
Scientific, Inc.) and exposure to a Fuji X-ray film (Fujian Gutian
Yuanhang Medical Co., Ltd.). Bands were imaged using a G:Box gel
imaging system (Syngene), and protein expression quantified using
ImageJ 3.5 software (National Institutes of Health).
Statistical analysis
Data were presented as the mean ± standard deviation
from at least three independent in vitro experiments.
Differences between groups were analyzed using one-way ANOVA,
followed by Tukey's post hoc test. All statistical analyses were
performed using SPSS version 12.0 software (SPSS, Inc.). P<0.05
was considered to indicate a statistically significant
difference.
Results
Characterization of NSPCs derived from
fetal rat cortex
Primary NSPCs were cultured as aforementioned. After
2–3 days, NSPCs proliferated and formed secondary neurospheres
(Fig. 1A), and were characterized
by expression of nestin (a marker of NSPCs). Nestin-positive
immunoreactivity was observed in 88.02±4.39% of cells in the
neurospheres (Fig. 1B). To
identify the neural differentiating potential, the NSPCs were
dissociated into single cells by trypsin, plated onto
poly-D-lysine-coated coverslips and cultured in differentiating
medium (DMEM/F12 supplemented with 1% N2, 2% B27 and 1% FBS,
lacking bFGF and EGF). Immunostaining revealed that neuron-specific
class III β-tubulin (Tuj1; a marker of immature neurons) and glial
fibrillary acidic protein (a marker of astrocytes) immunoreactivity
was observed in cultured cells (Fig.
1C and D).
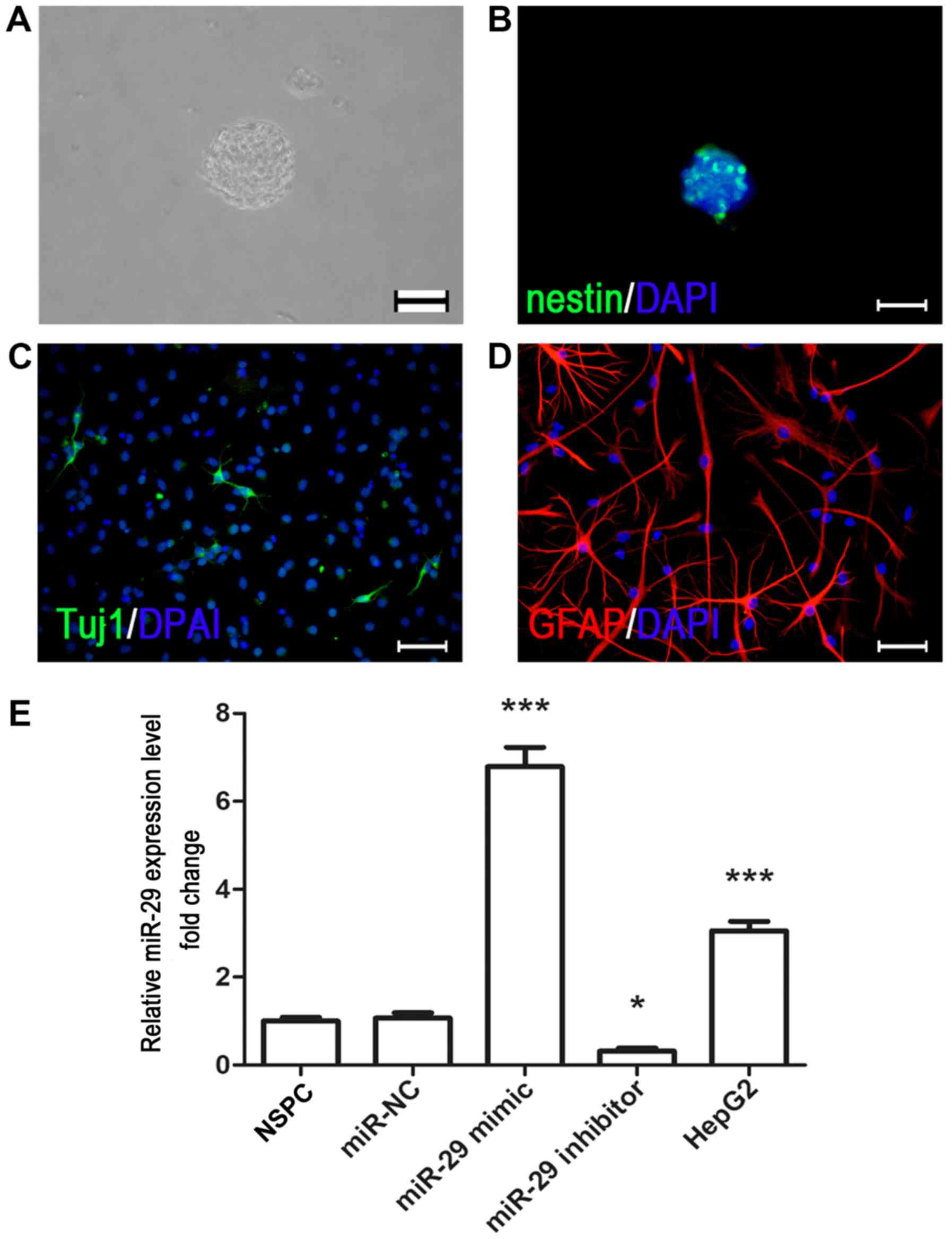 | Figure 1.Culture and immunocytochemical
characterization of fetal rat cortex NSPCs. (A) NSPCs were isolated
from the E14.5 fetal rat cortex, and neurospheres were formed
following 3–5 days of cell proliferation. Scale bar, 200 µm. (B)
Nestin (green) immunostaining of NSPCs was conducted in
neurospheres under proliferating conditions. Scale bar, 200 µm.
NSPCs were differentiated into (C) Tuj1- and (D)
GFAP-immunopositive cells by removal mitogens and adding 1% fetal
bovine serum. Scale bar, 40 µm. Nuclei were counterstained with
DAPI (blue). (E) Reverse transcription-quantitative PCR analysis
indicated that miR-29 mimic increased the expression of miR-29,
whereas miR-29 inhibitor downregulated its expression. The liver
cancer cell line HepG2 was used as a positive control. *P<0.05,
***P<0.001 vs. miR-NC. NSPC, neural stem/progenitor cell;
miR-29, microRNA-29; miR-NC, microRNA negative control; Tuj1,
neuron-specific class III β-tubulin; GFAP, glial fibrillary acidic
protein. |
miR-29 promotes the viability of
NSPCs
To investigate the effects of miR-29 on NSPCs, cells
were transfected with miR-NC, miR mimic or inhibitor. It was
revealed that transfection with miR mimic significantly upregulated
the expression of miR-29 compared with miR-NC, whereas miR-29
inhibitor significantly downregulated expression (Fig. 1E). Then, CCK-8 assays were
conducted at various timepoints following transfection of NSPCs
with miR-NC, miR-29 mimic or miR-29 inhibitor. It was revealed that
cell viability was significantly increased at 24 h following
transfection with miR-29 compared with at 0 h, whereas the
viability of control and miR-NC-transfected NPSCs was only
significantly increased compared with 0 h at the 48-h point
(Fig. 2A-C). Conversely, the
viability of miR-29 inhibitor-transfected cells did not
significantly alter across the 72-h assay period (Fig. 2D). The results suggested that
overexpression of miR-29 promoted the viability of NSPCs.
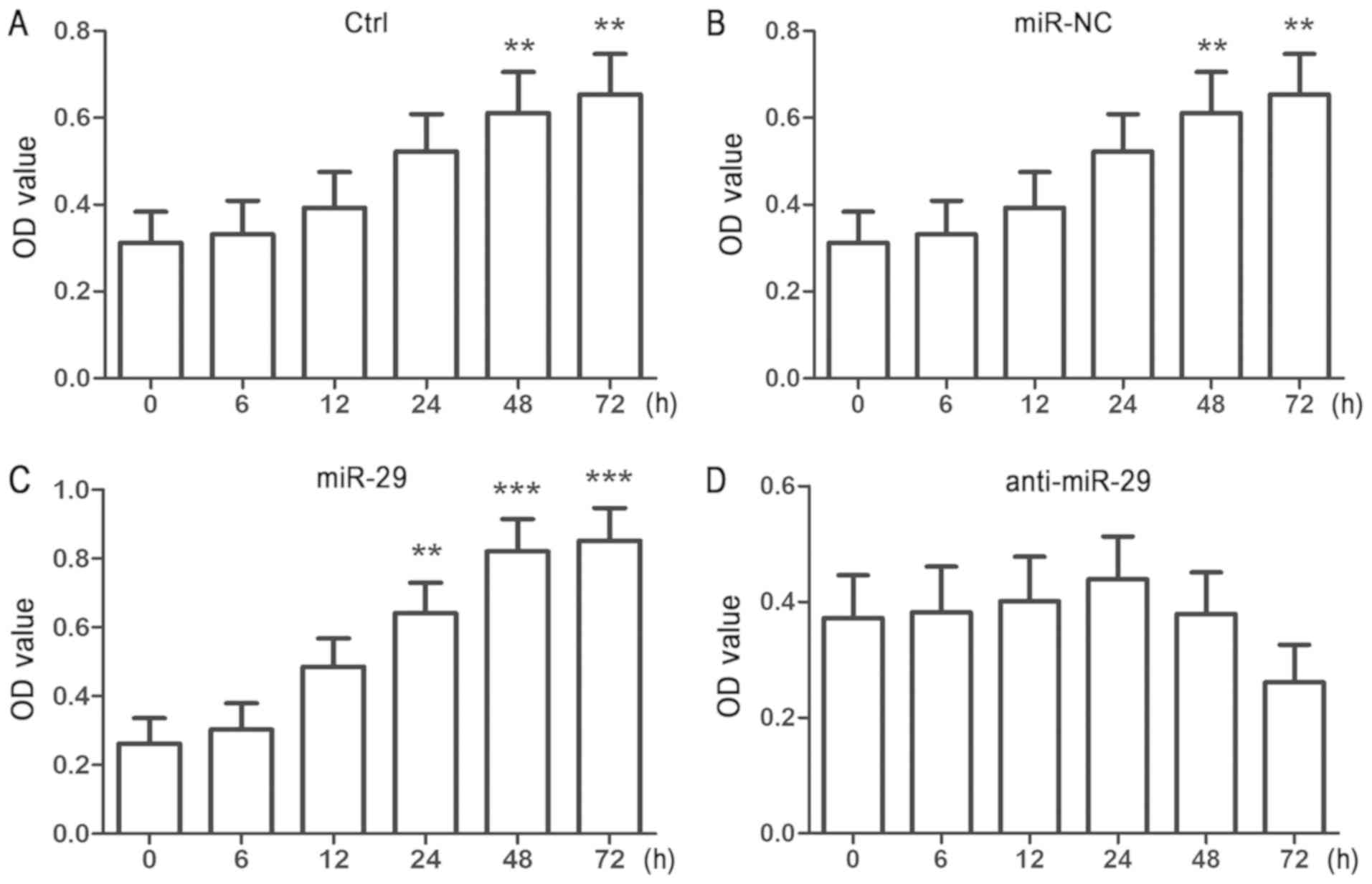 | Figure 2.Overexpression of miR-29 increases the
viability of cultured NSPCs. Single adhesive cultured NSPCs were
transfected for 6 h with (A) Ctrl, (B) miR-NC, (C) miR-29 mimic or
(D) anti-miR-29 using Lipofectamine 2000. Cell viability was
detected by a Cell Counting Kit-8 assay at the end of each time
point (6, 12, 24 and 72 h). Data are presented as the mean ±
standard deviation of three independent experiments. **P<0.01,
***P<0.001 vs. 0 h. NSPC, neural stem/progenitor cell; miR-29,
microRNA-29; anti-miR-29, miR-29 inhibitor; miR-NC, microRNA
negative control; Ctrl, miR-free medium; OD, optical density. |
miR-29 promotes the expression of
PTEN
PTEN protein catalyzes the dephosphorylation of the
3′phosphate of the inositol ring of phosphatidylinositol (3,4,5)-trisphosphate (PIP3),
converting it to phosphatidylinositol (4,5)-bisphosphate (PIP2)
(14). A previous study reported
that PTEN has a role in regulating NSPC behavior (15). To investigate whether miR-29
influenced the expression of PTEN, NSPCs were transfected with
miR-29 mimic or miR-29 inhibitor. Following treatment for various
durations (5, 15, 30, 60 and 120 h), total protein was collected
and PTEN expression was analyzed via western blotting. The results
indicated that the overexpression of miR-29 significantly
downregulated the expression of PTEN in a time-dependent manner
(Fig. 3A). Of note, knockdown of
miR-29 induced opposing effects on the expression of PTEN (Fig. 3B). To further investigate the
effects of miR-29 on PTEN, VO-OHpic trihydrate (VO-OH; a potent,
selective and reversible PTEN inhibitor) was added to the culture
medium. NSPCs were transfected with miR-NC, miR-29 mimic or miR-29
inhibitor; miR-29 inhibitor-treated cells were pretreated with 2 µM
VO-OH. As VO-OH was dissolved in DMSO, the same volume of DMSO
(0.5%) was added to the control group as a negative control group.
There were no significant differences observed between the control,
DMSO and miR-NC groups, indicating that DMSO and miR-NC did not
influence the expression of PTEN. miR-29 mimic-transfected cells
exhibited significantly downregulated expression of PTEN, whereas
miR-29 inhibitor upregulated PTEN expression. Of note, treatment
with VO-OH significantly eliminated the effects of anti-miR-29 on
PTEN levels (Fig. 3C). It is
hypothesized that a possible reason for PTEN suppression is that
VO-OH (a highly potent inhibitor of PTEN) could inhibit the
activation of AKT pathway, which may feedback to PTEN and suppress
the PTEN expression. The results indicated that miR-29 regulated
the expression of PTEN in NSPCs, and that overexpression of miR-29
inhibited the expression of PTEN.
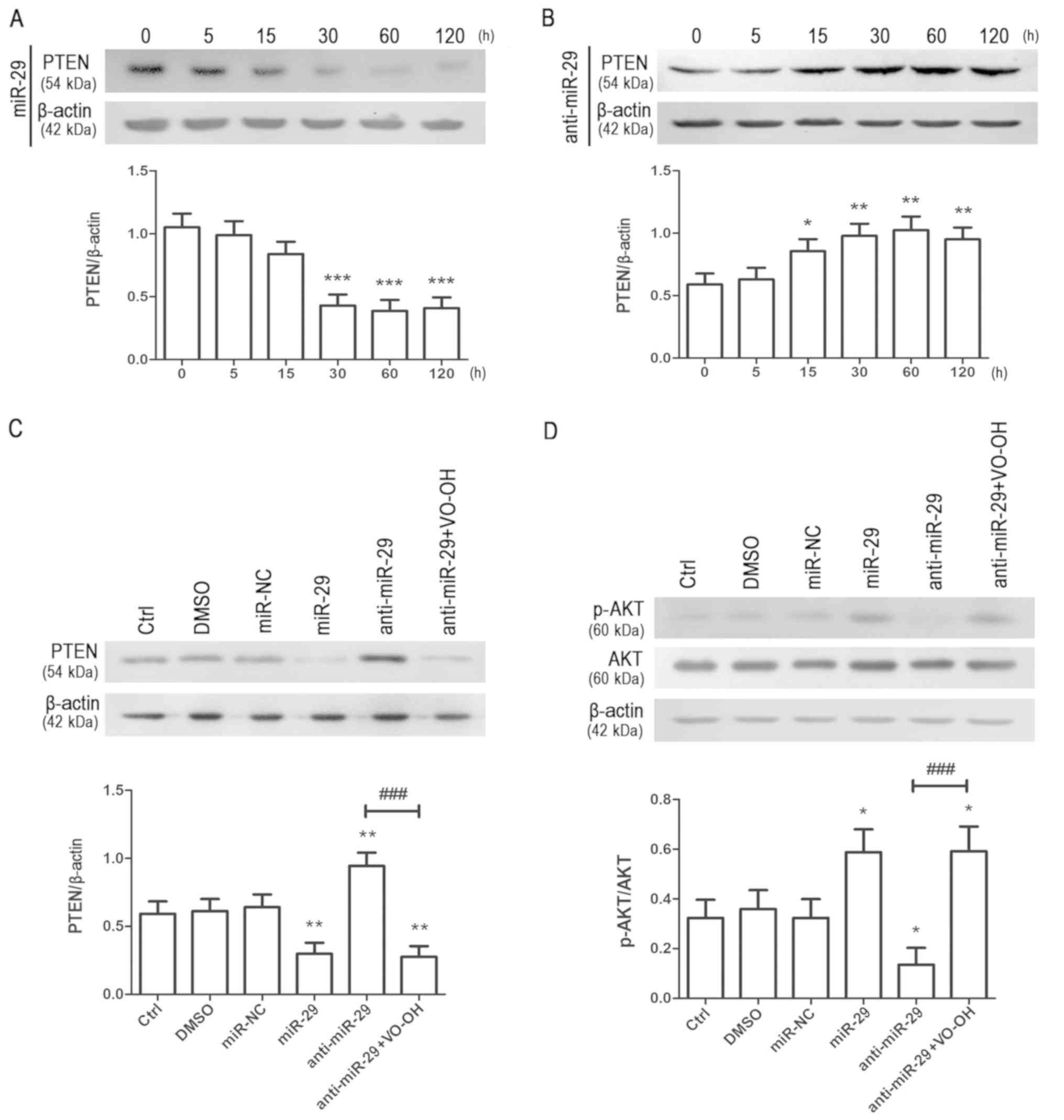 | Figure 3.miR-29 regulates the PTEN/AKT
signaling pathway in NPSCs. Single adhesive cultured NSPCs were
transfected with (A) miR-29 mimic or (B) miR-29 inhibitor using
Lipofectamine 2000. After transfection, NSPCs protein were
collected at the end of each treatment and detected by western blot
analysis after each time point (5, 15, 30, 60, 120 h). Data are
presented as the mean ± standard deviation of three independent
experiments. *P<0.05, **P<0.01, ***P<0.001 vs. 0 h. (C) To
further investigate the effects of miR-29 on PTEN, the
PTEN-specific inhibitor VO-OH was added to the medium before
transfection and the medium after transfection still contained
VO-OH. PTEN expression was quantified and normalized to β-actin.
Data are presented as the mean ± standard deviation of three
independent experiments. **P<0.01 vs. DMSO;
###P<0.001 vs. anti-miR-29. (D) Expression of p-AKT
and AKT as determined by western blot analysis; p-AKT expression
was normalized to total AKT expression. Data are presented as the
mean ± standard deviation of three independent experiments.
*P<0.05 vs. DMSO, ###P<0.001 vs. anti-miR-29.
NSPC, neural stem/progenitor cell; miR-29, microRNA-29;
anti-miR-29, miR-29 inhibitor; miR-NC, microRNA negative control;
Ctrl, treatment with miRNA-free medium; DMSO, dimethyl sulfoxide;
PTEN, phosphatase and tensin homologue deleted on chromosome 10;
VO-OH, VO-OHpic trihydrate; p-, phosphorylated. |
Additionally, the activation of AKT (a kinase
downstream of PTEN) was analyzed via western blotting.
Overexpression of miR-29 promoted the phosphorylation of AKT,
whereas knockdown of miR-29 induced opposing effects (Fig. 3D). Furthermore, VO-OH significantly
attenuated the effects of miR-29 inhibitor on AKT activation. The
findings indicated that miR-29 affected the activation of the AKT
signaling pathway.
miR-29 promotes the proliferation of
NSPCs by regulating PTEN
To investigate whether the effects of miR-29 on the
proliferation of cells were mediated by PTEN, BrdU and cell cycle
assays were conducted in the presence of VO-OH. NSPCs were
transfected as described. BrdU staining analysis revealed that both
DMSO and miR-NC did not significantly affect the proliferation of
NSPCs compared with the control (Fig.
4A and B). Conversely, overexpression of miR-29 significantly
increased the number of BrdU-positive cells compared with the
control, whereas knockdown of miR-29 induced opposing effects;
however, treatment with VO-OH significantly eliminated the effects
of miR-29 inhibitor on the number of BrdU-positive cells (Fig. 4A and B). The results of the cell
cycle assay were similar to those from the BrdU assay.
Overexpression of miR-29 significantly increased the PI of NSPCs
compared with the control, whereas knockdown of miR-29 decreased
the PI, an effect that was significantly reversed by treatment with
VO-OH (Fig. 4C). Collectively,
these results suggested that miR-29 promoted the proliferation of
NSPCs in a PTEN-dependent manner.
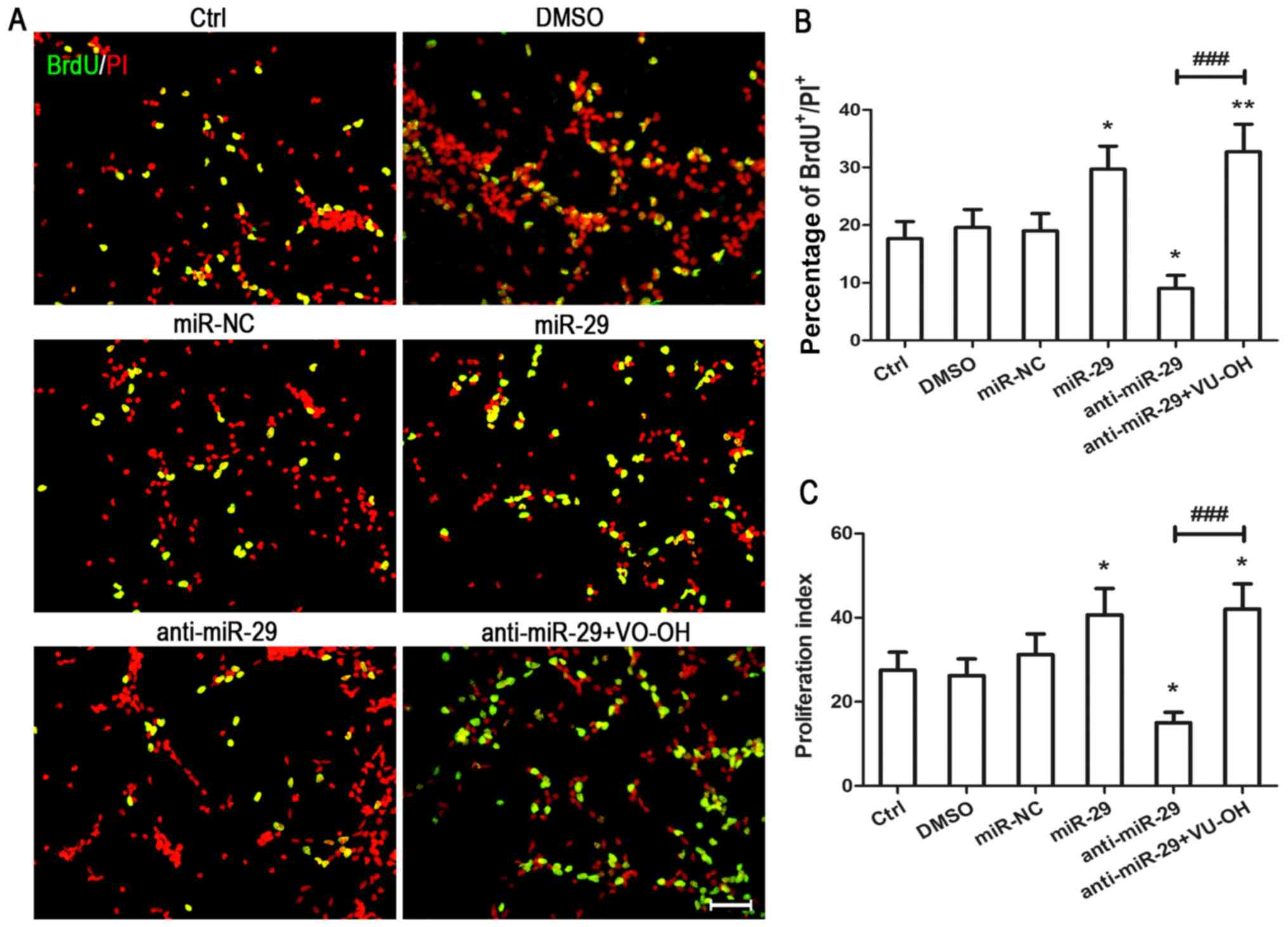 | Figure 4.Overexpression of miR-29 promotes the
proliferation of NSPCs. Single adhesive cultured NSPCs were
transfected for 6 h with miR-NC, miR-29 mimic or miR-29 inhibitor
using Lipofectamine 2000. In addition, the PTEN-specific inhibitor
VO-OH was used to treat anti-miR-29 cells. (A) Proliferating cells
were detected by BrdU incorporation and immunostaining.
BrdU-labeled cells, green; propidium iodide, red. Scale bars=200
µm. (B) Data from three independent experiments (n=3) are presented
as the percentage of BrdU-positive cells in the total population of
propidium iodide-positive cells. *P<0.05, **P<0.01 vs. DMSO;
###P<0.001 vs. anti-miR-29. (C) Proliferation index
of transfected cells as determined by cell cycle analysis.
Proliferation index=(S +
G2/M)/[G0/G1 +
(S+G2/M)]. Data are presented as the mean ± standard
deviation of three independent experiments. *P<0.05 vs. DMSO
group; ###P<0.001 vs. anti-miR-29. NSPC, neural
stem/progenitor cell; miR-29, microRNA-29; anti-miR-29, miR-29
inhibitor; miR-NC, microRNA negative control; Ctrl, treatment with
miRNA-free medium; DMSO, dimethyl sulfoxide; PTEN, phosphatase and
tensin homologue deleted on chromosome 10; VO-OH, VO-OHpic
trihydrate; p-, phosphorylated. |
Discussion
PTEN protein is one of the most frequently mutated
tumor suppressor genes in human cancer; however, previous studies
reported that PTEN serves important roles not only in cancer cells,
but also in NSPCs (15,16). Such findings suggest that PTEN
could be potential novel treatments to promote CNS repair. A number
of factors regulate the intracellular expression of PTEN, including
miR-29; previous studies demonstrated that miR-29 significantly
downregulated the expression of PTEN in various types of cell
(17–19). However, the effects of miR-29 on
PTEN in NSPCs remain unclear. In the present study, it was
demonstrated that overexpression of miR-29 decreased PTEN
expression in a time-dependent manner, and that PTEN inhibitor
eliminated the effects of miR-29 inhibitor. Furthermore, the
results indicated that miR-29 promoted NPSC proliferation by
inhibiting the expression of PTEN. These findings suggested that
miR-29 may be a molecular target for regulating the proliferation
of NSPCs.
It was previously demonstrated that PTEN protein
exhibits at least two biochemical functions: It has both lipid
phosphatase and protein phosphatase activity (20). As a phosphatase, PTEN protein
specifically catalyzes the dephosphorylation of the 3′phosphate on
the inositol ring of PIP3, converting it to
PIP2. This dephosphorylation leads to the inhibition of
the AKT signaling pathway (21).
AKT is an important intercellular signal molecule involved in the
regulation of the proliferation, differentiation and survival of
numerous types of cell, including NSPCs (22,23).
Activation of AKT promoted neurogenesis and the proliferation of
retinal progenitor cells in vitro and in vivo
(13,23). In the present study, it was
revealed that overexpression of miR-29 increased the
phosphorylation levels of AKT. Furthermore, PTEN arrests cell cycle
progression at the G1/S phase, mediated at least
partially via the upregulation of the cyclin-dependent kinase
inhibitor p27 (24). As such, it
is proposed that miR-29 may promote the proliferation of NSPCs by
suppressing the expression of PTEN protein and thereby promoting
the activation of AKT. For protein phosphatase activity, PTEN does
not only affect the PI-3-K signaling pathway; it is involved in the
persistent enhancement of neural stem cell self-renewal without
exhaustion, and the inhibition of the growth factor-stimulated
mitogen-activated protein kinase (MAPK) signaling pathway (25,26).
The MAPK and AKT signaling pathways contribute to the proliferation
of NSPCs, and activation of MAPK signaling pathway may promote cell
proliferation (27). Intercellular
signaling pathways form an intersecting biochemical network that,
once activated, can result in a series of biological effects,
including proliferation, apoptosis and differentiation (28). Cross-talk exists between the kinase
cascades, in which the inhibition of one kinase cascade may lead to
the activation of another, and vice versa (29). Therefore, whether other signal
pathways are involved in the effects of miR-29 on NPSC
proliferation requires further investigation.
miR-29 was one of the first miRNAs detected in the
human genome, and is upregulated in all types of human malignancy;
overexpression of miR-29 may promote proliferation and invasion,
and reduce cell apoptosis (30).
This previous study showed miR-29 family members share a common
seed region sequence and are predicted to target largely
overlapping sets of genes. However, the miR-29 family members
exhibit differential regulation in several cases and different
subcellular distribution, suggesting their functional relevance may
not be identical (30,31). It may be one of reason why the
function of miR-29 is different between NSPCs and some types of
cancer cells. Previous studies demonstrated that miR-29 regulated
cell behavior not only by regulating the expression of PTEN
protein; a number of tumor-suppressor genes have also been
identified as targets for miR-29, supporting its proposed
pro-proliferative role in cells (31). These genes include various
components of the p53 network, c-Myc and Smad, all of which are
involved in tumorigenesis, cell cycle control and apoptosis
(32,33). Additionally, an increasing number
of miR-29 targets are being identified, and novel regulatory
mechanisms of miR-29 are being determined. Therefore, it is
possible that the effects of miR-29 on the proliferation of NSPCs
were mediated by other targets, in addition to or instead of PTEN.
The present study reported that overexpression of miR-29 increased
the viability of NPSCs and reduced the expression of PTEN. The
proliferation of NSPCs was further promoted by miR-29, as indicated
by increases in the number of BrdU-positive cells and the PI. Of
note, VO-OH, a PTEN-specific inhibitor, attenuated the
anti-proliferative effects of miR-29 inhibitor on NSPCs.
Furthermore, the proliferation effects of miR-29 might be due to
suppression the expression of PTEN in cultured rat NSPCs. These
data may provide a theoretical basis and reference for the clinical
treatment of CNS diseases. However, additional research must be
conducted to explain the precise mechanism by which miR-29 promotes
proliferation of NSPCs.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81000030) and Fundamental
Research funds for the Central Universities (grant no.
xzy012019104).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YG, HQ and YH designed the experiments. YH
supervised the research. YG, HQ and ZL performed most of
experiments. YG drafted the manuscript. HQ and YH revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Animal Care and Use Regulation of Xi'an Jiaotong University Health
Science Center.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CNS
|
central nervous system
|
|
NSPCs
|
neural stem/progenitor cells
|
|
PTEN
|
phosphatase and tensin homologue
deleted on chromosome 10
|
|
BrdU
|
5-bromo-2-deoxyuridine
|
|
EGF
|
epidermal growth factor
|
|
bFGF
|
fibroblast growth factor
|
References
|
1
|
Bond AM, Ming GL and Song H: Adult
mammalian neural stem cells and neurogenesis: Five decades later.
Cell Stem Cell. 17:385–395. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Taupin P: Adult neurogenesis in the
mammalian central nervous system: Functionality and potential
clinical interest. Med Sci Monit. 11:RA247–RA252. 2005.PubMed/NCBI
|
|
3
|
Zhao C, Deng W and Gage FH: Mechanisms and
functional implications of adult neurogenesis. Cell. 132:645–660.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Temple S: The development of neural stem
cells. Nature. 414:112–117. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carthew RW and Sontheimer EJ: Origins and
mechanisms of miRNAs and siRNAs. Cell. 136:642–655. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cui Y, Li T, Yang D, Li S and Le W: miR-29
regulates Tet1 expression and contributes to early differentiation
of mouse ESCs. Oncotarget. 7:64932–64941. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xiong Y, Fang JH, Yun JP, Yang J, Zhang Y,
Jia WH and Zhuang SM: Effects of microRNA-29 on apoptosis,
tumorigenicity, and prognosis of hepatocellular carcinoma.
Hepatology. 51:836–845. 2010.PubMed/NCBI
|
|
8
|
Fráguas MS, Eggenschwiler R, Hoepfner J,
Schiavinato JL, Haddad R, Oliveira LH, Araújo AG, Zago MA,
Panepucci RA and Cantz T: MicroRNA-29 impairs the early phase of
reprogramming process by targeting active DNA demethylation enzymes
and Wnt signaling. Stem Cell Res. 19:21–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hilz S, Fogarty EA, Modzelewski AJ, Cohen
PE and Grimson A: Transcriptome profiling of the developing male
germ line identifies the miR-29 family as a global regulator during
meiosis. RNA Biol. 14:219–235. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Z, Ma W, Wang L, Gong H, Tian Y,
Zhang J, Liu J, Lu H, Chen X and Liu Y: Activation of type 4
metabotropic glutamate receptor attenuates oxidative stress-induced
death of neural stem cells with inhibition of JNK and p38 MAPK
signaling. Stem Cells Dev. 24:2709–2722. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen L, Liu M, Luan Y and Liu Y, Zhang Z,
Ma B, Liu X and Liu Y: BMP6 protects retinal pigment epithelial
cells from oxidative stressinduced injury by inhibiting the MAPK
signaling pathways. Int J Mol Med. 42:1096–1105. 2018.PubMed/NCBI
|
|
13
|
Zhang Z, Hu F, Liu Y, Ma B, Chen X, Zhu K,
Shi Y, Wei T, Xing Y, Gao Y, et al: Activation of type 5
metabotropic glutamate receptor promotes the proliferation of rat
retinal progenitor cell via activation of the PI-3-K and MAPK
signaling pathways. Neuroscience. 322:138–151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yuan TL and Cantley LC: PI3K pathway
alterations in cancer: Variations on a theme. Oncogene.
27:5497–5510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zheng H, Ying H, Yan H, Kimmelman AC,
Hiller DJ, Chen AJ, Perry SR, Tonon G, Chu GC, Ding Z, et al: p53
and Pten control neural and glioma stem/progenitor cell renewal and
differentiation. Nature. 455:1129–1133. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao L, Liu P, Gill K, Reece EA, Cheema AK
and Zhao Z: Identification of novel cell survival regulation in
diabetic embryopathy via phospholipidomic profiling. Biochem
Biophys Res Commun. 470:599–605. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin X, Zhou X, Liu D, Yun L, Zhang L, Chen
X, Chai Q and Li L: MicroRNA-29 regulates high-glucose-induced
apoptosis in human retinal pigment epithelial cells through PTEN.
In Vitro Cell Dev Biol Anim. 52:419–426. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zanotti S, Gibertini S, Curcio M, Savadori
P, Pasanisi B, Morandi L, Cornelio F, Mantegazza R and Mora M:
Opposing roles of miR-21 and miR-29 in the progression of fibrosis
in Duchenne muscular dystrophy. Biochim Biophys Acta.
1852:1451–1464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zou H, Ding Y, Shi W, Xu X, Gong A, Zhang
Z and Liu J: MicroRNA-29c/PTEN pathway is involved in mice brain
development and modulates neurite outgrowth in PC12 cells. Cell Mol
Neurobiol. 35:313–322. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tamguney T and Stokoe D: New insights into
PTEN. J Cell Sci. 120:4071–4079. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Janku F, Hong DS, Fu S, Piha-Paul SA,
Naing A, Falchook GS, Tsimberidou AM, Stepanek VM, Moulder SL, Lee
JJ, et al: Assessing PIK3CA and PTEN in early-phase trials with
PI3K/AKT/mTOR inhibitors. Cell Rep. 6:377–387. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kondo T, Funayama M, Tsukita K, Hotta A,
Yasuda A, Nori S, Kaneko S, Nakamura M, Takahashi R, Okano H, et
al: Focal transplantation of human iPSC-derived glial-rich neural
progenitors improves lifespan of ALS mice. Stem Cell Reports.
3:242–249. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Le Belle JE, Orozco NM, Paucar AA, Saxe
JP, Mottahedeh J, Pyle AD, Wu H and Kornblum HI: Proliferative
neural stem cells have high endogenous ROS levels that regulate
self-renewal and neurogenesis in a PI3K/Akt-dependant manner. Cell
Stem Cell. 8:59–71. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu H, Goel V and Haluska FG: PTEN
signaling pathways in melanoma. Oncogene. 22:3113–3122. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gregorian C, Nakashima J, Le Belle J, Ohab
J, Kim R, Liu A, Smith KB, Groszer M, Garcia AD, Sofroniew MV, et
al: Pten deletion in adult neural stem/progenitor cells enhances
constitutive neurogenesis. J Neurosci. 29:1874–1886. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mulholland DJ, Kobayashi N, Ruscetti M,
Zhi A, Tran LM, Huang J, Gleave M and Wu H: Pten loss and RAS/MAPK
activation cooperate to promote EMT and metastasis initiated from
prostate cancer stem/progenitor cells. Cancer Res. 72:1878–1889.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang X, Zhu LL, Zhao T, Wu LY, Wu KW,
Schachner M, Xiao ZC and Fan M: CHL1 negatively regulates the
proliferation and neuronal differentiation of neural progenitor
cells through activation of the ERK1/2 MAPK pathway. Mol Cell
Neurosci. 46:296–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Torsvik A and Bjerkvig R: Mesenchymal stem
cell signaling in cancer progression. Cancer Treat Rev. 39:180–188.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hausenloy DJ, Mocanu MA and Yellon DM:
Cross-talk between the survival kinases during early reperfusion:
Its contribution to ischemic preconditioning. Cardiovasc Res.
63:305–312. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kriegel AJ, Liu Y, Fang Y, Ding X and
Liang M: The miR-29 family: Genomics, cell biology, and relevance
to renal and cardiovascular injury. Physiol Genomics. 44:237–244.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Zhang X, Li H, Yu J and Ren X: The
role of miRNA-29 family in cancer. Eur J Cell Biol. 92:123–128.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He Y, Huang C, Lin X and Li J: MicroRNA-29
family, a crucial therapeutic target for fibrosis diseases.
Biochimie. 95:1355–1359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan B, Guo Q, Fu FJ, Wang Z, Yin Z, Wei YB
and Yang JR: The role of miR-29b in cancer: Regulation, function,
and signaling. Onco Targets Ther. 8:539–548. 2015.PubMed/NCBI
|














