Introduction
The occurrence of postoperative cognitive
dysfunction (POCD) is closely associated with the inflammatory
response of the central nervous system (CNS) (1). POCD is characterized by progressive
deterioration of cognitive function and reduced self-care ability.
Permanent cognitive disabilities can develop into neurodegenerative
diseases similar to Alzheimer's disease and dementia, and may lead
to the loss of independent living abilities (2). Trauma caused by surgery is regarded
as a major cause of POCD in the senior population (3), but its neurobiological basis is
largely unknown. A previous study demonstrated that the learning
ability of mice is reduced, and interleukin 1β (IL-1β) expression
in the hippocampus is significantly increased following
lipopolysaccharide (LPS) injection into the peripheral blood of
mice (4). Cao et al
(5) and Wuri et al
(6) separately performed partial
hepatectomies on rats and mice; they found decreased learning and
memory ability, and increased inflammatory factors in the
hippocampus in the postoperative animals (5,6).
Dexmedetomidine is a commonly used α2 receptor
agonist with sedative and analgesic effects. Its α2:α1 ratio is
1,600:1, and its affinity is eight times greater than that of
clonidine (7). It produces
sedative, hypnotic and anxiolytic effects by acting on α2 receptors
in the locus coeruleus of the brain stem (8). A previous study revealed that
dexmedetomidine exhibits anti-inflammatory effects at multiple
sites (9), including in lung,
kidney and rat sepsis models. It exhibits neuroprotective effects
in animal models, including ischemic brain injury and spinal cord
injury models (10). Additionally,
a previous study demonstrated that dexmedetomidine may inhibit
inflammation caused by LPS-induced microglial activation (11). A previous study indicated that
postoperative cognitive impairment is associated with inflammation
of the CNS (12). The activation
of microglia and NF-κB can lead to overexpression of inflammatory
factors, including tumor necrosis factor α (TNF-α) and IL-1β, which
may cause postoperative cognitive decline (13). NF-κB is involved in the regulation
of neuroinflammation, such as that associated with cerebral
ischemia and hypoxia. It is a key molecule of the inflammatory
response, which mediates the expression of inflammatory factors in
multiple signaling pathways (14).
A previous study demonstrated that activated NF-κB can induce the
expression of inflammatory factors in microglia (15).
In the present study, a model of hepatectomy in rats
was selected to simulate cognitive dysfunction following surgery.
The effect of dexmedetomidine on the expression levels of TNF-α,
IL-1β and NF-κB in the hippocampus was examined to explore whether
dexmedetomidine may inhibit the inflammatory response in the CNS,
and its possible mechanism of action.
Materials and methods
Ethics approval
The present study was approved by the Institutional
Animal Care and Use Committee of Zhejiang Hospital.
Establishment of a rat model of
POCD
A partial hepatectomy was used to establish a POCD
model in aged rats. A total of 80 male Sprague Dawley rats (age, 18
months; weight, 500–600 g) were provided by the Experimental Animal
Center of Zhejiang Hospital. The animals were housed at 22±2°C, a
relative humidity of 45–75% and with a 12 h light-dark cycle. Food
and water were freely accessible. Preoperative fasting was carried
out for 12 h. The rats were administered inhaled isoflurane
(1.5–2.0%) for anesthesia. Tracheal intubation was performed and
mechanical ventilation was used in the rats. The rats were
anesthetized throughout the operation by exposure to a mixture of
air (21% O2 with 79% N2) and 1.5–2.0%
isoflurane. Following disinfection, a 3-cm vertical incision was
made at the lower edge of the xiphoid process, and the left hepatic
lobe was dissected. The left hepatic lobe was probed and ligated to
the root, and subsequently resected. The incision was infiltrated
with 0.25% bupivacaine (Nanjing Chemlin Chemical Industry Co.,
Ltd.) and sutured with a 3-0 suture. The procedure was performed
aseptically and the surgery time was controlled at ~30 min. After
the rats returned to consciousness, they were returned to the
animal room.
Animal grouping
The upper limit of the 95% CI of the mean training
duration of rats in the Y-maze test was used to decide whether the
rats that had undergone surgery had cognitive dysfunction,
according to a previously described method (16). Among the 80 rats that received
hepatectomy, 72 rats developed POCD. These 72 rats were randomly
divided into four groups: i) Blank control group; ii) low-dose
anesthesia group; iii) high-dose anesthesia group; and iv) surgery
model group. The blank control group was not treated. In a
transparent anesthetic chamber, rats were exposed for 4 h to 1.5%
isoflurane (Baxter) in 2 l/min of 100% oxygen (Renyuan Chem) as the
carrying gas, or to vehicle (2 l/min of 100% oxygen), 24 h after
hepatectomy. At the outlet of the chamber, the gas composition
(concentrations of isoflurane, oxygen and CO2) within
the chamber was continuously analyzed by a gas monitor
(Datex-Ohmeda; GE Healthcare). Following anesthesia, the rats
received 100% oxygen until they recovered to consciousness. The
heart rate and oxygen saturation of the rats were monitored to
maintain normal heart rate and >90% oxygen saturation during
surgery. Dexmedetomidine (10 and 30 µg/kg; Nanjing Chemlin Chemical
Industry Co., Ltd.) was intraperitoneally injected 1 h after
surgery for the low- and high-dose anesthesia groups, respectively.
The dose selection was based on previous studies (17,18).
All rats received an intramuscular injection of penicillin (200,000
units) daily for 3 days after surgery to prevent infection. A total
of 6 rats were taken from each group on the 1st, 3rd and 7th day
after treatment to perform a Y-maze test. Following anesthesia with
2% pentobarbital sodium (50 mg/kg; intraperitoneal), the
hippocampus was quickly dissected and separately immersed into
liquid nitrogen and maintained at −80°C prior to analysis.
Peripheral blood was collected and placed into an anticoagulant
tube. Following the intervention, the rats were sacrificed by
cervical dislocation. The supernatant of the solution in the
anticoagulant tube was removed by centrifugation (8,000 × g for 20
min at 4°C). The extracted hippocampal tissues and supernatants
were stored in a refrigerator at −80°C for subsequent use.
Behavioral testing
A behavior test (Y-maze) was generally designed to
confirm whether POCD developed in rats suffering from surgical
trauma. A total of 6 rats were taken from each group at 1, 3 and 7
days after treatment, and a Y-maze test was performed between 9:00
and 12:00 a.m. The Y-maze test was performed using a previously
described method with minor modification (19). The Y-maze was divided into three
arms, labeled as I, II and III, and the angle between the three
arms was 120°. The size of each arm was 40×30×15 cm, and the length
of the equilateral triangle encircled by the three arms was 15 cm.
The light source and camera were set 2 m above the center of the Y
maze. A cloth was installed on the outside of the maze to prevent
external light interference. The three arms of the Y maze were
randomly set as start arm, novel arm and the other arm. Prior to
testing, the novel arm was blocked with a septum, and the rat was
placed in the start arm and allowed to move freely in the start arm
and the other arm. The rat was removed after 10 min of adaptation.
After 1 h, the test was started: The novel arm was opened, and the
rat was placed in the start arm and allowed to freely move among
the three arms for 5 min. The shuttle path was recorded. At the
bottom of the labyrinth, wood chips were placed on the bottom of
the maze. Following each training, the wood chips in the three arms
were mixed and re-paved. After each test, the labyrinth was cleaned
to remove odors and residues to prevent any influence on the test
results. In the experiment, different rats were randomly arranged
with different start, novel and other arms. During the training and
testing of the same rat, the settings were not changed. The entire
process of the test was observed and recorded by the camera
directly above the maze. The motion path of test was recorded, and
the total entries and the spontaneous alternation score were
calculated. The calculation method was as follows: Assume that the
novel arm is A, the start arm is B, and the other arm is C. For
example, the motion path during the test of the rat is BACABCBAB.
During the test, the rat passes through 9 arms, thus the number of
shuttles is 9. Continuous ABC, BCA and CAB were recorded as 1 point
at a time. In this example, there were BAC, CAB, ABC and CBA; the
alternation score was 4, and the alternation ratio formula was: The
spontaneous alternation score (%)=[(number of alternation)/(total
arm entries-2)]x100%. In this example, the alternation scoring
rate=4/7×100%=57.14%. The total number of passing arms in the
Y-maze is considered as an index reflecting the exercise ability of
the rats.
ELISA for detection of IL-1β and
TNF-α
An ELISA was used to detect the expression levels of
IL-1β and TNF-α in hippocampal tissues. Frozen hippocampus samples
from four POCD rats from each group were mixed with an appropriate
amount of physiological saline. They were fully homogenized and
centrifuged at a speed of 8,000 × g for 10 min at 4°C. The
supernatant was collected and tested using a rat IL-1β ELISA kit
(cat. no. ab100768; Abcam) or a rat TNF-α ELISA kit (cat. no.
ab100785; Abcam), according to the manufacturer's protocol.
Determining NF-κB p65 expression in
the hippocampus by western blotting
Protein extraction was performed using RIPA buffer
and a protease inhibitor cocktail [1.4% SDS, 2% dithiothreitol, 120
mM Tris-Cl (pH 6.8), 7 M urea, 2 M sulfourea, 20 mM DTT, 3.20 mM
Tris (pH 7.5), 1.5 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100,
2.5 mM sodium pyrophosphate, 1 mM β-glycerophosphate]. A
bicinchoninic acid assay kit (Sigma-Aldrich; Merck KGaA) was used
to determine protein concentration in samples from four POCD rats.
After sample buffer was added, the samples were boiled at 95°C for
10 min. Then, the proteins (30 µg/sample/well) were separated using
10% polyacrylamide gel electrophoresis. After electrophoresis,
proteins were transferred onto PVDF membranes at 100 V
transfer-molded voltage for 45–70 min. Subsequently, samples were
incubated at room temperature for 1 h with 5% BSA (Shanghai
Rebiosci Biotechnology Co., Ltd.), and then incubated with rabbit
anti-NF-κB/p65 (1:1,000; cat. no. ab16502; Abcam) and mouse
anti-β-actin primary antibodies (1:1,000; cat. no. ab8226; Abcam),
at 4°C overnight. Following that, samples were washed with TBS with
Tween-20 three times (5 min each). A horseradish peroxidase
secondary antibody (1:3,000; cat. no. ab6721; Abcam) was added for
incubation at room temperature for 1 h. Subsequently, membranes
were washed three times (5 min/time). Development was performed
using SuperSignal West Femto chemiluminescence reagent (Thermo
Fisher Scientific, Inc). β-actin was used as an internal reference.
Bands were visualized with a Bio-Rad Gel Doc EZ imager (Bio-Rad
Laboratories, Inc.). ImageJ software (version 1.8.0_112; National
Institutes of Health) was used to analyze the intensity of the
target bands.
Immunofluorescent staining for
detection of activation of microglial cells
Ionized calcium-binding adapter molecule 1 (Iba-1)
is a biomarker of microglia and macrophages (20). By labeling microglia with Iba-1
antibody, the activated microglia may be quantified. Frozen
hippocampal tissues were cut into 2–3 mm slices and loaded into
embedding cassettes. The tissues were submerged and fixed in a
solution of neutral buffered formalin for at least 48 h at room
temperature. When using zinc fixatives (formalin-zinc sulfate
fixative; Beijing Solarbio Science and Technology Co., Ltd.), the
same sized tissues (2–3 mm), were fixed in a zinc fixation buffer
for 24 h at room temperature. Tissues were then fixed in paraffin
for 10–15 min at room temperature. The paraffin embedded sections
of hippocampal tissue of four POCD rats from each group were
dewaxed, hydrated and antigen-repaired. To enhance the
accessibility of the antibody to antigens, antigen retrieval was
performed prior to incubation with the primary antibody. Briefly,
slides were placed in citrate buffer in a slide container and the
container was placed in a metal container filled with distilled
water in a decloaker. The decloaker was heated to a temperature of
122°C and was allowed to naturally air cool to 89°C. The lid was
then opened for further cooling. Slides were blocked using 5% goat
serum (Abcam) in PBS for 10 min at room temperature and then
incubated with primary anti-Iba-1 (1:500; cat. no. ab178847; Abcam)
at 4°C overnight. Following incubation with fluorescent secondary
antibody (AlexaFluor488-conjugated anti-rabbit; 1:1,000; cat. no.
A27034; Invitrogen; Thermo Fisher Scientific, Inc.) for 2 h in the
dark, the nucleus was stained with DAPI (1 µg/ml) and mounted for
10 min at room temperature, and observed under a confocal
fluorescence microscope (magnification, ×400). A total of five
independent fields were selected and the activated microglia and
total microglia in the visual field were counted: Microglial
activation rate = activated microglia number/total microglia number
×100%. The microglial activation rates from five fields were
averaged as the microglial activation rate of the specimen.
Statistical analysis
All data were analyzed by GraphPad Prism version 6
statistical software (GraphPad Software, Inc.). Each experiment was
repeated three times. Measurement data are presented as the mean ±
SD. One-way ANOVA followed by a Least Significant Difference post
hoc test was applied for comparisons among multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Alterations in the cognitive function
of rats in each group
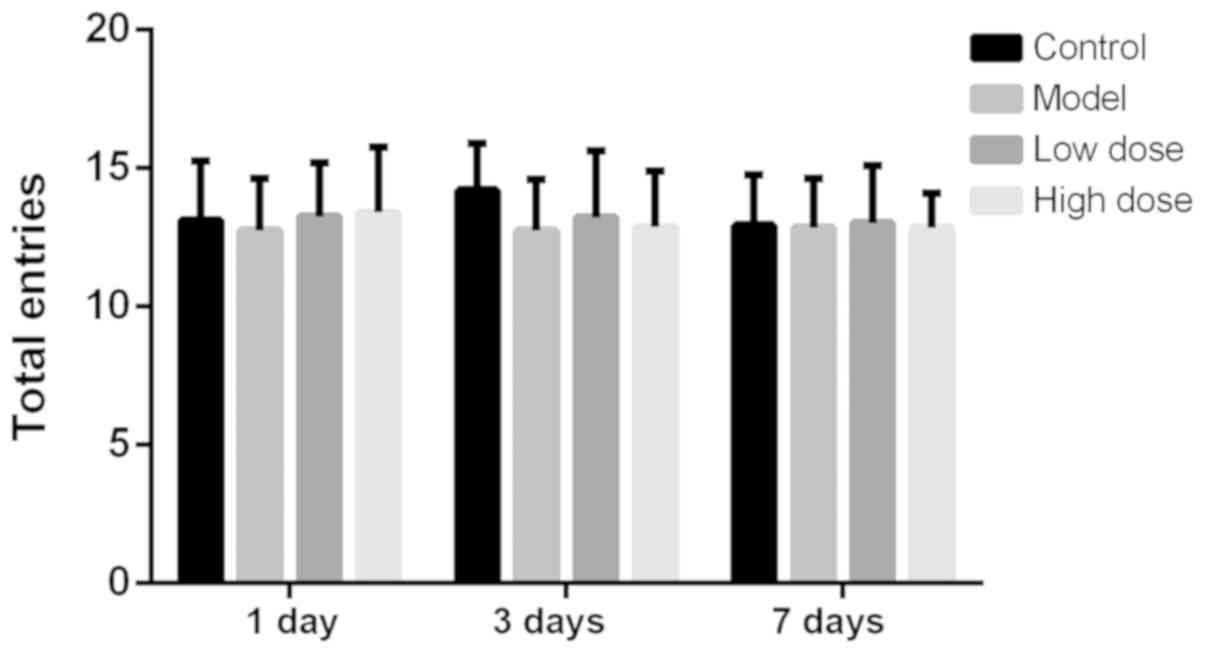
The total number of arms a rat passes in the Y-maze
test is considered to be an index reflecting the exercise ability
of the rats (17). The number of
rats passing alternative arms at each time point is shown in
Fig. 1. There was no statistical
difference in the number of rats passing alternative arms in each
group at 1, 3 or 7 days after surgery. This indicated that neither
anesthesia nor partial hepatectomy affected the exercise activity
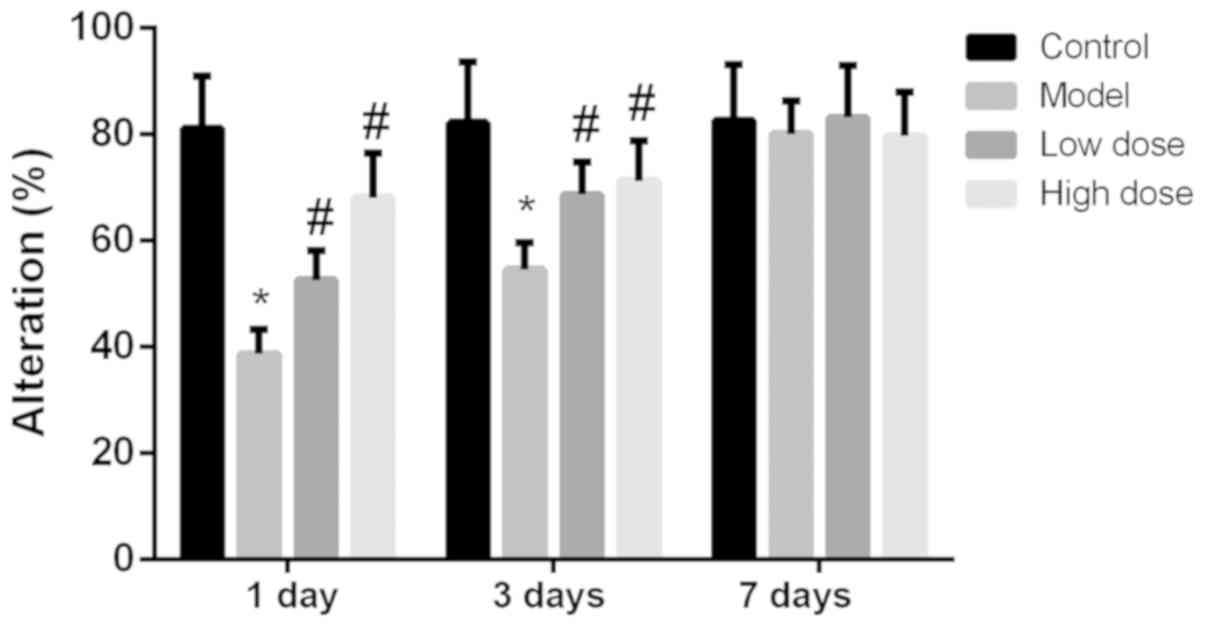
of rats. The alternation scoring rate of each group of rats at 1, 3
and 7 days is shown in Fig. 2. The
alternation score of the model group was significantly lower than
that of the control group at 1 and 3 days (P<0.05). Compared
with the model group, the alternation scores of the high-dose and
low-dose anesthesia groups were significantly higher at 1 and 3
days (P<0.05), particularly in the high-dose group. On the
seventh day, there was no statistically significant difference
identified in the alternation score among the rats in each group
(P>0.05). These results indicated that anesthesia and surgery
did not alter the exercise ability of the rats, but decreased the
cognitive function of the rats.
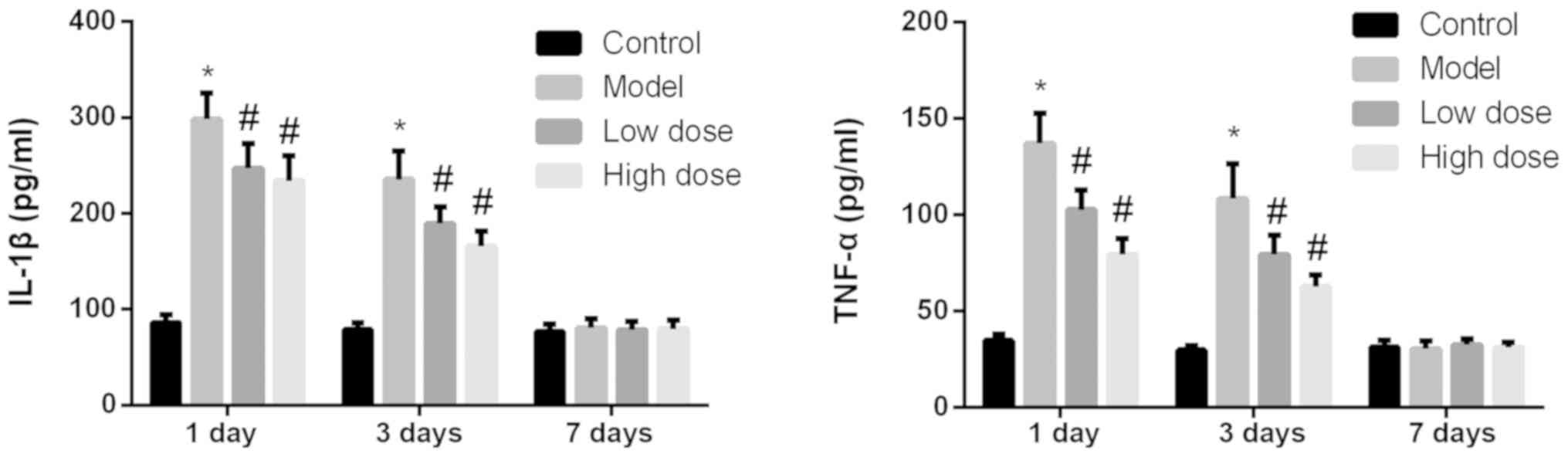
Anesthesia and surgery lead to
decreased expression levels of IL-1β and TNF-α in the rat
hippocampus
The expression levels of IL-1β and TNF-α in the
hippocampi of rats from each group were detected by ELISA. As shown
in Fig. 3, at 1 and 3 days after
surgery, the expression levels of IL-1β and TNF-α in the model
group were significantly higher than those in the control group
(P<0.05). The expression levels of IL-1β and TNF-α in the
high-dose and low-dose anesthesia groups decreased significantly
(P<0.05), and the drop in the high-dose group was slightly more
pronounced. On day 7 after surgery, the expression levels of IL-1β
and TNF-α were low in all groups and there were no significant
differences identified among the groups (P>0.05).
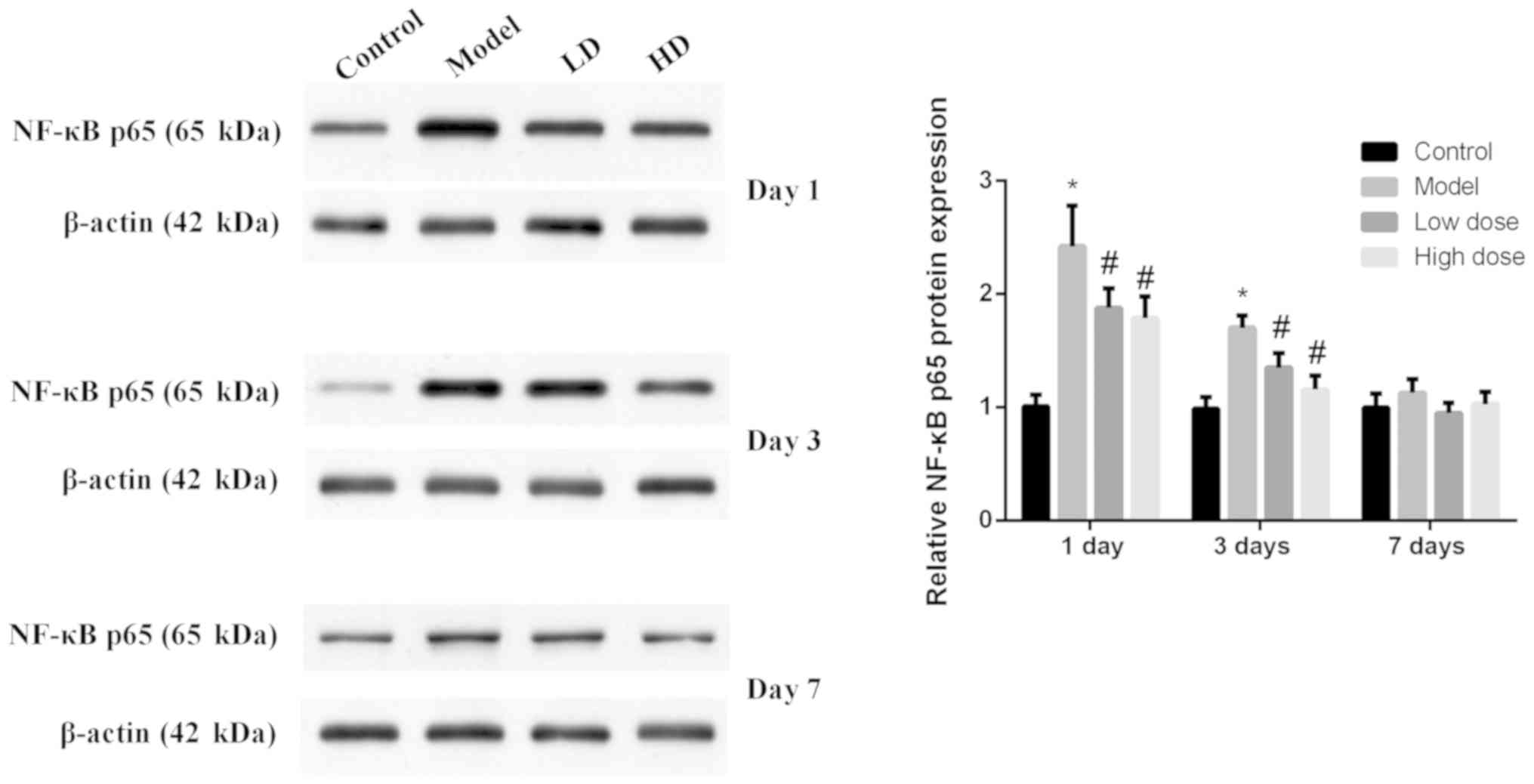
Expression levels of NF-κB p65 in rat
hippocampus tissues at each time point for each group
The expression levels of NF-κB p65 in rat
hippocampus tissues were detected by western blotting (Fig. 4). NF-κB p65 expression in the model
group was significantly increased compared with the control group
at 1 and 3 days after surgery (P<0.05). The expression levels of
NF-κB p65 in the high-dose and low-dose groups were significantly
decreased compared with the model group (P<0.05), and the
decrease in the high-dose group was more obvious. At 7 days after
surgery, the expression levels of NF-κB p65 relatively low and
there was no significant difference identified among the groups
(P>0.05).
Activation of microglial cells in rat
brains
By labeling microglia with Iba-1 antibody, activated
microglia may be quantified. The Iba-1 antibody immunofluorescence
staining results are shown in Fig.
5. At 1 and 3 days after surgery, the microglial cells in the
brain tissues of rats in the control group were in an inactive
state, the cell bodies were small and irregular protrusions were
observed. By contrast, the microglial cells in the brain tissues of
rats in the model group were large, the cell bodies were round, and
they exhibited an activation state of deformation and phagocytosis.
On day 1, the quantification of immunofluorescence staining
revealed that the proportion of activated microglial cells was
79.54±5.52% in the model group, which was significantly higher than
that of the control group (P<0.05). The activation of microglia
in brain tissue from the high-dose and low-dose anesthesia groups
was inhibited, and the activation rates were 21.84±2.03 and
39.45±3.55%, respectively, which were significantly lower than that
of the model group (P<0.05). The activation of microglia in
brain tissue was decreased to 55.16±5.23% in the model group and to
15.97±1.26% (low-dose) and 10.65±1.12% (high dose), respectively,
at 3 days after surgery. Microglia activation in the low-dose and
high-dose groups was significantly lower compared with the model
group at 1 and 3 days. At 7 days after surgery, there was no
statistically significant difference observed in the activation
rates of microglial cells in the brain tissues of each group
(P>0.05). These results indicated that microglia were activated
by dexmedetomidine.
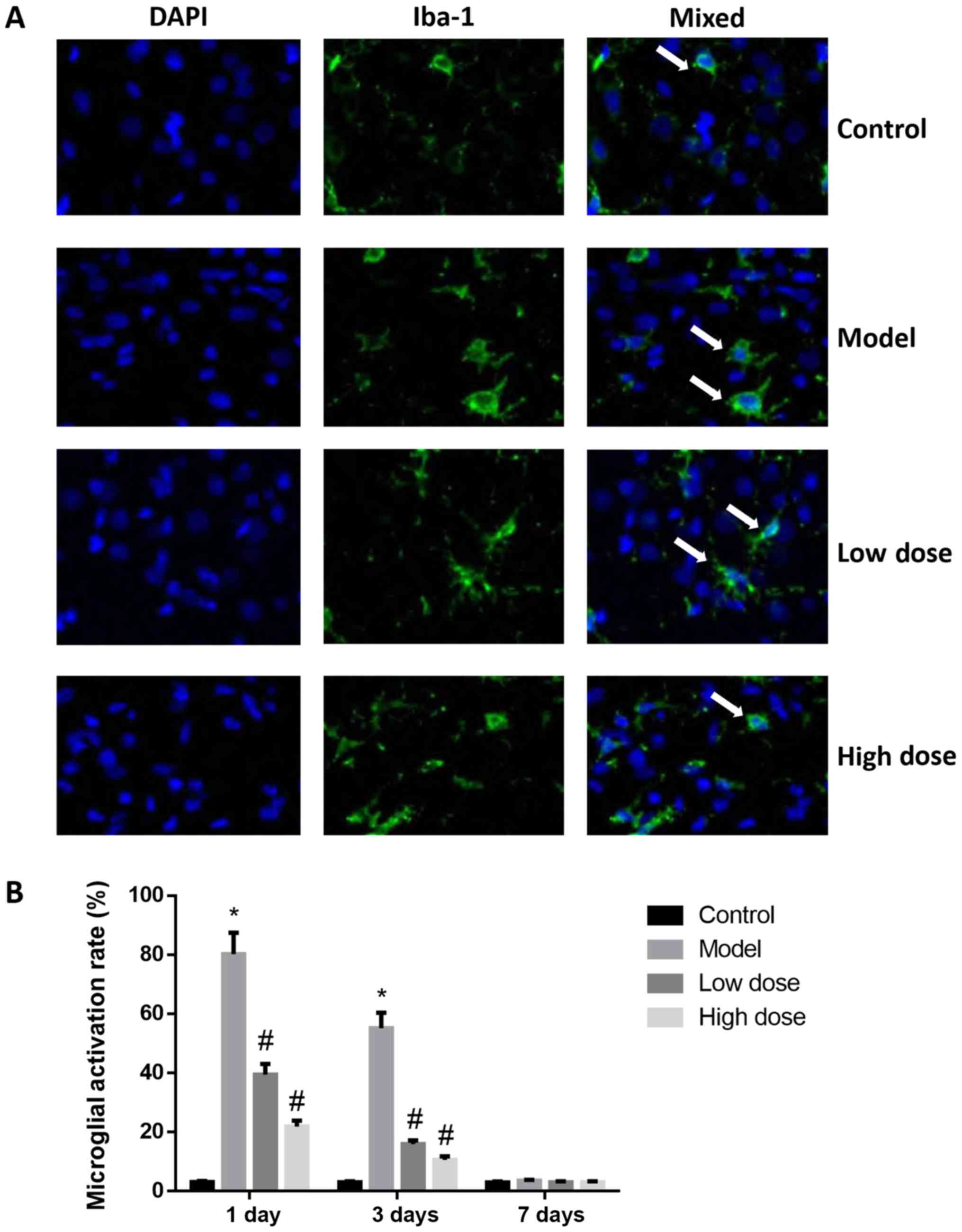 | Figure 5.Activation of microglia in the brains
of rats following surgery. (A) Immunofluorescence staining of
microglia in the hippocampus at 1 day after surgery. Magnification,
×400. Arrows indicate activated microglia. (B) Microglial
activation rates at 1, 3 and 7 days after surgery. At 1 day after
surgery, the microglial cells in the brain tissues of rats in the
control group were in an inactive state, cell bodies were small
with irregular protrusions. By contrast, the microglial cells in
the brain tissues of rats in the model group were large, cell
bodies were round and they exhibited an activation state of
deformation and phagocytosis. At day 1, the quantification of
immunofluorescence staining revealed that the proportion of
activated microglial cells was 79.54±5.52% in the model group,
which was significantly higher than that of the control group
(P<0.05). The activation of microglia in the brain tissue from
the high-dose and low-dose anesthesia groups was inhibited, the
activation rates were 21.84±2.03 and 39.45±3.55%, respectively,
which were significantly lower than that of the model group
(P<0.05). At 3 days after surgery, the activation of microglia
in brain tissue was decreased to 55.16±5.23% in the model group, to
15.97±1.26% in the low-dose and to 10.65±1.12% high-dose groups. At
7 days after surgery, there was no statistically significant
difference observed in the activation rates of microglial cells in
the brain tissues of each group (P>0.05). These results
indicated that microglia were activated by dexmedetomidine.
*P<0.05 vs. control group; #P<0.05 vs. model
group. Iba-1, ionized calcium-binding adapter molecule 1. |
Discussion
Tissue injury caused by surgical trauma leads to the
activation of the immune system, and release of inflammatory
factors (21). The amount of
inflammatory factor release is associated with the degree of
surgical trauma (22). To
understand whether surgical trauma could affect cognitive function
in rats, a partial hepatectomy was performed and a Y-maze test was
conducted. The results revealed a non-significant difference in the
total entry numbers of rats passing the Y-maze in the control and
model groups. However, the alternation score of rats in the model
group was significantly lower than that of the control group at 1
and 3 days after surgery, and dexmedetomidine treatment
significantly increased this score. At 7 days, the alternation
score in rats of all groups returned to the level of the control
group. These results indicated that cognitive function was
decreased following hepatectomy, and dexmedetomidine could protect
postoperative cognitive function in the short term.
Under physiological conditions, the presence of the
blood-brain barrier prevents the entry of macromolecule proteins
into brain tissue. However, the permeability of the blood-brain
barrier increases under certain circumstances, such as in ischemia,
trauma, tumors, neurological conditions associated with neuronal
hyper-excitability, epileptic seizures and spreading
depolarizations. In the case of POCD, a large number of
inflammatory factors and immune cells from the peripheral blood can
enter into the brain (23). A
previous study demonstrated that cutting the glossopharyngeal nerve
can block the thermal response of the soft palate induced by
injection of LPS (24).
Additionally, the fever caused by intraperitoneal injection of LPS
may be decreased after the vagus nerve is cut (25). These findings suggest that primary
sensory nerves may mediate inflammatory factors and indirectly
affect the brain tissue. Peripheral inflammatory mediators act
inside the brain tissue, causing microglial cells to be rapidly
activated, and to release a large number of inflammatory factors
(26). In addition, neurons and
peripheral lymphocytes entering the brain can release a variety of
inflammatory factors, including IL-1β, TNF-α and interferon γ,
which, in turn, activate microglia cells and further stimulate
signaling transduction pathways, resulting in a cascade of
amplified inflammatory responses (27). The inflammatory response in the
brain is involved in numerous processes of degenerative alterations
in the nervous system. On one hand, inflammation can resist the
damage to the body caused by external stimuli; on the other hand,
inflammation can damage body tissues through various mechanisms,
including degeneration, necrosis, metabolic dysfunction of the
parenchymal cells in the inflammatory foci, and mechanical
obstruction and compression caused by inflammatory exudates may
cause dysfunction of the inflammatory organs. Pain can also affect
the active function of the brain (28). Accumulation of a large number of
inflammatory mediators and other toxicants in the brain can cause
reversible or irreversible damage to the brain tissue, and impaired
cognitive function. The hippocampus exhibits widely-expressed
receptors for inflammatory factors and is more sensitive to
abnormal increases in inflammatory cytokines (29). IL-1β and TNF-α can directly or
indirectly penetrate the blood-brain barrier, and they are a class
of inflammatory factors with a wide range of biological activities
(30). Subsequent to entering the
brain, these inflammatory factors cause cell edema, affecting the
function of synaptic connections, causing an inflammatory response
in the CNS and leading to impairments of cognitive function. If the
concentration of IL-1β in the hippocampus is high, it may affect
the plasticity of the synapse, and thus affect long-term
potentiation, resulting in impaired learning and memory (31). IL-1β and TNF-α can stimulate actin
in cells other than intracranial neurons, leading to the
regeneration of actin, which serves an important role in the
process of neurodegenerative diseases (32,33).
In the present study, IL-1β and TNF-α expression in rat hippocampi
were measured by an ELISA, and NF-κB p65 expression was measured by
western blotting. The results revealed that their expression levels
were increased following hepatectomy (for 1–3 days), which was
consistent with previous studies (4,8,34).
Additionally, the present study demonstrated that dexmedetomidine
could significantly reduce their expression levels (between 1 and 3
days), and this reduction was concentration-dependent.
Microglia are macrophage-like cells in the CNS which
perform immune surveillance and defense functions in the brain
(35). Under normal conditions,
the microglia are in a resting state, their cell bodies are small
and there are long protrusions surrounding the cells. In this
state, microglia lack phagocytic function (36). Microglia can respond to subtle
pathological alterations in the nervous system. Microglia can be
activated by infection, trauma and ischemia, and perform innate
immune functions, including the induction of inflammation,
cytotoxic effects and modulation of T cell responses via antigen
presentation (37). Activated
microglia can release inflammatory factors, including TNF-α, IL-1β,
IL-6, nitric oxide and prostaglandin E2 (38). These inflammatory factors exhibit a
strong toxic effect on neurons and reactivate microglia,
aggravating or expanding the inflammatory response of the CNS
(39). Therefore, inhibiting the
expression of central inflammatory factors may serve a role in
improving postoperative cognitive function, and the results of the
present study supported this hypothesis. The present study revealed
that microglia were activated after hepatectomy and returned to a
resting state at 7 days. Dexmedetomidine could significantly reduce
the activation of microglia in POCD rats in a
concentration-dependent manner. These results indicated that
microglia responded to inflammatory alterations and were involved
in POCD development, which was consistent with previous studies
(13,35,39).
Dexmedetomidine is commonly used as an α2-agonist in
clinical practice (8). A previous
study reported that the anti-inflammatory effects of
dexmedetomidine may be associated with its central sympathomimetic
effects and the activation of cholinergic anti-inflammatory
pathways (40). However, its
mechanism of action has not yet been elucidated. Hoffman et
al (41) applied
dexmedetomidine to a rat model of cerebral ischemia and found that
there is a dose-associated decrease in blood catecholamine content
and histopathological improvement in rats, and this improvement is
associated with decreased sympathetic nerve activity. Ma et
al (34) reported that
dexmedetomidine inhibits systemic inflammatory responses and
improves survival in septic shock rats. Additionally, Fang et
al (42) demonstrated the
anti-inflammatory effects of dexmedetomidine in rat spinal cord
injury models. In clinical applications, Yang and Hong (43) reported that dexmedetomidine
sedation can significantly reduce the expression levels of
pro-inflammatory cytokines IL-1β, TNF-α and IL-6 in the plasma in
critically ill patients. The present study demonstrated that
dexmedetomidine served an anti-inflammatory role, inhibited the
expression of IL-1β, TNF-α and NF-κB p65, which are crucial for
microglial activation and cognitive dysfunction, and that its
effect was concentration-dependent. However, during the present
study, it was noted that the NF-κB p65 expression should be
assessed by separating the nucleus and cytoplasm; in order to be
more exact, a nuclear/cytoplasmic fractionation is required prior
to protein quantification. The present study was also limited to
observation and simple detection methods. Further studies are
required to investigate how dexmedetomidine suppresses inflammation
and improves postoperative cognitive function.
In conclusion, the present study found that the
intraperitoneal administration of dexmedetomidine may effectively
inhibit the release of inflammatory mediators in the CNS and
improve the cognitive function of aged rats. Therefore, the present
study may provide a potential method for the treatment of POCD in
older patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JQ designed the study. NC and XC analyzed the data
and wrote the manuscript. JX performed experiments, including
western blotting, ELISA and immunofluorescent staining. CW
established the animal model and carried out the behavioral tests.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures were approved by the Ethics Committee
of Zhejiang Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xu J, Dong H, Qian Q, Zhang X, Wang Y, Jin
W and Qian Y: Astrocyte-derived CCL2 participates in
surgery-induced cognitive dysfunction and neuroinflammation via
evoking microglia activation. Behav Brain Res. 332:145–153. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Berger M, Nadler JW, Browndyke J, Terrando
N, Ponnusamy V, Cohen HJ, Whitson HE and Mathew JP: Postoperative
cognitive dysfunction: Minding the gaps in our knowledge of a
common postoperative complication in the elderly. Anesthesiol Clin.
33:517–550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ballard C, Jones E, Gauge N, Aarsland D,
Nilsen OB, Saxby BK, Lowery D, Corbett A, Wesnes K, Katsaiti E, et
al: Optimised anaesthesia to reduce post operative cognitive
decline (POCD) in older patients undergoing elective surgery, a
randomised controlled trial. PLoS One. 7:e374102012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen J, Buchanan JB, Sparkman NL, Godbout
JP, Freund GG and Johnson RW: Neuroinflammation and disruption in
working memory in aged mice after acute stimulation of the
peripheral innate immune system. Brain Behav Immun. 22:301–311.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cao XZ, Ma H, Wang JK, Liu F, Wu BY, Tian
AY, Wang LL and Tan WF: Postoperative cognitive deficits and
neuroinflammation in the hippocampus triggered by surgical trauma
are exacerbated in aged rats. Prog Neuropsychopharmacol Biol
Psychiatry. 34:1426–1432. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wuri G, Wang DX, Zhou Y and Zhu SN:
Effects of surgical stress on long-term memory function in mice of
different ages. Acta Anaesthesiol Scand. 55:474–485. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Devasya A and Sarpangala M:
Dexmedetomidine: A review of a newer sedative in dentistry. J Clin
Pediatr Dent. 39:401–409. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Keating GM: Dexmedetomidine: A review of
its use for sedation in the intensive care setting. Drugs.
75:1119–1130. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Celik F, Göcmez C, Kamaşak K, Tufek A,
Guzel A, Tokgoz O, Fırat U and Evliyaoğlu O: The comparison of
neuroprotective effects of intrathecal dexmedetomidine and
metilprednisolone in spinal cord injury. Int J Surg. 11:414–418.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Koca U, Olguner ÇG, Ergür BU, Altekin E,
Taşdöğen A, Duru S, Girgin P, Gündüz K, Cilaker Mıcılı S, Güzeldağ
S and Akkuş M: The effects of dexmedetomidine on secondary acute
lung and kidney injuries in the rat model of intra-abdominal
sepsis. ScientificWorldJournal. 2013:2926872013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yeh CH, Hsieh LP, Lin MC, Wei TS, Lin HC,
Chang CC and Hsing CH: Dexmedetomidine reduces lipopolysaccharide
induced neuroinflammation, sickness behavior, and anhedonia. PLoS
One. 13:e01910702018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Han C, Fu R and Lei W: Beneficial effects
of dexmedetomidine on early postoperative cognitive dysfunction in
pediatric patients with tonsillectomy. Exp Ther Med. 16:420–426.
2018.PubMed/NCBI
|
|
13
|
Yang Z, Liu Y, Yuan F, Li Z, Huang S, Shen
H and Yuan B: Sinomenine inhibits microglia activation and
attenuates brain injury in intracerebral hemorrhage. Mol Immunol.
60:109–114. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Frakes AE, Ferraiuolo L, Haidet-Phillips
AM, Schmelzer L, Braun L, Miranda CJ, Ladner KJ, Bevan AK, Foust
KD, Godbout JP, et al: Microglia induce motor neuron death via the
classical NF-κB pathway in amyotrophic lateral sclerosis. Neuron.
81:1009–1023. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park J, Min JS, Kim B, Chae UB, Yun JW,
Choi MS, Kong IK, Chang KT and Lee DS: Mitochondrial ROS govern the
LPS-induced pro-inflammatory response in microglia cells by
regulating MAPK and NF-κB pathways. Neurosci Lett. 584:191–196.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen SQ, Wang PJ, Ten GJ, Zhan W, Li MH
and Zang FC: Role of myo-inositol by magnetic resonance
spectroscopy in early diagnosis of Alzheimer's disease in APP/PS1
transgenic mice. Dement Geriatr Cogn Disord. 28:558–566. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu YS, Xiong YF, Luo FQ and Min J:
Dexmedetomidine protects rats from postoperative cognitive
dysfunction via regulating the GABAB R-mediated
cAMP-PKA-CREB signaling pathway. Neuropathology. 39:30–38. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu Z, Wang D, Zhou Z, Chen Q, Zhang D,
Chen S, Jiang H, Jia C and Liu X: Dexmedetomidine attenuates renal
and myocardial Ischemia/Reperfusion injury in a dose-dependent
manner by inhibiting inflammatory response. Ann Clin Lab Sci.
49:31–35. 2019.PubMed/NCBI
|
|
19
|
Tucker LB, Fu AH and McCabe JT:
Performance of male and female C57BL/6J mice on motor and cognitive
tasks commonly used in pre-clinical traumatic brain injury
research. J Neurotrauma. 33:880–894. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shoham S, Linial M and Weinstock M:
Age-induced spatial memory deficits in rats are correlated with
specific brain region alterations in microglial morphology and gene
expression. J Neuroimmune Pharmacol. 14:251–262. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hsu WC, Yu CH, Kung WM and Huang KF:
Enhancement of matrix metalloproteinases 2 and 9 accompanied with
neurogenesis following collagen glycosaminoglycan matrix
implantation after surgical brain injury. Neural Regen Res.
13:1007–1012. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lord JM, Midwinter MJ, Chen YF, Belli A,
Brohi K, Kovacs EJ, Koenderman L, Kubes P and Lilford RJ: The
systemic immune response to trauma: An overview of pathophysiology
and treatment. Lancet. 384:1455–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Podjaski C, Alvarez JI, Bourbonniere L,
Larouche S, Terouz S, Bin JM, Lécuyer MA, Saint-Laurent O,
Larochelle C, Darlington PJ, et al: Netrin 1 regulates blood-brain
barrier function and neuroinflammation. Brain. 138:1598–1612. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Romeo HE, Tio DL and Taylor AN: Effects of
glossopharyngeal nerve transection on central and peripheral
cytokines and serum corticosterone induced by localized
inflammation. J Neuroimmunol. 136:104–111. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Le Maitre E, Revathikumar P, Estelius J
and Lampa J: Increased recovery time and decreased LPS
administration to study the vagus nerve stimulation mechanisms in
limited inflammatory responses. J Vis Exp. Mar 29–2017.doi:
10.3791/54890. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hoogland IC, Houbolt C, van Westerloo DJ,
van Gool WA and van de Beek D: Systemic inflammation and microglial
activation: Systematic review of animal experiments. J
Neuroinflammation. 12:1142015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gyoneva S, Davalos D, Biswas D, Swanger
SA, Garnier-Amblard E, Loth F, Akassoglou K and Traynelis SF:
Systemic inflammation regulates microglial responses to tissue
damage in vivo. Glia. 62:1345–1360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kang YC, Zhang L, Su Y, Li Y, Ren WL and
Wei WS: MicroRNA-26b regulates the microglial inflammatory response
in Hypoxia/Ischemia and affects the development of vascular
cognitive impairment. Front Cell Neurosci. 12:1542018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fakhoury M: Role of immunity and
inflammation in the pathophysiology of neurodegenerative diseases.
Neurodegener Dis. 15:63–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim DW, Lee JC, Cho JH, Park JH, Ahn JH,
Chen BH, Shin BN, Tae HJ, Seo JY, Cho JH, et al: Neuroprotection of
ischemic preconditioning is mediated by Anti-inflammatory, Not
Pro-inflammatory, Cytokines in the Gerbil hippocampus induced by a
subsequent lethal transient cerebral ischemia. Neurochem Res.
40:1984–1995. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Loftis JM, Valerio J, Taylor J, Huang E,
Hudson R, Taylor-Young P, Chang M, Ho SB, Dieperink E, Miranda JL
and Hauser P: S100B and inflammatory cytokine levels in blood as
potential markers of blood-brain barrier damage and psychiatric
impairment in comorbid hepatitis C viral infection and alcohol use
disorder. Alcohol Clin Exp Res. Jun 28–2018.doi: 10.1111/acer.13796
(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Farbood Y, Sarkaki A, Dianat M, Khodadadi
A, Haddad MK and Mashhadizadeh S: Ellagic acid prevents cognitive
and hippocampal long-term potentiation deficits and brain
inflammation in rat with traumatic brain injury. Life Sci.
124:120–127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Amor S, Peferoen LA, Vogel DY, Breur M,
van der Valk P, Baker D and van Noort JM: Inflammation in
neurodegenerative diseases-an update. Immunology. 142:151–166.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ma Y, Yu XY and Wang Y: Dose-related
effects of dexmedetomidine on immunomodulation and mortality to
septic shock in rats. World J Emerg Med. 9:56–63. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Suzumura A: Neuron-microglia interaction
in neuroinflammation. Curr Protein Pept Sci. 14:16–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bogie JF, Stinissen P and Hendriks JJ:
Macrophage subsets and microglia in multiple sclerosis. Acta
Neuropathol. 128:191–213. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li C, Chen T, Zhou H, Zhang C, Feng Y,
Tang F, Hoi MP, He C, Zheng Y and Lee SM: Schisantherin A
attenuates neuroinflammation in activated microglia: Role of Nrf2
activation through ERK phosphorylation. Cell Physiol Biochem.
47:1769–1784. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Peng J, Zhang P, Zheng H, Ren YQ and Yan
H: Dexmedetomidine reduces hippocampal microglia inflammatory
response induced by surgical injury through inhibiting NLRP3. Chin
J Traumatol. 22:161–165. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li JW, Zong Y, Cao XP and Tan L and Tan L:
Microglial priming in Alzheimer's disease. Ann Transl Med.
6:1762018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Daskalopoulos EP, Malliou F, Rentesi G,
Marselos M, Lang MA and Konstandi M: Stress is a critical player in
CYP3A, CYP2C, and CYP2D regulation: Role of adrenergic receptor
signaling pathways. Am J Physiol Endocrinol Metab. 303:E40–E54.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hoffman WE, Kochs E, Werner C, Thomas C
and Albrecht RF: Dexmedetomidine improves neurologic outcome from
incomplete ischemia in the rat. Reversal by the alpha 2-adrenergic
antagonist atipamezole. Anesthesiology. 75:328–332. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fang B, Li XQ, Bi B, Tan WF, Liu G, Zhang
Y and Ma H: Dexmedetomidine attenuates blood-spinal cord barrier
disruption induced by spinal cord ischemia reperfusion injury in
rats. Cell Physiol Biochem. 36:373–383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang D and Hong JH: Dexmedetomidine
modulates Histamine-induced Ca(2+) Signaling and Pro-inflammatory
cytokine expression. Korean J Physiol Pharmacol. 19:413–420. 2015.
View Article : Google Scholar : PubMed/NCBI
|