Introduction
Hepatocellular carcinoma (HCC) is a common form of
liver cancer associated with high mortality. It is estimated that
~600,000 new cases are diagnosed annually worldwide; HCC is
relatively common in Asia-Pacific countries and sub-Saharan Africa
(1). Hepatitis B virus (HBV)
infection is believed to be the most common cause of HCC
development, with the clinical course of HBV infection often
progressing from chronic hepatitis B (CHB) to liver cirrhosis (LC)
and then HCC (1). It is estimated
that ~2-10% of CHB patients develop LC, some of which subsequently
develop HCC; however, some HBV carriers can also spontaneously
eliminate the virus (2). HBV
infection is common in China due socio-economic factors. As a
consequence, the incidence of HCC in China is relatively high,
contributing to ~422,100 deaths annually (3).
The apolipoprotein B mRNA-editing catalytic
polypeptide-like 3 (APOBEC3, A3) gene cluster is located in
chromosome region 22q13.1 to q13.2 (4). This gene cluster encodes seven
proteins, including A3A, A3B, A3C, A3DE, A3F, A3G and A3H (5) and is reported to perform important
roles in various biological processes, including the innate immune
response to viral infections (4,6).
Among the seven protein family members, A3A and A3B are able to
restrict the infection of a broad range of viruses, including
parvovirus, HBV, hepatitis C virus, herpesvirus, human
papillomavirus and human immunodeficiency virus 1 (HIV-1) (7–13).
A3H is the most polymorphic member of the A3 subfamily, as it has
various single nucleotide polymorphism (SNP) combinations that
influence protein stability during resistance to HIV-1 infection
(14). Besides their role in viral
restriction, the dysregulation and hypermutation of A3 genes has
recently been linked to carcinogenesis (15). In particular, a 29.5 kb germline
deletion of A3A/B was associated with an increased risk of
various cancer types, including breast and ovarian cancer. However,
the effect has been inconsistent in different populations and for
different types of cancer. For example, it has been suggested that
the deletion of A3A/B was associated with an increased risk
of breast cancer in European women (16), Chinese women (17) and southeast Iranian women (18). However, this association was not
observed in Swedish (19) or
Moroccan (20) populations, or in
the general European population (21). Few studies have investigated the
association between A3H polymorphisms and cancer risk. Zhu
et al (22) reported that
the T allele of the rs139293 A3H SNP was associated with
reduced lung cancer risk in a Chinese population; therefore,
further studies are required to confirm the associations between
A3A, A3B and A3H polymorphisms and HCC risk.
The present study evaluated the associations between
the SNPs of A3A, A3B and A3H, and the development of
chronic HBV and HBV-related HCC in a Han Chinese population.
Materials and methods
Study population
Between January 2012 and July 2016, a total of 654
patients from the First Hospital of Jilin University were enrolled
in the present study, including 104 patients with CHB, 265 patients
with HBV-related LC and 285 patients with HBV-related HCC. The
criteria used to diagnose CHB, HBV-related LC and HBV-related HCC
have been defined previously (23). Hepatitis A-, C-, D- or E-positive
patients and those with HIV were excluded. In addition, patients
who had suffered another organ malignancy in the past 5 years, had
combined autoimmune diseases, or had other liver diseases, such as
intra- and extra-hepatic bile duct stones, alcoholic liver diseases
and hemorrhagic liver diseases, were also excluded. General
characteristics, including gender, age, smoking history, drinking
history, HBV infection history and treatment history, were gathered
using a standardized questionnaire. Whole blood (5 ml) was
collected from veins of each patient within 48 h of hospital
admission and their hepatitis B profile was compiled, including
hepatitis B e antigen (HBeAg), hepatitis B e antibody (HBeAb),
anti-hepatitis B core antigen (HBc), anti-HBe, hepatitis C, HBV DNA
quantification, liver function, renal function, α-fetoprotein,
blood lipids, blood glucose, blood routine, coagulation routine and
abdominal color Doppler ultrasound (or liver computed tomography or
magnetic resonance imaging). Patients were also assessed using the
Child-Pugh score (24,25) and those with HCC underwent
Barcelona clinic liver cancer staging (26). The present study was approved by
the First Hospital Ethical Committee of Jilin University and
written informed consent was obtained from all participants.
SNP selection and genotyping
A3A, A3B and A3H SNPs were selected
from the functional regions of the exon, promoter and untranslated
regions (UTRs) by GeneView (27)
based on Hapmap (https://www.genome.gov/10001688/international-hapmap-project)
and the 1,000 Genomes database (http://www.internationalgenome.org/), with a minor
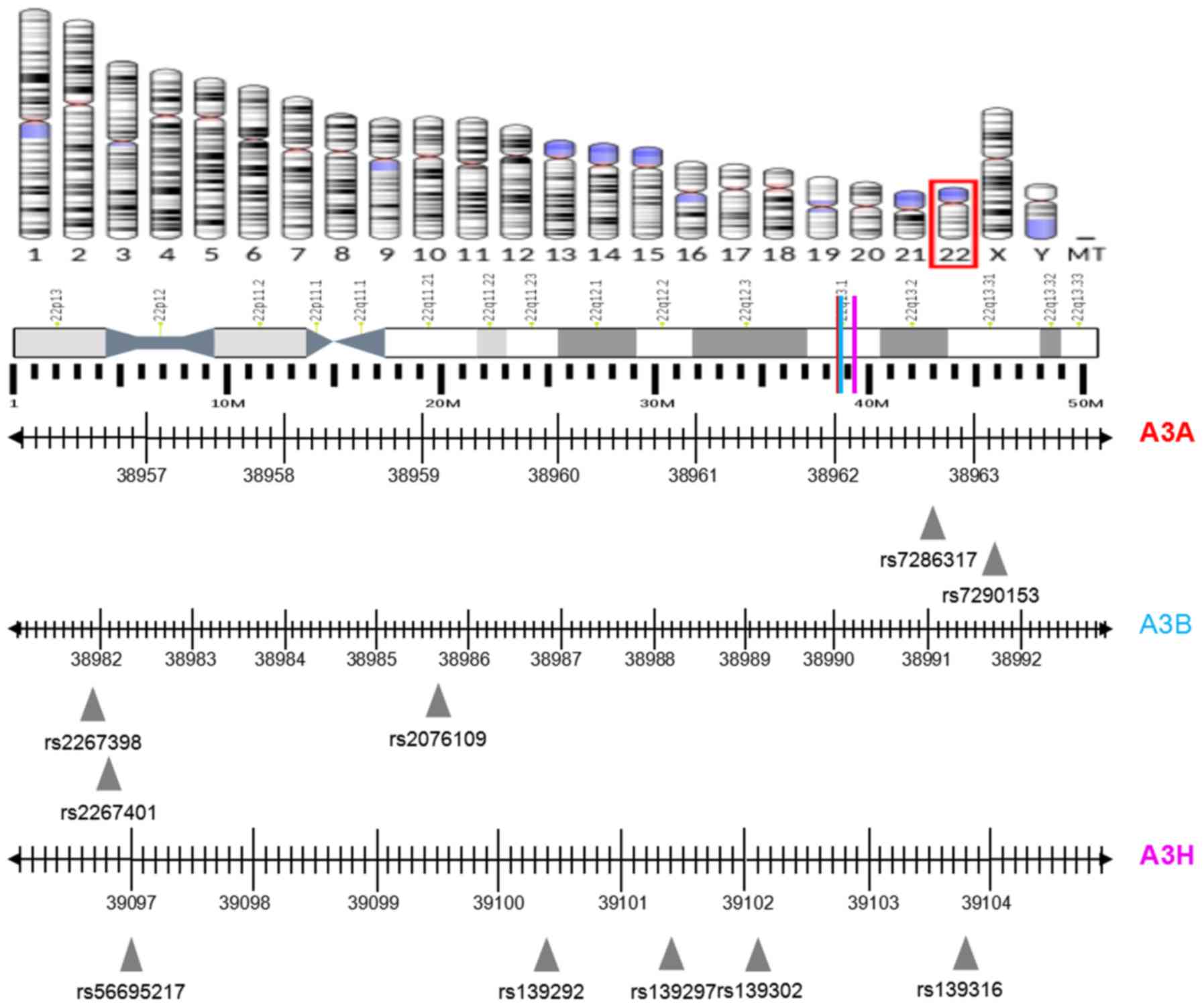
allele frequency of >10%. The SNPs rs7286317 and rs7290153 were
selected for A3A since they are located in the
microRNA-binding site of the 3´UTR. The SNPs rs2267398 and
rs2267401, located in the transcription factor-binding site of the
promoter region, were selected for A3B due to their
potential roles in gene transcription, while the SNP rs2076109 was
selected as it is a missense mutation that may regulate gene
function by altering the protein structure. The SNPs rs56695217,
rs139302, rs139297, rs139316 and rs139292 were selected for
A3H because rs56695217 is located in the transcription
factor-binding site, and the others are missense mutations.
Haplotype analysis was performed using Haploview version 4.2
(http://www.broad.mit.edu/mpg/haploview) with rs2076109
(A3B), rs139297 (A3H), rs139302 (A3H) and
rs139316 (A3H) tag-SNPs. The locations of the A3A,
A3B and A3H genes and the selected SNPs are shown in
Fig. 1.
Genomic DNA was isolated from whole blood using a
blood genomic DNA kit (Sigma-Aldrich; Merck KGaA), according to the
manufacturer's instructions. SNP genotyping was performed using a
MassArray system (Sequenom), according to the manufacturer's
protocol. All SNP primers were designed using Assay Designer
(http://assay.archerdx.com/, version 3.2;
Table I).
 | Table I.Primer sequences for SNP
genotyping. |
Table I.
Primer sequences for SNP
genotyping.
| Gene | SNPs | Primer
sequence | Annealing
temperature (°C) |
|---|
| APOBEC3A | rs7286317 | F:
5′-ACGTTGGATGGTCAGGAGATCGAGACCATC-3′ | 45.1 |
|
|
| R:
5′-ACGTTGGATGCACGCCTGGCTAATTTTTTG-3′ |
|
|
| rs7290153 | F:
5′-ACGTTGGATGGGAAGATTCTTAATTTTGTG-3′ | 45.5 |
|
|
| R:
5′-ACGTTGGATGGATTATGCTCAATATTCTCAG-3′ |
|
| APOBEC3B | Rs2267398 | F:
5′-ACGTTGGATGTTCTCCCTTCCTTGGTGTCG-3′ | 46.1 |
|
|
| R:
5′-ACGTTGGATGATGCGTCCCCTCTTCCAAC-3′ |
|
|
| rs2267401 | F:
5′-ACGTTGGATGTCTCTCAGCTGGGTCTGGA-3′ | 52.4 |
|
|
| R:
5′-ACGTTGGATGGGACCCAACGGAATTGCAAA-3′ |
|
|
| rs2076109 | F:
5′-ACGTTGGATGAGAGGAAGCACATTTCTGCG-3′ | 49.6 |
|
|
| R:
5′-ACGTTGGATGTGCTCCCCCTCTCAGAGCAT-3′ |
|
| APOBEC3H | Rs56695217 | F:
5′-ACGTTGGATGCCTTGTAATTTGCCCACCTC-3′ | 47.0 |
|
|
| R:
5′-ACGTTGGATGAAGAACAAAGGCCAGATGCG-3′ |
|
|
| Rs139292 | F:
5′-ACGTTGGATGTCAGCTGGTAACACAAGAGG-3′ | 58.2 |
|
|
| R:
5′-ACGTTGGATGAGCCGAAACATTCCGCTTAC-3′ |
|
|
| Rs139297 | F:
5′-ACGTTGGATGTTGCACCAGTGGTAGTACAG-3′ | 48.9 |
|
|
| R:
5′-ACGTTGGATGGCTGGTTGACTTCATCAAGG-3′ |
|
|
| Rs139302 | F:
5′-ACGTTGGATGCAGGACAGTGCCTCACCTT-3′ | 49.1 |
|
|
| R:
5′-ACGTTGGATGCCTTCAACCCCTATAAGATG-3′ |
|
|
| Rs139316 | F:
5′-ACGTTGGATGCCAGGGAAAGTCATCTTGAG-3′ | 46.7 |
|
|
| R:
5′-ACGTTGGATGAAGAAGTTTGCAGCTTGGAC-3′ |
|
Statistical analysis
All data were analyzed using SPSS version 21.0 (IBM
Corp.). Continuous variables are expressed as the mean ± standard
deviation or as the median and the interquartile range (25 and
75%). Categorical variables are expressed as a percentage (%).
Differences among multiple groups were compared using analysis of
variance and the least significant difference multiple comparisons
test. Haplotype analysis was performed using Unphased version 3.1.4
(28). The two-sided χ2
test or Fisher's exact test was used to compare allele
distributions. Multivariate logistic regression analysis was
performed to calculate odds ratios and 95% confidence intervals
after adjusting the factors of smoking, drinking and gender
differences. P<0.05 was considered to indicate a statistically
significant difference.
Results
General characteristics of the study
population
The main general and clinical characteristics of the
study population are summarized in Table II. No statistical differences were
observed between the sex, age, or the percentage of smokers and
alcohol consumers in the CHB and LC patient groups (P<0.05). In
comparison, the median age and percentages of smokers and alcohol
consumers were significantly higher for HCC patients than for CHB
patients (P=0.006, 0.013 and 0.008, respectively); however, no
significant difference was observed in their sex distributions.
Furthermore, no significant differences were observed in the sex,
age and percentage of alcohol consumers between the LC and HCC
patients (P<0.05), but the percentage of smokers differed
significantly (P<0.001).
| Table II.General and clinical characteristics
of study subjects. |
Table II.
General and clinical characteristics
of study subjects.
|
Characteristics | CHB n=104 |
P-valuea | LC n=265 |
P-valueb | HCC n=287 |
P-valuec | Reference
ranges |
|---|
| Sex (M/F) | 84/20 | 0.823 | 210/55 | 0.063 | 246/41 | 0.286 | – |
| Aged | 47 (43,53) | 0.368 | 49 (41.5,56) | 0.901 | 50 (46,56) | 0.006 | – |
|
Smokinge |
| 0.862 |
| <0.001 |
| 0.013 | – |
| Have
ever smoked | 37 (35.6) |
| 91 (34.3) |
| 144 (50.2) |
|
|
| Have
never smoked | 67 (64.4) |
| 174 (65.7) |
| 143 (49.8) |
|
|
| Alcohol
consumptione |
| 0.142 |
| 0.123 |
| 0.008 | – |
| Have
ever consumed alcohol | 28 (26.9) |
| 93 (35.1) |
| 120 (41.8) |
|
|
| Have
never consumed alcohol | 76 (73.1) |
| 172 (64.9) |
| 167 (58.2) |
|
|
| Serum
HBV-DNAe |
| 0.002 |
| 0.004 |
| 0.107 | – |
|
Positive | 98 (94.2) |
| 211 (79.6) |
| 254 (88.5) |
|
|
|
Negative | 6 (5.8) |
| 55 (20.4) |
| 33 (11.5) |
|
|
| HBV load, log10
(IU/ml)d | 6.1 (4.2, 7.3) | <0.001 | 4.6 (2.1, 6.3) | 0.565 | 4.6 (3.1, 6.0) | <0.001 | 1.3–8.2 |
| HBeAge |
| 0.814 |
| 0.359 |
| 0.295 | – |
|
Positive | 43 (48.9) |
| 99 (46.7) |
| 100 (42.2) |
|
|
|
Negative | 45 (51.1) |
| 113 (53.3) |
| 137 (57.8) |
|
|
| ALT
(U/l)d | 172 (53,
496.5) | <0.001 | 42 (24, 86) | 0.715 | 43.5 (27.8,
69.3) | <0.001 | 13.0–35.0 |
| AST
(U/l)d | 99 (39.5,
249.5) | <0.001 | 47 (31, 90) | <0.001 | 62.0
(38.0,110.0) | 0.005 | 7.0–40.0 |
| ALP
(U/l)d | 87 (67,122.8) | 0.724 | 89(68,125.5) | <0.001 | 129.5
(86.0,190.5) | <0.001 | 50.0–135.0 |
| GGT
(U/l)d | 93 (38.3,
161.8) | <0.001 | 49.5 (27,
100.8) | <0.001 | 112.5 (51.3,
254.3) | 0.018 | 7.0–45.0 |
| Prealbumin
(g/l)d | 0.16 (0.13,
0.20) | <0.001 | 0.12 (0.09,
0.16) | 0.192 | 0.13 (0.08,
0.17) | <0.001 | 0.18–0.39 |
| Albumin
(g/l)d | 37.5 (32.0,
41.2) | <0.001 | 30.5 (25.3,
36.0) | <0.001 | 33.1 (28.3,
37.3) | <0.001 | 40.0–55.0 |
| Total bilirubin
(µmol/l)d | 19.6 (13.3,
48.0) | 0.035 | 27.6 (16.2,
61.4) | 0.314 | 25.9 (16.5,
44.2) | 0.101 | 0.0–8.6 |
| Cholinesterase
(U/l)d | 6,019 (4,343,
8,281) | <0.001 | 3,289 (2,356,
4,744) | 0.012 | 3,918.0 (2,488.3,
5,762.0) | <0.001 | 4,300–12,000 |
| Platelet count
(×109/l)d | 145 (117, 188) | <0.001 | 77 (53, 121) | <0.001 | 118.5 (78.8,
172.0) | <0.001 | 100–300 |
No significant differences were observed between the
HBeAg positive rate or alkaline phosphatase (ALP) level of the CHB
and LC patients. However, the serum HBV-DNA positive rate, HBV
load, and alanine transaminase (ALT), aspartate transaminase (AST)
and glutamyl transpeptidase (GGT) levels of the CHB patients were
significantly higher compared with those of the LC patients
(P<0.05), suggesting that hepatocellular damage was more severe
in CHB patients. Furthermore, the prealbumin, albumin and
cholinesterase levels, and the platelet count were all
significantly higher in CHB patients compared with LC patients
(P<0.05), while the total bilirubin level was significantly
lower in CHB patients compared with LC patients (P<0.05). The
HBeAg positive rate, HBV load, ALT level, prealbumin level and
total bilirubin level did not differ significantly between LC and
HCC patients (P<0.05). Additionally, the levels of AST, ALP,
GGT, albumin and cholinesterase, and the platelet count were all
significantly lower in LC patients compared with HCC patients
(P<0.05), suggesting that the LC patients had a lower level of
hepatocellular damage than the HCC patients.
Associations between genotype and
allele frequency in A3A, A3B and A3H SNPs
The genotype and allele frequency of the A3A
polymorphisms in CHB patients and healthy individuals are displayed
in Table III. No significant
associations were detected between the genotype and allele
frequency of the two A3A SNPs (rs7286317 and rs7290153) and
chronic hepatitis B progression or HCC occurrence (P<0.05).
Furthermore, as shown in Tables
IV and V, no significant
associations were observed between the three A3B SNPs
(rs2267398, rs2267401 and rs2076109) or the five A3H SNPs
(rs56695217, rs139302, rs139297, rs139316 and rs139292) and chronic
hepatitis B progression or HCC occurrence (P<0.05).
| Table III.Genotype and allele frequencies of
two SNPs of APOBEC3A. |
Table III.
Genotype and allele frequencies of
two SNPs of APOBEC3A.
| A, Rs7286317
genotype and allele |
|---|
|
|---|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|---|
|
|
|
|
|
|---|
| SNP | N (%) | OR (95%Cl) |
P-valuea | N
(%) | OR (95%Cl) |
P-valueb | N (%) | OR (95%Cl) |
P-valuec |
|---|
| Number
detected | n=102 |
|
| n=265 |
|
| n=285 |
|
|
| AA | 75 (73.5) | 1 |
| 197 (74.3) | 1 |
| 200 (70.2) | 1 |
|
| AG | 27 (26.5) | 0.95
(0.56,1.60) | 0.85 | 68 (25.7) | 1.30
(0.89,191) | 0.18 | 85 (29.8) | 1.24
(0.74,2.08) | 0.42 |
| A Allele | 177 (86.8) | 1 |
| 462 (87.2) |
|
| 485 (85.1) | 1 |
|
| G Allele | 27 (13.2) | 0.96
(0.60,1.56) | 0.88 | 68 (12.8) | 0.84
(0.60,1.18) | 0.32 | 85 (14.9) | 0.87
(0.55,1.39) | 0.56 |
|
| B, Rs7290153
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=97 |
|
| n=246 |
|
| n=267 |
|
|
| CC | 78 (80.4) | 1 |
| 198 (80.5) | 1 |
| 217 (81.3) | 1 |
|
| CT | 14 (14.4) | 0.88
(0.44,1.75) | 0.72 | 31 (12.6) | 0.83
(0.48,1.47) | 0.52 | 30 (11.2) | 0.78
(0.39,1.57) | 0.48 |
| TT | 5 (5.2) | 1.30
(0.46,3.70) | 0.62 | 17 (6.9) | 1.02
(0.51,2.04) | 0.95 | 20 (7.5) | 1.41
(0.51,3.92) | 0.51 |
| CT+TT | 19 | 0.99
(0.55,1.80) | 0.97 | 48 | 0.90
(0.57,1.41) | 0.65 | 50 | 0.95
(0.52,1.73) | 0.86 |
| C Allele | 170 (87.6) | 1 |
| 427 (86.8) | 1 |
| 464 (86.9) | 1 |
|
| T Allele | 24 (12.4) | 1.0
(0.65,1.78) | 0.77 | 65 (13.2) | 1.01
(0.70,1.45) | 0.96 | 70 (13.1) | 0.94
(0.57,1.54) | 0.79 |
| Table IV.Genotype and allele frequencies of
three SNPs of APOBEC3B. |
Table IV.
Genotype and allele frequencies of
three SNPs of APOBEC3B.
| A, Rs2267398
genotype and allele |
|---|
|
|---|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|---|
|
|
|
|
|
|---|
| SNP | N (%) | OR (95%Cl) |
P-valuea | N (%) | OR (95%Cl) |
P-valueb | N (%) | OR (95%Cl) |
P-valuec |
|---|
| Number
detected | n=96 |
|
| n=256 |
|
| n=272 |
|
|
| CC | 28 (29.2) | 1 |
| 90 (35.2) | 1 |
| 93 (34.2) | 1 |
|
| CT | 50 (52.1) | 0.75
(0.44,1.29) | 0.31 | 125 (48.8) | 1.11
(0.75,1.63) | 0.61 | 135 (49.6) | 0.85
(0.50,1.47) | 0.57 |
| TT | 18 (18.8) | 0.71
(0.35,1.44) | 0.34 | 41 (16.0) | 1.15
(0.68,1.94) | 0.61 | 44 (16.2) | 0.77
(0.38,1.56) | 0.47 |
| CT+TT | 68 | 0.74
(0.44,1.24) | 0.26 | 166 | 1.12
(0.77,1.61) | 0.56 | 179 | 0.83
(0.50,1.39) | 0.49 |
| C Allele | 106 (55.2) | 1 |
| 305 (59.6) | 1 |
| 321 (59.0) | 1 |
|
| T Allele | 86 (44.8) | 0.84
(0.60,1.17) | 0.30 | 207 (40.4) | 0.98
(0.76,1.25) | 0.85 | 223 (41.0) | 1.17
(0.84,1.63) | 0.36 |
|
| B, Rs2267401
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=102 |
|
| n=265 |
|
| n=284 |
|
|
| GG | 25 (24.5) | 1 |
| 58 (21.9) |
|
| 63 (22.2) |
|
|
| GT | 29 (28.4) | 0.93
(0.49,1.77) | 0.82 | 65 (24.5) | 0.89
(0.53,1.46) | 0.62 | 61 (21.5) | 0.88
(0.47,1.69) | 0.69 |
| TT | 48 (47.1) | 1.25
(0.70,2.22) | 0.45 | 142 (53.6) | 1.00
(0.65,1.54) | 0.99 | 160 (56.3) | 1.29
(0.72,2.29) | 0.39 |
| GT+TT | 77 | 1.13
(0.66,1.93) | 0.67 | 207 | 0.97
(0.64,1.46) | 0.87 | 221 | 1.13
(0.66,1.93) | 0.65 |
| G Allele | 79 (38.7) | 1 |
| 181 (34.2) | 1 |
| 187 (32.9) | 1 |
|
| T Allele | 125 (61.3) | 1.22
(0.87,1.70) | 0.25 | 349 (65.8) | 0.95
(0.74,1.22) | 0.67 | 381 (67.1) | 1.00
(0.71,1.42) | 0.98 |
| C, Rs2076109
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=94 |
|
| n=234 |
|
| n=246 |
|
|
| AA | 24 (25.5) | 1 |
| 57 (24.4) | 1 |
| 54 (22.0) | 1 |
|
| AG | 20 (21.3) | 1.30
(0.65,2.61) | 0.46 | 62 (26.5) | 1.12
(0.67,1.89) | 0.67 | 64 (26.0) | 1.70
(0.83,3.50) | 0.15 |
| GG | 50 (53.2) | 0.95
(0.53,1.71) | 0.87 | 115 (49.1) | 0.13
(0.72,1.79) | 0.59 | 128 (52.0) | 1.21
(0.67,2.21) | 0.53 |
| AG+GG | 70 | 1.05
(0.61,1.83) | 0.86 | 177 | 1.13
(0.73,1.74) | 0.58 | 192 | 1.03
(0.63,1.69) | 0.89 |
| A Allele | 68 (36.2) | 1 |
| 176 (37.6) | 1 |
| 172 (35.0) | 1 |
|
| G Allele | 120 (63.8) | 0.94
(0.66,1.37) | 0.73 | 292 (62.4) | 0.89
(0.69,1.16) | 0.39 | 320 (65.0) | 1.35
(0.76,2.39) | 0.31 |
| Table V.Genotype and allele frequencies of
five SNPs of APOBEC3H. |
Table V.
Genotype and allele frequencies of
five SNPs of APOBEC3H.
| A, Rs56695217
genotype and allele |
|---|
|
|---|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|---|
|
|
|
|
|
|---|
| SNP | N (%) | OR (95%Cl) |
P-valuea | N (%) | OR (95%Cl) |
P-valueb | N (%) | OR (95%Cl) |
P-valuec |
|---|
| Number
detected | n=89 |
|
| n=235 |
|
| n=241 |
|
|
| CC | 11 (12.4) | 1 |
| 29 (12.3) |
|
| 26 (10.8) |
|
|
| CG | 76 (85.4) | 1.03
(0.49,2.17) | 0.94 | 203 (86.4) | 1.26
(0.71,2.25) | 0.43 | 214 (88.8) | 1.17
(0.54,2.56) | 0.69 |
| GG | 2 (2.2) | 0.60
(0.09,4.13) | 0.60 | 3 (1.3) | 0.34
(0.03,3.64) | 0.36 | 1 (0.4) | 0.11
(0.01,1.37) | 0.08 |
| CG+GG | 78 | 1.02
(0.48,2.15) | 0.96 | 206 | 1.25
(0.70,2.22) | 0.45 | 215 | 1.14
(0.53,2.47) | 0.74 |
| C Allele | 98 (55.1) | 1 |
| 261 (55.5) | 1 |
| 266 (55.2) | 1 |
|
| G Allele | 80 (44.9) | 0.98
(0.69,1.39) | 0.91 | 209 (44.5) | 0.99
(0.76,1.27) | 0.92 | 216 (44.8) | 1.00
(0.71,1.42) | 0.98 |
|
| B, Rs139292
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=95 |
|
| n=247 |
|
| n=250 |
|
|
| DEL | 42 (44.2) | 1 |
| 111 (44.9) | 1 |
| 112 (44.8) | 1 |
|
| CAA.DEL | 53 (55.8) | 0.97
(0.60,1.57) | 0.90 | 134 (54.3) | 1.04
(0.72,1.49) | 0.84 | 137 (54.8) | 1.03
(0.91,0.63) | 0.91 |
| CAA | 0 (0.0) | 0.73
(0.66,0.80) | 1.00 | 2 (0.8) | 0.50
(0.60,7.09) | 0.68 | 1 (0.4) | 0.73
(0.66,0.80) | 1.00 |
| DEL+CAA | 53 | 0.99
(0.61,1.60) | 0.95 | 136 | 1.03
(0.72,1.48) | 0.86 | 138 | 1.03
(0.63,1.69) | 0.89 |
| DEL Allele | 137 (72.1) | 1 |
| 356 (72.1) | 1 |
| 361 (72.2) | 1 |
|
| CAA Allele | 53
(27.9) | 0.69
(1.00,1.46) | 1.00 | 138 (27.9) | 0.99
(0.75,1.31) | 1.00 | 139 (27.8) | 0.99
(0.69,1.45) | 1.00 |
|
| C, Rs139297
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=102 |
|
| n=263 |
|
| n=280 |
|
|
| CC | 41 (40.2) | 1 |
| 115 (43.7) | 1 |
| 118 (42.1) | 1 |
|
| CG | 16 (15.7) | 0.84
(0.42,1.68) | 0.63 | 38 (14.4) | 0.95
(056,1.62) | 0.85 | 36 (12.9) | 0.85
(0.42,1.72) | 0.65 |
| GG | 45 (44.1) | 0.87
(0.52,1.44) | 0.58 | 110 (41.8) | 1.78
(0.81,1.70) | 0.39 | 126 (45.0) | 1.07
(0.64,1.79) | 0.79 |
| CG+GG | 61 | 0.86
(0.54,1.38) | 0.53 | 143 | 1.12
(0.79,1.58) | 0.53 | 174 | 1.01
(0.63,1.63) | 0.97 |
| C Allele | 98 (49.4) | 1 |
| 268 (49.8) |
|
| 272 (77.0) | 1 |
|
| G Allele | 106 (50.6) | 0.89
(0.64,1.23) | 0.48 | 258 (50.2) | 1.15
(0.89,1.48) | 0.28 | 288 (23.0) | 0.98
(0.71,1.35) | 0.90 |
|
| D, Rs139302
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=89 |
|
| n=229 |
|
| n=255 |
|
|
| CC | 34 (38.2) | 1 |
| 86 (37.6) | 1 |
| 81 (31.8) | 1 |
|
| CG | 20 (22.5) | 1.1
(0.57,2.11) | 0.79 | 56 (24.5) | 1.38
(0.86,2.20) | 0.18 | 74 (29.0) | 1.53
(0.80,2.94) | 0.20 |
| GG | 35 (39.3) | 0.98
(0.55,1.72) | 0.93 | 87 (30.8) | 1.25
(0.82,1.91) | 0.30 | 100 (39.2) | 1.31
(0.73,2.33) | 0.37 |
| CG+GG | 55 | 1.02
(0.61,1.70) | 0.94 | 143 | 1.30
(0.89,1.90) | 0.18 | 174 | 1.39
(0.83,2.24) | 0.21 |
| C Allele | 88 (49.4) | 1 |
| 228 (49.8) | 1 |
| 236 (46.3) | 1 |
|
| G Allele | 90 (50.6) | 0.99
(0.70,1.40) | 0.94 | 230 (50.2) | 0.87
(0.68,1.12) | 0.28 | 274 (53.7) | 0.88
(0.63,1.24) | 0.47 |
|
| E, Rs139316
genotype and allele |
|
|
| CHB patients
(n=104) | LC patients
(n=265) | HCC patients
(n=287) |
|
|
|
|
|
| SNP | N (%) | OR
(95%Cl) |
P-valuea | N (%) | OR
(95%Cl) |
P-valueb | N (%) | OR
(95%Cl) |
P-valuec |
|
| Number
detected | n=103 |
|
| n=263 |
|
| n=281 |
|
|
| CC | 17 (16.5) | 1 |
| 43 (16.3) | 1 |
| 37 (13.2) | 1 |
|
| CT | 46 (44.7) | 1.02
(0.52,1.97) | 0.96 | 117 (44.5) | 0.26
(0.75,2.10) | 0.39 | 133 (47.3) | 1.36
(0.69,2.69) | 0.38 |
| TT | 40 (38.8) | 1.01
(0.51,2.00) | 0.97 | 103 (39.2) | 1.24
(0.73,2.10) | 0.42 | 111 (39.5) | 1.40
(0.69,2.77) | 0.37 |
| CT+TT | 86 | 1.01
(0.54,1.89) | 0.97 | 220 | 1.25
(0.77,2.03) | 0.37 | 244 | 1.37
(0.72,2.60) | 0.34 |
| C Allele | 80 (38.3) | 1 |
| 203 (38.6) | 1 |
| 207 (36.8) | 1 |
|
| T Allele | 126 (61.2) | 1.01
(0.73,1.41) | 0.95 | 323 (61.4) | 0.93
(0.73,1.86) | 0.55 | 355 (63.2) | 1.09
(0.78,1.51) | 0.61 |
Haplotype analysis of A3A, A3B and
A3H
Haplotype analysis was also performed on the two
A3A SNPs, three A3B SNPs and five A3H SNPs
using Unphased version 3.1.4. No haplotypes were found for the two
A3A SNPs or five A3H SNPs (data not shown). The
distribution of the A3B haplotype
rs2267398-rs2267401-rs2076109 was significantly different between
the LC and HCC groups (Table VI).
The C-G-G haplotype was used as a reference, with the results
showing that the mutant C-T-A, C-T-G, T-G-G and T-T-G haplotypes of
rs2267398-rs2267401-rs2076109 were associated with a lower risk of
HCC compared with the reference haplotype (Table VII).
| Table VI.Distributions of SNPs of
apolipoprotein B mRNA-editing catalytic polypeptide-like 3B in the
different groups. |
Table VI.
Distributions of SNPs of
apolipoprotein B mRNA-editing catalytic polypeptide-like 3B in the
different groups.
| Groups | SNPs | χ2 | df | P-value |
|---|
| CHB vs. LC |
rs2267398-rs2267401 | 3.87 | 3 | 0.276 |
|
|
rs2267398-rs2076109 | 5.25 | 3 | 0.153 |
|
|
rs2267401-rs2076109 | 2.51 | 3 | 0.472 |
|
|
rs2267398-rs226740-rs2076109 | 7.33 | 6 | 0.291 |
| CHB vs. HCC |
rs2267398-rs2267401 | 1.01 | 3 | 0.798 |
|
|
rs2267398-rs2076109 | 0.149 | 3 | 0.985 |
|
|
rs2267401-rs2076109 | 0.405 | 3 | 0.939 |
|
|
rs2267398-rs2267401-rs2076109 | 1.764 | 6 | 0.940 |
| LC vs. HCC |
rs2267398-rs2267401 | 8.210 | 3 | 0.042 |
|
|
rs2267398-rs2076109 | 1.368 | 3 | 0.713 |
|
|
rs2267401-rs2076109 | 3.278 | 3 | 0.351 |
|
|
rs2267398-rs2267401-rs2076109 | 14.25 | 6 | 0.027 |
| Table VII.Analysis of the
rs2267398-rs2267401-rs2076109 haplotypes of apolipoprotein B
mRNA-editing catalytic polypeptide-like 3B in patients with LC and
HCC. |
Table VII.
Analysis of the
rs2267398-rs2267401-rs2076109 haplotypes of apolipoprotein B
mRNA-editing catalytic polypeptide-like 3B in patients with LC and
HCC.
| Haplotype | LC (%) | HCC (%) | OR (95%CI) | P-value |
|---|
| C-G-G | 8 (1.1) | 9 (5.6) | 1 |
|
| C-T-A | 254 (33.8) | 56 (34.6) | 0.19 (0.07,
0.53) | <0.001 |
| C-T-G | 191 (25.4) | 35 (21.6) | 0.16 (0.06,
0.45) | <0.001 |
| T-G-A | 2 (0.3) | 0 (0.0) | 0.47
(0.28,0.78) | 0.474 |
| T-G-G | 246 (32.7) | 50 (30.9) | 0.18 (0.07,
0.49) | <0.001 |
| T-T-A | 25 (3.3) | 10 (6.2) | 0.36
(0.11,1.18) | 0.087 |
| T-T-G | 26 (3.5) | 2 (1.2) | 0.07 (0.01,
0.38) | 0.001 |
Discussion
It is estimated that ~55% of HCC cases are
associated with CHB (29,30). Members of the A3 protein family
have been reported to edit the HBV genome and reduce HBV
replication in vivo and in vitro (31,32).
However, the effects of the SNPs of A3 genes have not yet been
evaluated in a Chinese population. To the best of the authors'
knowledge, the present study is the first to investigate the
association between A3A, A3B and A3H SNPs and the
development of CHB and HBV-related HCC in a Chinese population.
There were two major findings of the present study: i) The
rs7286317 and rs7290153 SNPs of A3A, and the rs56695217,
rs139292, rs139297, rs139302 and rs139316 SNPs of A3H, had
no relationship with CHB progression or HCC development; and ii)
the rs2267398, rs2267401 and rs2076109 SNPs of A3B may not
affect the likelihood of CHB progression or HCC development.
However, the C-T-A, C-T-G, T-G-G and T-T-G haplotypes of
rs2267398-rs2267401-rs2076109 were associated with a lower risk of
HCC development than the reference haplotype C-G-G.
APOBEC cytosine deaminases are known to confer
innate immunity against retroviruses by generating lethal
hypermutations in viral genomes (33). Köck and Blum (31) assessed the ability of A3G,
A3C and A3H to edit HBV genomes, finding that each gene
could edit HBV DNA and that each protein was likely to contribute
(to varying degrees) to genome modification in human liver cells.
Previously, it was demonstrated that the A3G rs8177832 SNP
was associated with a decreased risk of CHB infection and HCC,
while the rs2011861 SNP was associated with an increased risk of
HCC (23). Furthermore, it has
been shown that A3A is an efficient HBV DNA editor, while
A3A and A3B serve crucial roles in inducing the
degradation of HBV covalently closed circular DNA (34). Therefore, it was speculated that
these three genes may be associated with disease progression
following HBV infection.
The present study analyzed the association between
A3A, A3B and A3H SNPs and the progression of HBV
infection. A total of 654 patients were included in the study,
consisting of 104 patients with CHB, 265 patients with HBV-related
LC and 285 patients with HBV-related HCC. However, the results
demonstrated that the SNPs of these three genes were not associated
with disease progression following HBV infection. Haplotype
analysis suggested that the C-T-A, C-T-G, T-G-G and T-T-G
haplotypes of rs2267398-rs2267401-rs2076109 were associated with a
lower risk of HCC compared with the reference haplotype C-G-G. It
was hypothesized that this may be due to the linkage between
different functional genes. Previous studies have shown that
APOBEC-specific mutations are common in tumor genomes (35,36)
and that the expression level of APOBEC mRNA is positively
correlated with the APOBEC-specific mutation rate (37). In vitro, A3B has been shown
to promote the proliferation of the hepatoma cell line HepG2 by
upregulating the expression of heat shock protein 1 (38). Therefore, A3B may be the
predominant APOBEC-specific mutation-inducing gene in the
development of primary liver cancer. Notably, clinical data have
demonstrated that the deletion of ~29.5 kb between A3A exon
5 and A3B exon 8 causes the loss of the entire A3B
coding region and increases the risk of HCC (39,40).
Furthermore, genome sequencing has revealed that A3B
deletion can increase the APOBEC-specific mutation rate in the
tumor genome (38). Consequently,
it has been hypothesized that A3B gene deletion may cause
the expression of A3AΔA3B (A3A after A3B deletion) to be more
stable and efficient (41,42) and that A3AΔA3B may be the
predominant mutagenic factor. Therefore, haplotype changes may
affect HCC occurrence by altering the gene expression and editing
the functions of A3A and A3B. However, the exact mechanisms by
which this occurs requires further investigation.
The present study had a number of limitations.
First, healthy controls were not enrolled in this study to evaluate
the effect of the A3A, A3B and A3H SNPs on
susceptibility to HBV infection. Second, some disease factors were
not considered in the present study, such as the age at which HBV
infection occurred, which is closely associated with the outcome of
HBV infection (43). However,
exact HBV infection age data are not available from most places in
China due to socioeconomic factors. According to previous studies,
~90% of infants infected perinatally become chronic carriers,
unless vaccinated at birth. The risk of CHB decreases to 30% in
children infected between ages 1 and 4 years, and to <5% in
persons infected as adults (44–46).
Therefore, most patients with CHB infection are likely to have been
infected in infancy. Since it was not possible to acquire the exact
infection age, the present study assumed the age of patients as the
length of infection. Therefore, age-matched patients with CHB,
HBV-related LC and HBV-related HCC were recruited. Third, the
associations were analyzed solely by statistical analysis and were
not validated experimentally. Therefore, further studies using
larger sample sizes from different populations alongside
experimental validation should be conducted to verify the results
of the present study.
In conclusion, the present study demonstrated that
there was no association between the rs7286317 and rs7290153 SNPs
of A3A, the rs2267398, rs2267401 and rs2076109 SNPs of
A3B, and the rs56695217, rs139292, rs139297, rs139302 and
rs139316 SNPs of A3H and CHB progression or HCC development.
However, the C-T-A, C-T-G, T-G-G and T-T-G haplotypes of
rs2267398-rs2267401-rs2076109 were associated with a lower risk of
HCC than the reference haplotype C-G-G.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Youth
Development Science Foundation of the First Hospital of Jilin
University (grant no. JDYY82017023).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XH, JN and PG conceived and designed the study. XH,
HX and XW acquired the data. HX and JN analyzed and interpreted the
data. JW performed the statistical analysis. XH drafted the
manuscript. JN and PG revised the manuscript for important
intellectual content. All authors given final approval of the
version to be published.
Ethics approval and consent to
participate
The present study was approved by the First Hospital
Ethical Committee of Jilin University. Written informed consent was
obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
APOBEC3
|
apolipoprotein B mRNA-editing
catalytic polypeptide-like 3
|
|
SNPs
|
single nucleotide polymorphisms
|
|
HBV
|
hepatitis B virus
|
|
HCC
|
hepatocellular carcinoma
|
|
CHB
|
chronic hepatitis B
|
|
LC
|
liver cirrhosis
|
|
HIV-1
|
human immunodeficiency virus 1
|
|
HBeAg
|
hepatitis B e antigen
|
|
HBeAb
|
hepatitis B e antibody
|
|
UTRs
|
untranslated regions
|
|
OR
|
odds ratio
|
|
CI
|
confidence intervals
|
References
|
1
|
Sawai H, Nishida N, Khor SS, Honda M,
Sugiyama M, Baba N, Yamada K, Sawada N, Tsugane S, Koike K, et al:
Genome-wide association study identified new susceptible genetic
variants in HLA class I region for hepatitis B virus-related
hepatocellular carcinoma. Sci Rep. 8:79582018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chu CM: Natural history of chronic
hepatitis B virus infection in adults with emphasis on the
occurrence of cirrhosis and hepatocellular carcinoma. J
Gastroenterol Hepatol. 15 (Suppl):E25–E30. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Henderson S and Fenton T: APOBEC3 genes:
Retroviral restriction factors to cancer drivers. Trends Mol Med.
21:274–284. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Teng B, Burant CF and Davidson NO:
Molecular cloning of an apolipoprotein B messenger RNA editing
protein. Science. 260:1816–1819. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Swanton C, McGranahan N, Starrett GJ and
Harris RS: APOBEC enzymes: Mutagenic fuel for cancer evolution and
heterogeneity. Cancer Discov. 5:704–712. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Doehle BP, Schafer A and Cullen BR: Human
APOBEC3B is a potent inhibitor of HIV-1 infectivity and is
resistant to HIV-1 Vif. Virology. 339:281–288. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Prasetyo AA, Sariyatun R, Revion o, Sari
Y, Hudiyon o, Haryati S, Adnan ZA, Harton o and Kageyama S: The
APOBEC3B deletion polymorphism is associated with prevalence of
hepatitis B virus, hepatitis C virus, Torque Teno virus, and
Toxoplasma gondii co-infection among HIV-infected individuals. J
Clin Virol. 70:67–71. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Berger G, Durand S, Fargier G, Nguyen XN,
Cordeil S, Bouaziz S, Muriaux D, Darlix JL and Cimarelli A:
APOBEC3A is a specific inhibitor of the early phases of HIV-1
infection in myeloid cells. PLoS Pathog. 7:e10022212011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakaya Y, Stavrou S, Blouch K, Tattersall
P and Ross SR: In vivo examination of mouse APOBEC3- and human
APOBEC3A- and APOBEC3G-mediated restriction of parvovirus and
herpesvirus infection in mouse models. J Virol. 90:8005–8012. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Minkah N, Chavez K, Shah P, Maccarthy T,
Chen H, Landau N and Krug LT: Host restriction of murine
gammaherpesvirus 68 replication by human APOBEC3 cytidine
deaminases but not murine APOBEC3. Virology. 454-455:215–226. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Henry M, Guetard D, Suspene R, Rusniok C,
Wain-Hobson S and Vartanian JP: Genetic editing of HBV DNA by
monodomain human APOBEC3 cytidine deaminases and the recombinant
nature of APOBEC3G. PLoS One. 4:e42772009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Warren CJ, Xu T, Guo K, Griffin LM,
Westrich JA, Lee D, Lambert PF, Santiago ML and Pyeon D: APOBEC3A
functions as a restriction factor of human papillomavirus. J Virol.
89:688–702. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
OhAinle M, Kerns JA, Li MM, Malik HS and
Emerman M: Antiretroelement activity of APOBEC3H was lost twice in
recent human evolution. Cell Host Microbe. 4:249–259. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gansmo LB, Romundstad P, Hveem K, Vatten
L, Nik-Zainal S, Lønning PE and Knappskog S: APOBEC3A/B deletion
polymorphism and cancer risk. Carcinogenesis. 39:118–124. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xuan D, Li G, Cai Q, Deming-Halverson S,
Shrubsole MJ, Shu XO, Kelley MC, Zheng W and Long J: APOBEC3
deletion polymorphism is associated with breast cancer risk among
women of European ancestry. Carcinogenesis. 34:2240–2243. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Long J, Delahanty RJ, Li G, Gao YT, Lu W,
Cai Q, Xiang YB, Li C, Ji BT, Zheng Y, et al: A common deletion in
the APOBEC3 genes and breast cancer risk. J Natl Cancer Inst.
105:573–579. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rezaei M, Hashemi M, Hashemi SM, Mashhadi
MA and Taheri M: APOBEC3 deletion is associated with breast cancer
risk in a sample of southeast iranian population. Int J Mol Cell
Med. 4:103–108. 2015.PubMed/NCBI
|
|
19
|
Gohler S, Da Silva Filho MI, Johansson R,
Enquist-Olsson K, Henriksson R, Hemminki K, Lenner P and Försti A:
Impact of functional germline variants and a deletion polymorphism
in APOBEC3A and APOBEC3B on breast cancer risk and survival in a
Swedish study population. J Cancer Res Clin Oncol. 142:273–276.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Marouf C, Gohler S, Filho MI, Hajji O,
Hemminki K, Nadifi S and Försti A: Analysis of functional germline
variants in APOBEC3 and driver genes on breast cancer risk in
Moroccan study population. BMC Cancer. 16:1652016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Klonowska K, Kluzniak W, Rusak B,
Jakubowska A, Ratajska M, Krawczynska N, Vasilevska D, Czubak K,
Wojciechowska M, Cybulski C, et al: The 30 kb deletion in the
APOBEC3 cluster decreases APOBEC3A and APOBEC3B expression and
creates a transcriptionally active hybrid gene but does not
associate with breast cancer in the European population.
Oncotarget. 8:76357–76374. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu M, Wang Y, Wang C, Shen W, Liu J, Geng
L, Cheng Y, Dai J, Jin G, Ma H, et al: The eQTL-missense
polymorphisms of APOBEC3H are associated with lung cancer risk in a
Han Chinese population. Sci Rep. 5:149692015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
He XT, Xu HQ, Wang XM, He XS, Niu JQ and
Gao PJ: Association between polymorphisms of the APOBEC3G gene and
chronic hepatitis B viral infection and hepatitis B virus-related
hepatocellular carcinoma. World J Gastroenterol. 23:232–241. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pons F, Varela M and Llovet JM: Staging
systems in hepatocellular carcinoma. HPB (Oxford). 7:35–41. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cholongitas E, Papatheodoridis GV, Vangeli
M, Terreni N, Patch D and Burroughs AK: Systematic review: The
model for end-stage liver disease-should it replace Child-Pugh's
classification for assessing prognosis in cirrhosis? Aliment
Pharmacol Ther. 22:1079–1089. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Selby LK, Tay RX, Woon WW, Low JK, Bei W,
Shelat VG, Pang TC and Junnarkar SP: Validity of the barcelona
clinic liver cancer and hong kong liver cancer staging systems for
hepatocellular carcinoma in singapore. J Hepatobiliary Pancreat
Sci. 24:143–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dohi H, Ishizuka M, Minoshima S and
Shimizu N: GeneView: Multi-language human gene mapping library with
a graphical user interface. Comput Appl Biosci. 9:459–464.
1993.PubMed/NCBI
|
|
28
|
Dudbridge F: Likelihood-based association
analysis for nuclear families and unrelated subjects with missing
genotype data. Hum Hered. 66:87–98. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cougot D, Neuveut C and Buendia MA: HBV
induced carcinogenesis. J Clin Virol. 34 (Suppl 1):S75–S78. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kock J and Blum HE: Hypermutation of
hepatitis B virus genomes by APOBEC3G, APOBEC3C and APOBEC3H. J Gen
Virol. 89:1184–1191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Suspene R, Guetard D, Henry M, Sommer P,
Wain-Hobson S and Vartanian JP: Extensive editing of both hepatitis
B virus DNA strands by APOBEC3 cytidine deaminases in vitro and in
vivo. Proc Natl Acad Sci USA. 102:8321–8326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chiu YL and Greene WC: The APOBEC3
cytidine deaminases: An innate defensive network opposing exogenous
retroviruses and endogenous retroelements. Annu Rev Immunol.
26:317–353. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lucifora J, Xia Y, Reisinger F, Zhang K,
Stadler D, Cheng X, Sprinzl MF, Koppensteiner H, Makowska Z, Volz
T, et al: Specific and nonhepatotoxic degradation of nuclear
hepatitis B virus cccDNA. Science. 343:1221–1228. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Roberts SA, Lawrence MS, Klimczak LJ,
Grimm SA, Fargo D, Stojanov P, Kiezun A, Kryukov GV, Carter SL,
Saksena G, et al: An APOBEC cytidine deaminase mutagenesis pattern
is widespread in human cancers. Nat Genet. 45:970–976. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alexandrov LB, Nik-Zainal S, Wedge DC,
Aparicio SA, Behjati S, Biankin AV, Bignell GR, Bolli N, Borg A,
Børresen-Dale AL, et al: Signatures of mutational processes in
human cancer. Nature. 500:415–421. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Burns MB, Temiz NA and Harris RS: Evidence
for APOBEC3B mutagenesis in multiple human cancers. Nat Genet.
45:977–983. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xu R, Zhang X, Zhang W, Fang Y, Zheng S
and Yu XF: Association of human APOBEC3 cytidine deaminases with
the generation of hepatitis virus B × antigen mutants and
hepatocellular carcinoma. Hepatology. 46:1810–1820. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang T, Cai J, Chang J, Yu D, Wu C, Yan
T, Zhai K, Bi X, Zhao H, Xu J, et al: Evidence of associations of
APOBEC3B gene deletion with susceptibility to persistent HBV
infection and hepatocellular carcinoma. Hum Mol Genet.
22:1262–1269. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
He X, Li J, Wu J, Zhang M and Gao P:
Associations between activation-induced cytidine
deaminase/apolipoprotein B mRNA editing enzyme, catalytic
polypeptide-like cytidine deaminase expression, hepatitis B virus
(HBV) replication and HBV-associated liver disease (Review). Mol
Med Rep. 12:6405–6414. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chan K, Roberts SA, Klimczak LJ, Sterling
JF, Saini N, Malc EP, Kim J, Kwiatkowski DJ, Fargo DC, Mieczkowski
PA, et al: An APOBEC3A hypermutation signature is distinguishable
from the signature of background mutagenesis by APOBEC3B in human
cancers. Nat Genet. 47:1067–1072. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Caval V, Suspene R, Shapira M, Vartanian
JP and Wain-Hobson S: A prevalent cancer susceptibility APOBEC3A
hybrid allele bearing APOBEC3B 3′UTR enhances chromosomal DNA
damage. Nat Commun. 5:51292014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gerlich WH: Medical virology of hepatitis
B: How it began and where we are now. Virol J. 10:2392013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hyams KC: Risks of chronicity following
acute hepatitis B virus infection: A review. Clin Infect Dis.
20:992–1000. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
McMahon BJ, Alward WL, Hall DB, Heyward
WL, Bender TR, Francis DP and Maynard JE: Acute hepatitis B virus
infection: Relation of age to the clinical expression of disease
and subsequent development of the carrier state. J Infect Dis.
151:599–603. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ott JJ, Stevens GA, Groeger J and Wiersma
ST: Global epidemiology of hepatitis B virus infection: New
estimates of age-specific HBsAg seroprevalence and endemicity.
Vaccine. 30:2212–2219. 2012. View Article : Google Scholar : PubMed/NCBI
|