Introduction
Rheumatoid arthritis (RA), which affects
approximately 1% of the population of the world, is characterized
by chronic inflammation and the destruction of the cartilage and
bone in various joints. RA is usually associated with disability
and systemic complications (1).
The extracellular matrix (ECM) is an important structural component
that maintains the integrity of bone tissue. In patients with RA,
degradation of the ECM by matrix metalloproteinases (MMPs) is
considered essential to joint destruction (2) and MMPs also have important roles in
the erosion of joint bone and cartilage (3).
MicroRNAs (miRNAs) are small, conserved, non-coding
RNA molecules that regulate approximately 30% of human genes and
influence several cellular processes, including cell growth,
differentiation, proliferation, and death (4). Numerous studies have demonstrated the
involvement of miRNAs in RA development (5), and these molecules have emerged as
important contributing factors in RA pathophysiology. Some
microRNAs, for example, miRNA-203 (6) and miRNA-155 (7), can affect RA development by
regulating the levels of MMPs that participate in the processes of
bone and joint erosion and destruction. Previously, we determined
that the levels of miR-145-5p are significantly increased in
peripheral blood mononuclear cells (PBMCs) and synovial tissue from
patients with RA compared with healthy control PBMCs and
osteoarthritis (OA) tissue samples. Micro-computed tomography (CT)
of the joints, the expression levels of mRNA and histopathological
analysis of arthritic joints revealed that the miR-145-5p agomir
aggravates cartilage erosion (8);
however, whether miR-145-5p is related to the levels of MMPs in RA
patients and whether this miRNA is involved in bone erosion remains
unknown.
In the present study, it was revealed that
overexpression of miR-145-5p increased expression of MMP-3, MMP-9,
and MMP-13 in RA-fibroblast-like synoviocytes (FLS) from patient
samples. Furthermore, a chemical inhibitor of nuclear factor κB
(NF-κB) signaling, BAY11-7082, significantly attenuated the
expression of MMP-9. Overexpression of miR-145-5p increased p65
nuclear localization and levels of phosphorylated p65 (p-p65),
while significantly reducing those of IkB-α. These results
demonstrated that miR-145 can promote increased MMP-9 levels in
patients with RA by targeted activation of the NF-κB pathway.
Materials and methods
Isolation and culture of FLS from
patients with RA
RA patients (n=30; female, 22; male, 8; median age,
56.2 years; age range, 21–61 years) who were admitted to the
Department of Rheumatology and Immunology, Tianjin Medical
University General Hospital (Tianjin, China), between October 2015
and January 2017, were enrolled in the present study. To isolate
synovial fibroblasts, synovial tissue specimens obtained
immediately after opening the knee-joint capsule were minced and
digested with dispase at 37°C for 60 min. After washing, the cells
were cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), supplemented with 100
IU/ml penicillin or streptomycin and 10% fetal calf serum (HyClone;
GE Healthcare Life Sciences, Logan, UT, USA). RA-FLS cultures were
maintained at 37°C under humid conditions with 5% CO2.
All FLS cultures were subjected to experimental procedures between
passages 4 and 6.
Transfection of FLS
miR-145-5p mimic (cat. no. miR10000437), miR-145-5p
inhibitor (cat. no. miR20000437), mimic negative control (cat. no.
miR01101), and inhibitor negative control (cat. no.miR02101) were
provided by Guangzhou RiboBio Co., Ltd. (Guangzhou, China). Cells
(2×106) were transfected with miR-145-5p inhibitor
(final concentration, 100 nM), miR-145-5p mimic (final
concentration, 50 nM), and corresponding controls using RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.), following the
manufacturer's instructions. Western blot analyses of factors in
the NF-κB, p53/MDM2, JNK, and mitogen-activated protein kinase
(MAPK) pathways were performed using proteins extracted from cells
treated with miR-145-5p mimic and control mimic for 24 h. Chemical
inhibitors were added to cultures 2 h prior to stimulation with
miR-145-5p. Subsequently, the cells were used for functional assays
or RNA/protein extraction.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol® reagent (Takara Bio, Inc., Otsu,
Japan) was used to isolate total RNA, including miRNA from FLS. A
FastQuant RT Kit (Tiangen Biotech Co., Ltd., Beijing, China) was
used to reverse-transcribe RNA into complementary DNA, following
the manufacturer's instructions. SYBR-Green PCR Master Mix (Takara
Bio, Ic.) was used to perform RT-qPCR, on a Light Cycler 480 (Roche
Diagnostics, Indianapolis, IN, USA) with the following program:
95°C for 30 sec, followed by 40 cycles at 95°C for 5 sec, 60°C for
30 sec, 95°C for 5 sec, and 60°C for 1 min. Specific primers were
synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). A
forward primer was designed based on the mature miR-145-5p DNA
sequence (5′-GUCCAGUUUUCCCAGGAAUCCCU-3′) from miRBase (http://www.mirbase.org), whereas a universal 3′
reverse primer (5′-AGTGCAGGGTCCGAGGTATT-3′; Guangzhou RiboBio Co.,
Ltd.) was used. Other primers were as follows: MMP-1 forward,
5′-TTCTACATGCGCACAAATCC-3′ and reverse, 5′-ACCGGACTTCATCTCTGTCG-3′;
MMP-3 forward, 5′-GCGCCCTGGTCCTGGTGTCCAT-3′ and reverse,
5′-GAAACCACAATTCTGTCTTTCAC-3′; MMP-9 forward,
5′-CCTTCACTTTCCTGGGTAAG-3′ and reverse, 5′-CCATTCACGTCGTCCTTATG-3′;
MMP-13 forward, 5′-TCCTGATGTGGGTGAATACAATG-3′ and reverse,
5′-GCCATCGTGAAGTCTGGTAAAAT-3′. GAPDH (forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′) and U6 (forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′
were used as internal controls for mRNA and miRNA, respectively.
Relative quantification was performed based on differences in
cross-threshold values (ΔCq) between the gene of interest and the
endogenous control [i.e., Cq(gene of interest)-Cq (endogenous
control)]. Relative expression was calculated using the comparative
threshold cycle method (9).
Enzyme-linked immunosorbent assay
(ELISA)
The concentrations of MMP-1 (cat. no. 70-EK1M01-96),
MMP-3 (cat. no. 70-EK1M03-96), MMP-9 (cat. no. 70-EK1M09-96) and
MMP-13 (cat. no. 70-EK1M132) were assessed in cell supernatants by
ELISA using the SensoLyte ELISA kit according to the instructions
of the manufacturer (MultiSciences (Lianke) Biotech Co., Ltd.,
Hangzhou, China).
Western blot analysis
Cultured FLS were lysed in radioimmunoprecipitation
assay buffer supplemented with a protease inhibitor cocktail (both
from Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China), and boiled. The protein concentration was measured using a
BCA Protein Assay kit (Thermo Fisher Scientific, Inc.). Equal
volumes (25 µg) of concentrated samples were separated by sodium
dodecyl sulfate polyacrylamide gel electrophoresis on 8–10% gels,
before transfer to polyvinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA). Non-fat dried milk powder in Tris
buffered saline with 0.1% Tween-20 was used to block membranes at
room temperature for 2 h, and membranes were then incubated with
primary antibodies at 4°C overnight. Incubation with horseradish
peroxidase-conjugated secondary antibody (1:5,000; cat. no. 7074S;
Cell Signaling Technology, Inc., Danvers, MA, USA) for 1 h at room
temperature was used to assess the protein levels. Enhanced
chemiluminescence (Amersham Life Sciences; GE Healthcare Life
Sciences, Little Chalfont, UK) reagent was used to visualize
signals, and individual protein band intensities were quantified
using ImageJ software (version 1.6.0; National Institutes of
Health, Bethesda, MD, USA). Primary antibodies used were as
follows: MMP-1 (1:1,000; cat. no. ab137332), MMP-3 (1:1,000; cat.
no. ab52915), MMP-9 (1:1,000; cat. no. ab38898), MMP-13 (1:3,000;
cat. no. ab39012), IκBα (1:2,000; cat. no. ab32518), p65 (1:3,000;
cat. no. ab32536), and p-p65 (1:700; Ser536; cat. no. ab28856) were
all purchased from Abcam (Cambridge, MA, USA). β-actin antibody
(1:2,000; cat. no. ab8227; Abcam) was applied as a loading
control.
Induction of CIA
Male DBA/1 mice (15–20 g; 8 weeks old) were used in
the present study. A total of 30 animals were housed at 24±1°C with
a 12:12-h light/dark cycle and a relative humidity of 56±5%.
Animals were provided access to food and water ad libitum,
and were fed a commercial diet according to the guidelines of the
Animal Ethical and Welfare Committee. All mice were housed
individually. Lyophilized bovine type II collagen (Chondrex, Inc.,
Redmond, WA, USA) was dissolved overnight at 4°C in 0.05 M acetic
acid under constant stirring. Next, the collagen was emulsified
using Freund's complete adjuvant (Chondrex, Inc.) to provide a
final concentration of 2 mg/ml. Mice were injected in the tail with
0.1 ml of emulsion on day 1 and booted intraperitoneally on day 21
with collagen. On day 30, an equal volume of mir-145-5p agomir or
scrambled miRNA agomir (15 nM; Guangzhou RiboBio Co., Ltd.) was
injected in the tail vein of mice at 1-week intervals. Four weeks
after the first injection, the mice were sacrificed by injecting
pentobarbital sodium at a dose of 100 mg/kg, then various
indications such as respiration, heartbeat, pupil reflection, and
nerve reflex were observed in the mice, and when all the
aforementioned reactions disappeared, death was judged. Next, the
right hind limb was dissected and synovial tissue was collected for
subsequent experiments.
Immunohistochemistry (IHC)
IHC analysis of MMPs was conducted using 3-µm-thick
sections from 8-week-old male DBA/1 mice, obtained from Hua Fukang
Co., Ltd. (Beijing, China). Sections were fixed in 4% methanol at
4°C for 24 h, washed in PBS and treated with retrieval solution
(Dako; Agilent Technologies, Inc., Santa Clara, CA, USA) for 20 min
at 90°C. Following incubation in 5% BSA (cat. no. SW3015; Beijing
Solarbio Science & Technology Co., Ltd.) at 4°C for 30 min, the
sections were treated with MMP-1 (1:1,000; cat. no. ab137332),
MMP-3 (1:1,000; cat. no. ab52915), MMP-9 (1:1,000; cat. no.
ab38898) and MMP-13 (1:3,000; cat. no. ab39012) primary antibodies
overnight at 4°C. Sections incubated with Alexa Fluor®
568-conjugated rabbit anti-goat immunoglobulin G (1:1,000; cat. no.
A-11079; Molecular Probes; Thermo Fisher Scientific, Inc.) as a
secondary antibody for 1 h at room temperature. Antibodies were
diluted with 5% BSA. DAPI solution was applied for 5 min at room
temperature for nuclear staining. Negative controls were prepared
in the same manner, but without primary antibody. Images were
captured using an Olympus IX2-UCB microscope (magnification, ×40;
Olympus Corporation, Tokyo, Japan).
Investigation of signaling
pathways
Specific inhibitors of various signaling pathways
were purchased from Beijing Solarbio Science & Technology Co.,
Ltd. and used to analyze the effects of these pathways on levels of
MMP-3, MMP-9, and MMP-13. Briefly, specific inhibitors of NF-κB (20
µM, BAY11-7082), p53/MDM2 (10 µM, Nutlin-3), JNK (10 µM, SP600125),
and MAPK (10 µΜ, SB203580) pathways (all from Beyotime Institute of
Biotechnology, Haimen, China) were added to FLS cultures 2 h prior
to miR-145-5p stimulation, following the manufacturer's
instructions. RT-qPCR and western blotting were used to assess the
expression levels of MMP-3, MMP-9, and MMP-13 as
aforementioned.
Immunofluorescence
FLS cells were fixed in 4% methanol at 4°C for 15
min, washed three times (5 min each) with PBS, permeabilized with
0.1% Triton X-100 for 15 min, then, incubated overnight at 4°C with
anti-NF-κB p65 primary antibody (1:100; cat. no. ab32536; Abcam),
followed by incubation with FITC-labeled secondary antibody (cat.
no. ab7086; Abcam) at room temperature for 1 h. Then, the cells
were counterstained with DAPI for 5 min. Images were captured using
a Leica TCS SP5 confocal fluorescence microscope (magnification,
×80). The negative control was prepared in the same manner without
the primary antibody.
Target sequence prediction
TargetScan (version 7.2; http://www.targetscan.org/vert_72/docs/help.html)
and mirDB (version 6.0; http://www.mirdb.org/download.html) were employed to
search for putative target sequences of miR-145-5p in the NF-κB
pathway.
Statistical analysis
Data were expressed as the mean ± standard error of
the mean (SEM) from at least three independent experiments. The
difference among groups was determined by analysis of variance
(ANOVA) followed by Bonferroni's test, for qPCR experiments and
western blotting, and differences between the two groups were
analyzed using a two-tailed Student's t-test. Differences were
considered statistically significant for P-values <0.05
(P<0.05, P<0.01, P<0.001).
Results
Overexpression of miR-145-5p enhances
MMP-3, MMP-9, and MMP-13 levels in RA-FLS
FLS samples isolated from synovial tissue derived
from patients with RA were used to investigate the effect of
miR-145-5p on synoviocytes in vitro. FLS were incubated with
miR-145-5p mimic, miR-145-5p inhibitor, and their corresponding
negative controls for 48 h. MMP-3, MMP-9, and MMP-13
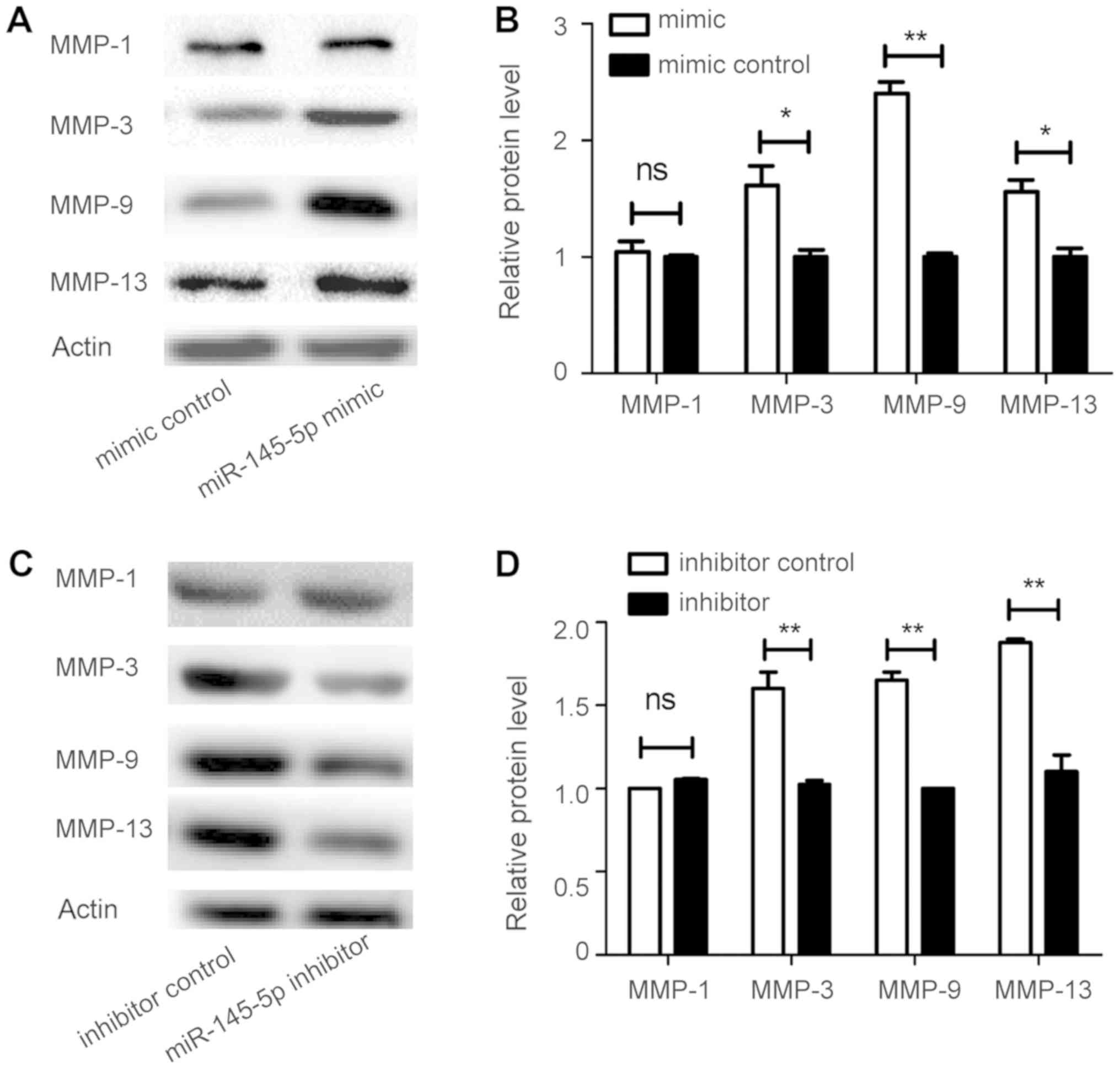
RNA levels were significantly increased by miR-145-5p mimic, while
they were downregulated by miR-145-5p inhibitor; however, no change
in MMP-1 was detected (Fig. 1A
and B). Consistent with the changes in secreted protein levels
in cell culture supernatants (Fig.
1C-F), levels of intracellular MMP-3, MMP-9, and MMP-13
proteins were markedly enhanced by treatment with miR-145-5p mimic
(Fig. 2A and B), but decreased
after treatment with miR-145-5p inhibitor (Fig. 2C and D). These results indicated
that miR-145-5p regulates various MMPs in FLS.
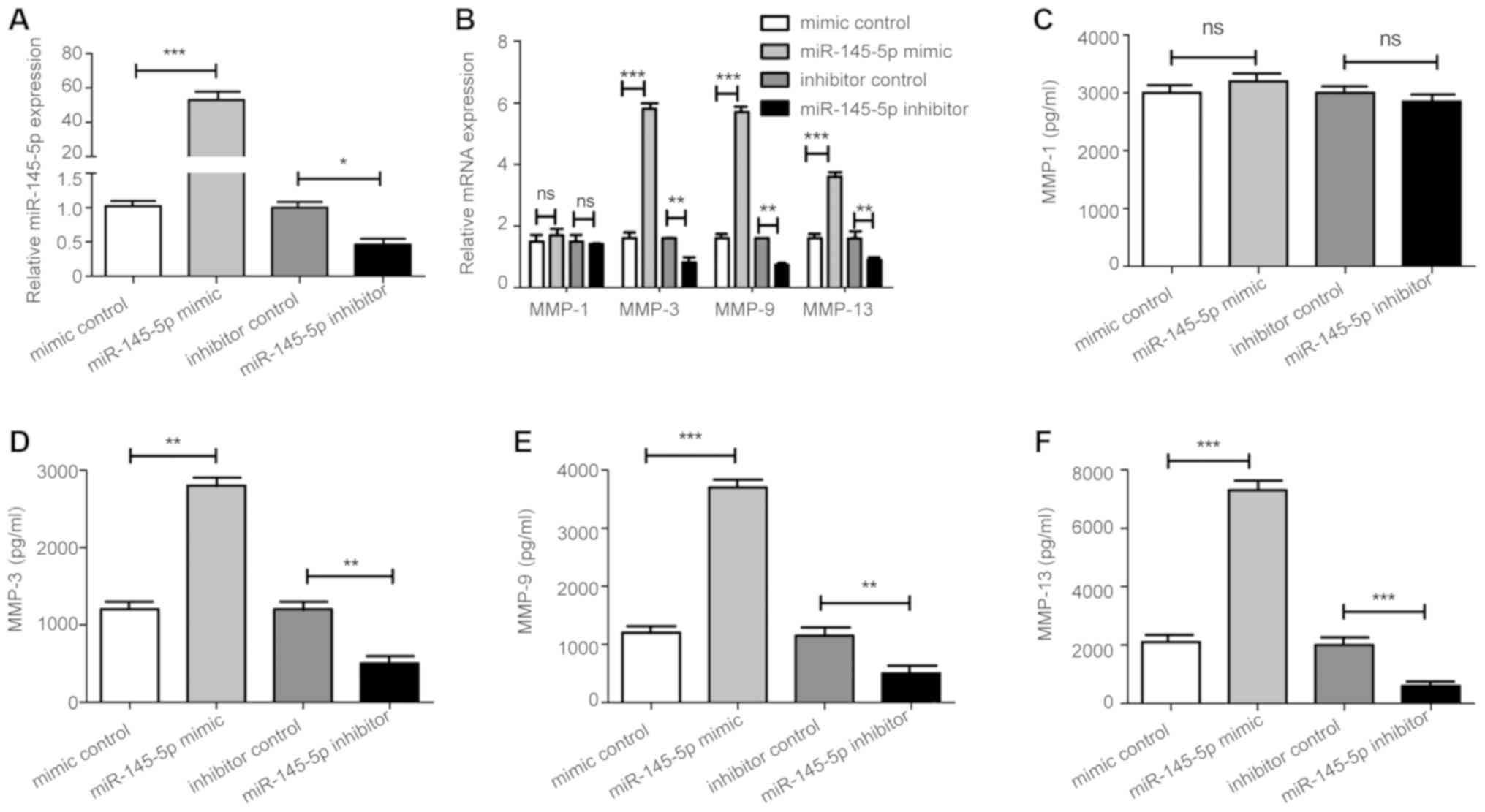 | Figure 1.miR-145-5p regulates MMPs. (A) After
transfection with the miR-145-5p mimic, miR-145-5p inhibitor, and
respective controls, levels of miR-145-5p in RA-FLS were assessed
by RT-qPCR. (B) Assessment of intracellular levels of MMPs in
RA-FLS after transfection with miR-145-5p mimic, miR-145-5p
inhibitor, and respective controls by RT-qPCR and (C-F) ELISA. This
experiment was repeated at least three times with similar results;
one set of representative results is presented. *P<0.05,
**P<0.01, ***P<0.001 vs. control. Values are expressed as the
mean ± SD. MMPs, matrix metalloproteinases; RA, rheumatoid
arthritis; FLS, fibroblast-like synoviocytes; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; ELISA,
enzyme-linked immunosorbent assay. |
Different mRNA expression levels of
MMP-1, MMP-3, MMP-9, and MMP-13 in collagen-induced arthritis
mice
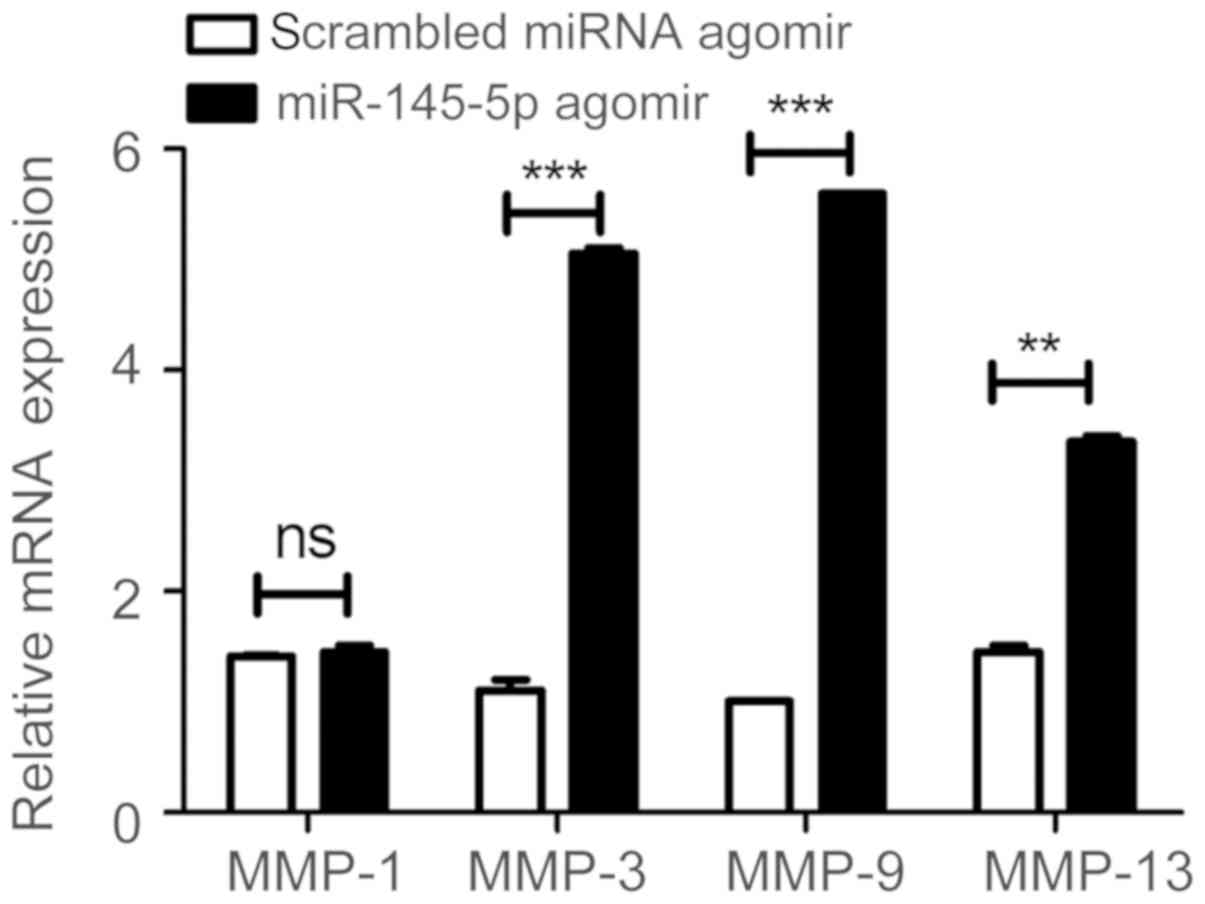
miR-145-5p agomir or scrambled miRNA agomir was
injected into CIA mice in the tail vein. It was observed that
miR-145-5p agomir significantly upregulated MMP-3, MMP-9, and
MMP-13 compared with the mice treated with the scrambled miRNA
agomir, however, the level of MMP-1 underwent no apparent
alteration (Fig. 3).
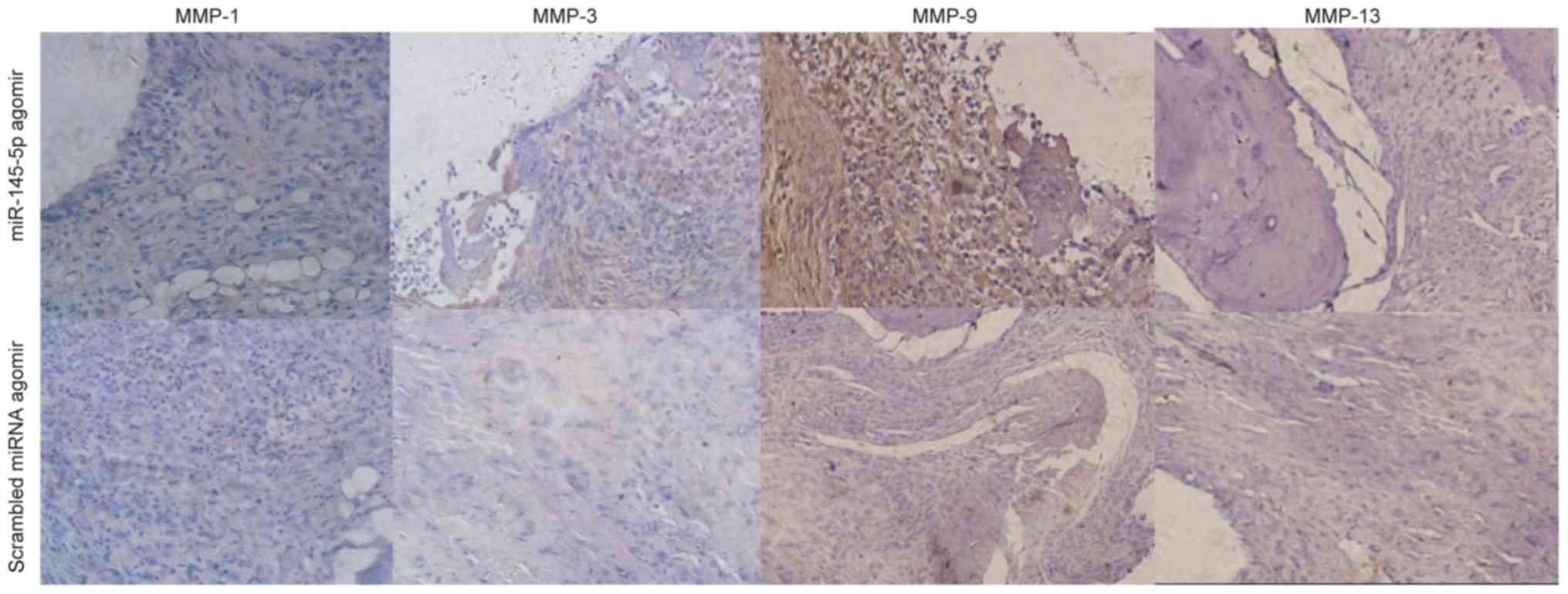
MMP-1, MMP-3, MMP-9, and MMP-13 levels
in miR-145-5p-treated mouse joints
IHC analysis (Fig.
4) revealed increased MMP-3, MMP-9, and MMP-13 levels in the
hind ankles of mice treated with miR-145-5p agomir, compared with
those in the right hind ankles of control mice.
miR-145-5p abrogates stimulation of
MMP-9 expression by NF-κB inhibitors
To explore whether there is a direct functional
connection among miR-145-5p, cell signaling pathways, and MMP
expression, the effects of NF-κB, p53/MDM2, MAPK and JNK signaling
pathways were assessed using specific inhibitors targeting NF-κB
(BAY11-7082, 20 µM), p53/MDM2 (Nutlin-3, 10 µM), MAPK (SB203580, 10
µM), and JNK (SP600125, 10 µM). In RA-FLS stimulated by miR-145-5p,
the NF-κB inhibitor, BAY11-7082, significantly inhibited MMP-9
levels compared to those in the controls (Fig. 5A-C), while inhibitors of the other
three cell signaling pathways had no marked effect on the levels of
MMP-3, MMP-9, or MMP-13 (Fig.
5D-F).
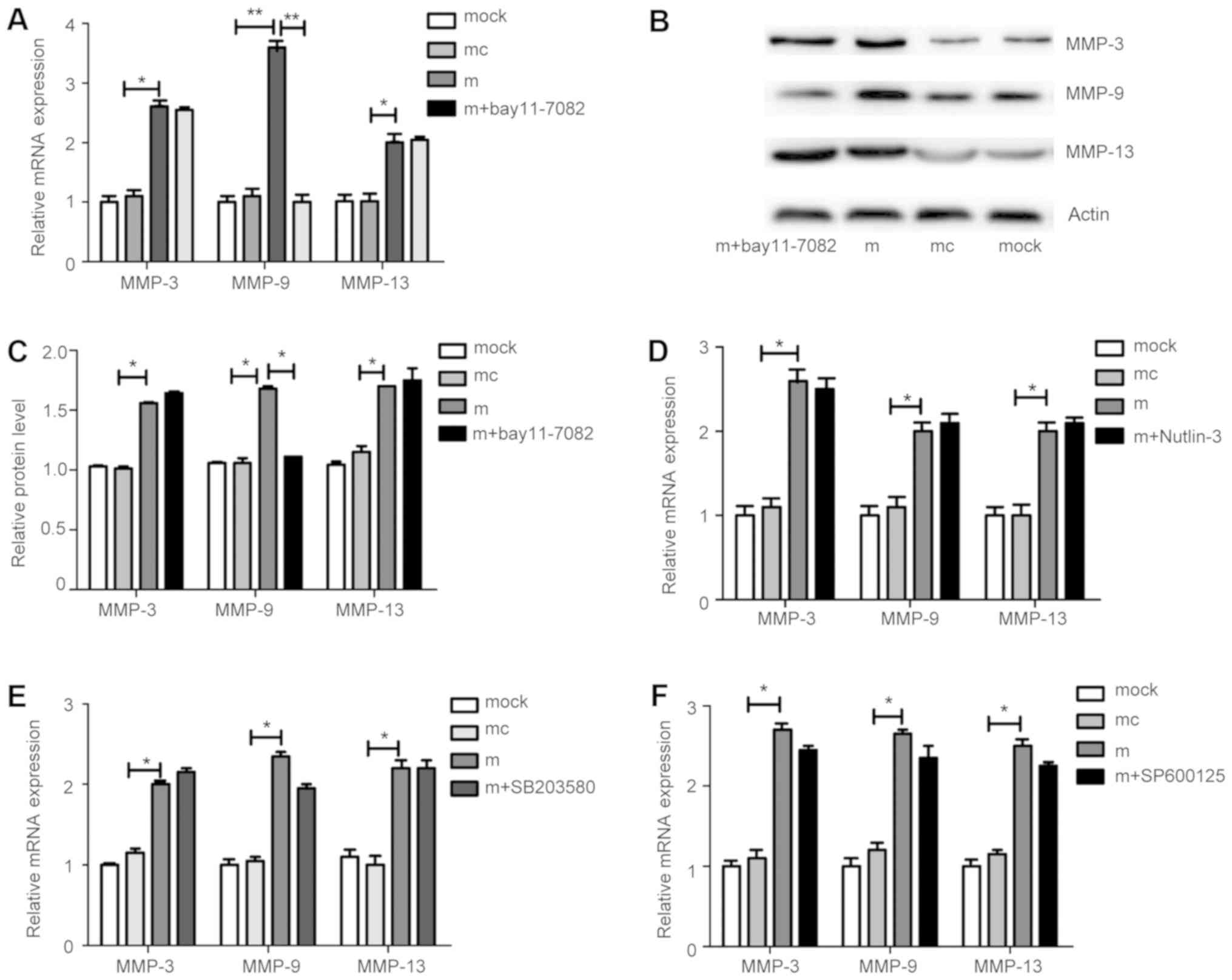 | Figure 5.Changes in MMP levels following
treatment with JNK, MAPK, NF-κB, and p53/MDM2 pathway inhibitors in
FLS overexpressing miR-145-5p. (A and B) An inhibitor of the NF-κB
pathway significantly suppressed levels of MMP-9, but not MMP-3 or
MMP-13. (C) Quantification data are presented for the corresponding
groups. (D-F) Inhibitors of the p53/MDM2, MAPK, and JNK pathways
had no effect on MMP-3, MMP-9, or MMP-13 levels. *P<0.05,
**P<0.01. Results are presented as the mean ± SD of three
independent experiments. MMP, matrix metalloproteinase; NF-κB,
nuclear factor κB; FLS, fibroblast-like synoviocytes. |
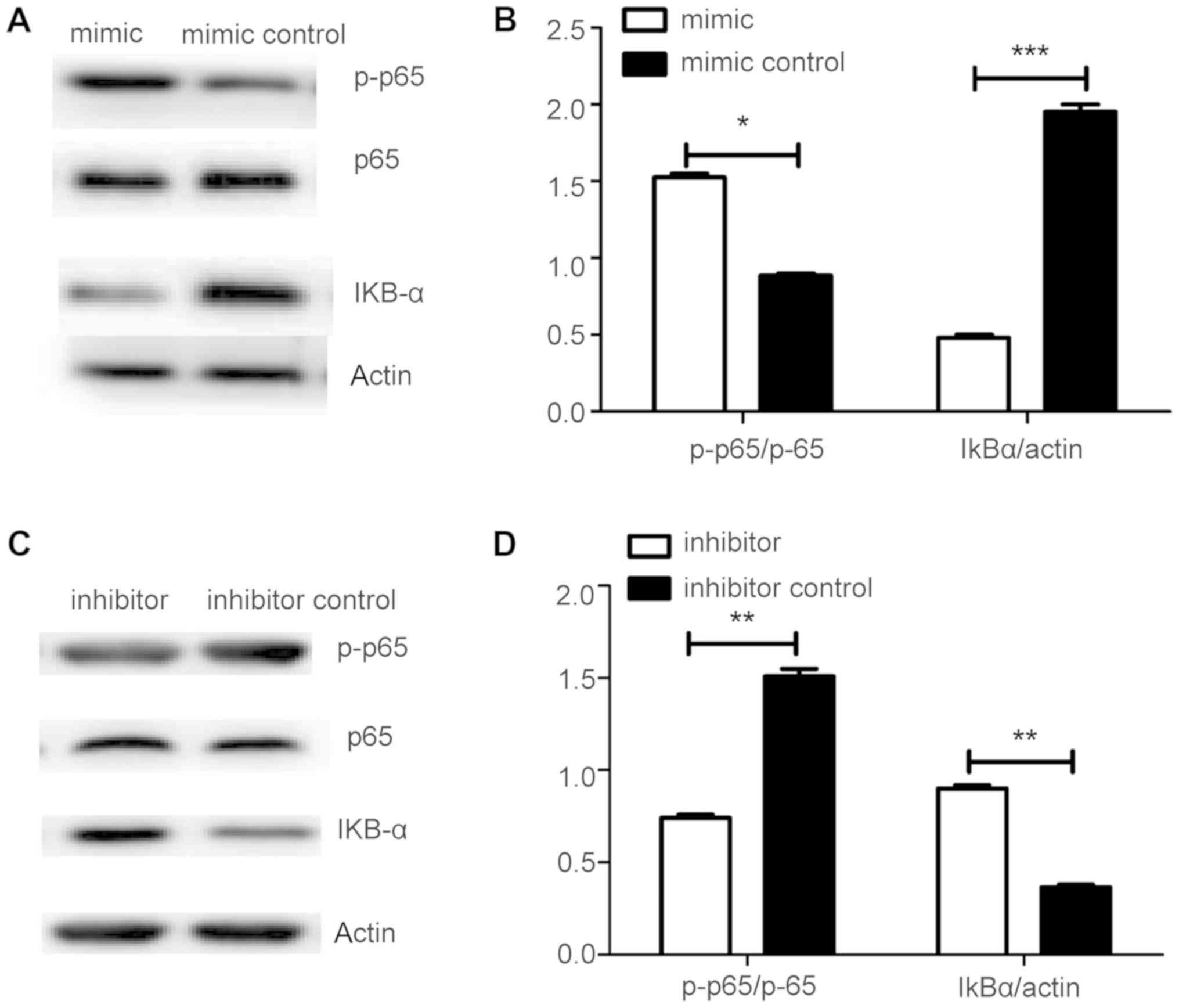
miR-145-5p activates the NF-ΚB
signaling pathway in FLS samples from patients with RA
To explore the mechanism underlying the involvement
of miR-145-5p in MMP activation, the effects of the key NF-κB
pathway factors were assessed, including p65, p-p65, and IkB-a, in
response to miR-145-5p control of MMP-9 synthesis. Levels of p-p65
were significantly increased, while those of IkB-a were markedly
decreased in response to miR-145-5p (Fig. 6A and B). Furthermore, reduction of
miR-145-5p led to marked downregulation of p-p65 levels, while it
increased those of IkB-a (Fig.
6C-D). In conclusion, miR-145-5p can contribute to arthritis
progression by activating NF-κB signaling, which controls MMP-9
expression.
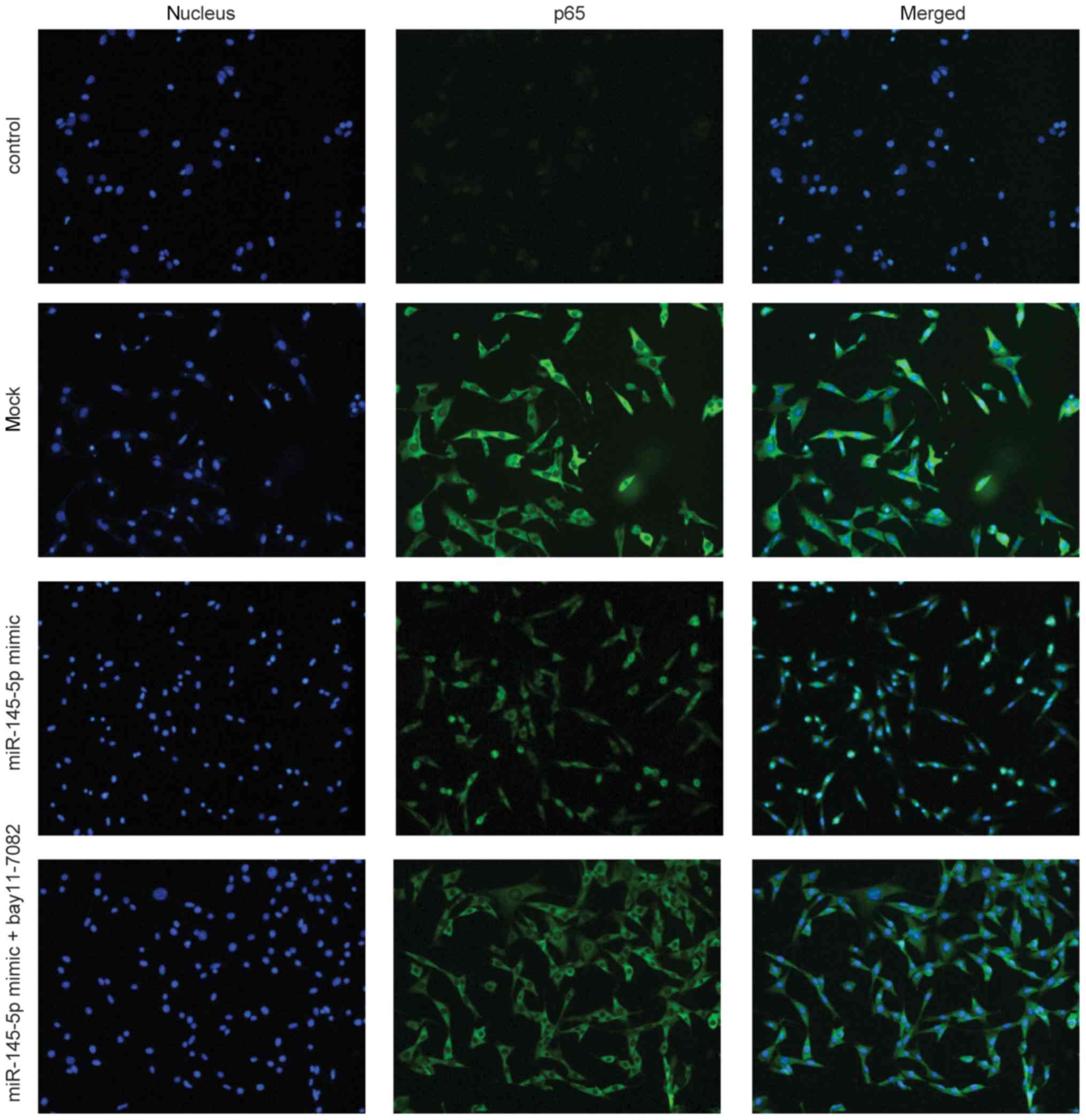
miR-145-5p activates NF-κB by
promoting p65 translocation from the cytoplasm to the nucleus
An immunofluorescence assay was conducted to
investigate p65 nuclear translocation in FLS. miR-145-5p induced
NF-κB p65 nuclear translocation in FLS; however, this change was
reversed by treatment with the NF-κB pathway inhibitor, BAY11-7082
(Fig. 7). These findings
demonstrated that BAY11-7082 can attenuate miR-145-5p-induced NF-κB
activation.
Discussion
During normal physiological processes, the synthesis
and degradation of joint-cartilage matrix proteins occurs in a
state of dynamic equilibrium; however, in patients with RA, this
equilibrium becomes disrupted. Matrix protein degradation and
excessive bone resorption lead to the loss of cartilage matrix
proteins and bone, and consequent gradual loss of joint integrity
(10). The function of MMPs is
mainly the degradation of type II collagen and proteoglycans;
however, their aberrant expression can also promote erosion and
destruction of articular cartilage in patients with RA (11).
Patients with RA have increased levels of various
MMPs in their synovial fluid and peripheral blood, with these
levels closely related to disease activity and bone and joint
destruction (12). Erosion of the
articular cartilage, the ECM of which is comprised of proteoglycans
and collagen (mainly type II collagen), is the most prominent
manifestation of RA, and loss of structural integrity directly
affects the mechanics of the joint (13).
Aggrecanase, which is secreted by synoviocytes, can
degrade the proteoglycan matrix, while MMP-9 can degrade type II
collagen. Giannelli et al (14) previously revealed that, in an
arthritic mouse model, MMP-2 and MMP-9 have roles in arthritis
pathogenesis. Various signaling pathways can promote MMP activity,
including MAPK, signal transduction and transcriptional activator
(STAT), and SMAD (15)
pathways.
miR-146b was revealed to reduce glioma cell invasion
by targeting MMP-16 gene expression in human patients with glioma
(16). Stanczyk et al
(7) and Wu et al (17) revealed that miR-155 regulated MMP-1
and MMP-3 levels by acting on the target molecule, SOCS1, to
activate the APK/JNK signaling pathway, thereby affecting
osteoblast and osteoclast metabolism. Furthermore, Akhtar et
al (18) revealed that
negative regulation of miR-27b expression by activation of MAPK and
NF-κB signaling may be necessary for IL-1β-mediated stimulation of
MMP-13 production in OA chondrocytes.
In the present study, overexpression of miR-145-5p
in RA-FLS significantly increased MMP-3, MMP-9, and MMP-13 levels.
Analysis of MMP levels after the addition of JNK, MAPK, NF-κB, or
p53-MDM2 pathway inhibitors demonstrated that only MMP-9 levels
were significantly decreased by the NF-κB inhibitor,
BAY11-7082.
NF-κB is a transcription factor that regulates the
expression of multiple genes and a key signaling factor in the
control of inflammation, synovial hyperplasia, and matrix
degeneration. This transcription factor also regulates the
expression of many pro-inflammatory genes (19,20).
An NF-κB binding site was first identified in the MMP-9 promoter
and associated with TNF-α gene induction, and subsequently reported
to mediate the synergistic effects of growth factors and cytokines
on MMP-9 (21). p65 is a key
factor in the NF-κB pathway, and p-p65 can serve as a marker for
NF-κB activation (22), while
IkB-α is an inhibitor of NF-κB, and decreased IκB-α levels are also
indicative of NF-κB pathway activation (23).
Herein, the regulatory effects of miR-145-5p on MMPs
were assessed in bone-induced rheumatoid arthritis. The results
revealed that miR-145-5p overexpression significantly increased
MMP-9 levels and enhanced p65 nuclear translocation, while
increasing the levels of p-p65 and decreasing those of IkB-α; these
effects were reversed by the NF-κB inhibitor, BAY11-7082. In
conclusion, these studies suggest that miR-145 promotes increased
MMP-9 levels in patients with RA by activating the NF-κB pathway,
which accelerates the process of bone erosion, increasing disease
severity of RA patients; however, predictions using two informatics
software tools, TargetScan and miRDB, failed to identify a direct
target of miR-145-5p in the NF-κB pathway (data not shown),
suggesting that miR-145-5p may not affect MMP levels by directly
regulating this pathway, and that other cytokines may also be
involved in the regulatory process. For example, it has been
reported that tumor necrosis factor-α (TNF-α), interleukin (IL)-1
and IL-6 can play an important role in cartilage catabolism as
major pro-inflammatory cytokines (24). These cytokines may activate more
signaling pathways to form a complex regulatory network. Therefore,
further studies should be performed to confirm the results.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the
National Natural Science Foundation of China (Grant/Award ‘Project
no. 81330029’), Tianjin Medical University General Hospital
(Tianjin, China).
Availability of data and materials
All datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YHY conceived and designed the experiments. XXW, KT,
YYW, YQC, and MCY performed the cell culture, RT-qPCR, and IHC
experiments, and interpreted the data. XXW, KT, YW, JW, and CGG
performed the ELISA and western blotting experiments and
interpreted the data. XXW wrote the manuscript. All authors read
and approved the final manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Tianjin Medical University General Hospital (no.
IRB2015-YX-081) and the Animal Ethical and Welfare Committee (AEWC;
no. TMUaMEC 2018035). Informed consent was obtained from all
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CIA
|
collagen-induced arthritis
|
|
ECM
|
extracellular matrix
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
FLS
|
fibroblast-like synoviocytes
|
|
IHC
|
immunohistochemical
|
|
MAPK
|
mitogen-activated protein kinase
|
|
miRNAs
|
microRNAs
|
|
MMP
|
matrix metalloproteinase
|
|
NF-κB
|
nuclear factor-κΔ
|
|
OA
|
osteoarthritis
|
|
PBMCs
|
peripheral blood mononuclear cells
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
RA
|
rheumatoid arthritis
|
|
STAT
|
signal transduction and
transcriptional activator
|
References
|
1
|
McInnes IB and O'Dell JR:
State-of-the-art: Rheumatoid arthritis. Ann Rheum Dis.
69:1898–1906. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cunnane G, Fitzgerald O, Beeton C, Cawston
TE and Bresnihan B: Early joint erosions and serum levels of matrix
metalloproteinase 1, matrix metalloproteinase 3, and tissue
inhibitor of metalloproteinases in rheumatoid arthritis. Arthritis
Rheum. 44:2263–2274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Murphy G, Knäuper V, Atkinson S, Butler G,
English W, Hutton M, Stracke J and Clark I: Matrix
metalloproteinases in arthritic disease. Arthritis Res. 4 (Suppl
3):S39–S49. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schanen BC and Li X: Transcriptional
regulation of mammalian miRNA genes. Genomics. 97:1–6. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tavasolian F, Abdollahi E, Rezaei R,
Momtazi-Borojeni AA, Henrotin Y and Sahebkar A: Altered expression
of microRNAs in rheumatoid arthritis. J Cell Biochem. 119:478–487.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stanczyk J, Ospelt C, Karouzakis E, Filer
A, Raza K, Kolling C, Gay R, Buckley CD, Tak PP, Gay S and Kyburz
D: Altered expression of microRNA-203 in rheumatoid arthritis
synovial fibroblasts and its role in fibroblast activation.
Arthritis Rheum. 63:373–381. 2014. View Article : Google Scholar
|
|
7
|
Stanczyk J, Pedrioli DM, Brentano F,
Sanchez-Pernaute O, Kolling C, Gay RE, Detmar M, Gay S and Kyburz
D: Altered expression of microRNA in synovial fibroblasts and
synovial tissue in rheumatoid arthritis. Arthritis Rheum.
58:1001–1009. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Y, Wang X, Yang M, Ruan W, Wei W, Gu
D, Wang J, Guo X, Guo L and Yuan Y: miR-145-5p increases osteoclast
numbers in vitro and aggravates bone erosion in collagen-induced
arthritis by targeting osteoprotegerin. Med Sci Monit.
24:5292–5300. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta c(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xia Y: The role of RLX/RXFP1, MMP9/MMP13
and RANKL/OPG in arthritic cartilage and bone destruction and
Chinese medicine treatment [d]. Huazhong Univ Sci Technol; 2012
|
|
11
|
Gravallese EM, Darling JM, Ladd AL, Katz
JN and Glimcher LH: In situ hybridization studies of stromelysin
and collagenase messenger RNA expression in rheumatoid synovium.
Arthritis Rheum. 34:1076–1084. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nakada M, Nakamura H, Ikeda E, Fujimoto N,
Yamashita J, Sato H, Seiki M and Okada Y: Expression and tissue
localization of membrane-type 1, 2, and 3 matrix metalloproteinases
in human astrocytic tumors. Am J Pathol. 154:417–428. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dai SM, Shan ZZ, Nishioka K and Yudoh K:
Implication of interleukin 18 in production of matrix
metalloproteinases in articular chondrocytes in arthritis: Direct
effect on chondrocytes may not be pivotal. Ann Rheum Dis.
64:735–742. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giannelli G, Erriquez R, Iannone F,
Marinosci F, Lapadula G and Antonaci S: MMP-2, MMP-9, TIMP-1 and
TIMP-2 levels in patients with rheumatoid arthritis and psoriatic
arthritis. Clin Exp Rheumatol. 22:335–338. 2004.PubMed/NCBI
|
|
15
|
Ni S, Li C, Xu N, Liu X, Wang W, Chen W,
Wang Y and van Wijnen AJ: Follistatin-like protein 1 induction of
matrix metalloproteinase 1, 3 and 13 gene expression in rheumatoid
arthritis synoviocytes requires MAPK, JAK/STAT3 and NF-κB pathways.
J Cell Physiol. 234:454–463. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia H, Qi Y, Ng SS, Chen X, Li D, Chen S,
Ge R, Jiang S, Li G, Chen Y, et al: microRNA-146b inhibits glioma
cell migration and invasion by targeting MMPs. Brain Res.
1269:158–165. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu T, Xie M, Wang X, Jiang X, Li J and
Huang H: miR-155 modulates TNF-α-inhibited osteogenic
differentiation by targeting SOCS1 expression. Bone. 51:498–505.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Akhtar N, Rasheed Z, Ramamurthy S,
Anbazhagan AN, Voss FR and Haqqi TM: MicroRNA-27b regulates the
expression of matrix metalloproteinase 13 in human osteoarthritis
chondrocytes. Arthritis Rheum. 62:1361–1371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hammaker D, Sweeney S and Firestein GS:
Signal transduction networks in rheumatoid arthritis. Ann Rheum
Dis. 62 (Suppl 2):ii86–ii89. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miagkov AV, Kovalenko DV, Brown CE,
Didsbury JR, Cogswell JP, Stimpson SA, Baldwin AS and Makarov SS:
NF-kappaB activation provides the potential link between
inflammation and hyperplasia in the arthritic joint. Proc Nat Acad
Sci USA. 95:13859–13864. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He C: Molecular mechanism of
transcriptional activation of human gelatinase B by proximal
promoter. Cancer Lett. 106:185–191. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goon Goh FG, Sloss CM, Cunningham MR,
Nilsson M, Cadalbert L and Plevin R: G-protein-dependent and
-independent pathways regulate proteinase-activated receptor-2
mediated P65 NFkappaB serine 536 phosphorylation in human
keratinocytes. Cell Signal. 20:1267–1274. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Q and Verma IM: NF-kappaB regulation in
the immune system. Nat Rev Immunol. 2:725–734. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
van de Loo FA, Joosten LA, van Lent PL,
Arntz OJ and van den Berg WB: Role of interleukin-1, tumor necrosis
factor alpha, and interleukin-6 in cartilage proteoglycan
metabolism and destruction. Effect of in situ blocking in murine
antigen- and zymosan-induced arthritis, Arthritis Rheum.
38:164–172. 1995.
|