Introduction
Gallbladder carcinoma is one of the tumor types with
the worst prognosis globally, with an average survival length of ~6
months and a 5-year survival rate <5% (1). There are two main factors associated
with the poor prognosis. Firstly, gallbladder carcinoma is not
sensitive to traditional chemotherapy or radiotherapy and surgery
is the only effective treatment However, due to the lack of
symptoms in the early stages of the disease, most patients do not
have the opportunity to receive a surgical treatment (2). Secondly, the biological
characteristics of gallbladder carcinoma lead to a high possibility
of recurrence or metastasis after surgery (3). Recent studies have shown that various
signaling pathways are associated with the development of
gallbladder carcinoma, including Hedgehog, PI3K/AKT/mTOR, Notch,
Erb-b receptor tyrosine kinases (ErbB) and the mitogen-activated
protein kinase (MAPK)/ERK pathway (4). However, their underlying mechanisms
remain unclear.
Myelin protein 0-like 1 (MPZL1), also known as PZR,
is a surface glycoprotein. It belongs to the immunoglobulin
superfamily and has an extracellular segment with significant
sequence homology with myelin protein 0 (a major structural protein
of myelin) (5–8). There are two immunoreceptor
tyrosine-based inhibitory motifs (ITIMs) in the intracellular
domain of MPZL1 that specifically interact with Src homology
phosphatase type-2 (SHP2) (6).
SHP2 is a tyrosine phosphatase containing two Src homology 2
domains encoded by the protein tyrosine phosphatase non-receptor
type 11 (PTPN11) gene (9). Studies
have confirmed that SHP2 is a cancer-promoting protein (10,11).
Mutations in the PTPN11 gene result in a variety of hematological
malignancies, juvenile myelomonocytic leukemia, and in the
development of certain solid tumors (12). SHP2 is thought to be regulatory
protein affecting tumor cell proliferation, differentiation and
survival through to a variety of signaling pathways, including
Ras/Raf/MEK/ERK, Janus kinase (JAK)-STAT, PI3K/AKT/mTOR and
Wnt/β-catenin signaling pathways (13).
As a highly conserved protein, MPZL1 is expressed in
a variety of cells, suggesting that it may have an important role
in basic cellular functions. A previous study found that MPZL1
protein may promote the fibronectin-dependent cell migration
process by recruiting and activating SHP2 protein (7). The binding of intracellular ITIMs of
MPZL1 protein to SHP2 may regulate cell migration, as MPZL1 lacking
ITIMs does not promote cell migration (8). Recent studies have also shown that
MPZL1 may promote the fibronectin-dependent migration of mouse
embryonic fibroblasts (8,14), and may be involved in
adhesion-dependent signaling (15,16).
However, the functional roles and clinical implications of MPZL1
amplification and overexpression in human cancers are largely
unknown.
There are reports regarding role of MPZL1 gene in
hepatocellular carcinoma. Jia et al (17) revealed that MPZL1 gene was
upregulated in liver cancer tissue, and its expression was
associated with clinical pathological parameters, such as age,
histological grade and intrahepatic metastasis of patients with
liver cancer. It also revealed that MPZL1 can activate cortactin
and promote the migration and metastasis of hepatoma cells via the
Src-mediated signaling pathway (17). Previous studies have also reported
that MPZL1 mediates the phosphorylation of cortactin by increasing
the phosphorylation levels of Src kinase, which may in turn lead to
the development of distant metastases of hepatocellular carcinoma,
such as lung metastasis (5,17,18).
Therefore, it may be possible that the upregulation of MPZL1 may
affect the invasiveness or metastatic ability of gallbladder
carcinoma cells.
To the best of our knowledge, there have been no
studies on the function and molecular mechanism of MPZL1 in
gallbladder carcinoma. Therefore, the present study aimed to
elucidate the function of MPZL1 expression in gallbladder carcinoma
tissues, and to further understand the effect of MPZL1 expression
on gallbladder carcinoma cell functions, including proliferation,
invasion, metastasis and apoptosis.
Materials and methods
Patients and tissue specimens
From January 2011 to June 2017, 82 patients
(male:female ratio, 52:30; median age, 63 years; age range, 44–77
years) with gallbladder carcinoma that received surgical treatment
at the Department of General Surgery, China-Japan Friendship
Hospital (Beijing, China) enrolled in the present study. The
carcinoma and paracarcinoma samples collected from all 82 patients
were stored in paraffin in the Department of Pathology. From
January 2016 to June 2017, an additional 20 paired samples of
gallbladder carcinoma and adjacent healthy tissue from 20
(male:female, 13:7; median age, 63 years; age range, 44–75 years)
of the 82 patients were snap frozen at −80°C. Moreover, from
January 2017 to June 2017, gallbladder samples of 20 patients
(male:female ratio, 12:8; median age, 62 years; age range, 45–73
years) with benign cholecystitis who received surgical treatment
were also collected and frozen as a normal control group. The
pathological diagnosis of all the 82 patients with gallbladder
carcinoma and the 20 patients with benign cholecystitis was clear.
None of the patients with gallbladder carcinoma that participated
in the present study had received radiotherapy, chemotherapy or any
other treatment prior to surgery. The Clinical Research Ethics
Committee of Chinese PLA General Hospital and China-Japan
Friendship Hospital approved the present study. All patients
provided written informed consent for the use of their medical
records and tissue specimens for research purposes.
RNA isolation and
reverse-transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated from tissue and GBC-SD cells
by using the Qiazol reagent (Qiagen GmbH). The concentration of
total RNA was determined via A260 measurement using the ND-1000
NanoDrop spectrophotometer (NanoDrop; Thermo Fisher Scientific,
Inc). A total of 500 ng total RNA was reverse transcribed using the
High Capacity cDNA Reverse Transcription kit (Thermo Fisher
Scientific, Inc.). The RT temperature protocol was 25°C for 10 min,
37°C for 120 min and 85°C for 5 min, followed by maintenance at
4°C. A total of 10 ng cDNA were used for qPCR using Power SYBR
Green PCR Master Mix (Applied Biosystems; Thermo Fisher Scientific,
Inc.), as per the manufacturer's instructions. The thermocycling
conditions were as follows: Initial denaturation at 95°C for 10
min; followed by 55 cycles of denaturation at 95°C for 30 sec and
60°C for 1 min; finally, annealing and elongation was performed by
increasing the temperature from 55°C to 95°C (30 sec/degree
increase). The primer sequences were as follows: MPZL1, forward
5′-TGGGCTGGAGACCTTGAC-3′ and reverse 5′-CCCACCACTACCCAAACTG-3′;
β-actin, forward 5′-CTTAGTTGCGTTACACCCTTTCTTG-3′ and reverse
5′-CTGTCACCTTCACCGTTCCAGTTT-3′. The experiment was repeated three
times and transcript levels were normalised to β-actin levels.
Transcript expression levels were calculated using the
2−ΔΔCq method (19).
mRNA microarray analysis
Total RNA (100 ng) from frozen paired carcinoma and
para-carcinoma tissues (n=4, samples selected from the 20 patients
cohort) were used for the expression analysis using Prime View
Human Gene Expression Array (Affymetrix; Thermo Fisher Scientific,
Inc.), according to the manufacturer's instructions. Briefly, the
GeneChip™ 3IVT labeling kit (Affymetrix; Thermo Fisher Scientific,
Inc.) was employed to label total RNA with biotin. To select genes
for microarray analysis, the GSE62335 microarray data were
downloaded from the Gene Expression Omnibus database, which was
deposited by Ma et al in 2014 (20). According to the expression profile
database, genes related to pathways and transcriptional
misregulation in cancer were selected. The chips were hybridized
overnight with, washed and stained using the GeneChip™
Hybridization, Wash and Stain kit (Affymetrix; Thermo Fisher
Scientific, Inc.). The chip was scanned using the GeneChip™ Scanner
3000 (Affymetrix; Thermo Fisher Scientific, Inc.). Gene Ontology
(GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway
analyses were used to detect differentially expressed genes. Genes
were selected for further evaluation if fold change was >2 and
P<0.05.
Immunohistochemical analysis
A total of 82 paraffin fixed carcinoma tissues from
gallbladder carcinoma patients were used for the
immunohistochemical analysis. The 20 paired frozen samples of
gallbladder carcinoma and 20 frozen gallbladder tissues in normal
control group were also used for immunohistochemical analysis. The
tissue sections were deparaffinized in xylene twice for 5 min at
room temperature, and rehydrated using a graded ethanol series
(100, 95, 80 and 70%; 5 min at room temperature for each
concentration). Subsequently, the endogenous peroxidase activity
was blocked by soaking in 0.3% hydrogen peroxide for 10 min at room
temperature. Thereafter, the sections were processed in 10 mmol/l
citrate buffer (pH 6.0) and were heated to 121°C in an autoclave
for 20 min for antigen retrieval. After rinsing with PBS (pH 7.2),
10% goat serum (Invitrogen; Thermo Fisher Scientific, Inc.) was
added and incubated at room temperature for 1 h to block
non-specific reactions. The sections were then incubated overnight
at 4°C with MPZL1 monoclonal antibody (cat. no. ab151541; diluted
1:100; Abcam). Subsequently, 25 µl horseradish peroxidase
(HRP)-labeled secondary antibody (goat anti-rabbit IgG H&L HRP;
cat. no. ab205718; diluted 1:100; Abcam) was added to each section
for 30 min at room temperature. Each section was then soaked in PBS
for 5 min and rinsed three times. All slides were processed using
the peroxidase anti-peroxidase method. After rinsing with PBS, the
peroxidase reaction was visualized by incubating the sections with
3,3-diamino-benzidine tetrahydrochloride for 5 min at room
temperature. The sections were then rinsed with water,
counterstained with hematoxylin (5%) for 1 min at room temperature,
dehydrated and coverslipped. All of the immunostained sections were
evaluated in a blinded manner by two independent and experienced
observers without any knowledge on the clinicopathological features
of the patients. Five fields (magnification, ×400) of each specimen
were selected randomly, and nuclear staining was examined using a
light microscope. For the interpretation of MPZL1 staining results,
the nuclear MPZL1 expression was scored as a percentage of positive
cells by two pathologists. Carcinoma tissues, paracarcinoma tissues
and gallbladder tissues in the normal control group showing nuclear
MPZL1 expression in ≥5% of cells were considered to be
MPZL1-positive.
Cell culture
The human gallbladder carcinoma GBC-SD cell line was
purchased from the Type Culture Collection of the Chinese Academy
of Sciences (Shanghai, China). Cells were maintained in RPMI-1640
medium supplemented with 10% heat-inactivated fetal calf serum, 2
mM L-glutamine, and 100 U/ml penicillin and streptomycin mixture
(all from Gibco; Thermo Fisher Scientific, Inc.), and incubated at
37°C with 5% CO2.
Western blot analysis
Tissue specimens and GBC-SD cell were lysed in the
lysis buffer (1 M Tris-HCl at pH 7.5, 1% Triton X-100, 1% Nonidet
P-40, 10% SDS, 0.5% sodium deoxycholate, 0.5 M EDTA, 10 µg/ml
leupeptin, 10 µg/ml aprotinin and 1 mM phenylmethylsulfonyl
fluoride) and then centrifuged at 4°C and 10,000 × g for 30 min to
collect the supernatant. Protein concentration was determined using
the Bio-Rad protein assay kit (Bio-Rad Laboratories, Inc.). Equal
amounts (60 µg/lane) of protein were separated by SDS-PAGE on 10%
gels, and transferred to polyvinylidine difluoride (PVDF) membranes
(EMD Millipore). Membranes were then blocked with 5% non-fat milk
in TBS with 0.05% Tween-20 for 2 h at room temperature.
Subsequently, the membrane was incubated for 12 h at 4°C with
primary antibodies, MPZL1 monoclonal antibody (cat. no. ab151541;
diluted 1:100; Abcam) for 1 h at room temperature. In addition,
β-actin antibody (cat. no. WL01845; WanleiBio) diluted in 5%
non-fat milk (1:1,000) was used to incubate the PVDF membrane at
4°C overnight. Subsequently, the PVDF membrane was washed in TBS
under agitation for 5 min; this step was repeated four times. The
membrane was then incubated with a secondary antibody (goat
anti-rabbit IgG HRP; cat. no. WLA023; diluted 1:5,000; WanleiBio)
diluted in 5% non-fat milk at 37°C for 45 min. Immunoblotting bands
were visualized via enhanced chemiluminescence (NEN Life Science
Products; PerkinElmer, Inc.). The band density was measured using a
computer-assisted image analysis system (Image Lab Software version
5.2.x; Bio-Rad Laboratories, Inc.), and band densities were
normalized against the β-actin levels.
MPZL1 overexpression and silence
TRIzol® was used to extract total RNA
from normal gallbladder epithelium. Total RNA was the reverse
transcribed using the High Capacity cDNA Reverse Transcription kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.) to obtain
human tissue cDNA library. The primer sequences used to amplify the
MPZL1 gene containing the sites for HindIII and XbaI
were: Forward 5′-AAGCTTCAGGTGGCGGAGAGATCAGAAG-3′ and reverse
5′-TCTAGAGACTTGTCCTTGCCTGGGTCTC-3′. PCR was then performed to
amplify the cDNA sequence with restriction sites using Platinum Taq
DNA Polymerase (cat. no. 10966034, Invitrogen; Thermo Fisher
Scientific, Inc.); the thermocycling conditions were the same as
for RT-qPCR., Subsequently 1% agarose gel electrophoresis was
performed to recover and purify the amplified cDNA fragment of
interest, and DNA was visualized using a gel imaging system (E-Gel
Imager; Invitrogen; Thermo Fisher Scientific, Inc.). After
enzymatic digestion and purification of the PCR amplification
product, it was cloned into a pCMV plasmid vector (cat. no. V51020;
Invitrogen; Thermo Fisher Scientific, Inc.) using a T4 DNA ligase
to connect the target sequence to the pCMV vector, thus generating
the pCMV-MPZL1 high expression vector. After transforming DH5α
competent cells (cat. no. 18263012 Invitrogen; Thermo Fisher
Scientific, Inc.) with the pCMV-MPZL1, a single clone was selected
and the correct construction of the vector was ensured by
sequencing.
Three pairs of small interfering RNA (siRNA) probes
targeting the MPZL1 mRNA were designed (WanleiBio) using the
chemical synthesis method (21),
and their transfection efficiency was verified. One pair of siRNA
molecules that was the most efficient at silencing the MPZL1 mRNA
(siRNA-1: 5′-GCACCUAUAUCUGUGAUGUTT-3′; siRNA-2:
5′-GUCAGAGUCUGUGGUGUAUTT-3′; siRNA-3: 5′-GAGAGUUUGUCACCAGUUATT-3′)
was selected. The pCMV-MPZL1 vector, blank vector, siMPZL1 and
scramble RNA (5′-CCCACCACTACCCAAACTG-3′) were transfected into
GBC-SD cells using Lipofectamine 2000. The concentration of
transfected DNA was 0.02 µg/µl and a total of 100 µl was added to
GBC-SD cells. Subsequent experiments were performed 16 h
post-transfection.
Cell proliferation assay
Cell proliferation was measured using Cell Counting
Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.), following the
manufacturer's instructions. Briefly, GBC-SD cells were seeded onto
96-well cell culture plates at a concentration of 2×104
cells/well in 100 µl fresh complete medium (cat. no. 12558011;
AmnioMax™ C100; Gibco). CCK-8 reagent (10 µl) was added to a subset
of wells and incubated for 2 h at 37°C, and the absorbance was
measured using an automated plate reader under the detection
wavelength of 450 nm. Each measurement was performed in triplicate
and the experiments were repeated twice.
Cell migration and invasion assay
Matrigel gel was thawed overnight at 4°C. The
Matrigel gel was placed on ice in a clean bench and diluted 1:3 in
serum-free medium. The Transwell chamber were placed in 24-well
plates, 40 µl pre-diluted Matrigel were added to the chamber
membrane and placed in a 37°C incubator for 2 h. The transfected
GBC-SD cells were cultured in 6-well plates (2×104
cells/well) at 37°C in an atmosphere containing 5% CO2
and saturated humidity for 48 h. The culture medium from each group
of cells was discarded, cells were washed three times with PBS
before being detached with 0.25% trypsin. Serum-free medium was
then added to create a single cell suspension. A total of 800 µl
culture medium containing 30% FBS was added to lower chamber of the
24-well Matrigel-coated Transwell, and 200 µl cell suspension
(2×104 cells/well) were added to the upper chamber. The
24-well plate was incubated in a cell incubator at 37°C under 5%
CO2 and saturated humidity for 12 h. Subsequently, the
lower chamber was washed twice with PBS, fixed with 4%
paraformaldehyde at room temperature for 20 min, stained with 0.5%
crystal violet dye at room temperature for 5 min, and rinsed with
distilled water. The cells that migrated to the lower layer of the
micro-porous membrane were counted under an inverted light
microscope (magnification, ×200). The number of cells in each
sample was the average number of cells across five fields.
Cell apoptosis assay
GBC-SD cell apoptosis was evaluated using an Annexin
V-FITC/propidium iodide (PI) staining assay. Flow cytometry
analysis of apoptotic cells was performed using an Annexin
V-FITC/PI staining kit (BD Biosciences). After washing with cold
PBS, the cells were re-suspended in binding buffer (100 mM HEPES,
pH 7.4, 100 mM NaCl and 25 mM CaCl2) followed by
staining with Annexin V-FITC/PI at room temperature in the dark for
15 min. Apoptotic cells were then evaluated by gating PI and
Annexin V positive cells on a FACSCalibur (BD Biosciences); data
were analyzed using CellQuest Pro software, version 5.1 (BD
Biosciences). Annexin V was set in the horizontal axis and PI was
set in the vertical axis. Early apoptotic cells were located in the
lower right quadrant of the flow cytometry plot.
Statistical analysis
Statistical analyses were performed using SPSS
software (version 20.0, IBM Corp.). Categorical variables were
compared using a χ2 or Fisher's exact test. A Student's
t test was applied for continuous variables with a normal
distribution. Multiple groups were compared using the
Bonferroni-corrected one-way ANOVA test. For paired samples, a
paired Student's t-test was applied. Bonferroni-corrected Student's
t-test was used to compare the paired carcinoma and paracarcinoma
tissues with control tissues. P<0.05 was considered to indicate
a statistically significant difference.
Results
Results of mRNA microarray analysis of
gallbladder carcinoma tissues and immunohistochemical analysis in
different Tumor-Node-Metastasis (TNM) stages
Of the 20,000 genes that were detected, genes were
selected for further evaluation if the fold change was >2, and
P<0.05. Using these criteria, 580 upregulated and 999
downregulated genes were identified in gallbladder carcinoma
tissues. Through GO analysis and KEGG pathway analysis of
differentially expressed genes, a total of 18 genes involved in
tyrosine phosphorylation were identified, of which 2 were
downregulated and 16 were upregulated, including MPZL1.
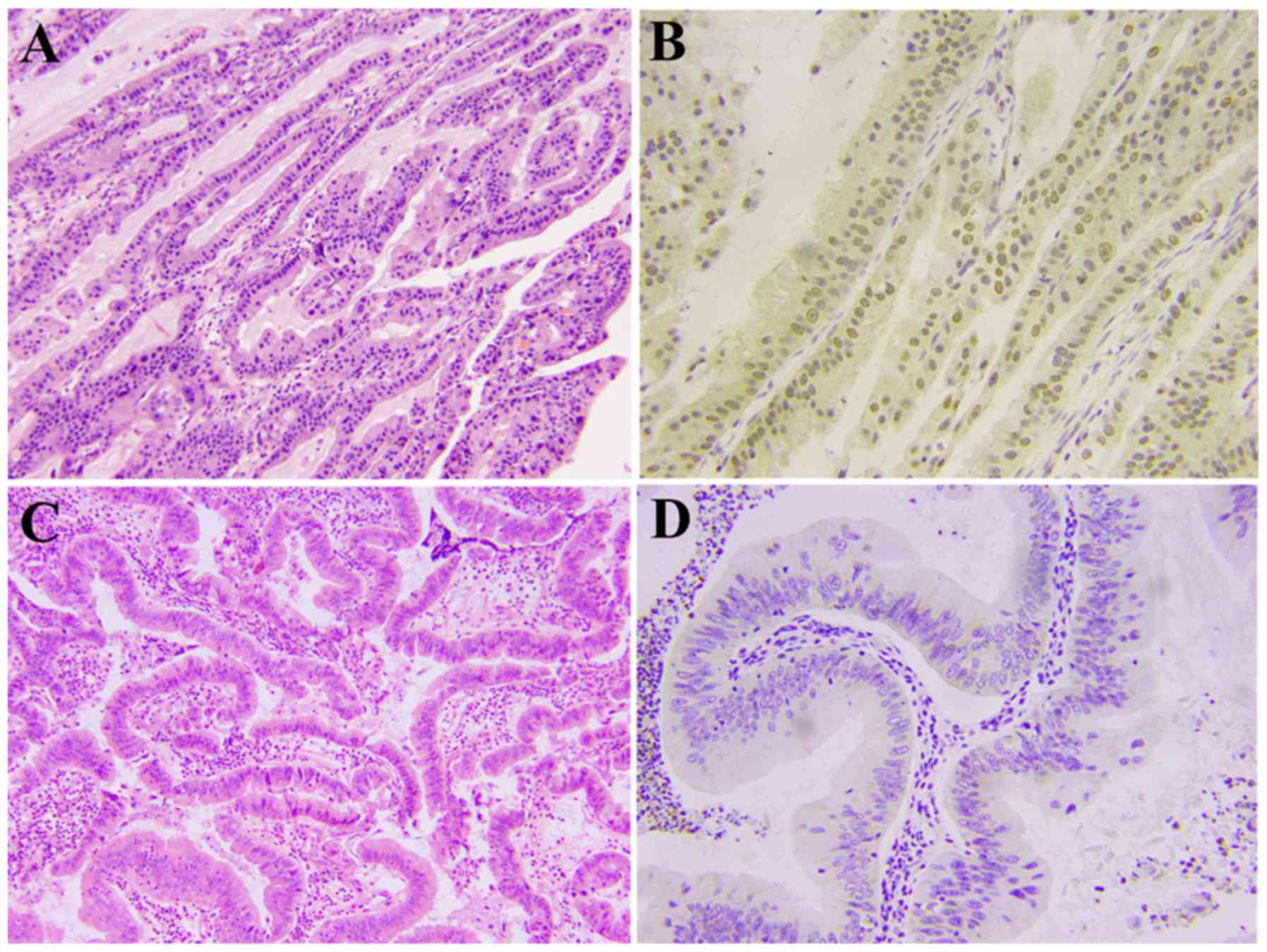
A total of 82 paraffin fixed tissues from
gallbladder carcinoma patients were collected for the
immunohistochemical analysis. The final pathological results of TNM
staging were also evaluated. The positive rates of MPZL1 across the
different T stages were 30.8% (4/13, T2), 55.9% (19/34, T3) and
77.1% (27/35, T4), respectively and the difference between these
groups was significant (P<0.01). The positive rates of MPZL1
across the different N stages were 32.3% (10/31, N negative) and
78.4% (40/51, N positive) and the difference between them was also
significant (P<0.01; Table I;
Fig. 1). The immunohistochemical
analysis results of 20 paracarcinoma tissues and 20 normal
gallbladder epithelial tissues were all negative for MPZL1
expression.
 | Table I.Results of immunohistochemical
analysis of samples from patients with gallbladder carcinoma. |
Table I.
Results of immunohistochemical
analysis of samples from patients with gallbladder carcinoma.
| A, T stage
(n=82) |
|---|
|
|---|
|
| Number of
patients |
|
|---|
|
|
|
|
|---|
| Group | All | MPZL1 (−) | MPZL1 (+) | P-value |
|---|
| II | 13 | 9 | 4 | 0.010 |
| III | 34 | 15 | 19 |
|
| IV | 35 | 8 | 27 |
|
|
| B, N stage
(n=82) |
|
|
| Number of
patients |
|
|
|
|
|
| Group | All | MPZL1
(−) | MPZL1
(+) | P-value |
|
| Negative | 31 | 21 | 10 | <0.001 |
| Positive | 51 | 11 | 40 |
|
MPZL1 mRNA and protein are highly
expressed in gallbladder carcinoma tissues
The 20 cases of gallbladder carcinoma and paired
paracarcinoma tissues were evaluated, and gallbladder tissue
specimens from patients with cholecystitis were also included as
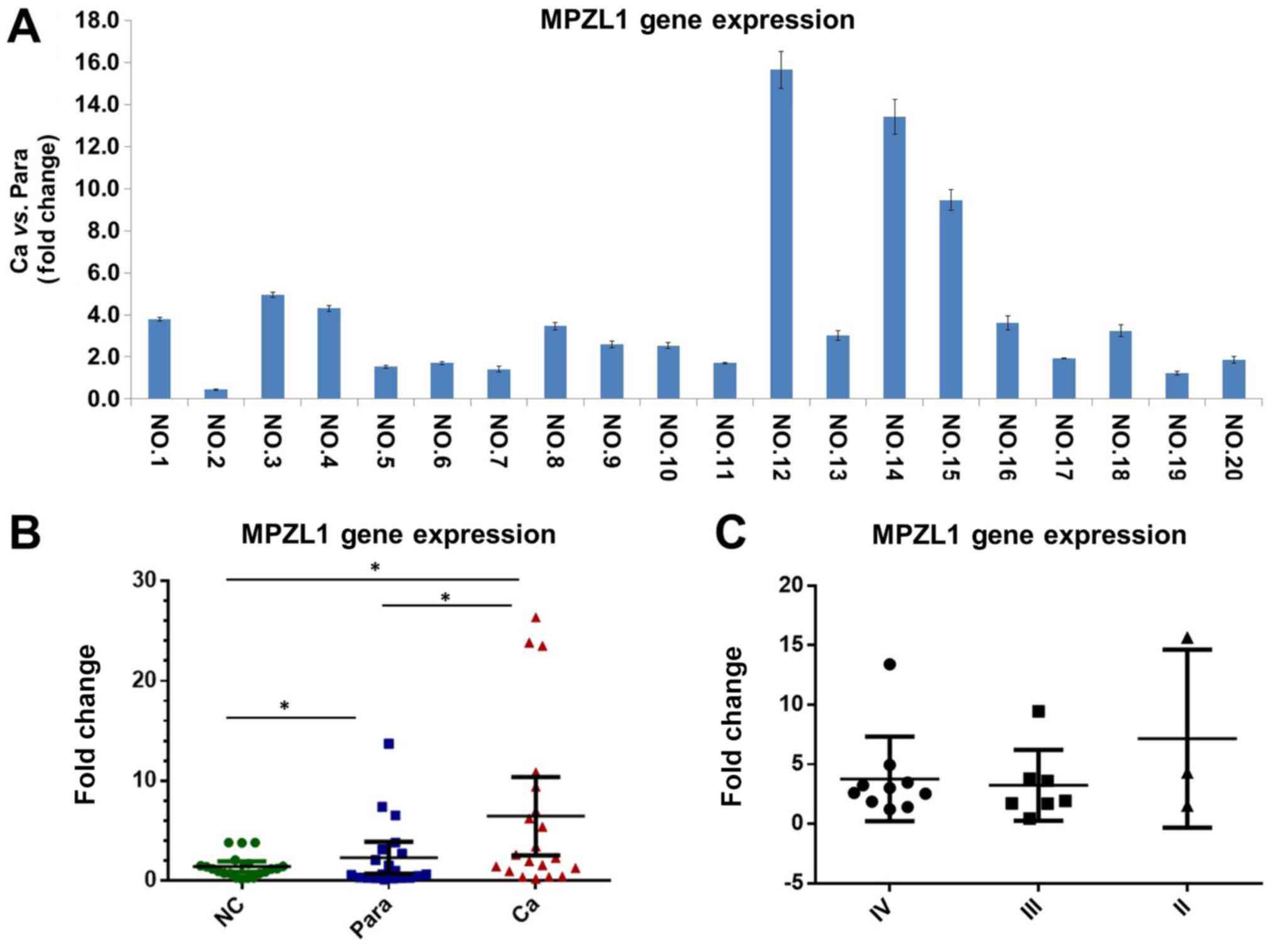
normal controls. The results of RT-qPCR suggested that the
expression of MPZL1 gene in gallbladder carcinoma tissues was
significantly higher than that in paracarcinoma tissues (Fig. 2A). Meanwhile, the expression of
MPZL1 gene in normal gallbladder epithelial tissue was
significantly lower than that in gallbladder carcinoma tissues and
paracarcinoma tissues (P<0.05; Fig.
2B). There was no significant difference in the expression
level of the MPZL1 gene in gallbladder carcinoma tissues with
different TNM staging (Fig.
2C).
The results of the western blotting showed that the
expression of MPZL1 protein in gallbladder carcinoma tissues was
significantly higher than that of paired paracarcinoma tissues and
randomly matched normal gallbladder epithelial tissues.
Densitometry analysis of the western blot results showed that the
expression of MPZL1 protein in gallbladder carcinoma was higher
compared with matched paired adjacent tissues (P<0.05) and
control gallbladder epithelial tissues (P<0.05), while the
levels of MPZL1 protein in the paracarcinoma tissues were not
markedly different from that of control tissues (P=0.23) (Fig. 3A). According to the different TNM
staging classification, the protein levels of MPZL1 protein in
stage IV gallbladder carcinoma were higher than that in stage III
gallbladder carcinoma (P<0.05; Fig.
3B). According to the T stage of gallbladder carcinoma, it was
also observed that the levels of MPZL1 protein in the T4 stage were
also higher than those in the T3 stage (P<0.05; Fig. 3C).
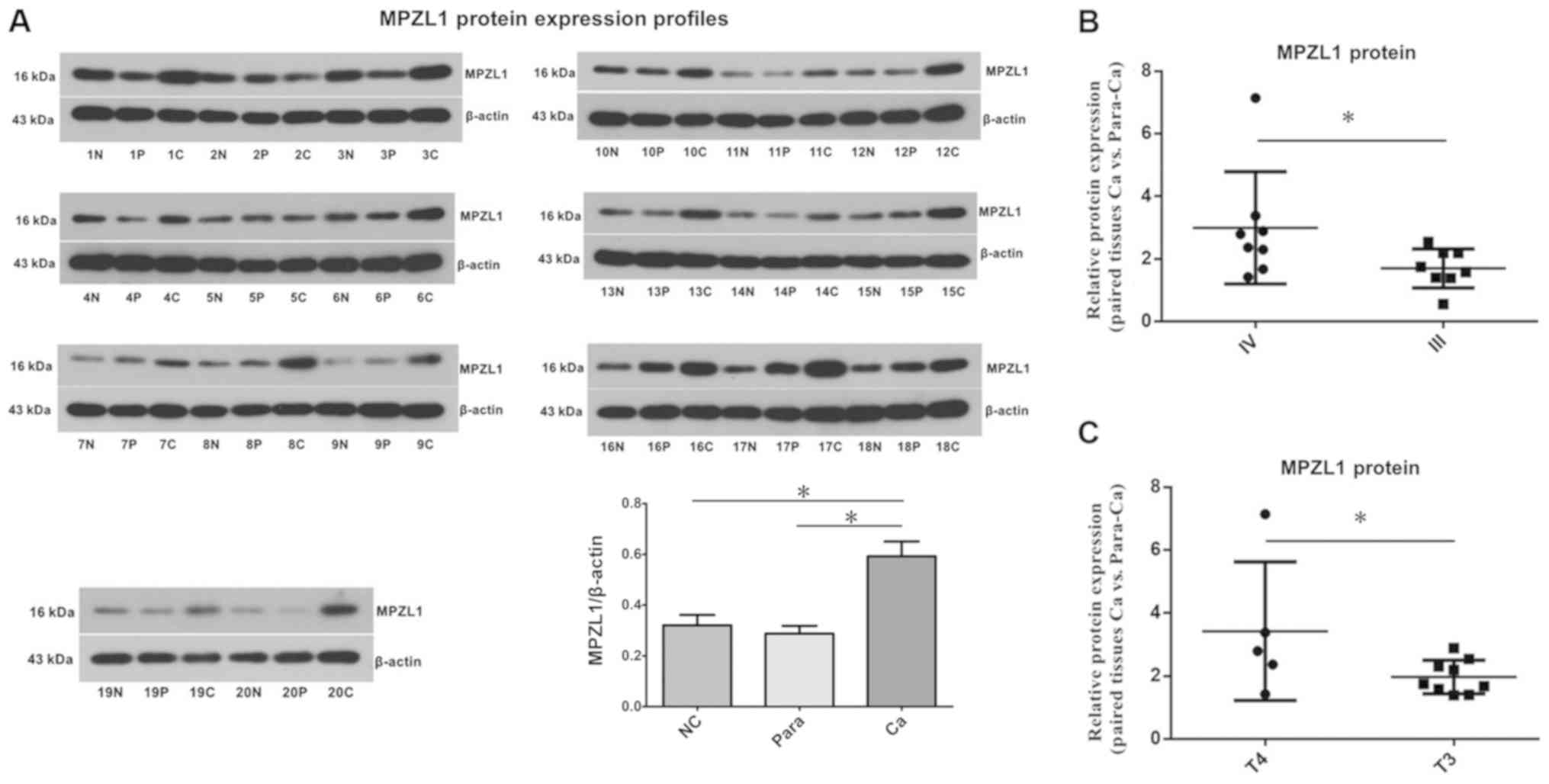 | Figure 3.Protein levels of MPZL1 in normal,
carcinoma and matched paraneoplastic gallbladder tissues. (A)
Western blot analysis of MPZL1 protein in carcinoma, paired
paracarcinoma and normal gallbladder tissue samples, using β-actin
as an internal reference. (B) Densitometry analysis of MPZL1
protein levels in gallbladder carcinoma tissue specimens of stage
III and stage IV according to Tumor-Node-Metastasis classification.
(C) Densitometry analysis of MPZL1 protein levels in gallbladder
carcinoma tissue samples of stages T4 and T3. *P<0.05. MPZL1,
myelin protein 0-like 1; N, normal gallbladder epithelium
(control); P, matched paracarcinoma gallbladder sample; C,
gallbladder carcinoma sample; Ca, carcinoma; Para-Ca,
paracarcinoma; NC, normal control group. |
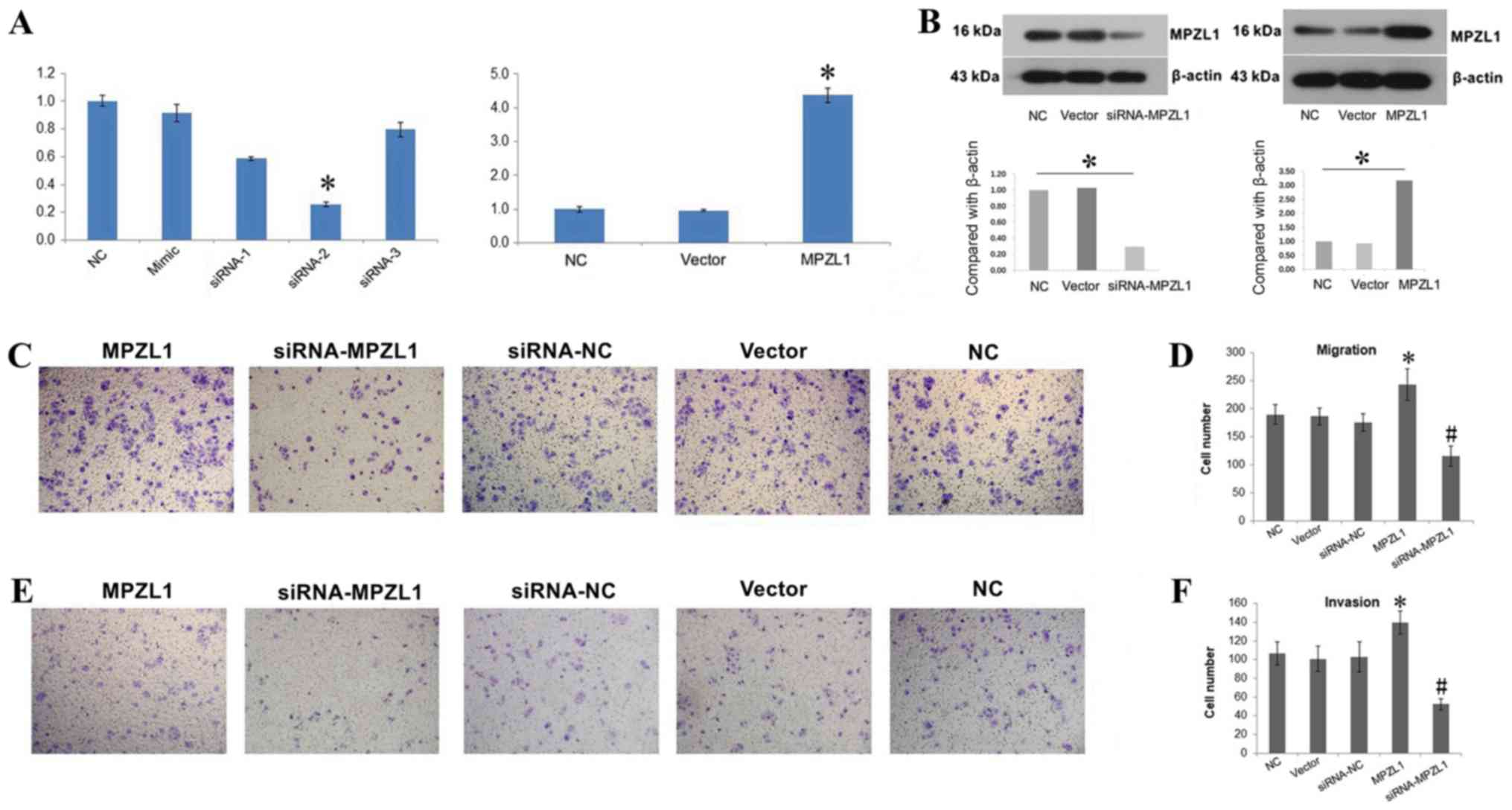
MPZL1 overexpression facilitates the
invasion and metastasis of GBC-SD cells
An overexpression vector and MPZL1 siRNA were used
to change the expression of MPZL1 gene in the gallbladder carcinoma
cell line GBC-SD. RT-qPCR and western blotting methods were
subsequently used to verify whether the gene expression and protein
levels of MZPL1 in GBC-SD cells, respectively, were affected by the
overexpression vector and siRNA. Of the three siRNAs tested,
siRNA-2 exhibited the highest efficiency and was therefore selected
for subsequent experiments. The results indicated that the
expression levels of MPZL1 mRNA and protein in GBC-SD cells
transfected with the MPZL1 overexpression vector were increased
(P<0.05), and the levels of MPZL1 mRNA and protein in GBC-SD
cells transfected with MPZL1 siRNA were decreased (P<0.05)
(Fig. 4A and B).
Both GBC-SD cells overexpressing or underexpressing
MPZL1 were evaluated in Transwell migration and invasion
experiments. The results showed that enhanced expression of MPZL1
gene increased the number of migrating GBC-SD cells. Conversely,
the downregulation of MPZL1 via siRNA led to a decrease in the
number of migrating GBC-SD cells (Fig.
4C and D). Regarding the Transwell invasion experiments, the
results confirmed that, compared with normal GBC-SD cells,
overexpression of MPZL1 gene increased the invasive ability of
GBC-SD cells, while MPZL1 downregulation decreased their invasive
ability (Fig. 4E and F).
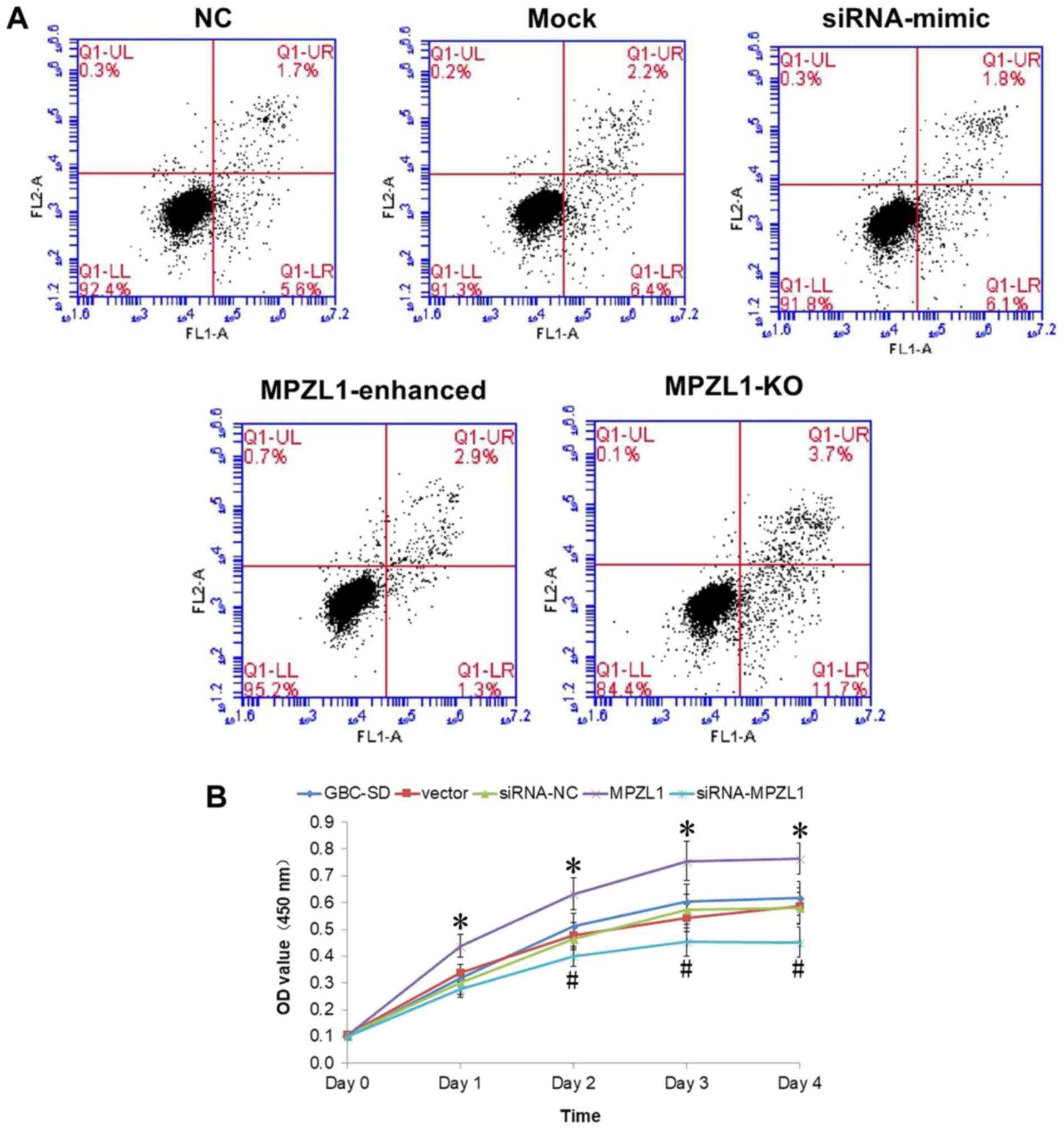
Enhancement of MPZL1 expression is
promotes the proliferation and survival of GBC-SD cells
GBC-SD cells overexpressing and downregulating MPZL1
were further tested in cell proliferation experiments. The results
showed that after siRNA transfection against MPZL1, the
proliferation of GBC-SD cells was significantly impaired
(P<0.05). By contrast, MPZL1 overexpression significantly
increased the proliferation ability of GBC-SD cells (P<0.05)
(Fig. 5B). The results of flow
cytometry analysis showed that only 1.3% of MPZL1 overexpressing
cells were in the early apoptosis stage (Q1-LR). On the other hand,
a total of 11.7% of GBC-SD cells with transfected with MPZL1 siRNA
were in the early apoptosis stage. In addition, the rate of normal
control cells in the early apoptosis stage was 5.6%. The proportion
of GBC-SD cells at the advanced apoptosis stage following MPZL1
overexpression was 2.9%, the proportion of cells in the advanced
apoptotic stage following MPLZ1 knockdown was 3.7% (Fig. 5A).
Discussion
Gallbladder carcinoma ranks as the gastrointestinal
cancer with the sixth highest incidence, with an annual incidence
rate of 2.2 per 100,000 people, and a mortality rate of 1.7 per
100,000 (1). Recent studies have
also confirmed the involvement of Hedgehog, PI3K/AKT/mTOR, Notch,
ErbB, MAPK/ERK and other signaling pathways in gallbladder
carcinoma development and progression (4,22–25).
However, the specific molecular mechanisms underlying gallbladder
carcinoma remain unknown, and further research is required.
Tyrosine phosphorylation, which has an important
role in the proliferation and migration of tumor cells, is a
reversible and dynamic process that requires the participation of
PTKs and PTPs (26). Studies have
also identified mutations in PTPs followed by changes in their
respective signaling pathways in a number of tumor types (27–30).
Although no PTP-related drugs are yet available, studies have
confirmed that PTP-associated factors, such as SHP2, dual
specificity protein phosphatase 1, acid phosphatase 1 and dual
specificity phosphatases have important tumor-promoting effects,
and may therefore be important anti-tumor targets (31–33).
However, to the best of our knowledge, there are no reports on the
role of PTP pathway-associated genes in gallbladder carcinoma.
Therefore, through a microarray gene expression analysis of four
gallbladder carcinoma tissue samples, the present study showed that
there were eight upregulated genes in gallbladder carcinoma tissues
associated with the PTP pathway, including MPZL1.
MPZL1 belongs to the immunoglobulin superfamily,
containing an extracellular domain, and significant sequence
homology with myelin P0 (5). There
are two immunoreceptor ITIMs in the intracellular portion that
specifically interact with SHP2 (6). Studies have also confirmed that SHP2
is a tumor promoting PTP (10–12),
involved in the proliferation, differentiation and survival of
tumor cells through a variety of signaling pathways, including
Ras/Raf/MEK/ERK, JAK-STAT, PI3K/AKT/mTOR and Wnt/β-catenin
(7,13,34,35).
Studies have also found that MPZL1 can promote
fibronectin-dependent cell migration process by recruiting and
activating SHP2 (8). A study has
also found that the expression of MPZL1 was upregulated in liver
cancer tissues, and through cortactin activation, was able to
promote the migration and metastasis of hepatoma cells through the
Src-signaling pathway (22).
Recently, other studies have documented a role for cortactin in the
promotion of cancer cell motility and invasion, including a
critical role in the development of invadopodia, actin-rich
subcellular protrusions associated with the degradation of the
extracellular matrix by cancer cells (36–38).
Moreover, MPZL1 has also been shown to activate Src kinase upon
stimulation with extracellular stimuli (39,40).
However, prior to the present study, the role of MPLZ1 in
gallbladder carcinoma remained unknown.
The present study verified that high levels of MPZL1
in gallbladder carcinoma tissues at the gene and protein levels via
RT-qPCR and western blotting. These suggested that the expression
of MPZL1 gene in gallbladder carcinoma tissues was significantly
higher than that in paracarcinoma tissues and normal gallbladder
epithelial tissues. The reason as to why some cases of
paracarcinoma tissues showed high expression of MPZL1 mRNA may be
associated with the proximity of the sampled tissue to the
carcinoma area or with the biological complexity of the samples.
The western blotting results also showed that MPZL1 protein levels
were higher in gallbladder carcinoma tissues than paracarcinoma
tissues and randomly matched normal gallbladder epithelial tissues.
The levels of MPZL1 protein in T4 stage samples were seemingly
higher than that the levels at the T3 stage. The results of
immunohistochemical analysis also confirmed that patients with
advanced gallbladder carcinoma were more likely to have higher
levels of MPZL1 positive cells, which indicated that MPZL1 may be
involved in the invasion and metastasis of gallbladder carcinoma
cells. The result of RT-qPCR failed to detect a difference in
different TNM stages, which did not match the results of western
blot and immunohistochemical analyses. The reason for this may be
the limited number of samples. Further experiments with a larger
group may be helpful to clarify this issue. In fact, MPZL1
overexpression in GBC-SD cells confirmed that the enhanced
expression of MPZL1 gene was beneficial GBC-SD cell migration and
invasion. Conversely, MPZL1 knockdown in the same cell line led to
a decreased migration and invasion ability. Moreover, the
proliferation ability of GBC-SD cells was reduced following MPLZ1
knockdown, and the enhancement of MPZL1 gene expression promoted
cell viability. Lastly, the flow cytometry analysis revealed that
MPZL1 overexpression GBC-SD cells appeared to reduce the early
activation of the apoptotic pathway, promoting cell survival.
In conclusion, the present findings revealed that
the levels of MPZL1 gene and protein were significantly higher in
gallbladder carcinoma tissues, particularly in patients with
advanced tumors. Overexpression of the MPZL1 gene also promoted
cancer GBC-SD cell invasion, metastasis, proliferation and
antiapoptotic ability. Further studies should focus on the possible
signaling pathways associated with MPZL1 to uncover the molecular
mechanisms underlying the early metastasis of gallbladder
carcinoma.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and RL conceptualized the study design and wrote
the manuscript. XL, JH and LL performed the experiments and
analyzed the data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by both Ethics
Committee of Chinese PLA General Hospital and China-Japan
Friendship Hospital (Beijing, China). All patients provided written
informed consent for the use of their medical records and tissue
specimens for research purposes.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MPZL1
|
myelin protein 0-like 1
|
|
ITIMs
|
immunoreceptor tyrosine-based
inhibitory motifs
|
|
SHP2
|
Src homology phosphatase type-2
|
|
PTP
|
protein tyrosine phosphatases
|
|
CCK
|
Cell Counting Kit
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
References
|
1
|
Bizama C, García P, Espinoza JA, Weber H,
Leal P, Nervi B and Roa JC: Targeting specific molecular pathways
holds promise for advanced gallbladder cancer therapy. Cancer Treat
Rev. 41:222–234. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Randi G, Malvezzi M, Levi F, Ferlay J,
Negri E, Franceschi S and La Vecchia C: Epidemiology of biliary
tract cancers: An update. Ann Oncol. 20:146–155. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goetze TO: Gallbladder carcinoma:
Prognostic factors and therapeutic options. World J Gastroenterol.
21:12211–12217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hundal R and Shaffer EA: Gallbladder
cancer: Epidemiology and outcome. Clin Epidemiol. 6:99–109.
2014.PubMed/NCBI
|
|
5
|
Yeh YT, Dai HY and Chien CY: Amplification
of MPZL1/PZR gene in hepatocellular carcinoma. Hepatobiliary Surg
Nutr. 3:87–90. 2014.PubMed/NCBI
|
|
6
|
Zhao ZJ and Zhao R: Purification and
cloning of PZR, a binding protein and putative physiological
substrate of tyrosine phosphatase SHP-2. J Biol Chem.
273:29367–29372. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taniguchi K and Karin M: IL-6 and related
cytokines as the critical lynchpins between inflammation and
cancer. Semin Immunol. 26:54–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zannettino AC, Roubelakis M, Welldon KJ,
Jackson DE, Simmons PJ, Bendall LJ, Henniker A, Harrison KL, Niutta
S, Bradstock KF and Watt SM: Novel mesenchymal and haematopoietic
cell isoforms of the SHP-2 docking receptor, PZR: Identification,
molecular cloning and effects on cell migration. Biochem J.
370:537–549. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Zhang F and Niu R: Functions of
Shp2 in cancer. J Cell Mol Med. 19:2075–2083. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Labbé DP, Hardy S and Tremblay ML: Protein
tyrosine phosphatases in cancer. Friends and foes! Prog Mol Biol
Transl Sci. 106:253–306. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang WQ, Lin Q, Zhuang X, Cai LL, Ruan
RS, Lu ZX and Tzeng CM: Structure, function, and pathogenesis of
SHP2 in developmental disorders and tumorigenesis. Curr Cancer Drug
Targets. 14:567–588. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chan G, Kalaitzidis D and Neel BG: The
tyrosine phosphatase Shp2 (PTPN11) in cancer. Cancer Metastasis
Rev. 27:179–192. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Grossmann KS, Rosário M, Birchmeier C and
Birchmeier W: The tyrosine phosphatase Shp2 in development and
cancer. Adv Cancer Res. 106:53–89. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roubelakis MG, Martin-Rendon E, Tsaknakis
G, Stavropoulos A and Watt SM: The murine ortholog of the SHP-2
binding molecule, PZR accelerates cell migration on fibronectin and
is expressed in early embryo formation. J Cell Biochem.
102:955–969. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Seda Eminaga and Anton M: Bennett: Noonan
syndrome-associated SHP-2/Ptpn11 mutants enhance SIRPα and PZR
tyrosyl phosphorylation and promote adhesion-mediated ERK
activation. J Biol Chem. 283:15328–15338. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kusano K, Thomas TN and Fujiwara K:
Phosphorylation and localization of protein-zero related (PZR) in
cultured endothelial cells. Endothelium. 15:127–136. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia D, Jing Y, Zhang Z, Liu L, Ding J,
Zhao F, Ge C, Wang Q, Chen T, Yao M, et al: Amplification of
MPZL1/PZR promotes tumor cell migration through Src-mediated
phosphorylation of cortactin in hepatocellular carcinoma. Cell Res.
24:204–217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu T, Liang L, Zhao X and Yin Y:
Structural and biochemical studies of the extracellular domain of
Myelin protein zero-like protein 1. Biochem Biophys Res Commun.
506:883–890. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma MZ, Kong X, Weng MZ, Zhang MD, Qin YY,
Gong W, Zhang WJ and Quan ZW: Long non-coding RNA-LET is a positive
prognostic factor and exhibits tumor-suppressive activity in
gallbladder cancer. Mol Carcinog. 54:1397–1406. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Caplen NJ, Parrish S, Imani F, Fire A and
Morgan RA: Specific inhibition of gene expression by small
double-stranded RNAs in invertebrate and vertebrate systems. Proc
Natl Acad Sci USA. 98:9742–9747. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yoon HA, Noh MH, Kim BG, Han JS, Jang JS,
Choi SR, Jeong JS and Chun JH: Clinicopathological significance of
altered Notch signaling in extrahepatic cholangiocarcinoma and
gallbladder carcinoma. World J Gastroenterol. 17:4023–4030. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bao RF, Shu YJ, Hu YP, Wang XA, Zhang F,
Liang HB, Ye YY, Li HF, Xiang SS, Weng H, et al: miR-101 targeting
ZFX suppresses tumor proliferation and metastasis by regulating the
MAPK/Erk and Smad pathways in gallbladder carcinoma. Oncotarget.
7:22339–22354. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang P, Guo Z, Wu Y, Hu R, Du J, He X,
Jiao X and Zhu X: Histone deacetylase inhibitors inhibit the
proliferation of gallbladder carcinoma cells by suppressing
AKT/mTOR signaling. PLoS One. 10:e01361932015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xie F, Xu X, Xu A, Liu C, Liang F, Xue M
and Bai L: Aberrant activation of Sonic hedgehog signaling in
chronic cholecystitis and gallbladder carcinoma. Hum Pathol.
45:513–521. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hunter T: Tyrosine phosphorylation: Thirty
years and counting. Curr Opin Cell Biol. 21:140–146. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hunter T: Protein kinases and
phosphatases: The yin and yang of protein phosphorylation and
signaling. Cell. 80:225–236. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Krause DS and Van Etten RA: Tyrosine
kinases as targets for cancer therapy. N Engl J Med. 353:172–187.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao S, Sedwick D and Wang Z: Genetic
alterations of protein tyrosine phosphatases in human cancers.
Oncogene. 34:3885–3894. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gaumann AK, Kiefer F, Alfer J, Lang SA,
Geissler EK and Breier G: Receptor tyrosine kinase inhibitors: Are
they Real Tumor Killers? Int J Cancer. 138:540–554. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tonks NK: Protein tyrosine
phosphatases-from housekeeping enzymes to master regulators of
signal transduction. FEBS J. 280:346–378. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Julien SG, Dubé N, Hardy S and Tremblay
ML: Inside the human cancer tyrosine phosphatome. Nat Rev Cancer.
11:35–49. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
He RJ, Yu ZH, Zhang RY and Zhang ZY:
Protein tyrosine phosphatases as potential therapeutic targets.
Acta Pharmacol Sin. 35:1227–1246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li M, Zhang Z, Li X, Ye J, Wu X, Tan Z,
Liu C, Shen B, Wang XA, Wu W, et al: Whole-exome and targeted gene
sequencing of gallbladder carcinoma identifies recurrent mutations
in the ErbB pathway. Nat Genet. 46:872–876. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao R, Fu X, Teng L, Li Q and Zhao ZJ:
Blocking the function of tyrosine phosphatase SHP-2 by targeting
its Src homology 2 domains. J Biol Chem. 278:42893–42898. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eminaga S and Bennett AM: Noonan
syndrome-associated SHP-2/Ptpn11 mutants enhance SIRPalpha and PZR
tyrosyl phosphorylation and promote adhesion-mediated ERK
activation. J Biol Chem. 283:15328–15338. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen L, Wang ZW, Zhu JW and Zhan X: Roles
of cortactin, an actin polymerization mediator, in cell
endocytosis. Acta Biochim Biophys Sin (Shanghai). 38:95–103. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sung BH, Zhu X, Kaverina I and Weaver AM:
Cortactin controls cell motility and lamellipodial dynamics by
regulating ECM secretion. Curr Biol. 21:1460–1469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eckert MA, Lwin TM, Chang AT, Kim J, Danis
E, Ohno-Machado L and Yang J: Twist1-induced invadopodia formation
promotes tumor metastasis. Cancer Cell. 19:372–386. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao R, Guerrah A, Tang H and Zhao ZJ:
Cell surface glycoprotein PZR is a major mediator of concanavalin
A-induced cell signaling. J Biol Chem. 277:7882–7888. 2002.
View Article : Google Scholar : PubMed/NCBI
|