Introduction
Lung cancer is a common malignant tumor worldwide.
There are 3 main types of lung cancer; ~85% of lung cancer cases
are non-small cell lung cancer (NSCLC) (1). The 5-year survival rate of patients
with some types of cancer has markedly improved during the last two
decades, however, that of patients with lung cancer remains low
(2). It has been reported that
>50% of patients with lung cancer are already at an advanced
stage upon initial diagnosis (3).
In addition, progress in the treatment of lung cancer is slow.
Therefore, new approaches and drugs to prevent and treat lung
cancer are urgently required in order to improve clinical
outcomes.
Peroxisome proliferator-activated receptor γ (PPARγ)
belongs to the nuclear hormone receptor superfamily and
translocates to the nucleus when ligands bind to it (4). PPARγ participates in multiple
physical and pathological processes, including inflammation,
adipocyte differentiation, and lipid and glucose metabolism
(5). A recent study has confirmed
that PPARγ also plays an important role in inhibiting proliferation
and development in lung cancer (6). Therefore, PPARγ agonists have become
a potential therapeutic drug candidate for the treatment of lung
cancer.
Natural traditional Chinese medicine products are
becoming popular in the search for antitumor drugs, both in China
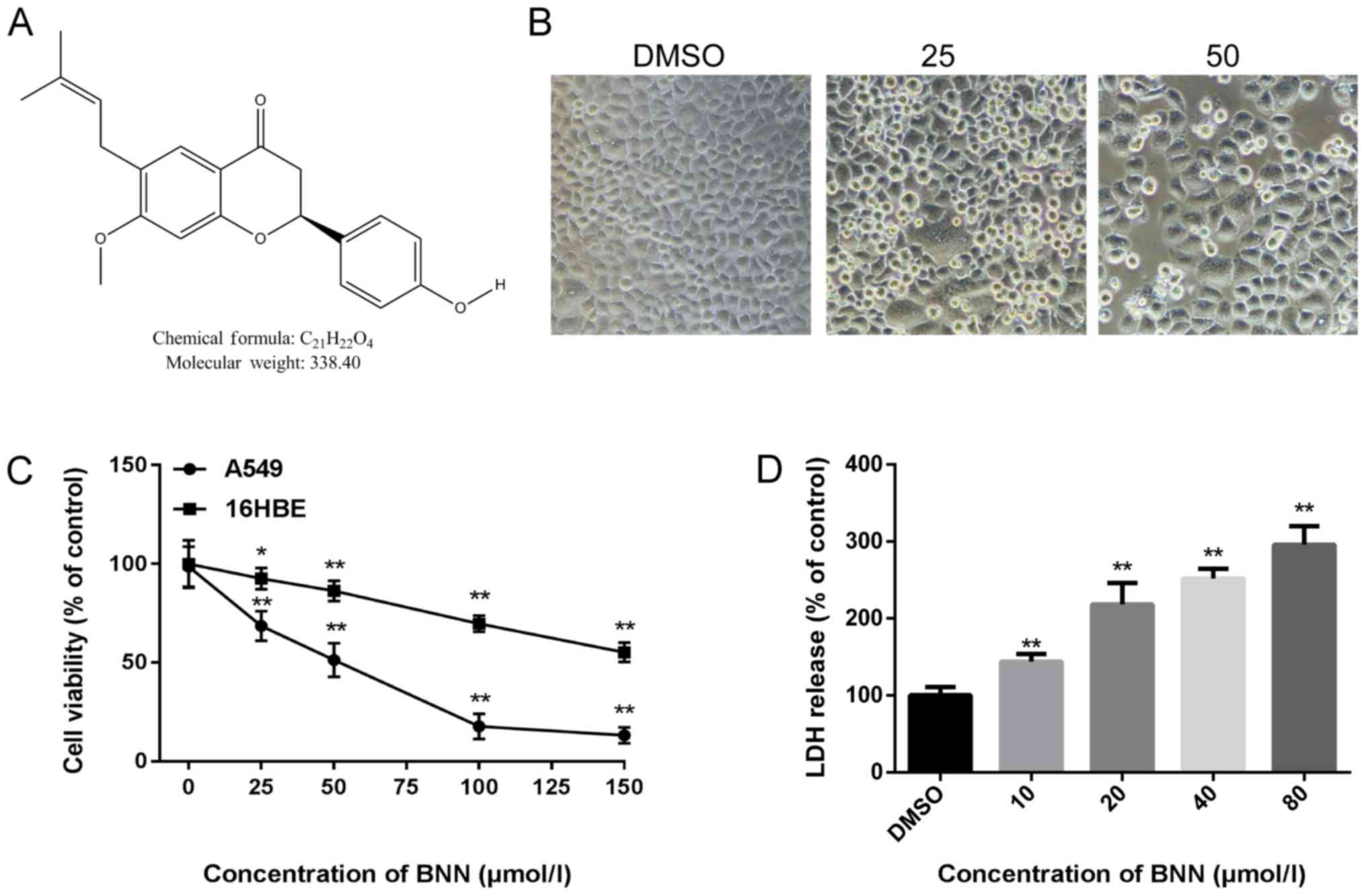
and the rest of the world. Bavachinin (BNN; Fig. 1A) is a naturally occurring compound
of Psoraleacorylifolia, which is widely used for the
treatment of various conditions, including eczema, psoriasis,
diabetes and cancer (7). Several
recent studies have indicated that BNN has PPARγ-activating
properties and is a PPARγ agonist (8,9). The
aim of the present study was to explore the antitumor effect of BNN
in NSCLC A549 cells, as well as the biomechanism involved.
Materials and methods
A549 cell culture
The human NSCLC A549 and human bronchial epithelial
16HBE cell lines were obtained from the Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences and cultured in DMEM
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin in 5% CO2 at 37°C. BNN was
obtained from Chengdu Herbpurify Co., Ltd. DMSO was purchased from
Beijing Solarbio Science & Technology Co., Ltd. The PPARγ
antagonists GW9662 and T0070907 were obtained from Beyotime
Institute of Biotechnology.
Cell Counting Kit-8 (CCK-8) assay
Both A549 and 16HBE cells were seeded on 96-well
culture plates (2×103 cells/well) and the cell culture
medium was removed 24 h later. Medium containing different
concentrations of BNN (0, 25, 50, 100 and 150 µmol/l) was added to
the cells. Following treatment for 24 h at 37°C, 10 µl CCK-8
solution was added to each well and incubated for 1 h at 37°C with
5% CO2. The absorbance value at 450 nm was then measured
using a SpectraMax iD3 microplate reader (Molecular Devices, LLC).
The IC50 was calculated by comparing the cell viability
with the BNN concentration.
Lactate dehydrogenase (LDH) release
assay
A549 cells were seeded on 96-well culture plates
(2×103 cells/well) and treated with different
concentrations of BNN (10, 20, 40 or 80 µM) or in the presence of
PPARγ antagonists GW9662 (20 µM) and T0070907 (10 µM) for 24 h.
Cell culture medium (100 µl) was collected for LDH determination
using an LDH cytotoxicity assay kit according to the manufacturer's
instructions (Beyotime Institute of Biotechnology). Foll owing the
reaction, the absorbance was read at a wavelength of 490 nm using
the SpectraMax iD3 microplate reader.
Measurement of intracellular reactive
oxygen species (ROS)
An ROS assay kit (Beyotime Institute of
Biotechnology) was used to measure intracellular ROS accumulation.
A549 cells were seeded on 6-well culture plates (8×103
cells/well) and treated with different concentrations of BNN (10,
20, 40 µM), with or without PPARγ antagonists GW9662 (20 µM) and
T0070907 (10 µM). The cells were incubated with 10 µM DCFH-DA for
20 min at 37°C, and images were captured using a fluorescence
microscope (Olympus Corporation). Cells were then collected and
measured using a SpectraMax iD3 microplate reader at an excitation
wavelength of 488 nm and an emission wavelength of 525 nm.
Immunofluorescence
Cells seeded in 48-well plates (3×103
cells/well)were fixed with 4% paraformaldehyde at room temperature
for 15 min and permeabilized with 0.3% Triton X-100 (Beyotime
Institute of Biotechnology) at room temperature for 30 min. The
cells were then blocked at 37°C for 1 h with Immunol Staining
Blocking Buffer (Beyotime Institute of Biotechnology), followed by
incubation with an anti-PPARγ antibody (16643-1-AP, 1:200;
ProteinTech Group, Inc.) at room temperature for 1 h. Next, cells
were incubated with an Alexa-488-conjugated secondary antibody
(SA00013-2, 1:500; ProteinTech Group, Inc.) at 37°C for 50 min.
Nuclei were stained with DAPI (Beyotime Institute of
Biotechnology). PPARγ was stained green and the nuclei were stained
blue. Fluorescent imaging was performed using a laser scanning
confocal microscope (Olympus Corporation).
Western blotting
Cells were lysed with RIPA buffer containing 1%
protease inhibitor and centrifuged at 14,000 × g for 20 min at 4°C.
The supernatants were collected and the protein concentrations were
measured by bicinchoninic acid assay kit (Beyotime Institute of
Biotechnology). The extraction of nuclear and cytoplasmic protein
was performed according to the manufacturer's instructions using
nuclear and cytoplasmic protein extraction kit (Beyotime Institute
of Biotechnology). Protein (20 µg) was applied to 10% SDS-PAGE, and
then transferred on to 0.45 µm PVDF membranes. The membranes were
incubated in blocking solution (5% non-fat milk) for 1 h at room
temperature and then incubated with the following primary
antibodies: Bax (50599-2-Ig; 1:2,000; Proteintech Group, Inc.),
Bcl-2 (12789-1-AP; 1:1,000, Proteintech Group, Inc.), PPARγ
(16643-1-AP; 1:2,000; ProteinTech Group, Inc), Caspase-3 (AC030;
1:2,000; Beyotime Institute of Biotechnology), Caspase-9
(10380-1-AP; 1:2,000; ProteinTech Group, Inc), GAPDH (60004-1-Ig;
1:3,000; Proteintech Group, Inc.) and Histone-H3 (17168-1-AP;
1:2,000; ProteinTech Group, Inc) overnight at 4°C. Then the
membranes were incubated with the horseradish peroxidase-conjugated
goat anti-rabbit IgG (A0208; 1:2,000; Beyotime Institute of
Biotechnology) for 1 h at room temperature. Western blotting bands
were visualized by enhanced chemiluminescent reagent (Beyotime
Institute of Biotechnology) and quantified using ImageJ software
(version 1.51j8; National Institutes of Health) and standardized
against GAPDH or Histone-H3 (nuclear protein quantification).
Drug affinity responsive target
stability (DARTS)
DARTS was performed as described in our previous
study (10). Cells were lysed with
M-PER lysis buffer (Thermo Fisher Scientific, Inc.) supplemented
with 1% phosphatase and protease inhibitors and centrifuged at
16,000 × g for 20 min at 4°C. Next, 10X TNC buffer (50 mM Tris·Cl,
50 mM NaCl, 10 mM CaCl2) was added to the supernatant at
room temperature for 10 min. The lysates were incubated with DMSO
or BNN (1, 10 or 100 µM) at room temperature for 1 h, followed by
incubation with 0.03 mg/ml pronase (Roche Diagnostics GmbH) at room
temperature for 30 min. The proteolysis was stopped using SDS
loading buffer. All samples were analyzed by western blotting.
Cellular thermal shift assay
(CETSA)
Cells treated with BNN (10 µM) or DMSO at 37°C for
24 h were collected, and the cell suspension was distributed into
0.2 ml PCR tubes, with 200 µl cell suspension in each tube. The PCR
tubes were heated at the designated temperature (42, 45, 48, 51 and
54°C) for 3 min. They were then removed and incubated at 4°C
immediately following heating. Cells were then lysed using cell
lysis buffer for western (Beyotime Institute of Biotechnology) and
analyzed by western blotting as described in the western blotting
methods above.
Statistical analysis
Statistical analysis of the data was performed using
GraphPad Prism v.6.0 (GraphPad Software, Inc.). Data are presented
as the mean ± standard deviation. Significance was determined by
one-way ANOVA, and Tukey's post hoc test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
BNN inhibits cell viability in human
A549 cells
In order to detect the toxicity of BNN, A549 cell
morphology following BNN exposure was first examined. When human
A549 cells were treated with BNN for 24 h, the cell number
decreased while the cell size increased. When the BNN concentration
was increased to 25 µM, the cell density was decreased. When the
BNN concentration was increased to 50 µM, the cell density was
markedly decreased, and a great number of dead cells were suspended
in the culture medium (Fig. 1B).
Next, a CCK-8 assay was performed to detect the cell viability
using the A549 and 16HBE cell lines following treatment with
different concentration of BNN (0, 25, 50, 100 or 150 µmol/l) for
24 h. As shown in Fig. 1C, the
proliferation of A549 cells was significantly suppressed by BNN in
a dose-dependent manner; BNN exerted less toxicity on 16HBE than on
A549 cells. To further evaluate the toxicity of BNN, the LDH
release from A549 cells treated with BNN was measured in order to
determine whether necrosis was involved in the BNN-induced decrease
in cell viability. LDH release is a typical property of cell
necrosis (11). As shown in
Fig. 1D, BNN dose-dependently
increased the release of LDH, which reflected the levels of
necrotic cell death. In addition, BNN increased the expression of
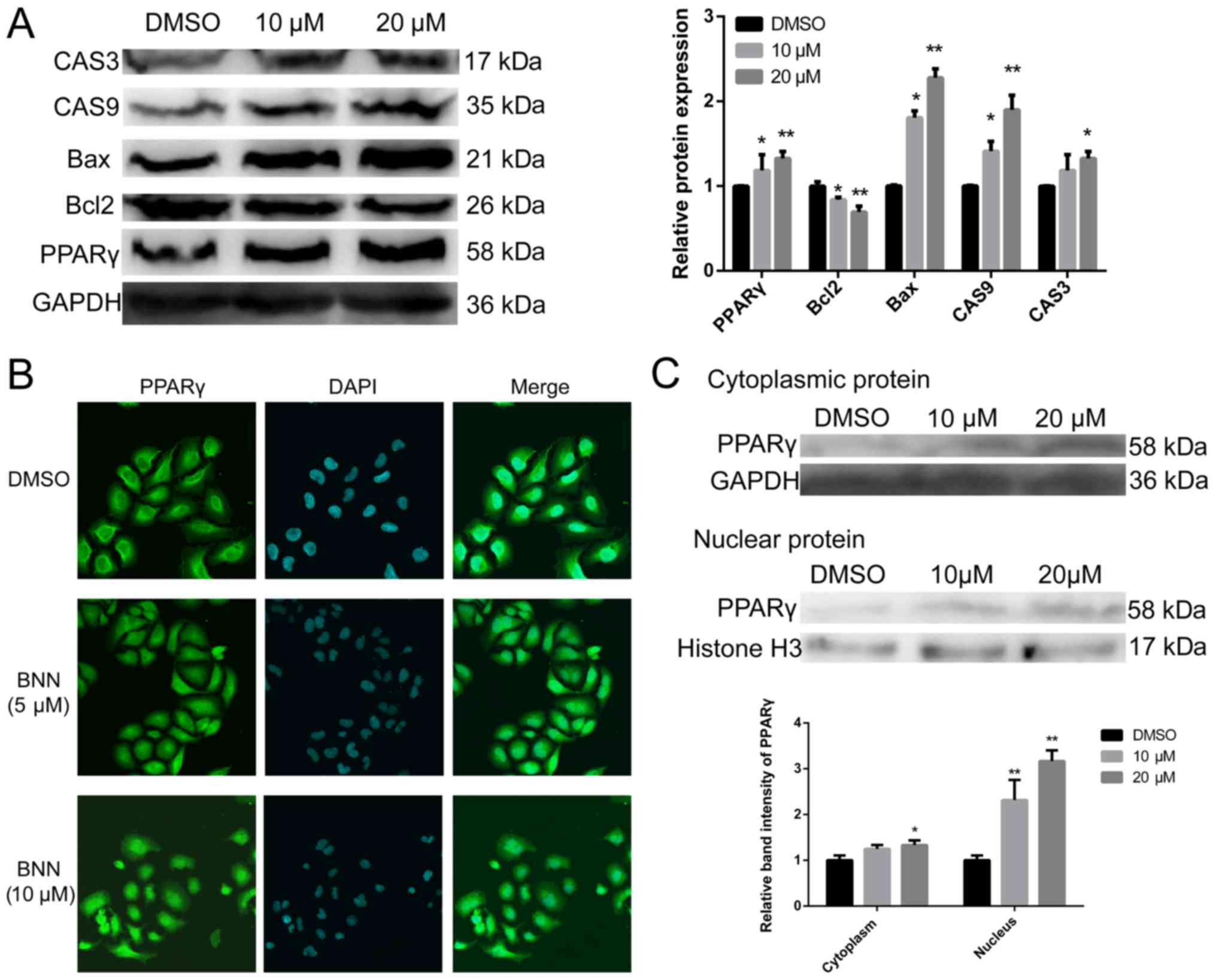
apoptosis-related factors caspase (CAS)-3, CAS9 and Bax, but
decreased the expression of Bcl-2 (Fig. 2A). These proteins act as pro- or
anti-apoptotic regulators in cellular activities (12).
BNN promotes PPARγ protein
expression
During carbohydrate and lipid metabolism, BNN
exhibits pan-PPAR activity (8). We
therefore speculated that BNN may exert a growth inhibition effect
on human A549 cells by activating PPARγ, which may serve as a
promising therapeutic target in lung cancer. As shown in Fig. 2A, BNN significantly (P<0.05)
increased PPARγ protein expression. PPARγ is a ligand-activated
nuclear transcription factor that belongs to the nuclear hormone
receptor superfamily (13). The
results of the immunofluorescence experiments showed that PPARγ
protein levels in both the cytoplasm and the nucleus were markedly
higher following treatment with BNN (Fig. 2B). To further confirm this result,
cytoplasmic and nuclear proteins were separated from cultured A549
cells following treatment with BNN. The results of western blotting
were consistent with those of immunofluorescence (Fig. 2C).
PPARγ directly binds BNN in A549
cells
Since BNN affected PPARγ expression at the protein
level, BNN may directly bind to PPARγ and act as a PPARγ agonist to
exert antitumor effects. To identify whether PPARγ directly binds
to BNN, DARTS and CETSA were employed to validate the affinity
between PPARγ and BNN. DARTS is a label-free strategy to identify
the small molecule targets that are stabilized by binding to small
molecules and can therefore be protected from proteolysis (14,15).
Following incubation with 0.03 mg/ml pronase for 30 min at room
temperature, significant (P<0.01) protection from the
proteolysis of PPARγ was observed in the A549 whole cell lysate in
the presence of 10 and 100 µM BNN (Fig. 3A). The physical interaction of BNN
with PPARγ was further investigated by CETSA, which is based on the
physical phenomenon of small molecule-induced thermal stabilization
of target proteins (16). The A549
cells were incubated with 10 µM BNN for 24 h and the collected
cells were heated. Compared with the DMSO-treated cell lysate, BNN
markedly changed the thermal stability of PPARγ at 42, 45, 48, 51
and 54°C (Fig. 3B).
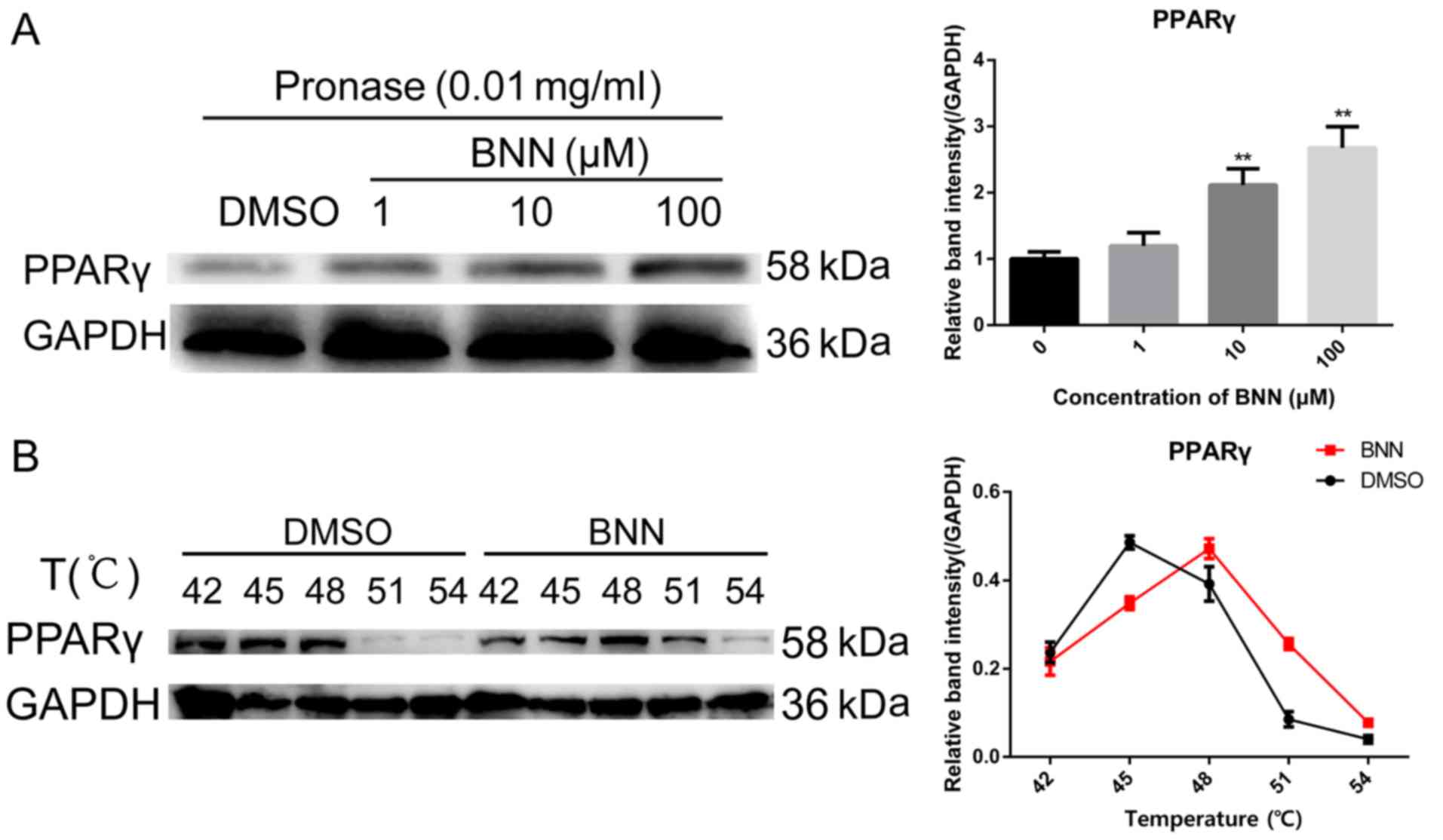 | Figure 3.BNN directly binds to PPARγ. (A) For
the DARTS assay, A549 cell lysates (5 mg/ml) were incubated with
BNN (1, 10 or 100 µM) or an equal volume of DMSO for 1 h at room
temperature, followed by digestion with pronase (0.03 mg/ml) for 30
min. (B) For CETSA experiments, A549 cells were incubated with BNN
(10 µM) or an equal volume of DMSO for 24 h, followed by heating at
the indicated temperatures. Cells were lysed, and the soluble
portion was analyzed by western blotting. The abundance of PPARγ
normalized to GAPDH is presented. **P<0.01 vs. 0 µmol/l BNN.
BNN, bavachinin; PPARγ, peroxisome proliferator-activated receptor
γ; DARTS, drug affinity responsive target stability; CETSA,
cellular thermal shift assay. |
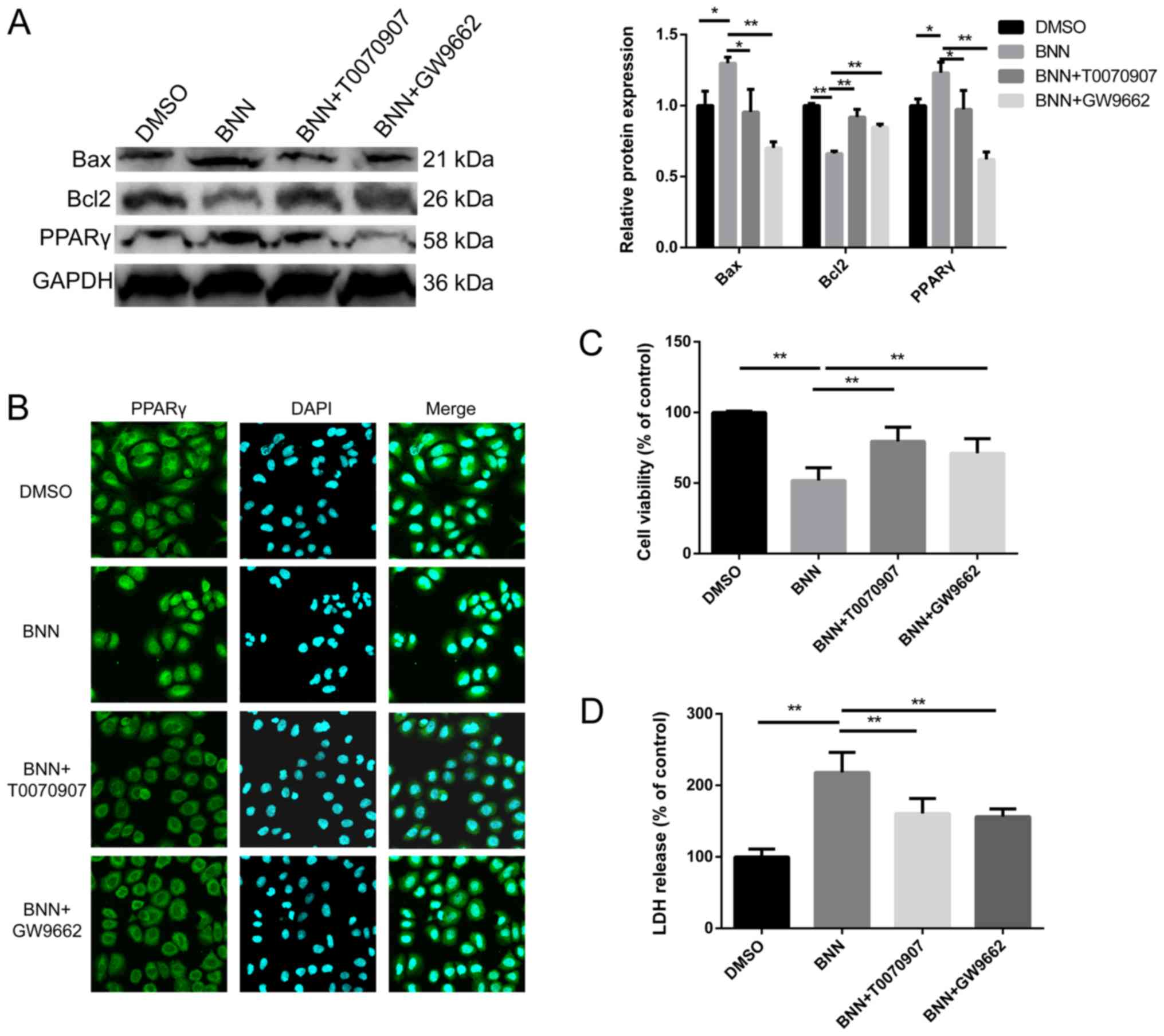
Effects of PPARγ antagonists on cell
toxicity and BNN-induced PPARγ expression
To clarify whether the effect of BNN on A549 was
dependent on PPARγ, cells were co-treated with BNN and PPARγ
antagonists. The BNN-induced alterations in the expression of the
apoptosis-related factors Bcl-2 and Bax were inhibited by the PPARγ
antagonists T0070907 and GW9662 (Fig.
4A). The upregulation of PPARγ and the BNN-induced
translocation of PPARγ to the nucleus were also restrained by
T0070907 and GW9662 (Fig. 4A and
B). Additionally, the present study investigated the impact of
T0070907 and GW9662 alone on A549, which had no significant effect
on A549 cell viability. In order to measure the effect of PPARγ
antagonists on cell toxicity, CCK-8 and LDH release assays were
performed. The cell viability of cells co-treated with PPARγ
antagonists and BNN was higher than that observed in the BNN only
group, while LDH release was decresed, when compared with the BNN
only group (Fig. 4C and D).
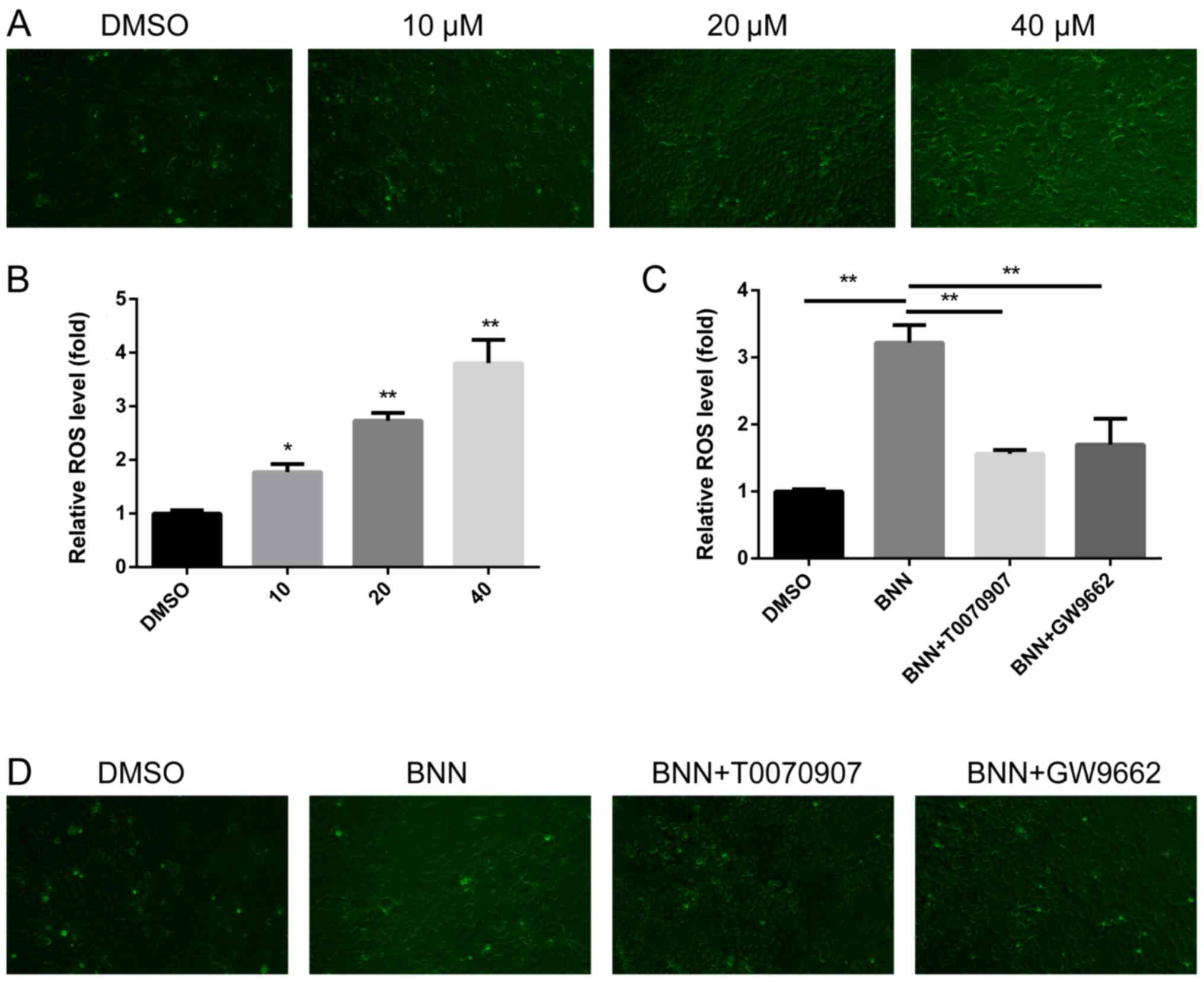
BNN induces ROS generation in a
PPARγ-dependent manner
ROS exerts a complex role in tumor survival and
proliferation; excessive ROS leads to DNA damage, which results in
cell necrosis and apoptosis (17).
The present study therefore investigated whether ROS participated
in the BNN-induced anti-cancer effect. ROS was detected using a
fluorescenct microscope, and the fluorescence intensity was then
measured using a microplate reader. BNN promoted ROS generation in
a dose-dependent manner (Fig. 5A and
B). In order to clarify whether the generation of ROS was
dependent on PPARγ, PPARγ antagonists were applied for
co-treatments with BNN. The upregulation of ROS induced by BNN
could be inhibited by PPARγ antagonists (Fig. 5C and D). These results indicated
that the generation of ROS in A549 cells was dependent on the
expression of PPARγ.
Discussion
Psoralea corylifolia is a popular
multipurpose medicinal plant that has been used to treat nephritis,
osteoporosis, hypertension, cardiovascular diseases and various
types of cancer (18). As one of
the main active ingredients of Psoralea corylifolia, BNN is
a novel natural PPARγ agonist. PPARγ is a nuclear transcription
factor and changes the expression of a series of genes upon its
activation. A previous study has indicated that PPARγ activation
plays a critical role in lung cancer development and progression by
modulating cell differentiation, proliferation, apoptosis and
motility (19). Therefore, PPARγ
may act as a promising therapeutic target in lung cancer, either as
a monotherapy or synergistic therapy.
The present study first confirmed the effect of BNN
on lung cancer A549 cell proliferation. The CCK-8 and LDH release
assays revealed that BNN caused a marked toxic effect on A549
cells. In order to examine cell activity, the protein expression of
apoptosis-related factors, Bax, Bcl-2, CAS3 and CAS9, was also
detected; the results indicated that BNN also promoted the
apoptosis of A549 cells. These results confirmed that BNN can act
as a therapeutic target for lung cancer. To explore the mechanism
of BNN cytotoxicity in lung cancer cells, PPARγ protein expression
was measured. BNN not only upregulated the expression of PPARγ at
the protein level, but also promoted the nuclear translocation of
PPARγ, which was confirmed by the separation of cytoplasmic and
nuclear proteins. These results indicated that BNN could activate
PPARγ, but the affinity between BNN and PPARγ remains unclear. The
DARTS and CETSA results confirmed that BNN could directly bind to
PPARγ.
To further explore the mechanism through which BNN
regulates A549 cell proliferation, the ROS level was also measured.
The results showed that BNN promoted the generation of ROS in a
dose-dependent manner. ROS is generated via the reduction of
molecular oxygen mainly formed in the mitochondrial respiratory
chain (20). In normal cells,
there is a balance between ROS and intracellular biochemical
antioxidants. However, excessive ROS causes oxidative damage to
intracellular biomacromolecules (21). In cancer cells, excessive ROS
production can lead to cell death, including necrotic cell death,
apoptosis, autophagy and ferroptosis (17). A previous study has confirmed that
PPARγ activation has a direct impact on ROS levels (22). The present results also showed that
BNN promoted the production of ROS in a PPARγ-dependent manner.
In conclusion, the present study demonstrated that
BNN induced A549 cell death by promoting PPARγ protein expression
and nuclear translocation. Furthermore, the present results also
suggested that BNN-induced PPARγ activation inhibited A549 cell
proliferation via an ROS dependent-mechanism. The results of the
present study provide a theoretical basis for the clinical
application of BNN in the treatment of NSCLC.
Acknowledgements
Not applicable.
Funding
The present study was funded by The Innovation
Project of Shandong Academy of Medical Sciences.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
KC conceived and designed the experiments, and wrote
the manuscript. LNG performed experiments. LY and CL performed the
statistical analysis and figure editing. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Parums DV: Current status of targeted
therapy in non-small cell lung cancer. Drugs Today (Barc).
50:503–525. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Blandin Knight S, Crosbie PA, Balata H,
Chudziak J, Hussell T and Dive C: Progress and prospects of early
detection in lung cancer. Open Biol. 7(pii): 1700702017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kahremany S, Livne A, Gruzman A,
Senderowitz H and Sasson S: Activation of PPARdelta: From computer
modelling to biological effects. Br J Pharmacol. 172:754–770. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Marion-Letellier R, Savoye G and Ghosh S:
Fatty acids, eicosanoids and PPAR gamma. Eur J Pharmacol.
785:44–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Reddy AT, Lakshmi SP and Reddy RC:
PPARgamma as a novel therapeutic target in lung cancer. PPAR Res.
2016:89725702016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin CH, Funayama S, Peng SF, Kuo CL and
Chung JG: The ethanol extraction of prepared Psoralea corylifolia
induces apoptosis and autophagy and alteres genes expression
assayed by cDNA microarray in human prostate cancer PC-3 cells.
Environ Toxicol. 33:770–788. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Feng L, Luo H, Xu Z, Yang Z, Du G, Zhang
Y, Yu L, Hu K, Zhu W, Tong Q, et al: Bavachinin, as a novel natural
pan-PPAR agonist, exhibits unique synergistic effects with
synthetic PPAR-γ and PPAR-α agonists on carbohydrate and lipid
metabolism in db/db and diet-induced obese mice. Diabetologia.
59:1276–1286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Du G, Zhao Y, Feng L, Yang Z, Shi J, Huang
C, Li B, Guo F, Zhu W and Li Y: Design, Synthesis, and
Structure-activity relationships of bavachinin analogues as
peroxisome proliferator-activated Receptor γ agonists. ChemMedChem.
12:183–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ge L, Cui Y, Cheng K and Han J:
Isopsoralen enhanced osteogenesis by targeting AhR/ERα. Molecules.
23(pii): E26002018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chan FK, Moriwaki K and De Rosa MJ:
Detection of necrosis by release of lactate dehydrogenase activity.
Methods Mol Biol. 979:65–70. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marimuthu P and Singaravelu K: Deciphering
the crucial residues involved in heterodimerization of Bak peptide
and anti-apoptotic proteins for apoptosis. J Biomol Struct Dyn.
36:1637–1648. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo L and Tabrizchi R: Peroxisome
proliferator-activated receptor gamma as a drug target in the
pathogenesis of insulin resistance. Pharmacol Ther. 111:145–173.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pai MY, Lomenick B, Hwang H, Schiestl R,
McBride W, Loo JA and Huang J: Drug affinity responsive target
stability (DARTS) for small-molecule target identification. Methods
Mol Biol. 1263:287–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lomenick B, Jung G, Wohlschlegel JA and
Huang J: Target identification using drug affinity responsive
target stability (DARTS). Curr Protoc Chem Biol. 3:163–180.
2011.PubMed/NCBI
|
|
16
|
Jensen AJ, Martinez Molina D and Lundbäck
T: CETSA: A target engagement assay with potential to transform
drug discovery. Future Med Chem. 7:975–978. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zou Z, Chang H, Li H and Wang S: Induction
of reactive oxygen species: An emerging approach for cancer
therapy. Apoptosis. 22:1321–1335. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Alam F, Khan GN and Asad M: Psoralea
corylifolia L: Ethnobotanical, biological, and chemical aspects: A
review. Phytother Res. 32:597–615. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Han EJ, Im CN, Park SH, Moon EY and Hong
SH: Combined treatment with peroxisome proliferator-activated
receptor (PPAR) gamma ligands and gamma radiation induces apoptosis
by PPARγ-independent up-regulation of reactive oxygen
species-induced deoxyribonucleic acid damage signals in non-small
cell lung cancer cells. Int J Radiat Oncol Biol Phys. 85:e239–e248.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sarniak A, Lipińska J, Tytman K and
Lipińska S: Endogenous mechanisms of reactive oxygen species (ROS)
generation. Postepy Hig Med Dosw (Online). 70:1150–1165. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Juránek I and Bezek S: Controversy of free
radical hypothesis: Reactive oxygen species-cause or consequence of
tissue injury? Gen Physiol Biophys. 24:263–278. 2005.PubMed/NCBI
|
|
22
|
Srivastava N, Kollipara RK, Singh DK,
Sudderth J, Hu Z, Nguyen H, Wang S, Humphries CG, Carstens R,
Huffman KE, et al: Inhibition of cancer cell proliferation by
PPARgamma is mediated by a metabolic switch that increases reactive
oxygen species levels. Cell Metab. 20:650–661. 2014. View Article : Google Scholar : PubMed/NCBI
|