Introduction
Morbidly adherent placenta (MAP) refers to abnormal
attachment of the placenta to the uterus. This includes abnormal
placenta attachment depth, including placenta accreta (PA), but not
placenta previa (PP), which is a condition caused by a low lying
placenta (1). PA refers to a
pathological condition where abnormal placental villi implant into
the uterine wall, due to the partial or total absence of the
decidual basal layer. Placental villi may not only invade the
uterine muscular layer, but may also penetrate the uterine wall
into the outer serosa layer, extending even to the bladder, making
dissection of the placenta from the uterus impossible (2). A previous study reported that the
incidence rate of PA has increased in the past 50 years (3). For patients with PA, dissection of
the entire placenta during delivery is difficult, which may lead to
intrapartum and postpartum hemorrhage. The average hemorrhage size
is large, and it is the main cause of perinatal emergency
hysterectomy (4). The maternal
mortality rate associated with PA is a serious obstetric
complication (5,6). It is necessary to identify key
proteins and pathways affecting placental implantation for further
prediction and treatment.
Zinc finger E-box-binding protein (ZEB) serves an
important role in cell differentiation and embryonic development
(7). It has been reported that ZEB
is associated with biological processes, including tumor invasion
and metastasis (8). Although
embryo implantation into the endometrium is similar to the invasion
mechanism of tumors, limited research has been conducted on ZEB in
pernicious PP (9). Zeb1 is a
nuclear transcription factor of zinc finger proteins. It can bind
to the E2 box on the promoter region of the E-cadherin-coding gene,
thus inhibiting E-cadherin transcription, inducing
epithelial-mesenchymal transition (EMT), and enhancing cell
invasion and metastasis; therefore, it may serve an important role
in cell differentiation and embryonic development, which are
important developmental processes (10,11).
There is also the process of EMT during embryo implantation and
development. Cell invasion of the endometrial layer is an important
factor that determines the success of embryo implantation. Previous
studies have reported that the expression of EMT markers, including
Zeb1, Zeb2, Twist family BHLH transcription factor and Snail family
transcriptional repressor 1 in the outer cell layer of trophoblasts
is increased at 22 days gestation (12). Therefore, it may be speculated that
Zeb1 is involved in regulation of trophoblastic invasion (13). However, to the best of our
knowledge, research into human chorionic villi is limited.
The present study used immunohistochemical methods
to detect the protein expression levels of Zeb1 in chorionic
tissues from 60 patients, including 20 patients with pernicious PP
and PA, 20 patients without PA (UPA) and 20 patients in late
pregnancy that delivered by cesarean section, in order to examine
the association between Zeb1 and PA, and to analyze the effects of
Zeb1 on PA.
Materials and methods
Sample collection
The placental samples were obtained from patients at
the Shengjing Hospital of China Medical University between January
2015 and January 2017 (Table I).
Among the patients, 20 had pernicious PP with PA (PA group). The PA
group consisted of patients with abnormal placental villi invading
the myometrium of the uterus, according to the International
Diagnostic Criteria of PA (14).
Pathological diagnosis was determined by observation of placental
villi having directly invaded the uterine myometrium on the excised
uterine specimens. The average age of the PA group was 27.38±3.98
years. The UPA group comprised 20 patients without PA. The UPA
group consisted of patients in which invasion of abnormal placental
villi into the myometrium of the uterus was not observed, according
to the International Diagnostic Criteria of PA. The average age of
the UPA group was 28.12±2.56 years. Furthermore, 20 placental
samples were obtained from patients in late pregnancy that
delivered by cesarean section (normal group); their average age was
26.99±3.31.
 | Table I.Patient characteristics. |
Table I.
Patient characteristics.
| Characteristic | PA | UPA | Normal |
|---|
| Number | 20 | 20 | 20 |
| Age (mean ±
standard deviation), years | 27.38±3.98 | 28.12±2.56 | 26.99±3.31 |
| BMI,
kg/m2 (mean ± standard deviation) | 22.56±4.24 | 22.01±5.01 | 21.47±3.31 |
| History of cesarean
section | 20 (100%) | 20 (100%) | 5 (25%) |
The selection criteria were as follows: i) Patients
had not entered labor; and ii) had not had a history of cesarean
section. The exclusion criteria for the three groups were as
follows: i) History of hypertension; ii) history of nephropathy and
diabetes; iii) history of anemia or other blood diseases; iv) the
current pregnancy was via vaginal delivery; v) fetal distress or
intrauterine growth retardation; and vi) a history of blood
transfusion and immunotherapy.
Tissue samples were obtained from patients
undergoing surgery at Shengjing Hospital of China Medical
University. Part of the excised tissue was fixed in 10% buffered
formalin (4°C, overnight) and part of the sample was snap-frozen at
−80°C for western blotting. Clinical samples were collected after
written informed consent was obtained from patients, and the study
was approved by the Ethics Committee at the Academic Medical Center
of Shengjing Hospital of China Medical University.
Immunohistochemical evaluation
Tissue sections were immunohistochemically stained
for Zeb1, vascular endothelial growth factor (VEGF) and E-cadherin.
Samples were fixed in 10% buffered formalin (4°C, overnight).
Serial 5 µm sections were cut using a microtome and affixed onto
positively charged slides. Tissues were deparaffinized and
rehydrated through graded xylene and in descending alcohol series.
The paraffin sections were subsequently de-waxed in water and
incubated with 3% H2O2 for 10 min at room
temperature. The sections were incubated with 3% goat serum (cat.
no. 16210064; Thermo Fisher Scientific, Inc.) at room temperature
for 10 min. Immunohistochemistry (IHC) was conducted using rabbit
primary antibodies against Zeb1 (1:800; sc-515797), VEGF (1:800;
sc-53462) and E-cadherin (1:500; sc-71007) (Santa Cruz
Biotechnology, Inc.) at 4°C overnight. An avidin-biotin-horseradish
peroxidase complex immunodetection kit (D110073-0500; Shanghai
Shenggong Biology Engineering Technology Services, Ltd.) was used
to detect the reaction according to the manufacturer's
instructions. Finally, the slides were dried, mounted with Canada
balsam, covered and were examined using a light microscope
(magnifications, ×100 and ×400). Staining intensity was scored on a
scale of 0–4, as follows: 0, no staining; 1, weak staining in 5–25%
of areas; 2, weak staining in 25–50% of areas; 3, moderate staining
in 51–75% of areas and 4, strong staining in >75% of areas.
Cell culture
HTR-8/sv neo cells and human umbilical vein
endothelial cells (HUVECs) were supplied by the China Center for
Type Culture Collection. The cells were cultured in DMEM
(Invitrogen; Thermo Fisher Scientific, Inc.) supplemented with 10%
FBS (Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C in an
atmosphere containing 5% CO2.
Transfection
To overexpress and silence Zeb1, 1×106
HTR-8/sv neo cells or HUVECs were plated into 6-well plates and
transfected with 2 µg pcDNA3.1 vector encoding Zeb1 or with 5 µl
Zeb1-targeted small interfering (si)RNA (si-Zeb1; 10 µmol/l;
5′-CCTCTCTGAAAGAACACATTA-3′; both Shanghai GeneChem Co., Ltd.)
using HiGene transfection reagent (Beyotime Institute of
Biotechnology), according to the manufacturer's instructions. The 2
µg pcDNA3.1 vector and 5 µl non-targeted siRNA (10 µmol/l;
5′-GCAGTTATCTGGAAGATCAGG-3′) were transfected into cells as
respective controls (Shanghai GeneChem Co., Ltd.). Further
experiments were carried out 24 h after transfection.
Cell viability assay
The MTT assay was employed to assess cell viability.
HTR-8/sv neo cells and HUVECs were cultured in 96-well plates at a
concentration of 1×104 cells/ml and were transfected
with a Zeb1 vector or si-Zeb1 for 12, 24, 36 or 48 h. A total of
0.01 ml MTT solution (5 mg/ml) was added to each well. After 4 h of
incubation at 37°C, the medium was replaced with 0.2 ml DMSO for 15
min. Subsequently, optical density was measured at a wavelength of
490 nm.
Cell cycle assay
In total, 1×106 HTR-8/sv neo cells or
HUVECs were seeded into 6-well plates and transfected with a Zeb1
vector or si-Zeb1 for 24 h. Cells were fixed in 75% ethanol at 4°C
overnight. Following staining using 10 µg/ml propidium iodide (PI)
and 100 µl RNase A (100 µg/ml; Beyotime Institute of Biotechnology)
for 20 min at room temperature, cell cycle progression was analyzed
by flow cytometry within 1 h. The cell cycle was then analyzed
using a flow cytometer (BD FACSCalibur; BD Biosciences) and FlowJo
V10 (FlowJo, LLC).
Apoptosis assay
In total, 1×106 HTR-8/sv neo cells or
HUVECs were seeded into 6-well plates and transfected with a Zeb1
vector or si-Zeb1 for 24 h. Cells were washed twice with cold PBS,
and stained with 5 µl Annexin V-fluorescein isothiocyanate
(FITC)/10 µl PI which were included in the Annexin V-FITC/PI
apoptosis kit (Beyotime Institute of Biotechnology). Cells were
incubated for 15 min at room temperature. Subsequently, 400 µl
binding buffer (Beyotime Institute of Biotechnology) was added to
each tube and the apoptosis rate was measured by flow cytometry
within 1 h. Apoptosis was then detected using a flow cytometer and
analyzed using FlowJo.
Transwell migration assay
In total, 1×105 HTR-8/sv neo cells or
HUVECs in 200 µl DMEM were transfected with Zeb1 vector or si-Zeb1
for 24 h in 100 µl FBS-free DMEM. Cells were then placed in the
upper chamber of a Transwell system, whereas the lower compartment
was filled with 600 µl DMEM containing 10% FBS. After 8 h of
incubation at 37°C, cells were fixed in 75% ethanol at 4°C
overnight. The cells that had migrated to the lower surface of the
filter were stained with 4% trypan blue (Beyotime Institute of
Biotechnology) for 20 min at room temperature and counted under a
light microscope (magnification, ×400).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from HTR-8/sv neo cells,
HUVECs or tissues using the RNA Isolation kit (Tiangen Biotech Co.,
Ltd.), according to the manufacturer's protocol, and samples were
stored at −80°C until subsequent use. The RNA Reverse Transcription
kit (Beyotime Institute of Biotechnology) was used for the RT of
RNA into cDNA using the following conditions: 37°C for 15 min, and
followed by 85°C for 5 sec. RT-qPCR was subsequently performed
using a SYBR® Green Real-time PCR Master Mix (Toyobo
Co., Ltd.). RT-qPCR was performed using MX3000P Real-time PCR
instrument (Agilent Technologies, Inc.), according to the
manufacturer's protocol, using the following conditions: 94°C for 5
min, and followed by 30 cycles of 94°C for 30 sec, 58–61°C for 30
sec. Primer sequences for detection of mRNA expression are shown in
Table II. The relative gene
expression levels were calculated using the 2−ΔΔCq
method (15). All experiments were
performed in triplicate.
| Table II.Primers used for reverse
transcription-quantitative PCR. |
Table II.
Primers used for reverse
transcription-quantitative PCR.
| Name | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Zeb1 |
GCACAACCAAGTGCAGAAG |
CATTTGCAGATTGAGGCTG |
| E-cadherin |
GGGGTCTGTCATGGAAGGTG |
CGACGTTAGCCTCGTTCTCA |
| N-cadherin |
CGGCCCGCTATTTGTCATCA |
TGCGATTTCACCAGAAGCCT |
| VEGF |
GCAAAAACGAAAGCGCAAG |
GGAGGCTCCAGGGCATTAGA |
| TRAIL-R2 |
CCACAAAGAATCAGGCATCA |
CCAGGTCGTTGTGAGCTTCT |
| TRAIL-R3 |
GATCGTCCCATCCCCACATC |
CTGCTCTGACCAAGGCTGAA |
| Cyclin D1 |
CCGAGGAGCTGCTGCAAATG |
CGTGCGGGGTCATTGCGGC |
| Bcl-2 |
CTTTGAGTTCGGTGGGGTCA |
GAAATCAAACAGAGGCCGC |
| GAPDH |
GAAGGCTGGGGCTCATTTG |
AGGGGCCATCCACAGTCTTC |
Western blot analysis
Tissues, HTR-8/sv neo cells and HUVECs were lysed in
lysis buffer [25 mM Tris (pH 7.6), 150 mM NaCl, 1% Nonidet P-40, 1
mM EDTA]. The cell lysates were collected by centrifugation at
12,000 × g for 10 min at 4°C. Total protein concentration was
quantified using a bicinchoninic acid protein assay kit (Applygen
Technologies, Inc.), and western blot analysis was performed. A
total of 30 µg protein were separated by 10% SDS-PAGE and were
transferred to a polyvinylidene difluoride membrane (Shanghai
Shenggong Biology Engineering Technology Service, Ltd.). Following
blocking with 3% BSA in TBS containing 0.1% Tween-20 for 3 h at
room temperature, the membranes were incubated overnight at 4°C
with primary antibodies. The following primary antibodies were
used: Zeb1 (1:1,000; cat. no. sc-515797; Santa Cruz Biotechnology,
Inc.), E-cadherin (1:1,000; cat. no. sc-71007; Santa Cruz
Biotechnology, Inc.), N-cadherin (1:1,000; cat. no. sc-393933;
Santa Cruz Biotechnology, Inc.), VEGF (1:1,000; cat. no. sc-53462;
Santa Cruz Biotechnology, Inc.), TRAIL-R2 (1:800; cat. no. ab8416;
Abcam), TRAIL-R3 (1:800; cat. no. ab2087; Abcam), Akt (1:1,000;
cat. no. sc-135829; Santa Cruz Biotechnology, Inc.), phosphorylated
AktSer473 (Aktp-Ser473; 1:1,000; cat. no.
sc-293125; Santa Cruz Biotechnology, Inc.), phosphorylated
AktTyr308 (Aktp-Tyr308; 1:1,000; cat. no.
sc-135650; Santa Cruz Biotechnology, Inc.), cyclin D1 (1:1,000;
cat. no. sc-4074; Santa Cruz Biotechnology, Inc.), Bcl-2 (1:1,000;
cat. no. sc-23960; Santa Cruz Biotechnology, Inc.) and GAPDH
(1:1,000; cat. no. sc-32233; Santa Cruz Biotechnology, Inc.) at 4°C
overnight. After several washes, the membranes were incubated with
an appropriate horseradish peroxidase-conjugated secondary antibody
(goat anti-mouse; 1:5,000; sc-2031; goat anti-rabbit; 1:5,000;
sc-2030; Santa Cruz Biotechnology, Inc.) for 1 h at room
temperature. Proteins bands were visualized using enhanced
chemiluminescence kits (GE Healthcare). After
Akt/E-cadherin/TRAIL-R2 antibodies were detected, the membrane was
stripped for 30 min at 55°C with a wash solution (50 mM Tris, 2%
SDS, 100 mM 2-mercaptoethanol), washed three times with TBS-10%
Tween-20, re-blocked and incubated with
Aktp-Ser473/pAktp-Tyr308/N-cadherin/TRAIL-R3/GAPDH
antibodies. All reactions were repeated at least in triplicate.
Cell treatment
The Akt activator insulin like growth factor 1
(IGF1; 0.1 µg; cat. no. ab9573; Abcam) (16–18)
was added to HTR-8/sv neo cells for 24 h to active Akt. The Akt
kinase inhibitor (cat. no. ab142088; Abcam; 0.5 µg) was added to
HTR-8/sv neo cells for 24 h to inhibit Akt. After 24 h of
treatment, western blot and RT-qPCR were carried out.
Statistical analysis
All reactions were repeated a minimum of three
times. All data are presented as the mean ± standard deviation.
Statistical analysis (two-tailed) of more than two groups was
performed by one-way analysis of variance, followed by Tukey's
post-hoc test for multiple comparisons. A Student's t-test was used
to compare differences between two groups. Statistical analyses
were conducted using GraphPad Prism 6 for Windows (version 6.05;
GraphPad Software, Inc.) P<0.05 was considered to indicate a
statistically significant difference.
Results
Zeb1 expression in placenta
tissues
The expression of Zeb1 in placenta tissues was
assessed by IHC (Fig. 1A). The
results demonstrated that Zeb1 expression was highest in the PA
group, and was higher in the UPA group compared with in the normal
group. In addition, IHC revealed that VEGF expression was higher,
whereas E-cadherin expression was lower in the PA group compared
with in the normal group (Fig. 1B and
C). Western blot analysis and RT-qPCR demonstrated that the
expression levels of Zeb1, N-cadherin, VEGF and TRAIL-R3
(P<0.05) were higher in the PA group compared with in the
control group, whereas E-cadherin and TRAIL-R2 expression were
lower (P<0.05; Fig. 1D and
E)
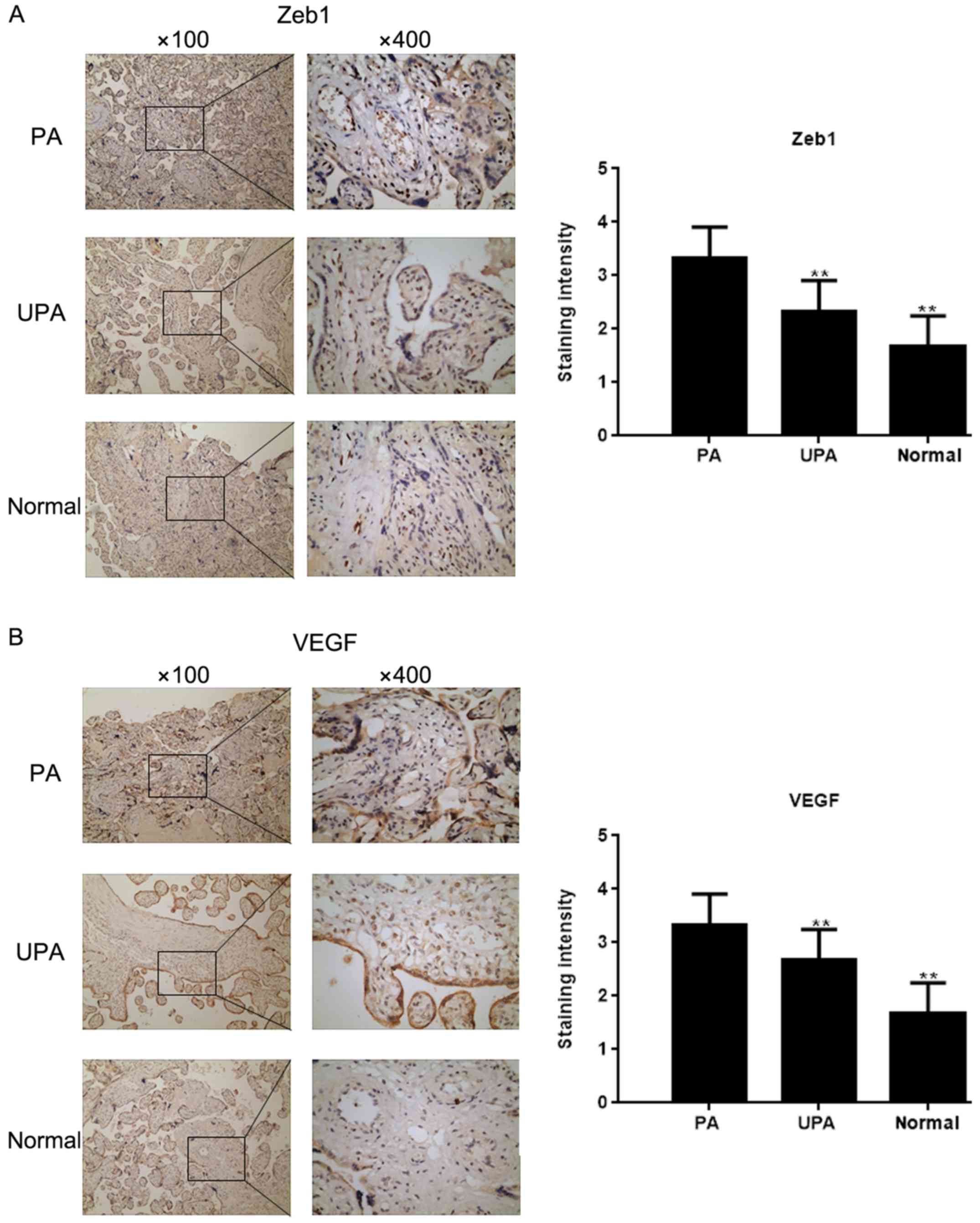 | Figure 1.Zeb1 expression in placenta tissues.
Immunohistochemical staining for the expression of (A) Zeb1 and (B)
VEGF in placenta tissues (magnification, ×100 and ×400).
**P<0.05 vs. PA tissues. E-cad, E-cadherin; N-cad, N-cadherin;
PA, placenta accrete; TRAIL-R, TNF receptor superfamily member;
UPA, placenta previa without PA; VEGF, vascular endothelial growth
factor; Zeb1, zinc finger E-box-binding homeobox 1. Zeb1 expression
in placenta tissues. Immunohistochemical staining for the
expression of (C) E-cad in placenta tissues (magnification, ×100
and ×400). **P<0.05 vs. PA tissues. (D and E) Expression of
Zeb1, E-cad, N-cad, VEGF, TRAIL-R2 and TRAIL-R3 in placenta tissues
was detected by western blot analysis and reverse
transcription-quantitative PCR **P<0.05 vs. PA tissues;
##P<0.05 vs. UPA tissues. E-cad, E-cadherin; N-cad,
N-cadherin; PA, placenta accrete; TRAIL-R, TNF receptor superfamily
member; UPA, placenta previa without PA; VEGF, vascular endothelial
growth factor; Zeb1, zinc finger E-box-binding homeobox 1. |
Si-Zeb1 inhibits the viability and
migration of HTR-8/sv neo cells
The MTT assay revealed that Zeb1-silencing
[depletion was confirmed using western blotting and RT-qPCR
(Fig. 2E and F)] inhibited the
viability of HTR-8/sv neo cells (P<0.05; Fig. 2A). Flow cytometry was used to
evaluate cell cycle progression of HTR-8/sv neo cells previously
exposed to si-Zeb1 for 24 h. si-Zeb1 induced a decrease in the
percentage of cells in S phase, and increased the number of cells
in G1 phase, compared with in the control group
(P<0.05; Fig. 2B). In order to
assess whether Zeb1 can inhibit apoptosis, flow cytometry was used
to assess HTR-8/sv neo cells. The results demonstrated there was an
increase in si-Zeb1-transfected apoptotic cells (P<0.05;
Fig. 2C). The effects of Zeb1 on
HTR-8/sv neo cell migration were assessed using Transwell assays.
The results showed that si-Zeb1 significantly inhibited migration
of HTR-8/sv neo cells (P<0.05; Fig.
2D). Western blot analysis and RT-qPCR revealed that
transfection with si-Zeb1 for 24 h decreased the expression levels
of N-cadherin, VEGF, TRAIL-R3, cyclin D1 and Bcl-2 (P<0.05), and
promoted the expression levels of E-cadherin and TRAIL-R2 in
HTR-8/sv neo cells (P<0.05; Fig. 2E
and F).
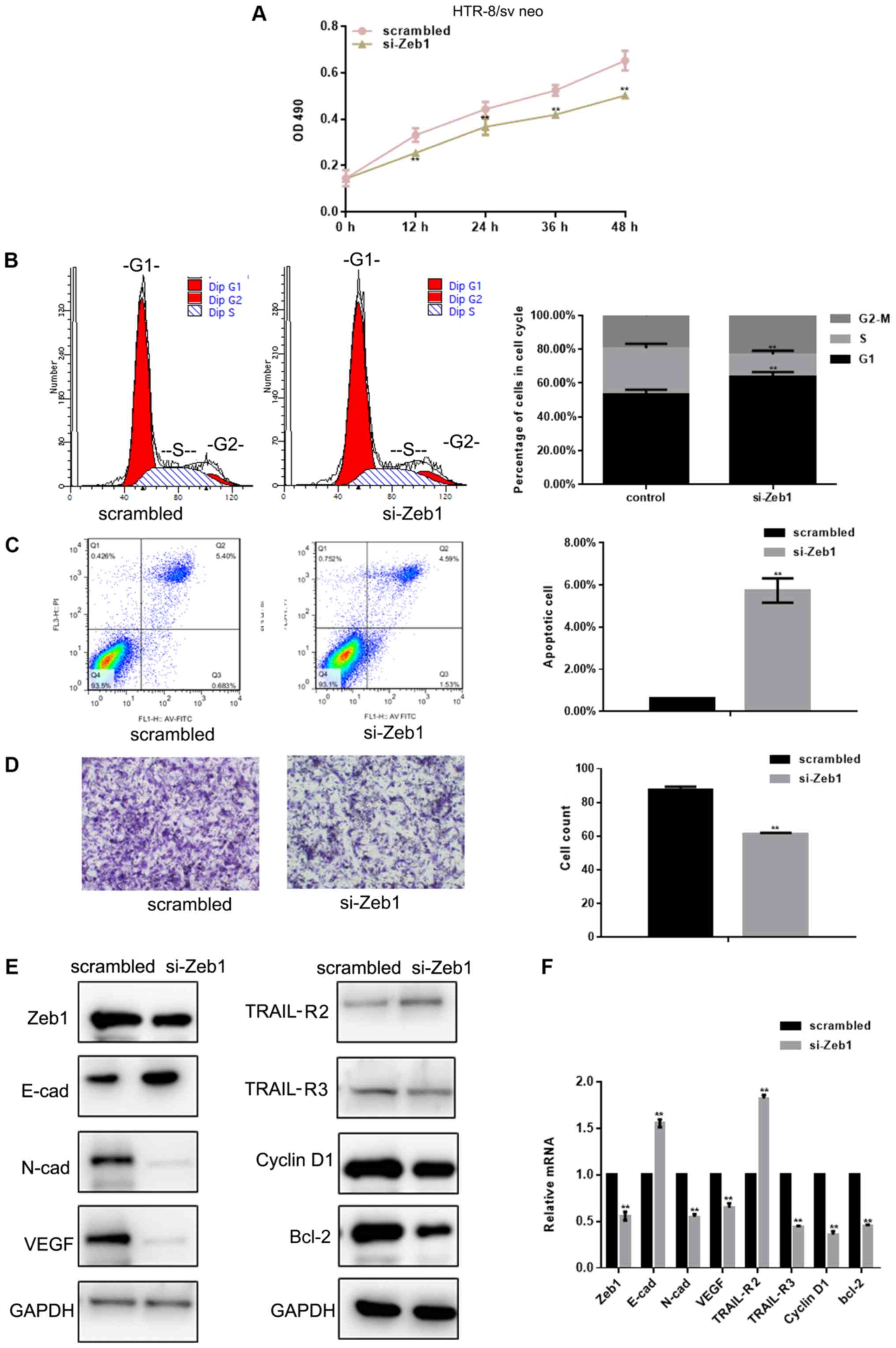 | Figure 2.si-Zeb1 inhibits the growth and
migration of HTR-8/sv neo cells. (A) Viability of HTR-8/sv neo
cells transfected with scrambled siRNA or si-Zeb1 was assessed at
different time points. (B) Flow cytometric analysis of cell cycle
progression of HTR-8/sv neo cells transfected with scrambled siRNA
or si-Zeb1 for 24 h. (C) Flow cytometric analysis of apoptosis of
HTR-8/sv neo cells transfected with scrambled siRNA or si-Zeb1 for
24 h. The Q3 region contains early apoptotic cells; these data were
used for statistical analysis. (D) Migration of HTR-8/sv neo cells
transfected with scrambled siRNA or si-Zeb1 for 24 h
(magnification, ×400). (E and F) Expression levels of Zeb1, E-cad,
N-cad, VEGF, TRAIL-R2, TRAIL-R3, cyclin D1 and Bcl-2, as detected
by western blot analysis and reverse transcription-quantitative
PCR, in HTR-8/sv neo cells transfected with scrambled siRNA or
si-Zeb1 for 24 h. All experiments were repeated in triplicate
**P<0.05 compared with scrambled control. E-cad, E-cadherin;
N-cad, N-cadherin; OD, optical density; si/siRNA, small interfering
RNA; TRAIL-R, TNF receptor superfamily member; VEGF, vascular
endothelial growth factor; Zeb1, zinc finger E-box-binding homeobox
1. |
HUVEC viability and migration are
inhibited by si-Zeb1
The results demonstrated that si-Zeb1 [depletion was
confirmed using western blotting and RT-qPCR (Fig. 3E and F)] inhibited the viability of
HUVECs by inhibiting G1/S transformation and promoting
apoptosis (P<0.05; Fig. 3A-C).
The results of a Transwell assay revealed that si-Zeb1 could
significantly inhibit the migration of HUVECs (Fig. 3D). Western blot analysis and
RT-qPCR showed that si-Zeb1 could inhibit the expression of
N-cadherin, VEGF, TRAIL-R3, cyclin D1 and Bcl-2 (P<0.05), and
promote the expression of E-cadherin and TRAIL-R2 in HUVECs
(P<0.05; Fig. 3E and F).
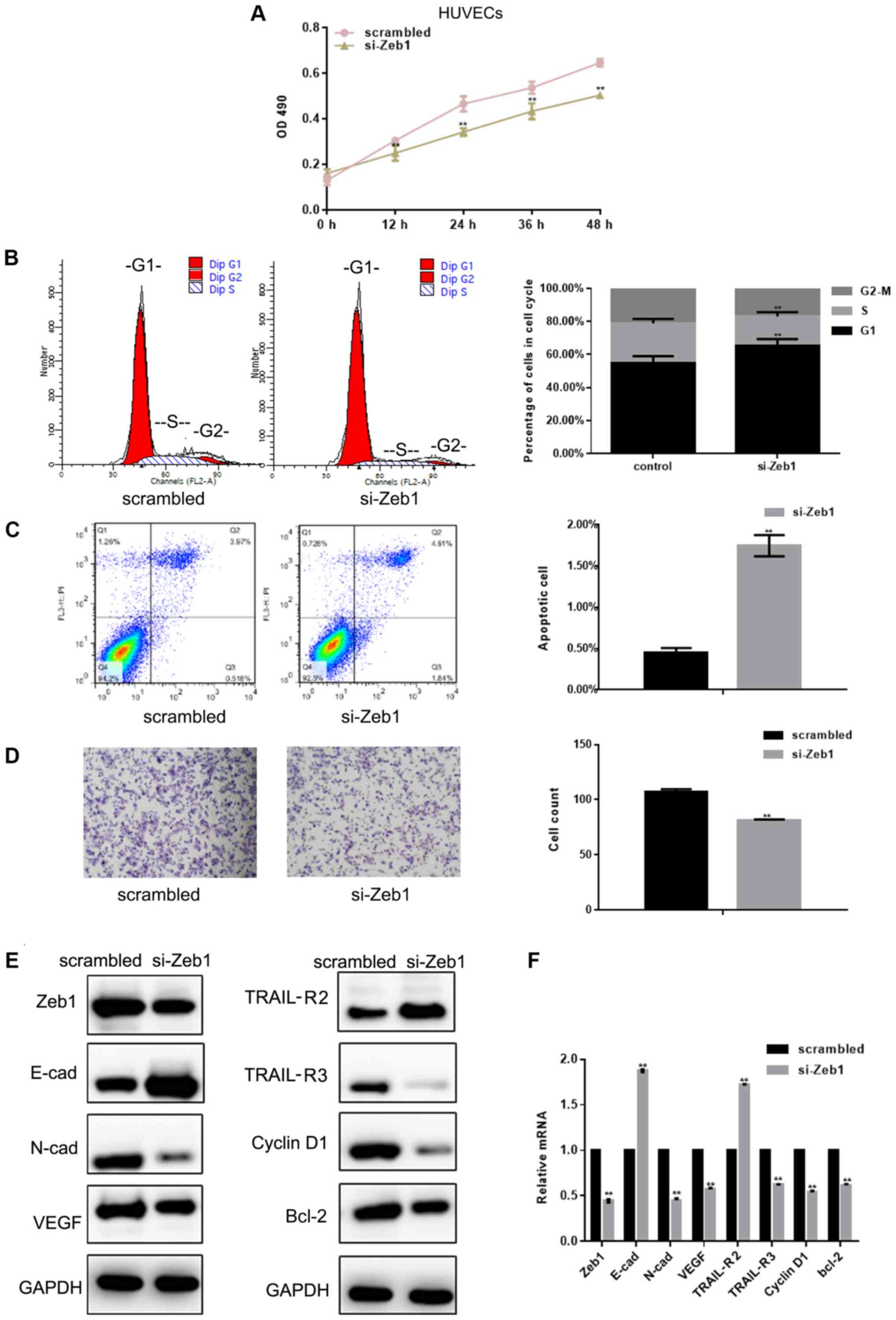 | Figure 3.HUVEC viability and migration are
inhibited by si-Zeb1. (A) Results of MTT viability assays in HUVECs
transfected with scrambled siRNA or si-Zeb1 at different time
points. (B) Flow cytometric analysis of cell cycle progression of
HUVECs transfected with scrambled siRNA or si-Zeb1 for 24 h. (C)
Flow cytometric analysis of apoptosis of HUVECs transfected with
scrambled siRNA or si-Zeb1 for 24 h. The Q3 region contains early
apoptotic cells; these data were used for statistical analysis. (D)
Migration of HUVECs transfected with scrambled siRNA or si-Zeb1 for
24 h (magnification, ×400). (E and F) Expression of Zeb1, E-cad,
N-cad, VEGF, TRAIL-R2, TRAIL-R3, cyclin D1 and Bcl-2, as detected
by western blot analysis and reverse transcription-quantitative
PCR, in HUVECs transfected with scrambled siRNA or si-Zeb1 for 24
h. All experiments were repeated in triplicate **P<0.05 compared
with scrambled control. E-cad, E-cadherin; HUVECs, human umbilical
vein endothelial cells; N-cad, N-cadherin; OD, optical density;
si/siRNA, small interfering RNA; TRAIL-R, TNF receptor superfamily
member; VEGF, vascular endothelial growth factor; Zeb1, zinc finger
E-box-binding homeobox 1. |
Zeb1 promotes cell growth and
migration through the Akt pathway
Since Zeb1 is involved in regulation of the Akt
pathway (19), this study examined
whether the role of Zeb1 in PA was partly mediated by the Akt
pathway. To achieve this, si-Zeb1 cells were treated with an Akt
activator and Zeb1 overexpressing cells were treated with an Akt
inhibitor. Western blot analysis and RT-qPCR revealed that IGF1
reversed the effects of si-Zeb1 on Aktp-Ser473,
Aktp-Tyr308, N-cadherin, VEGF, TRAIL-R3, cyclin D1,
Bcl-2, E-cadherin and TRAIL-R2 in HTR-8/sv neo cells (P<0.05;
Fig. 4A-D). Similar results were
observed in HUVECs, as determined by western blotting and RT-qPCR
(P<0.05; Fig. 5A-D).
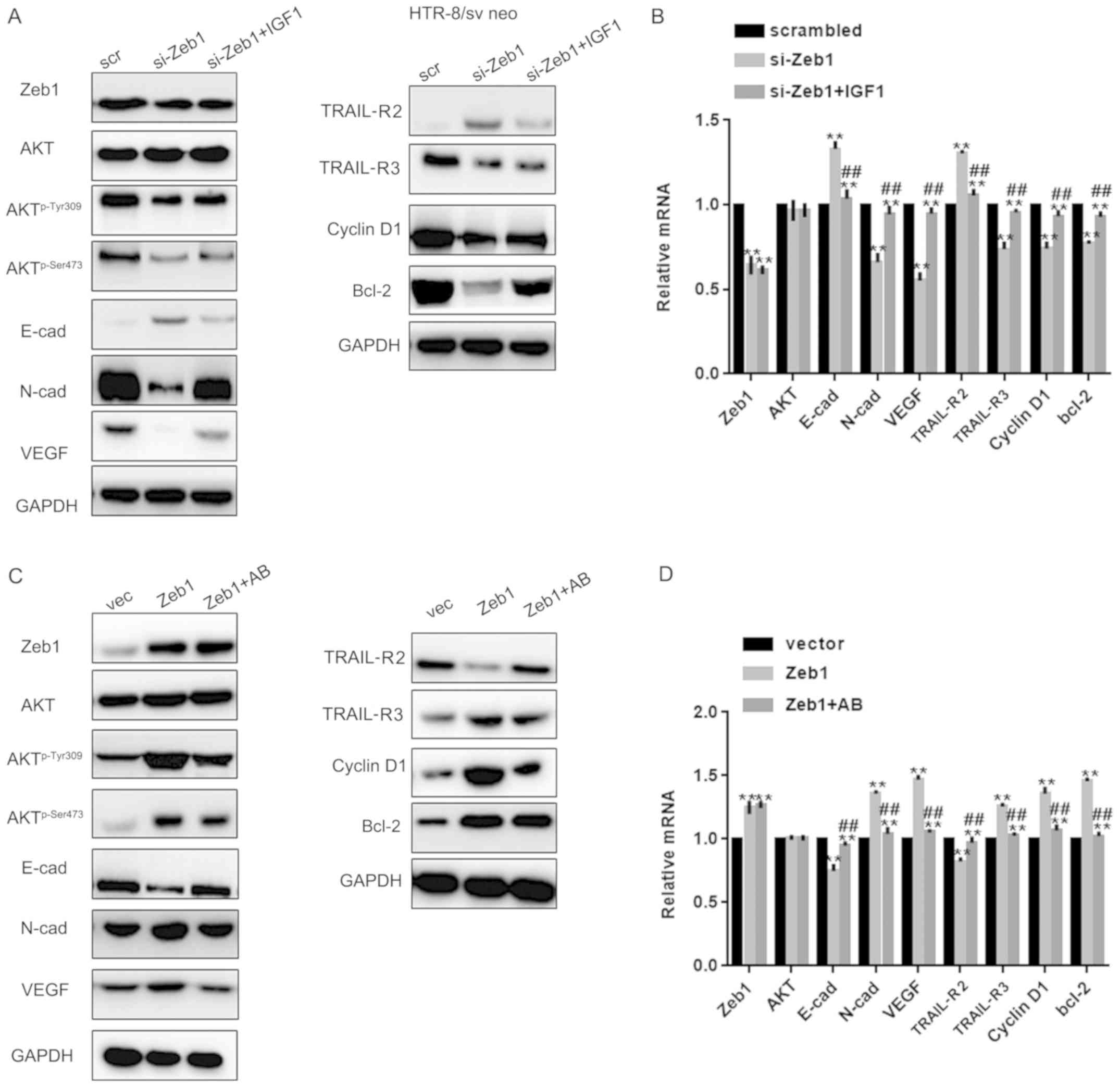 | Figure 4.Zeb1 promotes cell growth and
migration through the Akt pathway. (A and B) Western blotting and
reverse transcription-quantitative PCR analysis of Zeb1, E-cad,
N-cad, VEGF, TRAIL-R2, TRAIL-R3, Akt, Aktp-Ser473,
Aktp-Tyr308, cyclin D1 and Bcl-2 expression in HTR-8/sv
neo cells in the si-Zeb1, si-Zeb1 + IGF1 or scrambled groups.
**P<0.05 compared with scrambled group; ##P<0.05
compared with si-Zeb1 group. (C and D) Western blotting and reverse
transcription-quantitative PCR analysis of Zeb1, E-cad, N-cad,
VEGF, TRAIL-R2, TRAIL-R3, Akt, Aktp-Ser473,
Aktp-Tyr308, cyclin D1 and Bcl-2 expression in HTR-8/sv
neo cells in the Zeb1, Zeb1 + Akt kinase inhibitor and vector
groups **P<0.05 compared with vector group;
##P<0.05 compared with Zeb1 group.
Aktp-Ser473, phosphorylated AktSer473;
Aktp-Tyr308, phosphorylated AktTyr308; E-cad,
E-cadherin; IGF1, insulin like growth factor 1; N-cad, N-cadherin;
si, small interfering; TRAIL-R, TNF receptor superfamily member;
VEGF, vascular endothelial growth factor; Zeb1, zinc finger
E-box-binding homeobox 1. |
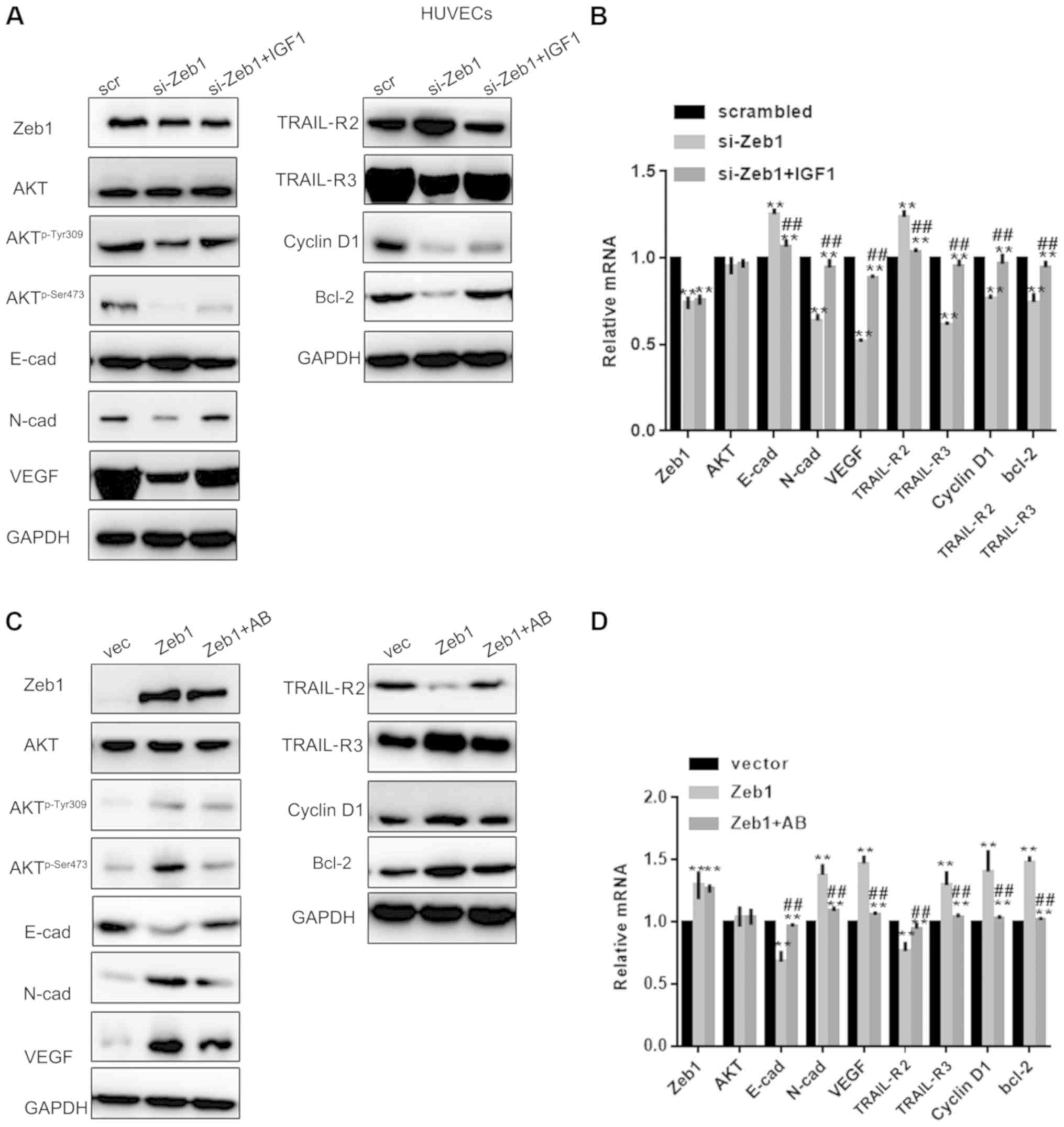 | Figure 5.Zeb1 promotes cell growth and
migration through the Akt pathway. (A and B) Western blotting and
reverse transcription-quantitative PCR analysis of Zeb1, E-cad,
N-cad, VEGF, TRAIL-R2, TRAIL-R3, Akt, Aktp-Ser473,
Aktp-Tyr308, cyclin D1 and Bcl-2 expression in HUVECs in
the si-Zeb1, si-Zeb1 + IGF1 or scrambled groups. **P<0.05
compared with scrambled group; ##P<0.05 compared with
si-Zeb1 group. (C and D) Western blotting and reverse
transcription-quantitative PCR analysis of, E-cad, N-cad, VEGF,
TRAIL-R2, TRAIL-R3, Akt, Aktp-Ser473,
Aktp-Tyr308, cyclin D1 and Bcl-2 expression in HUVECs in
the Zeb1, Zeb1 + Akt kinase inhibitor or vector groups **P<0.05
compared with vector group; ##P<0.05, compared with
Zeb1 group. Aktp-Ser473, phosphorylated
AktSer473; Aktp−Tyr308, phosphorylated
AktTyr308; E-cad, E-cadherin; HUVECs, human umbilical vein
endothelial cells; IGF1, insulin like growth factor 1; N-cad,
N-cadherin; si, small interfering; TRAIL-R, TNF receptor
superfamily member; VEGF, vascular endothelial growth factor; Zeb1,
zinc finger E-box-binding homeobox 1. |
Discussion
PA is a complex process, the underlying mechanism of
which remains unclear. The balance between normal placental villus
infiltration and decidual tissue reaction is the basis for a normal
pregnancy and a smooth delivery (1). It has been suggested that decidual
defects, excessive invasion of trophoblasts and abnormal
neovascularization of the uterus and placenta can lead to PA
(20). When PA occurs, placental
villi adhere tightly to the basal plate of the decidua basalis and
invade the myometrium of the uterus, indicating that the migratory
and invasive ability of trophoblasts outside villi is enhanced
(1,21). In the present study, it was
demonstrated that the expression of Zeb1 was significantly higher
in PA tissues compared with in the control and UPA groups. Zeb1 can
promote the proliferation and migration of trophoblasts and
umbilical vein endothelial cells (22). Therefore, it may be concluded that
Zeb1 serves an important role in PA.
PA is a process of tissue migration, in which EMT
and mesenchymal-epithelial transition (MET) serve important roles,
and the migratory ability of cells is greatly improved (23). Zeb1 may serve an important role in
placental implantation as a regulator of EMT. The process of EMT
involves several changes in protein expression (24). It has previously been reported that
Zeb1 can regulate E-cadherin gene expression to participate in the
regulation of EMT (7). When PA
occurs, the invasive ability of trophoblasts is increased and the
MET process cannot be completed, which is manifested by an increase
in the number and volume of invasive trophoblasts during PA
(9). In the present study, the
expression levels of Zeb1 and E-cad were detected in the three
patient groups: PA, UPA and normal. The results revealed that the
expression levels of Zeb1 were significantly upregulated, whereas
the expression levels of E-cadherin were downregulated in the PA
group. These results are consistent with previous studies (25,26),
which reported that Zeb1 and E-cadherin can participate in the
occurrence and development of placental implantation.
It has been suggested Zeb1 can induce angiogenesis
and promote the expression of VEGF, thereby promoting angiogenesis
(27). VEGF is involved in the
occurrence of preeclampsia. PA is also associated with vascular
abnormalities, and the process of PA can be regulated by Zeb1
(28). In this study, it was
indicated that the expression levels of Zeb1 and VEGF were
significantly upregulated in the PA group, thus indicating that
Zeb1 and VEGF may promote PA.
The Akt signaling pathway serves an important role
in embryogenesis (29).
Overactivation of the Akt signaling pathway has been shown to be
associated with macrosomia and embryonic development (30). In addition, overactivation of the
Akt signaling pathway can induce cell overproliferation and enhance
migratory abilities, which has been widely demonstrated in various
cells (31). EMT is the basic
process of embryonic development, tissue remodeling and wound
healing (30). In recent years,
EMT has gained increasing attention in cancer progression,
invasiveness and metastasis (31).
Through the EMT process, cells may acquire an invasive phenotype,
which may contribute to tissue invasion (32). Zeb1 has been reported to activate a
variety of downstream pathways, including PI3K, Smads and MAPK,
which are involved in Zeb1-induced EMT (33). In this study, the expression of
Zeb1, Akt, Aktp-Ser473, Aktp-Tyr308,
E-cadherin, N-cadherin, VEGF, TRAIL-R2, TRAIL-R3, cyclin D1 and
Bcl-2 were detected following the addition of the Akt activator
IGF1 to Zeb1-silenced cells. The results indicated that Zeb1
silencing could decrease the phosphorylation levels of Akt without
affecting the total protein levels of Akt, whereas IGF1 could
counteract the change in protein expression induced by Zeb1
silencing. In addition, treatment with an Akt kinase inhibitor
weakened the effects of Zeb1 overexpression on protein production
to some extent. These findings suggested that the regulatory effect
of Zeb1 on cellular biological function may be achieved through the
Akt signaling pathway to some extent.
In the present study, the sample size was limited,
and other possible signaling pathways that Zeb1 may regulate were
not screened; therefore, further examination is required. In
addition, there are further limitations to this study. Notably,
SC79, which is more specific than IGF-1, was not used as an Akt
activator. Therefore, in future studies, a more specific activator
of Akt should be used. Furthermore, IGF-1 can produce off-target
effects that do not involve Akt. Further analysis should be
conducted in a follow-up study.
In conclusion, this study demonstrated that Zeb1 may
promote placental implantation by activating the Akt signaling
pathway. The present findings support the potential of Zeb1 as a
novel diagnostic and treatment target for PA.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Key Research
and Development Program of Reproductive Health & Major Birth
Defects Control and Prevention (grant no. 2016YFC1000404 to Chong
Qiao) and he Science and Technology Project of Liaoning Provincial
Education Department (grant no. LS201611 for Na Li).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NL, TY, HL and WY performed the majority of the
experiments, and contributed to the writing of the manuscript. CQ
and CL designed the study and wrote the manuscript.
Ethics approval and consent to
participate
For the use of clinical materials for research
purposes, written consent and approval from patients were obtained
from the Shengjing Hospital of China Medical University. Patient
consent was obtained in writing according to institutional
regulations. Research involving human subjects, human material, or
human data was performed in accordance with the Declaration of
Helsinki and this study was approved by the Ethics Committee at the
Academic Medical Center of Shengjing Hospital of China Medical
University.
Patient consent to publication
Consent to publish was obtained from the
participants.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Roziana R, Kamarul Azhar K, Lau JH, Aina
MAA, Nadia R, Siti Nordiana A and Mohd Zulkifli K: Morbidly
adherent placenta: One-year case series in a tertiary hospital. Med
J Malaysia. 74:128–132. 2019.PubMed/NCBI
|
|
2
|
Balachandar K and Inglis E: The management
of severe pre-eclampsia and HELLP syndrome in a twin pregnancy with
a known morbidly adherent placenta: A case report. Case Rep Womens
Health. 22:e001142019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bamber JH and Sobers S: The Need to
Consider the Women's Perspective: Neuraxial anesthesia and cesarean
delivery for morbidly adherent placenta. Anesth Analg. 128:e56–e57.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lopes ES, Feitosa FEL, Brazil AV, de
Castro JDV, da Costa JIF, Araujo Júnior E, Peixoto AB and Carvalho
FHC: Assessment of sensitivity and specificity of ultrasound and
magnetic resonance imaging in the diagnosis of placenta accreta.
Rev Bras Ginecol Obstet. 41:17–23. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Furukawa S, Fujisaki M, Maki Y, Oohashi M,
Doi K and Sameshima H: Manual removal of placenta in women having
unpredictable adherent placenta. J Obstet Gynaecol Res. 45:141–147.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Katzman PJ, Blitman J and Metlay LA: Basal
chronic villitis and disorders of the placental basal plate: A
possible immunological link between hypertensive disorders of
pregnancy and morbidly adherent placenta. Pediatr Dev Pathol. Jan
21–2019.(Epub ahead of print). doi: 10.1177/1093526619825708.
View Article : Google Scholar
|
|
7
|
Wang Y, Wu Z and Hu L: The regulatory
effects of metformin on the [SNAIL/miR-34]:[ZEB/miR-200] system in
the epithelial-mesenchymal transition(EMT) for colorectal cancer
(CRC). Eur J Pharmacol. 834:45–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Egger JV, Lane MV, Antonucci LA, Dedi B
and Krucher NA: Dephosphorylation of the retinoblastoma protein
(Rb) inhibits cancer cell EMT via Zeb. Cancer Biol Ther.
17:1197–1205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mooney SM, Talebian V, Jolly MK, Jia D,
Gromala M, Levine H and McConkey BJ: The GRHL2/ZEB Feedback Loop-A
Key Axis in the regulation of EMT in breast cancer. J Cell Biochem.
118:2559–2570. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Noman MZ, Janji B, Abdou A, Hasmim M,
Terry S, Tan TZ, Mami-Chouaib F, Thiery JP and Chouaib S: The
immune checkpoint ligand PD-L1 is upregulated in EMT-activated
human breast cancer cells by a mechanism involving ZEB-1 and
miR-200. Oncoimmunology. 6:e12634122017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ohashi S, Natsuizaka M, Naganuma S, Kagawa
S, Kimura S, Itoh H, Kalman RA, Nakagawa M, Darling DS, Basu D, et
al: A NOTCH3-mediated squamous cell differentiation program limits
expansion of EMT-competent cells that express the ZEB transcription
factors. Cancer Res. 71:6836–6847. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rhodes LV, Tate CR, Segar HC, Burks HE,
Phamduy TB, Hoang V, Elliott S, Gilliam D, Pounder FN, Anbalagan M,
et al: Suppression of triple-negative breast cancer metastasis by
pan-DAC inhibitor panobinostat via inhibition of ZEB family of EMT
master regulators. Breast Cancer Res Treat. 145:593–604. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shu H, Chen H, Yang B, Chang Z, Xiong M
and Chen W: Aberrant expression of E-cadherin and integrin β-1 in
trophoblasts is associated with malignant gestational trophoblastic
diseases. Int J Gynecol Cancer. 23:749–754. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Al-Khan A, Bulmer JN, Chantraine F, Chen
CP, Chen Q, Collins S, Cotechini T, Fitzgerald JS, He M, Holland O,
et al: IFPA Meeting 2012 Workshop Report III: Trophoblast
deportation, gestational trophoblastic disease, placental
insufficiency and fetal growth restriction, trophoblast
over-invasion and accreta-related pathologies, placental thrombosis
and fibrinolysis. Placenta. 34 (Suppl):S11–S16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang Y, Jia L, Wang B, Diao S, Jia R and
Shang J: MiR-495/IGF-1/AKT Signaling as a novel axis is involved in
the Epithelial-to-Mesenchymal transition of oral squamous cell
carcinoma. J Oral Maxillofac Surg. 77:1009–1021. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu T, Lu MN, Chen B, Tong J, Mao R, Li SS,
Dai P, Tan YX and Xiyang YB: Electro-acupuncture-induced
neuroprotection is associated with activation of the IGF-1/PI3K/Akt
pathway following adjacent dorsal root ganglionectomies in rats.
Int J Mol Med. 43:807–820. 2019.PubMed/NCBI
|
|
18
|
Kuang WH, Dong ZQ, Tian LT and Li J: IGF-1
defends against chronic-stress induced depression in rat models of
chronic unpredictable mild stress through the PI3K/Akt/FoxO3a
pathway. Kaohsiung J Med Sci. 34:370–376. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Q, Zhou L, Guan Y, Cheng Y and Han
X: BENC-511, a novel PI3K inhibitor, suppresses metastasis of
non-small cell lung cancer cells by modulating β-catenin/ZEB1
regulatory loop. Chem Biol Interact. 294:18–27. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsubara S: Balloon uterine tamponade
device after peripartum hysterectomy for morbidly adherent
placenta. Obstet Gynecol. 133:1882019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shmakov RG, Vinitskiy AA, Chuprinin VD,
Yarotskaya EL and Sukhikh GT: Alternative approaches to surgical
hemostasis in patients with morbidly adherent placenta undergoing
fertility-sparing surgery. J Matern Fetal Neonatal Med.
32:2042–2048. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shin JO, Nakagawa E, Kim EJ, Cho KW, Lee
JM, Cho SW and Jung HS: miR-200b regulates cell migration via Zeb
family during mouse palate development. Histochem Cell Biol.
137:459–470. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kokkinos MI, Murthi P, Wafai R, Thompson
EW and Newgreen DF: Cadherins in the human
placenta-epithelial-mesenchymal transition (EMT) and placental
development. Placenta. 31:747–755. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bottalico B, Larsson I, Brodszki J,
Hernandez-Andrade E, Casslén B, Marsál K and Hansson SR:
Norepinephrine transporter (NET), serotonin transporter (SERT),
vesicular monoamine transporter (VMAT2) and organic cation
transporters (OCT1, 2 and EMT) in human placenta from pre-eclamptic
and normotensive pregnancies. Placenta. 25:518–529. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Duzyj CM, Buhimschi IA, Motawea H, Laky
CA, Cozzini G, Zhao G, Funai EF and Buhimschi CS: The invasive
phenotype of placenta accreta extravillous trophoblasts associates
with loss of E-cadherin. Placenta. 36:645–651. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Incebiyik A, Kocarslan S, Camuzcuoglu A,
Hilali NG, Incebiyik H and Camuzcuoglu H: Trophoblastic E-cadherin
and TGF-beta expression in placenta percreta and normal
pregnancies. J Matern Fetal Neonatal Med. 29:126–129. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guimaraes GC, Alves LA, Betarelli RP,
Guimarães CSO, Helmo FR, Pereira Júnior CD, Corrêa RRM and
Zangeronimo MG: Expression of vascular endothelial growth factor
(VEGF) and factor VIII in the gilt placenta and its relation to
fetal development. Theriogenology. 92:63–68. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Klettner A, Kaya L, Flach J, Lassen J,
Treumer F and Roider J: Basal and apical regulation of VEGF-A and
placenta growth factor in the RPE/choroid and primary RPE. Mol Vis.
21:736–748. 2015.PubMed/NCBI
|
|
29
|
Mathew S, Sundararaj S, Mamiya H and
Banerjee I: Regulatory interactions maintaining self-renewal of
human embryonic stem cells as revealed through a systems analysis
of PI3K/AKT pathway. Bioinformatics. 30:2334–2342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lv WM, Zhao Y, Yang G, Dong SY, Zhang GH,
Zhang Y, Li GZ and Cheng Y: Role of Ras, ERK, and Akt in
glucocorticoid-induced differentiation of embryonic rat
somatotropes in vitro. Mol Cell Biochem. 391:67–75. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Godoy-Parejo C, Deng C, Liu W and Chen G:
Insulin stimulates PI3K/AKT and cell adhesion to promote the
survival of individualized human embryonic stem cells. Stem Cells.
Apr 25–2019.(Epub ahead of print). doi: 10.1002/stem.3026.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Park JH and Han HJ: Caveolin-1 plays
important role in EGF-induced migration and proliferation of mouse
embryonic stem cells: Involvement of PI3K/Akt and ERK. Am J Physiol
Cell Physiol. 297:C935–C944. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tan Y, Xin X, Coffey FJ, Wiest DL, Dong LQ
and Testa JR: Appl1 and Appl2 are expendable for mouse development
but are essential for HGF-Induced akt activation and migration in
mouse embryonic fibroblasts. J Cell Physiol. 231:1142–1150. 2016.
View Article : Google Scholar : PubMed/NCBI
|














