Introduction
The coupling of osteogenesis and osteoclastogenesis
is mediated through intercellular signaling during bone remodeling
(1). Receptor activator of nuclear
factor-κB ligand (RANKL) is a central player in osteoblast-induced
osteoclast differentiation (2).
RANKL is expressed in osteoblasts and osteocytes and binds the
receptor activator of nuclear factor-κB on the surface of
osteoclast precursors to induce osteoclast differentiation through
the NF-κB pathway (2).
Osteoprotegerin (OPG) functions as a soluble decoy receptor of
RANKL, and thereby inhibits osteoclastogenesis (2).
Runt-related transcription factor 2 (RUNX2) is a
master transcription factor in osteogenesis (3). The targeted disruption of RUNX2
results in a complete loss of bone formation owing to the arrest of
osteoblast maturation (4).
Transgenic mice overexpressing RUNX2 in osteoblasts show increased
osteoclast differentiation and dramatically enhanced bone
resorption (5). The dual role of
RUNX2 in osteogenesis and osteoclastogenesis indicates its
involvement in integrating signals between osteoblasts and
osteoclasts (6). RUNX2
transactivates many essential genes in osteogenesis (7,8), yet
its role in osteoclastogenesis remains unclear. There are putative
RUNX2 binding sites in the RANKL promoter sequence (9), however, RUNX2 does not significantly
affect RANKL expression in osteoblasts (10). It has also been reported that RUNX2
may regulate osteoclastogenesis through RANKL trafficking (6). This observation suggests that RUNX2
may target genes required for the regulation of the intracellular
trafficking of RANKL.
A previous genomic analysis reported that the forced
expression of RUNX2 upregulated several genes related to
intracellular trafficking (11);
one of these genes encoded for lysosomal-associated protein
transmembrane 5 (LAPTM5). LAPTM5 is a transmembrane protein that
resides in lysosomes and functions as a regulator of protein
trafficking (12). The transport
of LAPTM5-positive vesicles from the Golgi to the lysosome is
modulated by its binding to the E3 ubiquitin ligase NEDD4, a
HECT-type E3 ligase that belongs to the Nedd4 family (13). LAPTM5 is involved in the negative
regulation of antigen receptor expression as a mediator of
lysosomal degradation (14–16).
The role of LAPTM5 in lysosomal function and its possible
regulation by RUNX2 led to the hypothesis that LAPTM5 may be a
modulator of RANKL trafficking in osteoblasts, the assessment of
which could further the understanding of the coupling of
osteogenesis and osteoclastogenesis. In the present study, this
hypothesis was initially tested by analyzing RUNX2 binding sites in
the LAPTM5 promoter to investigate their role in transcription
regulation. In addition, the role of LAPTM5 in the regulation of
RANKL expression was assessed.
Materials and methods
Cell culture
The bone marrow-derived mesenchymal pluripotent cell
line ST2 was a gift from Dr Xin Ye of Shandong University. The
murine monocyte/macrophage cell line RAW264.7 was purchased from
The Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences. Primary osteoblasts (POBs) were isolated from the
calvaria of female newborn C57BL/6 mice, as previously described
(17). A total of 12 mice were
purchased from Guangdong Medical Laboratory Animal Center. Cell
isolation was performed upon arrival of the mice. The procedure was
approved by The Institutional Animal Care and Use Committee of
Guangzhou Medical University. The cells were cultured in α-minimal
essential medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.).
The cells were co-cultured using Transwell chambers (Corning,
Inc.). For co-culture, RAW264.7 cells were cultured in 24-well
plates and ST2 cells were cultured in Transwell chambers and were
supplemented with 10 nM 1,25-(OH)2D3
(Sigma-Aldrich; Merck KGaA) and 100 nM dexamethasone
(Sigma-Aldrich; Merck KGaA). To explore the effect of co-culture on
osteoclastogenesis, RAW264.7 cells cultured with transwell
containing no ST2 cells served as the control.
Reverse transcription-quantitative PCR
(RT-qPCR)
The cells in culture plates were washed three times
with PBS. TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) was used to isolate the RNA. The concentration and purity of
the total RNA were measured using a UV spectrophotometer.
First-strand complementary DNA synthesis was performed using M-MLV
Reverse Transcriptase kit (Promega Corporation) according to the
manufacturer's instruction. qPCR was performed using a Real-Time
Quantitative PCR machine (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with ChamQ™ Universal SYBR® qPCR
Master Mix (Vazyme Biotech Co., Ltd.) based on SYBR-Green method.
The thermal condition was 40 cycles of denaturation at 95°C for 15
sec and annealing/extension at 60°C for 30 sec. The primers used
are listed in Table I. β-Actin was
used as the control gene. The 2−ΔΔCq method was used to
calculate the relative expression level of each mRNA (18).
 | Table I.Primers used in reverse
transcription-quantitative PCR. |
Table I.
Primers used in reverse
transcription-quantitative PCR.
| Target | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| Oscar |
CCTCCAGACTCCACCAGATA |
GGAAATAAGGCACAGGAAGG |
| TRAP |
GCCCCAAAGAAATGACCATC |
CTGTAAGTAAGCCCCTTGGT |
| Nfatc1 |
GTCTCTTTCCCCGACATCAT |
TCTCCAAGTAACCGTGTAGC |
| RUNX2 |
GACTGTGGTTACCGTCATGGC |
ACTTGGTTTTTCATAACAGCGGA |
| RANKL |
AGGCTGGGCCAAGATCTCTA |
GTCTGTAGGTACGCTTCCCG |
| LAPTM5 |
CGTACCTCAGGATGGCTGAC |
CAAGCTTCAAGTACGCTGGC |
| β-actin |
AGACCTCTATGCCAACACAG |
ACTCATCGTACTCCTGCTTG |
Western blotting
Cells were washed three times with PBS and lysed in
50 mM Tris-HCl buffer (pH 7.0) containing 150 mM NaCl, 1 mM EDTA,
1% Triton X-100 and a protease inhibitor cocktail. Total protein
concentrations were determined using the bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology). Protein samples
(20 µg) were separated by 8% SDS-PAGE, transferred to PVDF
membranes and soaked in a blocking solution (5% nonfat dry milk and
0.1% Tween-20) for 30 min at room temperature. The following
primary antibodies were used: RUNX2 (clone D1H7; 1:1,000; cat. no.
8486; Cell Signaling Technology, Inc.), LAPTM5 (clone H-178; 1:500;
cat. no. sc-134676; Santa Cruz Biotechnology, Inc.) and GAPDH
(1:5,000; cat no. HC301; Beijing Transgen Biotech Co., Ltd.). The
secondary antibodies used were Peroxidase-AffiniPure goat
anti-rabbit or anti-mouse immunoglobulin G (1:10,000; cat. nos.
111-035-003 and 115-035-003; Jackson ImmunoResearch Laboratories,
Inc.). Protein bands were visualized using an ECL kit (Forevergen).
GAPDH was used as an endogenous control.
ELISA
The concentration of RANKL in the medium was
determined using the Mouse TNFSF11/RANKL PicoKine™ ELISA kit (cat.
no. EK0843; Wuhan Boster Biological Technology, Ltd.) in accordance
with the manufacturer's instructions.
Fluorescence microscopy
Immunofluorescence staining was performed as
previously described (19).
Briefly, cell monolayers were fixed with 4% paraformaldehyde for 10
min and permeabilized with 0.01% Triton X-100 in PBS at room
temperature for 10 min. After blocking with 3% BSA at room
temperature for 1 h, cells were incubated with a primary anti-RANKL
mouse monoclonal antibody (1:50; cat. no. ab45039; Abcam) at 4°C
overnight, followed by incubation with an Alexa Fluor 555 donkey
anti-mouse secondary antibody (1:500; cat. no. ab150106; Abcam) in
the dark at room temperature for 1 h. The coverslips were mounted
using DAPI solution (Invitrogen; Thermo Fisher Scientific, Inc.).
Slides were viewed using a fluorescent microscope (Imager Z1; Zeiss
AG). The fluorescence intensity was analyzed using Image-Pro Plus
6.0 (Media Cybernetics, Inc.).
Tartrate-resistant acid phosphatase
(TRAP) staining
RAW264.7 cells were washed with PBS and fixed in 4%
paraformaldehyde at room temperature for 30 min. Cells were then
stained with a TRAP kit (Beyotime Institute of Biotechnology),
according to the manufacturer's protocol. Cells containing three or
more nuclei were counted as positive.
Transfections
The RUNX2 overexpression plasmid was synthesized
with LV003 vector by Forevergen. Small interfering (si) RNA
targeting RUNX2 (5′-CCACTTACCACAGAGCTAT-3′) and control sequences
siRNA (5′-UUCUCCGAACGUGUCACGUTT-3′) were purchased from Forevergen.
Short hairpin (sh) RNA (5′-GGTAAAGTGTCCTGTAGGTT-3′) targeting
LAPTM5 was purchased from General Biosystems, Inc. The shRNA
sequences were inserted into the pSicoR vector (Forevergen),
according to the manufacturer's protocol. Empty pSicoR vector was
used as control. Transfections were carried out using
lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.; 2 µg overexpression plasmid, 2 µg shRNA, 200 pmol
si RNA, per well in 6-well plate), according to the manufacturer's
protocol, respectively.
Construction of the LAPTM5
promoter-luciferase reporter plasmids
Promoter fragments of LAPTM5 were obtained from ST2
cells by PCR amplification using KOD-Plus-Neo with 10X PCR Buffer
for KOD-Plus-Neo (Toyobo Life Science). The following primers were
used: −714 to −1 forward, 5′-ATCGAGCTCGTTTTTTCATCTGTGCAATGGGG-3′
and reverse, 5′-CTAGCTAGCGGTGCGCAGTCCCCTCTTC-3′; and −1572 to −1
forward, 5′-ATCGAGCTCGATTAAGCTATCCCCCCAGTGC-3′ and reverse,
5′-CTAGCTAGCGGTGCGCAGTCCCCTCTTC-3′. The thermal condition was 35
cycles of 94°C for 20 sec, 58°C for 30 sec and 68°C for 2 min and
15 sec. The amplified fragment was isolated and purified following
1% agarose gel electrophoresis using the EasyPure Quick Gel
Extraction kit (Beijing TransGen Biotech) and digested with
SacI and NheI. The fragment was ligated into the
equivalent sites of the pGL3-BASIC vector (Promega Corporation) to
yield pGL3-714, which contains no RUNX2 binding sites, and
pGL3-1572, which contains several RUNX2 binding sites. A DNA
fragment (−1 to −1572) with a substitution mutation (−1176 to
−1171; ACCACA to ACTGTA) was obtained from GeneRay Biotech Co.,
Ltd. to construct pGL3-1572m.
Dual-luciferase reporter assays
Cells were co-transfected with LAPTM5 promoter
constructs and a Renilla luciferase plasmid (pRL-TK; Promega
Corporation) using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Cells were harvested 48 h after transfection, and the
activities of firefly and Renilla luciferases were assessed
using the Stop & Glo kit (Promega Corporation). A vector
without the promoter was used as a negative control. pGL3-1572 and
pGL3-1572m were co-transfected with the RUNX2 overexpression
plasmids, using an empty vector as a control.
Chromatin immunoprecipitation
(ChIP)
ChIP assays were carried out using an EZChIP kit
(cat. no. 17-371; Merck KGaA), according to the manufacturer's
protocol. Briefly, 1% formaldehyde was added to the medium to
crosslink DNA-bound proteins to chromatin. After incubation of 10
min at room temperature, unreacted formaldehyde was quenched with
0.125 mol/l glycine. Cells were harvested and resuspended in 1 ml
of SDS lysis buffer containing a protease inhibitor cocktail and
the DNA was sheared by sonication (amplitude: 20%; for 3 min and 5
sec ON, 10 sec OFF) (JY88-IIN Ultrasonic Homogenizer; Ningo Scientz
Biotechnology Co., Ltd.). The fragmented DNA was diluted 10-fold
with dilution buffer [0.01% SDS, 1% Triton X-100, 1.2 mmol/l EDTA,
167 mmol/l NaCl, 16.7 mmol/l Tris-HCl (pH 8.1)] containing protease
inhibitor cocktail (Merck KGaA). After preclearing with protein G
agarose slurry (Merck KGaA), 5% of the supernatant was collected as
input DNA. To the remaining supernatant, 5 µg RUNX2 antibody
(1:500; cat. no. 8486; Cell Signaling Technology, Inc.) or control
immunoglobulin G (1:500; cat. no. 2729; Cell Signaling Technology,
Inc.). was added and incubated at 4°C overnight. The
immunoprecipitated complex was centrifuged (5,000 × g for 1 min at
4°C) and washed with low salt, high salt, LiCl and TE buffers in
the kit (EZChIP, Merck KGaA), according to the manufacturer's
protocols. The complex was eluted from the antibody using a
solution of 1% SDS, 0.1 mol/l NaHCO3 and 200 mmol/l
NaCl. The DNA-protein crosslinking was reversed by incubation with
5 M NaCl at 65°C overnight. All samples were treated with RNase for
30 min and proteinase K at 37°C for 2 h. DNA was purified using
spin columns provided with the kit. Samples were subjected to qPCR
(as described above). Primers specific for the LAPTM5 promoter
region were used (Table II).
| Table II.Primers used in chromatin
immunoprecipitation. |
Table II.
Primers used in chromatin
immunoprecipitation.
| Name | Forward
(5′-3′) | Reverse
(5′-3′) | Product |
|---|
| P1 |
CGGTTCTCAACCTTCCTG |
ACAGTTATGAGGTAGCAACA | −1034 to −930 (105
bp) |
| P2 |
AACGCACAATCCCAGGTTTC |
GGGCTTCTCACACATCTCCA | −1262 to −1071 (192
bp) |
| P3 |
CTGGGGGCCGTTTCTAATCTC |
CCTGGGATTGTGCGTTCTTC | −1495 to −1247 (249
bp) |
Statistical analysis
Statistical analysis was conducted using IBM SPSS
version 22.0 for Windows (IBM Corp.). The experiments were repeated
three times. All data were presented as the mean ± SD. Statistical
differences between two groups were assessed using unpaired
Student's t-test. Statistical differences among multiple groups
were assessed using one-way ANOVA and the Newman-Keuls test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Altered expression of RUNX2, LAPTM5
and RANKL in ST2 cells co-cultured with RAW264.7 cells
ST2 and RAW264.7 cells were co-cultured using
Transwell chambers for 5 days. The expression levels of the
osteoclast-specific genes osteoclast-associated immunoglobulin-like
receptor (Oscar), TRAP and nuclear factor of activated T cells 1
(Nfatc1) were upregulated significantly in RAW264.7 cells following
co-culture with ST2 cells (Fig.
1A-C). The number of TRAP-positive RAW264.7 cells also
increased significantly in the co-culture (Fig. 1D-F). These results suggested that
the osteoclastic differentiation of RAW264.7 cells was enhanced
following co-culture with ST2 cells.
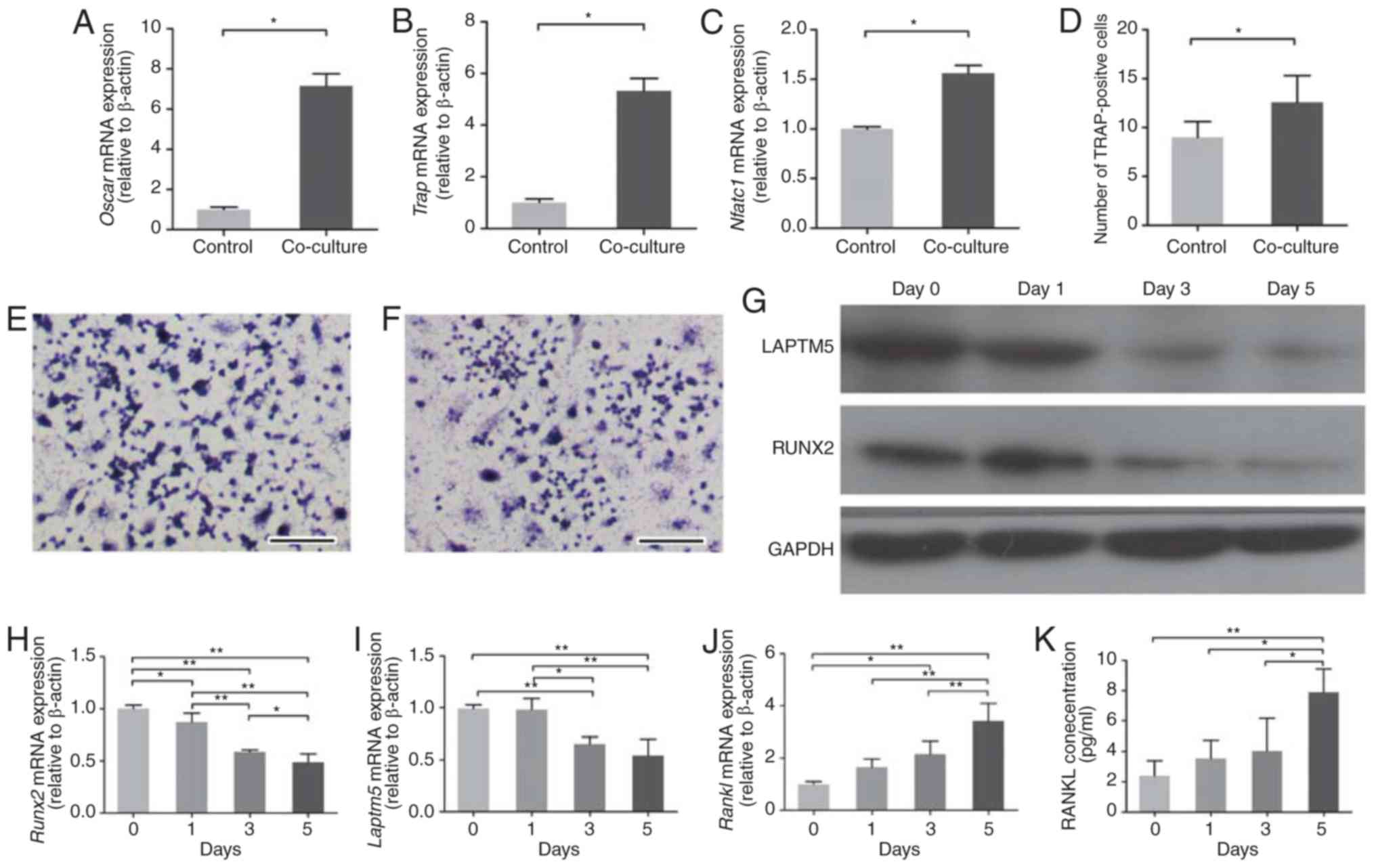 | Figure 1.Altered expression of RUNX2, LAPTM5
and RANKL in ST2 cells co-cultured with RAW264.7. (A) In the
RAW264.7 cells, the mRNA expression levels of Oscar, (B) TRAP and
(C) Nfatc1 were detected following co-culture with ST2 cells. (D)
The number of TRAP-positive RAW264.7 cells increased significantly
after 5 days of co-culture with ST2 cells. (E) Representative
TRAP-stained images of control and (F) co-cultured RAW264.7 cells.
Scale bar, 500 µm. (G) Western blotting of RUNX2 and LAPTM5 protein
expression levels in ST2 cells following the initiation of
co-culture. A representative image is shown from two independent
repeats. (H) mRNA expression levels of RUNX2, (I) LAPTM5 and (J)
RANKL were detected in ST2 cells following the indicated days of
co-culture. (K) Levels of secreted RANKL protein in the media were
detected by ELISA. Data are presented as the mean ± SD of three
independent experiments. *P<0.05, **P<0.01. Oscar,
osteoclast-associated immunoglobulin-like receptor; TRAP,
tartrate-resistant acid phosphatase; Nfatc1, nuclear factor of
activated T cells 1; RUNX2, runt related transcription factor 2;
LAPTM5, lysosomal-associated transmembrane protein 5; RANKL,
receptor activator of nuclear factor-κB ligand. |
Next, the effect of the co-culture on the ST2 cell
gene expression was examined. The mRNA expression levels of RUNX2,
LAPTM5 and RANKL in ST2 cells exhibited time-dependent alterations
following co-culture with RAW264.7 cells. Runx2 mRNA was
downregulated significantly on day 1 of co-culture, and the
downregulation continued over the following days (Fig. 1H). The protein levels of RUNX2
showed a similar trend (Fig. 1G).
The expression levels of LAPTM5 mRNA were downregulated
significantly on day 3 and 5 of co-culture (Fig. 1I). Accordingly, LAPTM5 protein
expression was also markedly decreased on day 5 (Fig. 1G). The mRNA expression levels of
RANKL were upregulated throughout the experiment (Fig. 1I). The secreted RANKL protein
levels in the culture medium were measured using ELISAs and the
results demonstrated that RANKL secreted levels increased
significantly on day 5 of o-culture (Fig. 1J).
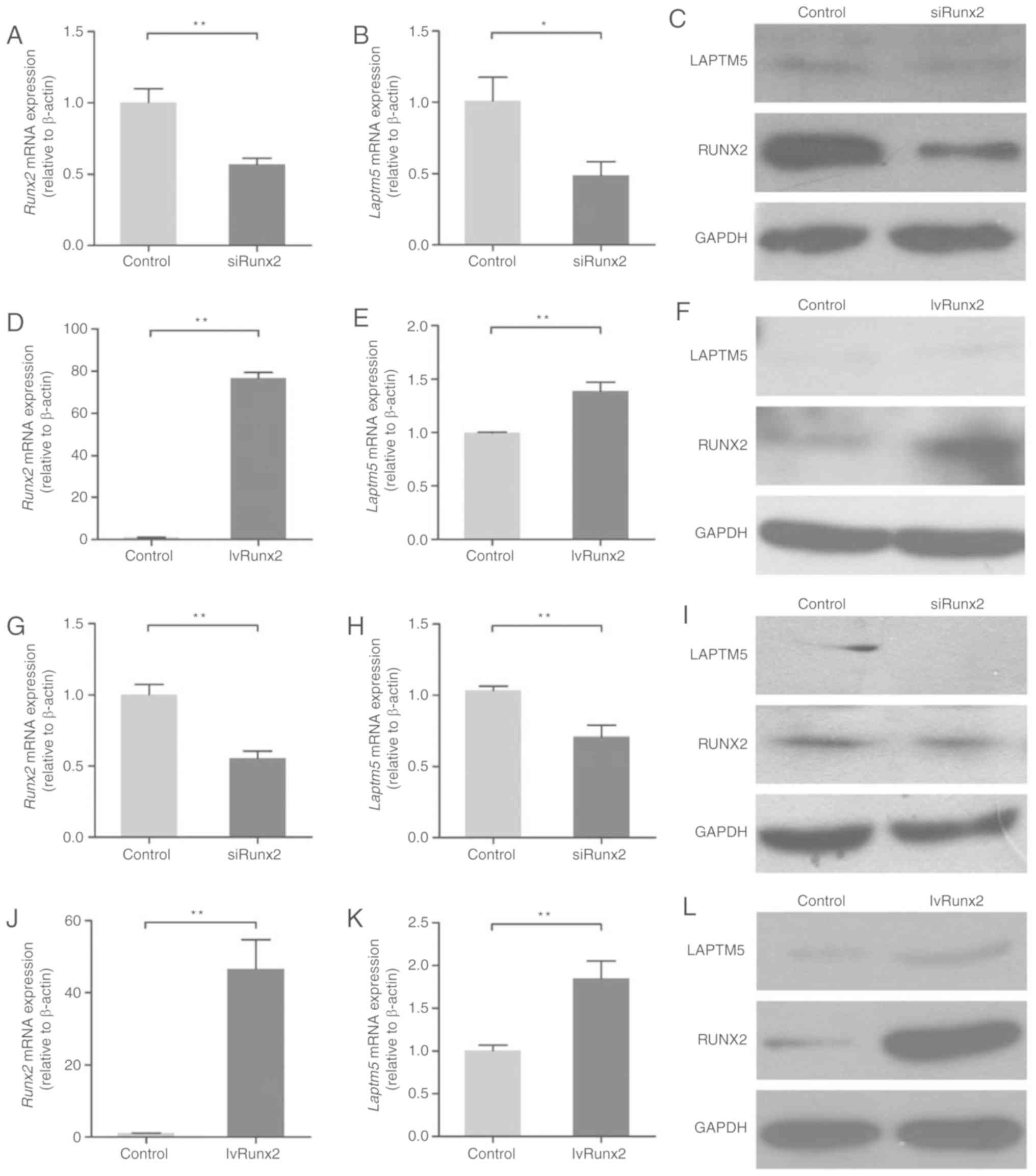
Expression of LAPTM5 is associated
with RUNX2
To explore the underlying association between RUNX2
and LAPTM5 expression, RUNX2 was silenced in ST2 cells using siRNA
(Fig. 2A and C). RUNX2 silencing
in ST2 cells resulted in reduced expression of LAPTM5 at the mRNA
and protein levels (Fig. 2B and
C). Overexpression of Runx2 in ST2 cells (Fig. 2D and F) resulted in a significant
increase in the expression of LAPTM5 at the mRNA and protein levels
(Fig. 2E and F). To further
investigate the association between these two genes, RUNX2 was also
silenced (Fig. 2G and I) or
overexpressed (Fig. 2J and L) in
POBs. The expression of LAPTM5 was decreased following RUNX2
silencing (Fig. 2H and I), while
the expression of LAPTM5 was increased following Runx2
overexpression (Fig. 2K and L).
These results suggested that the expression of LAPTM5 may be
associated with that of RUNX2.
RUNX2 regulates LAPTM5 expression by
binding to its promoter region
To test the hypothesis that LAPTM5 may be a target
gene regulated by RUNX2, the 5′-flanking region (−1 to −1500) of
the LAPTM5 gene was analyzed manually and three putative RUNX2
binding sites (ACCACA) were identified [-1012 to −1007/-980 to −975
(P1), −1176 to −1171 (P2), and −1456 to −1451 (P3); Fig. 3A].
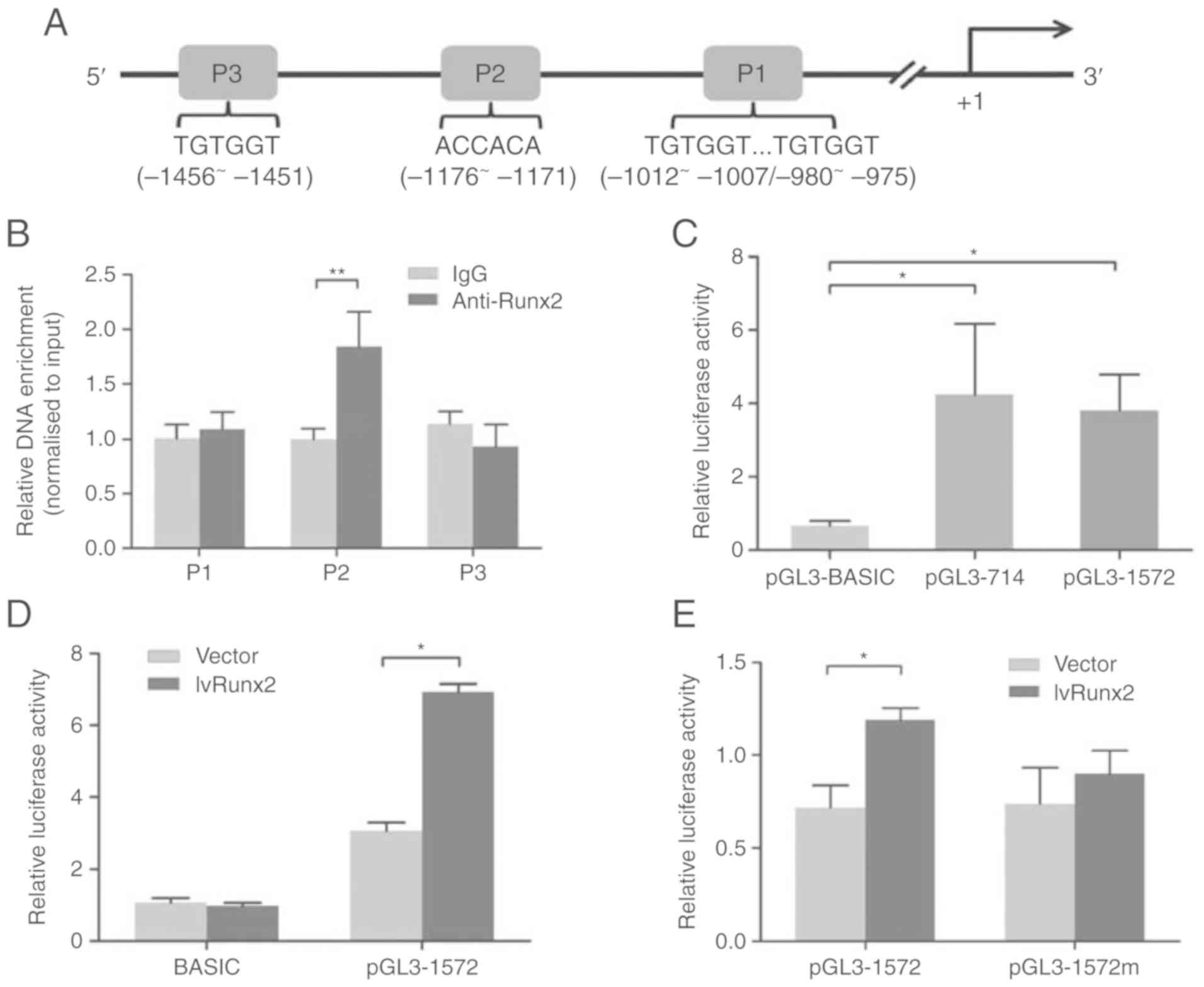 | Figure 3.RUNX2 regulates LAPTM5 expression by
binding its promoter region. (A) Schematic diagram of the LAPTM5
promoter region, where P1, P2 and P3 indicate putative RUNX2
binding sites. The transcription start site is marked as +1. (B)
Chromatin immunoprecipitation assays were used to assess RUNX2
binding to the LAPTM5 promoter in ST2 cells. (C) ST2 cells were
transfected with the pGL3-derived reporter constructs pGL3-714,
which contains no RUNX2 binding sites, or pGL3-1572, which contains
the P1, P2 and P3 binding sites. The empty vector pGL3-BASIC was
used as a control. The luciferase activity, normalized to
Renilla luciferase activity, was analyzed 48 h
post-transfection. (D) Cells were co-transfected with the pGL3-1572
vector (using the empty vector pGL3-BASIC as a control) alongside
the lvRUNX2 overexpression vector (using the empty LV003 vector as
a control). The luciferase activity, normalized to Renilla
luciferase activity, was analyzed 48 h post-transfection. (E) A
substitution mutation in the P2 site was introduced into the
pGL3-1572 vector, yielding the pGL3-1572m reporter. Cells
co-transfected with the pGL3-1572m and the lvRUNX2 overexpression
vector and relative luciferase activity was analyzed 48 h
post-transfection. Data are presented as the mean ± SD of two
independent experiments. *P<0.05, **P<0.01. RUNX2, runt
related transcription factor 2; LAPTM5, lysosomal-associated
transmembrane protein 5; IgG, immunoglobulin G. |
A ChIP assay was performed to determine whether
RUNX2 binds to the LAPTM5 promoter. DNA-protein complexes were
immunoprecipitated using a RUNX2 antibody. DNA enrichment in the
complexes was analyzed by qPCR. The results revealed that the
sequence containing P2 was enriched in DNA-protein immune
complexes, while those containing P1 and P3 were not (Fig. 3B), suggesting that RUNX2 was able
to bind the LAPTM5 promoter at the −1176 to −1171 position. Next,
dual-luciferase reporter assays were used to investigate the effect
of RUNX2 on LAPTM5 promoter activation. The relative luciferase
activities were significantly increased in cells transfected with
pGL3-1572 and pGL3-714 compared with the control group. There was
no significant difference between the activities of pGL3-714 and
pGL3-1572 (Fig. 3C). Considering
the putative RUNX2 binding sites, pGL3-1572 was used for further
study. The relative luciferase activity of pGL3-1572 in
RUNX2-overexpressing cells displayed a significant increase in
activity relative to the control, while pGL3-BASIC displayed no
significant alteration (Fig. 3D).
Based on the results of the ChIP assay, a substitution mutation in
the RUNX2 binding site (−1176 to −1171; ACCACA to ACTGTA) was
introduced into the pGL3-1572 reporter (pGL3-1572m). Overexpression
of Runx2 increased pGL3-1572 reporter activities significantly
compared with the control, however, the mutation diminished this
effect (Fig. 3E). Together, these
results demonstrated that RUNX2 transactivated the LAPTM5 gene by
binding to the LAPTM5 promoter at position −1176 to −1171.
LAPTM5 is involved in the trafficking
of RANKL
To investigate the relationship between the
expression of LAPTM5 and RANKL, LAPTM5 was silenced using shRNA
(Fig. 4A and B). The expression of
RANKL mRNA was not influenced by the knockdown of LAPTM5 (Fig. 4C). However, the concentration of
secreted RANKL protein in the culture medium was increased
significantly following LAPTM5 knockdown (Fig. 4D). RANKL was detected in the
cytoplasm using immunofluorescence staining (Fig. 4E). Notably, the fluorescence
intensity in the cytoplasm was significantly increased following
LAPTM5 knockdown (Fig. 4F),
suggesting that LAPTM5 knockdown enhanced RANKL accumulation in the
cytoplasm. Furthermore, the expression levels of Oscar, TRAP and
Nfatc1 were significantly upregulated (Fig. 4G-I) and the number of TRAP-positive
cells was significantly increased (Fig. 4J) in RAW264.7 cells when
co-cultured with LPTM5-depleted ST2 cells. Taken together, these
results suggested that LAPTM5 is involved in the trafficking of
RANKL.
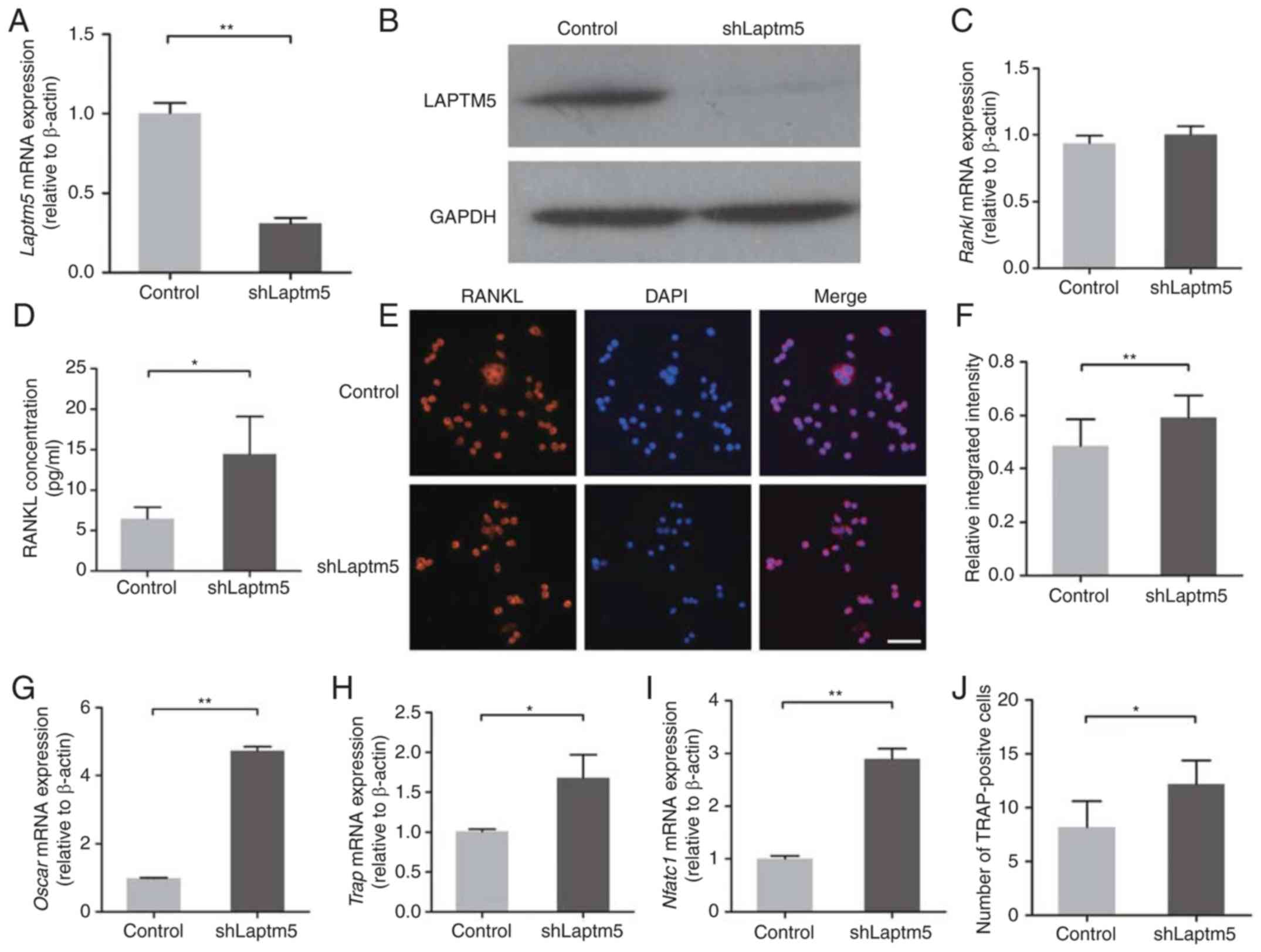 | Figure 4.LAPTM5 is involved in RANKL
trafficking. (A) LAPTM5 was silenced in ST2 cells using shRNA, and
successful knockdown was confirmed at the mRNA and (B) the protein
level. (C) RANKL mRNA expression levels exhibited no significant
alteration following LAPTM5 knockdown. (D) The concentration of
secreted RANKL protein in the culture medium was increased
significantly following LAPTM5 knockdown. (E) RANKL expression in
ST2 cells was determined using immunofluorescence staining. Nuclei
were stained with DAPI. Scale bar, 50 µm. (F) Relative integrated
intensity was calculated as the ratio of integrated fluorescence
intensity of RANKL in the cytoplasm to that of the total cell. All
cells in three random fields were analyzed using Image-Pro Plus
6.0. (G) mRNA expression levels of RUNX2, (H) TRAP and (I) Nftac1
were detected in RAW264.7 cells co-cultured with either control or
LAPTM5-silenced ST2 cells. (J) The number of TRAP-positive cells
increased significantly in RAW264.7 cells following co-culture with
LAPTM5-silenced ST2 cells. Cells transfected with empty vectors
were used as the control. Data are presented as the mean ± SD of
three independent experiments. *P<0.05, **P<0.01. RUNX2, runt
related transcription factor 2; LAPTM5, lysosomal-associated
transmembrane protein 5; RANKL, receptor activator of nuclear
factor-κB ligand; sh, short hairpin; Oscar, osteoclast-associated
immunoglobulin-like receptor; TRAP, tartrate-resistant acid
phosphatase; Nfatc1, nuclear factor of activated T cells 1. |
Discussion
The results of the present study indicated that
RUNX2 transactivated the expression of the LAPTM5 gene by binding
to its promoter region and that LAPTM5 was involved in the
expression trafficking of RANKL in ST2 cells.
In the current study, utilizing the co-culture of
ST2 and RAW264.7 cells, increased concentrations of RANKL in the
medium were observed. Subsequently, the expression levels of Oscar,
TRAP and Nfatc1 were demonstrated to be elevated in RAW264.7 cells.
These genes are RANKL-inducible and are generally considered to be
markers of osteoclast differentiation (20). In addition, an increased number of
TRAP-positive cells was detected following co-culture. These
results indicated that osteoclastogenic signals were induced by ST2
cells. By using this conventional co-culture model, a previous
study has indicated that soluble RANKL mediates osteoclastogenesis
(21).
In the presence of
1,25-(OH)2D3, RUNX2 and LAPTM5 were
downregulated in ST2 cells with time. The downregulation of RUNX2
can be attributed to the binding of the vitamin D receptor to the
promoter of RUNX2 (22), however,
the mechanism of regulation of LAPTM5 expression is not clear. As
it has been suggested that the forced expression of RUNX2
upregulates LAPTM5 expression (11), the relationship between RUNX2 and
LAPTM5 was explored in the present study by overexpressing and
depleting RUNX2. The results demonstrated that the expression of
LAPTM5 was regulated by RUNX2. To investigate the underlying
mechanism of how RUNX2 regulated LAPTM5 expression, luciferase
reporter and ChIP assays were conducted with a focus on the −1 to
−1500 5′-flanking region of the LAPTM5 gene. The ChIP assay results
revealed that RUNX2 directly bound to the −1176 to −1171 position
within the LAPTM5 promoter. The combination of the luciferase
reporter assay and the overexpression of RUNX2 revealed that RUNX2
could directly activate the LAPTM5 promoter and that this
activation was prevented by introducing a mutation in the binding
site at −1176 to −1171. These findings provided evidence that RUNX2
regulated the expression of LAPTM5 through its role as a
transcription factor. When validating this finding using luciferase
reporters, it was found that a reporter construct without the RUNX2
putative binding site exhibited an elevated level of activity. This
result indicated that the LAPTM5 gene is likely to be regulated by
multiple transcription factors.
The lysosomal system is not only involved in
osteogenesis but also regulates osteoclastogenesis (23). In differentiating osteoblasts,
lysosome levels are increased and are dispersed towards the cell
periphery (24). Bone
matrix-containing vesicles are partially associated with lysosomes
in osteoblasts (24,25). Amorphous calcium/phosphate crystals
are secreted through lysosomal exocytosis (26). The osteoclastogenic cytokine RANKL
is delivered to the cell surface through secretory lysosomes
(27,28) and OPG regulates RANKL-induced
osteoclastogenesis by mediating the transport of RANKL to the
lysosomal storage compartments (29). The fact that LAPTM5 is
transactivated by RUNX2, a transcription factor involved in both
osteogenesis and osteoclastogenesis, indicates that LAPTM5 may be
an important protein for understanding the role of the lysosomal
system in the coupling of osteogenesis and osteoclastogenesis.
In the present study, the upregulation of RANKL was
accompanied by the downregulation of LAPTM5 and RUNX2. To the best
of our knowledge, RUNX2 has not previously been shown to have a
role in RANKL transactivation (30), therefore, the relationship between
LAPTM5 and RANKL was examined in the present study. Knockdown of
LAPTM5 yielded no significant effect on the mRNA expression of
RANKL, however, it significantly increased the levels of RANKL
protein in the cytoplasm and its secretion into the culture medium.
These results suggested that the upregulation of RANKL could be
attributed to suppressed lysosomal proteolytic activity following
LAPTM5 silencing. This is consistent with a previous study that
reported the increased expression of the surface pre-B cell
receptor as a result of LAPTM5 deficiency (31). Future work is required to determine
whether LAPTM5 is involved in the lysosomal trafficking of RANKL to
the cell surface, or if it regulates the degradation of RANKL by
targeting it to lysosomal vesicles; this is specifically relevant
in the cellular context, in which RANKL expression on the plasma
membrane is lysosome-dependent (23).
The present study indicated that the downregulation
of RUNX2 may increase the expression of RANKL in the cytoplasm by
suppressing the expression of LAPTM5. These findings add to the
existing evidence that RUNX2 serves an important role in
osteoclastogenesis (32). However,
it appears that both the down and upregulation of RUNX2 can induce
osteoclastogenesis through the regulation of RANKL (6). Upregulation of RUNX2 promotes
osteoblast-induced osteoclastogenesis by regulating the association
of RANKL with the membrane (6).
Aside from regulation at the transcriptional level, the function of
RUNX2 is also regulated by additional mechanisms, including
posttranslational modifications under variable conditions (33). It was reported that the
phosphorylation of RUNX2 is required to induce gene expression in
response to mechanical stimuli, which is thought to regulate bone
remodeling (34); this finding
emphasizes the importance of the posttranslational modification of
Runx2 in the coupling of osteogenesis and osteoclastogenesis.
In conclusion, the present study demonstrated that
RUNX2 transactivated LAPTM5 gene expression and that LAPTM5 was
involved in the trafficking of RANKL. These findings suggested that
RUNX2 may regulate osteoclast differentiation through
lysosome-associated genes that modulate RANKL trafficking in bone
cells. The present results suggested a possible coupling mechanism
between osteogenesis and osteoclastogenesis.
Acknowledgements
Not applicable.
Funding
The present study was financially supported by The
Science Foundation for the Youth Scholars of Southern Medical
University (grant no. PY2015N018), The National Natural Science
Foundation of China (grant no. 81271187), The Science &
Technology Projects of Guangzhou (grant no. 201707010199) and The
Guangdong Provincial Science & Technology Projects (grant nos.
2013B021800319, 2014A020212239 and 2017A020215050).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
YG performed experiments, performed the statistical
analysis and drafted the manuscript. CL, WYL, PL, PY and WLL
performed the experiments. PX designed the study and drafted the
manuscript. XS designed the study and participated in the
experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Procedures involving animals were approved by The
Institutional Animal Care and Use Committee of Guangzhou Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Florencio-Silva R, Sasso GR, Sasso-Cerri
E, Simões MJ and Cerri PS: Biology of bone tissue: Structure,
function, and factors that influence bone cells. Biomed Res Int.
2015:4217462015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu W and Zhang X: Receptor activator of
nuclear factor-κB ligand (RANKL)/RANK/osteoprotegerin system in
bone and other tissues (review). Mol Med Rep. 11:3212–3218. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Long F: Building strong bones: Molecular
regulation of the osteoblast lineage. Nat Rev Mol Cell Biol.
13:27–38. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Komori T, Yagi H, Nomura S, Yamaguchi A,
Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, et al:
Targeted disruption of Cbfa1 results in a complete lack of bone
formation owing to maturational arrest of osteoblasts. Cell.
89:755–764. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Geoffroy V, Kneissel M, Fournier B, Boyde
A and Matthias P: High bone resorption in adult aging transgenic
mice overexpressing cbfa1/runx2 in cells of the osteob lastic
lineage. Mol Cell Biol. 22:6222–6233. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martin A, Xiong J, Koromila T, Ji JS,
Chang S, Song YS, Miller JL, Han CY, Kostenuik P, Krum SA, et al:
Estrogens antagonize RUNX2-mediated osteoblast-driven
osteoclastogenesis through regulating RANKL membrane association.
Bone. 75:96–104. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ducy P, Zhang R, Geoffroy V, Ridall AL and
Karsenty G: Osf2/Cbfa1: A transcriptional activator of osteoblast
differentiation. Cell. 89:747–754. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kern B, Shen J, Starbuck M and Karsenty G:
Cbfa1 contributes to the osteoblast-specific expression of type I
collagen genes. J Biol Chem. 276:7101–7107. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mori K, Kitazawa R, Kondo T, Maeda S,
Yamaguchi A and Kitazawa S: Modulation of mouse RANKL gene
expression by Runx2 and PKA pathway. J Cell Biochem. 98:1629–1644.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
O'Brien CA: Control of RANKL gene
expression. Bone. 46:911–919. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baniwal SK, Shah PK, Shi Y, Haduong JH,
Declerck YA, Gabet Y and Frenkel B: Runx2 promotes both
osteoblastogenesis and novel osteoclastogenic signals in ST2
mesenchymal progenitor cells. Osteoporos Int. 23:1399–1413. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Adra CN, Zhu S, Ko JL, Guillemot JC,
Cuervo AM, Kobayashi H, Horiuchi T, Lelias JM, Rowley JD and Lim B:
LAPTM5: A novel lysosomal-associated multispanning membrane protein
preferentially expressed in hematopoietic cells. Genomics.
35:328–337. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pak Y, Glowacka WK, Bruce MC, Pham N and
Rotin D: Transport of LAPTM5 to lysosomes requires association with
the ubiquitin ligase Nedd4, but not LAPTM5 ubiquitination. J Cell
Biol. 175:631–645. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ouchida R, Kurosaki T and Wang JY: A role
for lysosomal-associated protein transmembrane 5 in the negative
regulation of surface B cell receptor levels and B cell activation.
J Immunol. 185:294–301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Angenieux C, Waharte F, Gidon A,
Signorino-Gelo F, Wurtz V, Hojeij R, Proamer F, Gachet C, Van
Dorsselaer A, Hanau D, et al: Lysosomal-associated transmembrane
protein 5 (LAPTM5) is a molecular partner of CD1e. PLoS One.
7:e426342012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawai Y, Ouchida R, Yamasaki S, Dragone L,
Tsubata T and Wang JY: LAPTM5 promotes lysosomal degradation of
intracellular CD3ζ but not of cell surface CD3ζ. Immunol Cell Biol.
92:527–534. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen XQ, Geng YM, Liu P, Huang XY, Li SY,
Liu CD, Zhou Z and Xu PP: Magnitude-dependent response of
osteoblasts regulated by compressive stress. Sci Rep. 7:449252017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim EJ, Lee MY and Jeon YJ: Silymarin
inhibits morphological Changes in LPS-stimulated macrophages by
blocking NF-κB pathway. Korean J Physiol Pharmacol. 19:211–218.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song I, Kim JH, Kim K, Jin HM, Youn BU and
Kim N: Regulatory mechanism of NFATc1 in RANKL-induced osteoclast
activation. FEBS Lett. 583:2435–2440. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Furuya M, Kikuta J, Fujimori S, Seno S,
Maeda H, Shirazaki M, Uenaka M, Mizuno H, Iwamoto Y, Morimoto A, et
al: Direct cell-cell contact between mature osteoblasts and
osteoclasts dynamically controls their functions in vivo. Nat
Commun. 9:3002018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Drissi H, Pouliot A, Koolloos C, Stein JL,
Lian JB, Stein GS and van Wijnen AJ: 1,25-(OH)2-vitamin D3
suppresses the bone-related Runx2/Cbfa1 gene promoter. Exp Cell
Res. 274:323–333. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Honma M, Ikebuchi Y, Kariya Y, Hayashi M,
Hayashi N, Aoki S and Suzuki H: RANKL subcellular trafficking and
regulatory mechanisms in osteocytes. J Bone Miner Res.
28:1936–1949. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nabavi N, Urukova Y, Cardelli M, Aubin JE
and Harrison RE: Lysosome dispersion in osteoblasts accommodates
enhanced collagen production during differentiation. J Biol Chem.
283:19678–19690. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao H, Ito Y, Chappel J, Andrews NW,
Teitelbaum SL and Ross FP: Synaptotagmin VII regulates bone
remodeling by modulating osteoclast and osteoblast secretion. Dev
Cell. 14:914–925. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rohde M and Mayer H: Exocytotic process as
a novel model for mineralization by osteoblasts in vitro and in
vivo determined by electron microscopic analysis. Calcif Tissue
Int. 80:323–336. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kariya Y, Honma M, Aoki S, Chiba A and
Suzuki H: Vps33a mediates RANKL storage in secretory lysosomes in
osteoblastic cells. J Bone Miner Res. 24:1741–1752. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kariya Y, Honma M, Hanamura A, Aoki S,
Ninomiya T, Nakamichi Y, Udagawa N and Suzuki H: Rab27a and Rab27b
are involved in stimulation-dependent RANKL release from secretory
lysosomes in osteoblastic cells. J Bone Miner Res. 26:689–703.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Aoki S, Honma M, Kariya Y, Nakamichi Y,
Ninomiya T, Takahashi N, Udagawa N and Suzuki H: Function of OPG as
a traffic regulator for RANKL is crucial for controlled
osteoclastogenesis. J Bone Miner Res. 25:1907–1921. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
O'Brien CA, Kern B, Gubrij I, Karsenty G
and Manolagas SC: Cbfa1 does not regulate RANKL gene activity in
stromal/osteoblastic cells. Bone. 30:453–462. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kawano Y, Ouchida R, Wang JY, Yoshikawa S,
Yamamoto M, Kitamura D and Karasuyama H: A novel mechanism for the
autonomous termination of pre-B cell receptor expression via
induction of lysosome-associated protein transmembrane 5. Mol Cell
Biol. 32:4462–4471. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Haxaire C, Haÿ E and Geoffroy V: Runx2
controls bone resorption through the Down-regulation of the wnt
pathway in osteoblasts. Am J Pathol. 186:1598–1609. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jonason JH, Xiao G, Zhang M, Xing L and
Chen D: Post-translational regulation of Runx2 in bone and
cartilage. J Dent Res. 88:693–703. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li Y, Ge C, Long JP, Begun DL, Rodriguez
JA, Goldstein SA and Franceschi RT: Biomechanical stimulation of
osteoblast gene expression requires phosphorylation of the RUNX2
transcription factor. J Bone Miner Res. 27:1263–1274. 2012.
View Article : Google Scholar : PubMed/NCBI
|