Introduction
Cervical cancer is the most common malignant tumor
of the female reproductive system. Its incidence continues to
increase, while the average age of onset has decreased (1). At present, the methods of treating
cervical cancer are predominantly radiotherapy, platinum-based
chemotherapy and surgery. Although the traditional surgical method
remains the main effective approach towards treating cervical
cancer, it greatly affects the sexual and reproductive functions of
the patients (2). In addition,
although radiation equipment is constantly improving, and
chemotherapy drugs are continually being developed, radiotherapy
and chemotherapy continue to cause a series of systemic side
effects that result in a reduction in the quality of the patients'
lives (3). Therefore, the search
for a safe, effective and non-invasive treatment that can preserve
fertility and decrease the effects on the sexual function of young
patients is an important research direction.
Pulsed electric field (PEF) is a biomedical
engineering technique for tumor therapy that has been developed in
recent years. The effect of electrical pulses on the cell structure
and function, and as a biological treatment, has become a focus in
the biological electromagnetics field (4,5).
According to the different pulse durations, PEF may be classified
into millisecond (ms)-, microsecond (μs)-, nanosecond (ns)- and
picosecond (ps) PEF. A large number of studies have investigated
ms-, μs- and nsPEF.
It was demonstrated that when cells are exposed to
certain millisecond or microsecond pulses, a number of aqueous
channels form in the cell membrane, in a process known as
reversible electroporation (RE) (6). Electroporation enhances the membrane
permeability significantly, which is beneficial in enabling cells
to absorb drug molecules, genetic material, protein and other
macromolecular material, without damaging the intracellular
organelles, such as the nucleus and the mitochondria. Based on this
feature, the electroporation technique has been used for gene
transfection (7). The theory of
electroporation has also been used in a new therapeutic approach to
target tumors. This method, known as electrical chemotherapy (ECT),
combines chemotherapy with PEF to reduce the systemic use of
chemotherapy drugs and enhance the influx of chemotherapeutic
agents into cells. This therapeutic method has been successfully
used in various clinical therapies, including the treatment of
head, neck, skin, pancreatic and liver cancer (9,10).
However, if the electric field strength increases beyond a certain
level, irreversible electrical breakdown occurs in the cell
membrane, and the cancer cells are killed directly(11), a process termed irreversible
electrical breakdown (IREB). It has been demonstrated that when
microsecond pulses are applied in living pork liver, these induce
not only cell RE and IREB, but also cell apoptosis (12). Our previous study indicated that
IREB induced by a low voltage and with a higher number of pulses
ablated HeLa cells as effectively as a high voltage. The
combination of a low voltage and a higher number of pulses also
resulted in the desired outcome of a greater proportion of
apoptosis occurring than necrosis (13). A further reduction in the pulse
duration, to nanoseconds, leads to a series of cell responses, such
as apoptosis, intracellular calcium release and increased gene
expression, as well as DNA and chromosome damage, despite the outer
membrane remaining intact (14–17).
This technique is known as intracellular electromanipulation.
Although ms-, μs- and nsPEF have numerous biological
effects and clinical applications, they continue to require the use
of an invasive or minimally invasive electrode array (needles or
plate electrodes) to guide the puncture into the tumor tissue. In
addition, each type of PEF has different features that lead to
further limitations. As a result, these three types of PEF are not
able to meet the requirements that necessitate a treatment that
preserves fertility and minimizes the effect on the sexual function
of young patients in a safe, effective and non-invasive manner.
psPEF has an ultra-wideband spectrum [almost from
direct current (DC) to GHz], with a high time and spatial
resolution and a low signal distortion (18,19).
This means that there is the potential for psPEF to be transferred
to target deep tissue non-invasively and precisely, although a
method of achieving this transfer to the human body non-invasively
has not yet been established. Chongqing University (Chongqing,
China), the experimental cooperation unit for this study, has
developed an ultra-wideband impulse radiating antenna (IRA), which
is composed of an ellipsoid reflector and a single cone radiator.
This IRA is capable of transferring energy within a specific range,
converging the pulses of electromagnetic waves in the designated
area and destroying the target. The radiation energy is thus highly
concentrated in time and space. Based on the optical principle, the
IRA may potentially be used to transfer and focus the psPEF to
target deep biological tissue without trauma. This would enable
psPEF to kill tumor cells, while avoiding damage to the surrounding
healthy tissue. If successful, psPEF may be considered as a
non-invasive treatment for tumors (20). However, a limited number of studies
have investigated the biological effects induced in tumor cells by
psPEF.
Apoptosis, a type of programmed cell death, is
important in the normal functioning of the body; a departure from
normal apoptotic mechanisms may lead to malignant tumor formation
(21). We have previously studied
the antitumor effects of μsPEF and nsPEF. In this study, we exposed
HeLa cells to different psPEF conditions. MTT assay, flow cytometry
and transmission electron microscopy (TEM) were used to detect the
effects of the psPEF on HeLa cell growth inhibition, apoptosis and
the cell cycle. In addition, we investigated the interrelation
between the psPEF and biological effects induced in the HeLa cells,
by changing the psPEF parameters. The aim of this study was to
establish the parameters of psPEF that induced the killing effect
on tumor cells, in order to lay a foundation for subsequent
experiments in vivo and for clinical trials. As a result, a
non-invasive treatment that preserves the reproductive function of
young cervical cancer patients may be realized.
Materials and methods
Chemicals and reagents
RPMI-1640 medium and fetal bovine serum (FBS) were
purchased from Gibco (Grand Island, NY, USA). MTT and
dimethylsulfoxide (DMSO) were obtained from Sigma (St. Louis, MO,
USA), and the Annexin V-fluorescein isothiocyanate (FITC) apoptosis
detection kit was purchased from Nanjing KeyGen Biotech Co., Ltd.
(Nanjing, China).
Cell lines and cultures
Human cervical adenocarcinoma HeLa cells were
obtained from The Institute of Ultrasound Engineering in Medicine
of Chongqing Medical University (Chongqing, China). The cells were
cultured in RPMI-1640 medium supplemented with 10% FBS and 1%
penicillin-streptomycin at 37°C in a 5% humidified CO2
incubator. When the HeLa cells reached 70–80% confluence, cells
were subcultured or treated with psPEF.
psPEF treatment
Having been washed three times with
phosphate-buffered saline (PBS), cells were combined with 0.125%
trypsin-EDTA, and then centrifuged at 800 × g for 5 min. Cells were
then resuspended in fresh RPMI-1640 medium, at a concentration of
2×106 cells/ml. A total of 100 μl cell suspension was
subsequently placed into the cuvette and exposed to 800-psec pulses
with a frequency of 3 Hz, and a corresponding electric field
amplitude and pulse number. Cells that were loaded into cuvettes
and placed into the circuit without being pulsed were used as the
normal controls. The electric field amplitude and pulse width were
monitored throughout the procedures with a DP04054 oscilloscope
(Tektronix, Inc., Beaverton, OR, USA).
MTT assay
There were two purposes in performing the assay: i)
to establish the length of incubation time the cells required
post-pulses in order to achieve a level of maximum cell inhibition;
ii) to establish whether intense psPEF was capable of affecting
cell viability, and, if it was capable, whether there was any
correlation between electric field amplitude, pulse number and cell
inhibition. For the first purpose, cells were exposed to 800-psec
pulses at a frequency of 3 Hz, a pulse number of 2,000 and
different electric field amplitudes (100, 200, 300, 400, 500 and
600 kV/cm). Following the electrical pulse treatment, HeLa cells
were seeded at a density of 5,000 cells/well in 96-well plates
containing 150 μl RPMI-1640 medium with 10% FBS. Cells were
subsequently routinely cultured for 6, 12, 24, 36 and 48 h in a 5%
CO2 humidified incubator at 37°C, respectively. A
normal, control and a blank group (without cells) were included.
Following the corresponding culture time, 20 μl MTT (5 mg/ml) was
added to each well, and the cells were incubated for an additional
4 h. The culture medium was discarded by gentle aspiration and
replaced with 150 μl DMSO, prior to the plates being agitated for
10 min to dissolve the formazan crystals. The absorbance was then
measured with an EL×800 absorbance microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA) at a wavelength of 490 nm.
For the second purpose, the cells were divided into six groups
according to electric field amplitude (100, 200, 300, 400, 500 and
600 kV/cm), with each group being further divided into several
subgroups according to pulse number (1,000, 2,000 and 3,000). The
remaining parameters of each group were the same (800-psec pulses
and a frequency of 3 Hz). Cells of each group were treated with the
corresponding psPEF. Following treatment, the cells were incubated
in 96-well plates for 12 h. The subsequent steps in the procedure
corresponded to those in the MTT assay mentioned previously. This
assay was performed in quintuplicate and the experiments were
repeated a minimum of three times with similar results. As
established from the MTT results, we selected the electric field
amplitudes of 200, 400 and 600 kV/cm as the low-, middle- and
high-amplitude groups in the subsequent tests.
TEM analysis
Cell morphology was observed using TEM. Following
treatment with 800-psec pulses of PEF with electric field ampitudes
of 200, 400 and 600 kV/cm, respectively, a pulse number of 2,000
and a frequency of 3 Hz, the cells (including the untreated group)
were harvested and grown in RPMI-1640 medium containing 10% FBS for
12 h. Both floating and adherent cells were subsequently harvested
and centrifuged at 800 × g for 5 min, and then at 1,200 × g for 10
min. The cells were fixed overnight in 0.2 M sodium cacodylate
buffer solution (pH 7.4) containing 2% glutaraldehyde at 4°C. The
samples were then post-fixed in cacodylate-buffered 1% osmium
tetroxide, dehydrated, and embedded in Epon 812 for ultra-thin
sectioning. Following this, ultra-thin sections were stained with
uranyl acetate and lead citrate, and observed with a transmission
electron microscope (H-7500; Hitachi, Tokyo, Japan) at the
Chongqing Medical University Cell Imaging Facility. A total of 100
cells were sampled for each group.
Flow cytometry for cell apoptosis
analysis
Cell apoptosis was identified by flow cytometry with
an apoptosis detection kit. Groups were divided according to
electric field amplitude and pulse number, respectively. For the
division by electric field amplitude, the cells were exposed to
800-psec pulses with a frequency of 3 Hz, a pulse number of 2,000
and with various electric field amplitudes (200, 400 and 600
kV/cm). For the division by pulse number, the cells were exposed to
800-psec pulses with a frequency of 3 Hz, a 400 kV/cm electric
field amplitude and a range of pulse numbers (1,000, 2,000 and
3,000). Following their respective treatments, the cells, including
those in the control group, were collected and seeded in 6-well
plates containing 1.5 ml RPMI-1640 medium with 10% FBS, and then
cultured for 12 h in a 5% CO2 humidified incubator at
37°C. Following a period of 12 h, floating and adherent cells were
harvested from each well. These HeLa cells were then washed with
ice-cold PBS, and resuspended at a density of 1×106
cells/ml. A total of 100 μl cell suspension was added to the flow
tube and double-stained with Annexin V and propidium iodide (PI).
Following incubation for 15 min in the dark, the fluorescence in
the cells was quantitatively analyzed at an emission wavelength of
530 nm and an excitation wavelength of 480 nm, using a
fluorescence-activated cell sorting (FACS) Vantage SE flow
cytometer system (BD Biosciences, Franklin Lakes, NJ, USA).
Flow cytometry for cell cycle
analysis
In this study, HeLa cells were exposed to 800-psec
pulses with pulse number 2,000, various electric field amplitudes
(200, 400 and 600 kV/cm) and a frequency of 3 Hz. When the
electrical pulse treatment was complete, the HeLa cells, including
the control group, were seeded in 6-well plates containing 1.5 ml
RPMI-1640 medium with 10% FBS, and then cultured for 12 h in a 5%
CO2 humidified incubator at 37°C. Following a period of
12 h, floating and adherent cells were harvested from each well and
centrifuged at 800 × g for 5 min. The cells were then washed and
fixed with ice-cold alcohol (75%) for >24 h. Following a further
two washes, cells were incubated with PBS (pH 7.4), containing
RNase (5 units) and PI for 15 min at 37°C. Cell cycle distribution
was then measured with the FACSVantage SE flow cytometer (BD
Biosciences).
Statistical analysis
SPSS 17.0 statistical software (SPSS, Inc., Chicago,
IL, USA) was used for the statistical analysis. The statistical
data were presented as the mean ± standard deviation of values from
three independent experiments. Comparisons between groups were
performed by the student's t-test and one-way analysis of variance
(ANOVA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of psPEF on HeLa cell viability
following different culture times
Cell proliferation was determined by MTT assay. HeLa
cells were processed as described in Materials and methods. The
cell survival rate in normal control cells was taken as 100%
viability. The percentage of cell inhibition was determined as
follows: % cell inhibition = (absorbance of normal control cells -
absorbance of treated cells)/(absorbance of normal control cells -
absorbance of blank group) × 100. The growth inhibition of HeLa
cells at different culture times following various electric field
amplitude treatments, as assessed by MTT assay and calculated by
the previously mentioned formula, is shown in Fig. 1. In the electric field amplitude
groups (100, 200, 300 and 400 kV/cm), there were notable
differences in the growth inhibition rates of the HeLa cells in the
12-h groups (10.20±1.53, 19.10±2.873, 28.56±4.00 and 46.20±6.93%,
respectively) compared with the 6-h groups (4.50±0.63, 8.90±1.36,
15.21±2.28 and 22.12±3.32%, respectively) (P<0.01). There were
also significant differences in the growth inhibition rates between
the 6-h groups (35.23±5.28 and 50.56±7.58%) and 12-h groups
(56.12±8.42 and 71.23±10.60%) at 500 and 600 kV/cm, respectively
(P<0.05). However, there were no significant differences in the
growth inhibition rates between the 12- and 24-h groups
(P>0.05). No significant differences were noted between the 24-
and 36-h, or the 36- and 48-h groups (P>0.05). The results
demonstrated that at a given electric field amplitude, electrical
pulses achieved a plateau of maximum cell inhibition 12 h following
the pulse treatment. On the basis of these data, the cells were
routinely cultured for 12 h following the treatment in the
subsequent experiments.
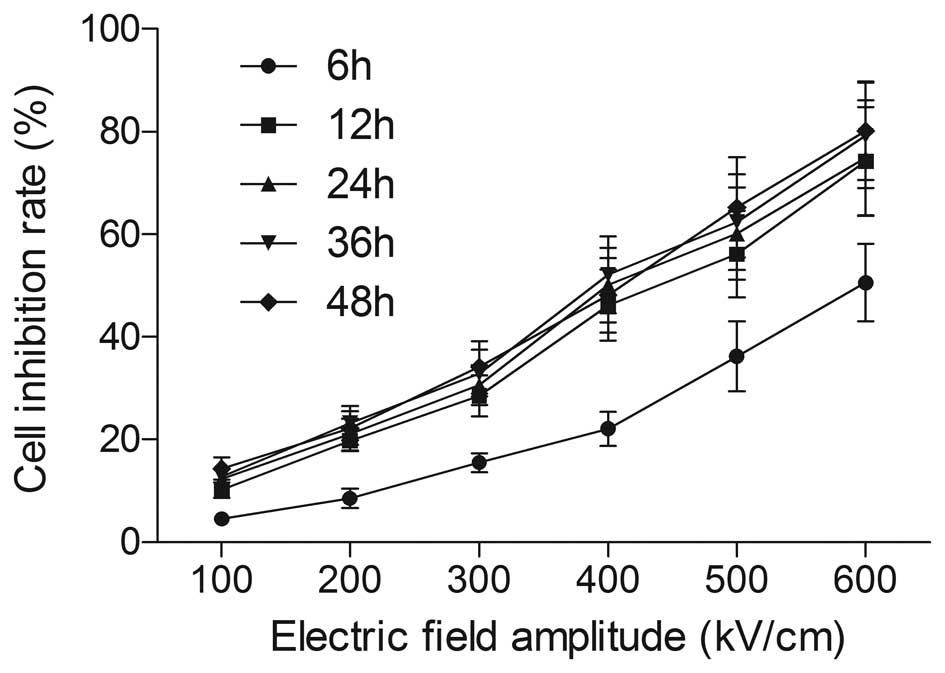 | Figure 1Effect of different culture times,
following picosecond pulsed electric field (psPEF) treatment with
different electric field amplitudes, on the growth inhibition of
HeLa cells, measured by MTT assay. Cells were treated with psPEF
with different amplitudes (100, 200, 300, 400, 500 and 600 kV/cm)
and cultured for different durations (6, 12, 24, 36 and 48 h)
following pulse treatment, respectively. Cell growth inhibition was
determined by MTT assay as described in Materials and methods. Data
are presented as the mean ± standard deviation of three separate
experiments. |
Effects of different picosecond
electrical pulse conditions on HeLa cell proliferation
As demonstrated in the results of the MTT assay
(Fig. 2), the growth inhibition
rates of the groups with a pulse number of 2,000 (10.2±1.50,
19.01±2.85, 28.56±3.68, 46.2±5.73, 56.12±9.19 and 71.23±4.48%) were
elevated in comparison with the groups with a pulse number of 1,000
(6.23±0.93, 13.2±1.98, 20.02±3.00, 36.51±5.25, 48.02±5.32 and
60.12±5.23%) (P<0.05). Results were recorded at electric field
amplitudes of 100–600 kV/cm, respectively, at 100-kV/cm intervals.
At a pulse number of 3,000, the growth inhibition rates in each
amplitude group were 13.5±2.03, 25.64±3.77, 38.84±5.30, 54.35±8.45,
70.24±12.34 and 83.4±10.723%, respectively. These rates were
significantly higher than the results in the groups with a pulse
number of 2,000 (P<0.05). The differences between the growth
inhibition rates of the groups with pulse numbers of 1,000 and
2,000 were particularly notable at amplitudes of 500 and 600 kV/cm
(P<0.01). Similarly, in each pulse number group (1,000, 2,000
and 3,000), the cell growth inhibition rate increased significantly
with increasing amplitude (P<0.05). The differences in the
growth inhibition rates were particularly of note between the 200
kV/cm groups (13.23±1.98, 19.01±2.85 and 25.64±3.77%) and the 100
kV groups (6.23±0.93, 10.2±1.50, and 13.50±2.03%; P<0.01). When
the electric field amplitude was 400 kV/cm, the cell growth
inhibition rates increased to 36.51±5.25, 46.2±5.73 and
59.31±8.45%, which were significantly higher than those in the 300
kV/cm groups (20.02±3.005, 28.56±3.685 and 38.84±5.305%,
P<0.01). The differences between the 500 and 600 kV/cm groups
were the same as the differences between the 300 and 400 kV/cm
groups. In the subsequent tests, 200, 400 and 600 kV/cm were
selected as the standard parameters of low-, middle- and high
amplitudes.
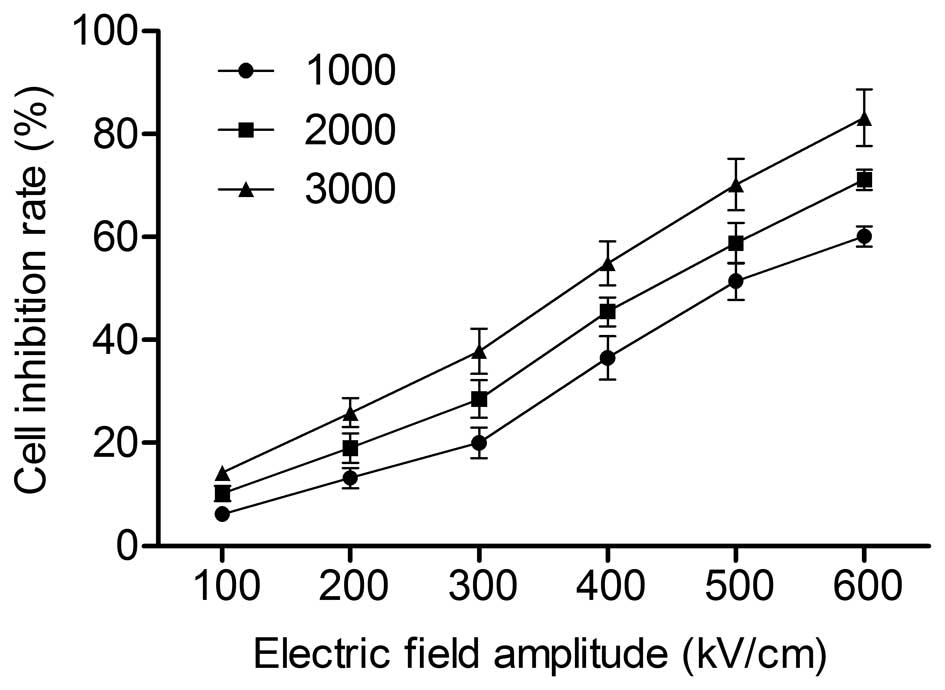 | Figure 2Effect of different electrical pulse
conditions on the viability of HeLa cells, measured by MTT assay.
Cells were treated with different electric field amplitudes (100,
200, 300, 400, 500 and 600 kV/cm), and then treated with different
pulse numbers (1,000, 2,000 and 3,000) in each electric field
amplitude group. Cell growth inhibition was determined by MTT
assay. Data are presented as the mean ± standard deviation of three
separate experiments. |
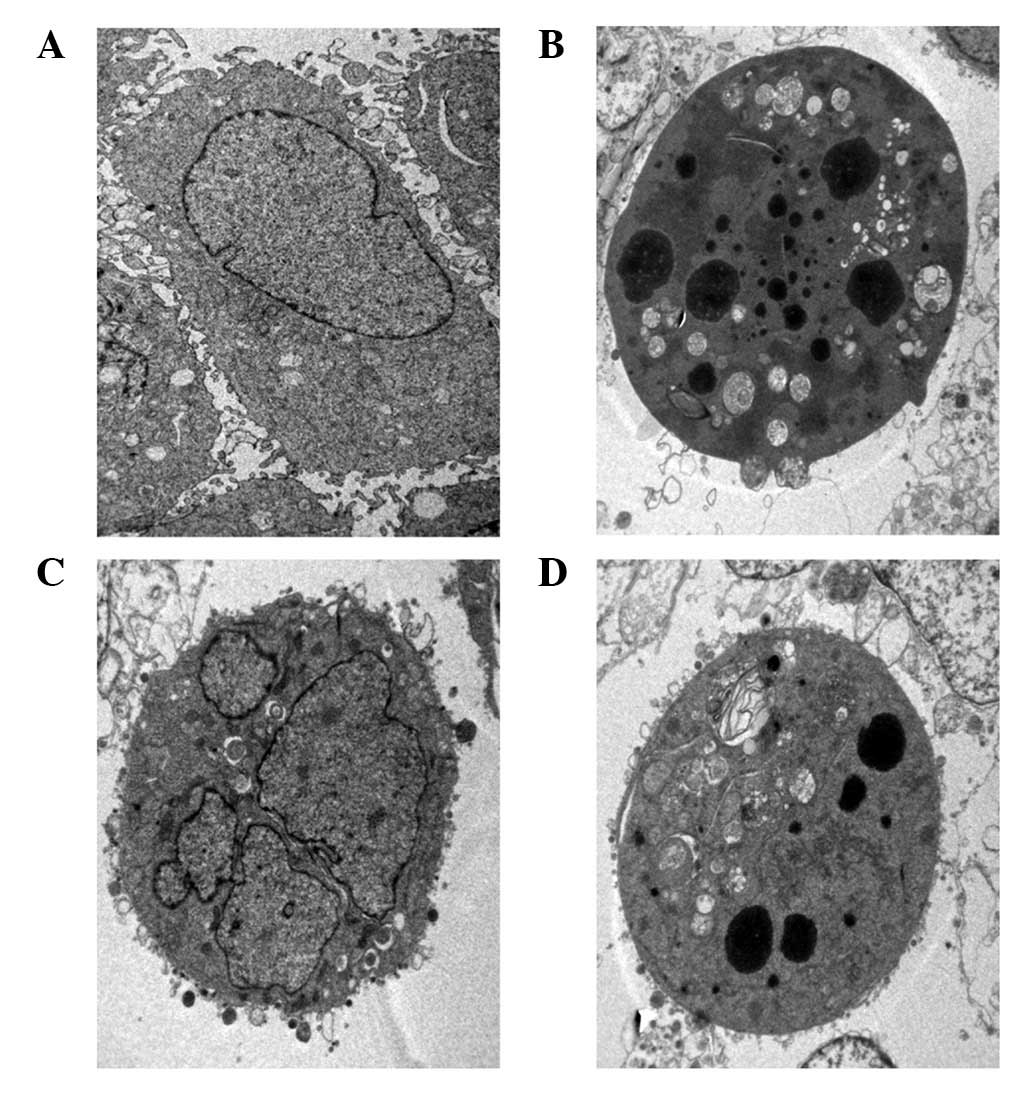
Ultrastructural observation
TEM was used to observe the ultrastructural changes
in the HeLa cells exposed to different electric field amplitudes.
As demonstrated in Fig. 3, the
normal HeLa cells were intact, with well-distributed chromatin and
a clear nuclear membrane (Fig.
3A). However, in response to psPEF exposure at electric field
amplitudes of 200, 400 and 600 kV/cm, the cells became reduced in
size and pyknotic, with an intact membrane, aggregated chromatin
and pseudopodia-like protrusions (Figs. 3B, C and D).
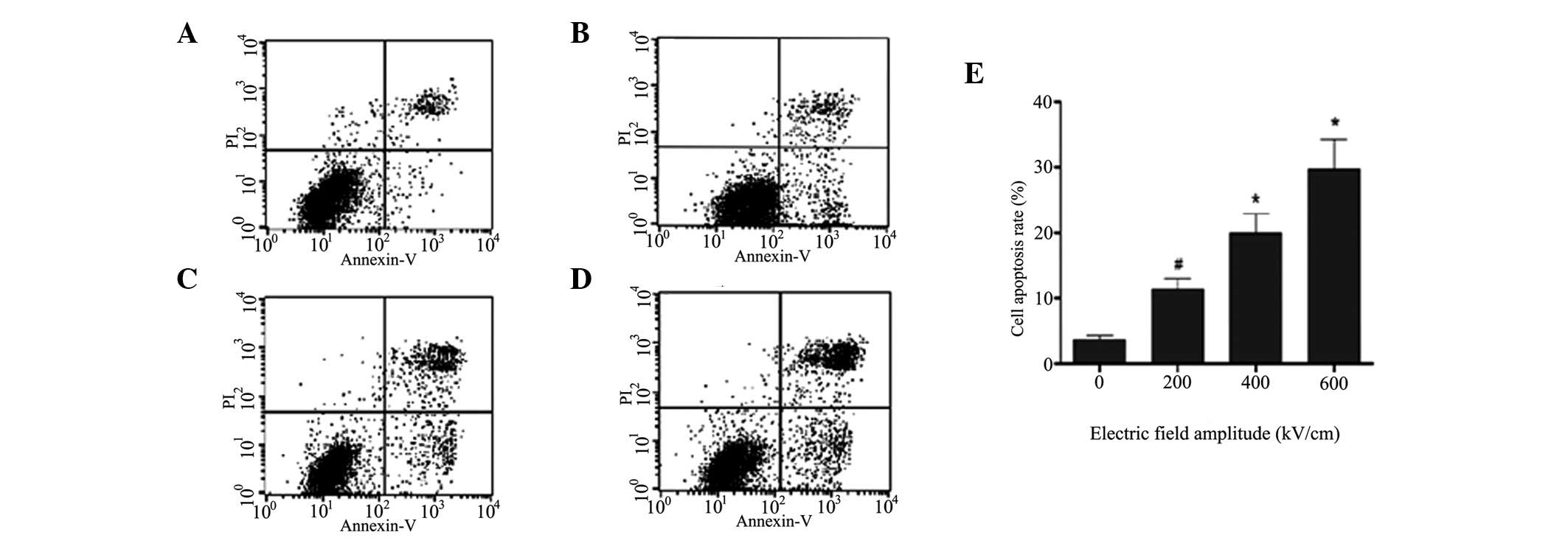
Cell apoptosis investigated by flow
cytometric analysis
With the aim of assessing whether apoptosis was
associated with the inhibition of proliferation induced by psPEF in
HeLa cells, cell apoptosis was detected by double staining the
cells with Annexin V and PI, following the electrical pulse
treatment and flow cytometric analysis. As is evident in Fig. 4, the rates of apoptosis in the
groups with pulse numbers of 1,000, 2,000 and 3,000 were
significantly higher (P<0.05) than those in the the untreated
group. The rate of apoptosis in the group with a pulse number of
1,000 (7.55±1.13%) was significantly higher (P<0.01) than that
in the control group (2.35±0.35%). The difference between the
groups with pulse numbers of 1,000 and 2,000 (14.75±2.21%) was also
notable (P<0.01). When the pulse number was increased to 3,000,
cell apoptosis increased to 19.94±2.29%, which was significantly
higher (P<0.05) than that in the group with a pulse number of
2,000.
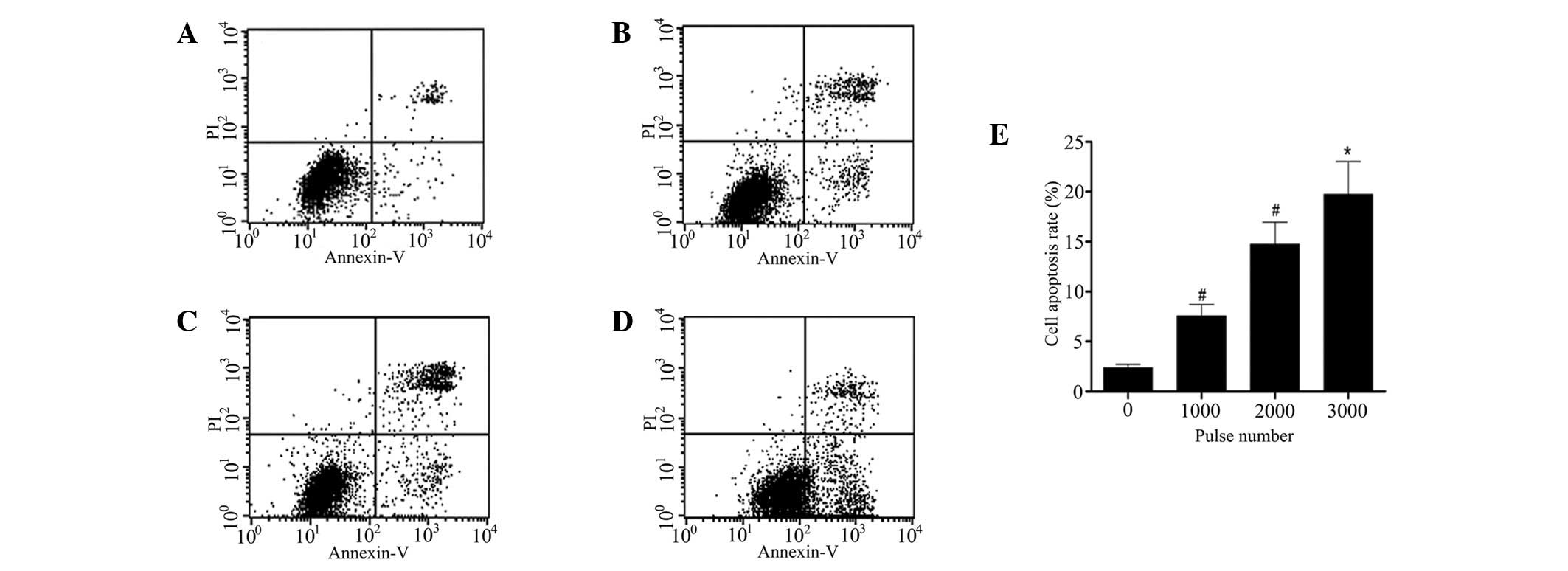 | Figure 4Apoptosis rate of HeLa cells under
different pulse numbers, tested by flow cytometry. Cells were
treated with different pulse numbers (1,000, 2,000 and 3,000),
harvested and double stained with Annexin V and propidium iodide
(PI), and then analyzed by flow cytometry. (A) Untreated group;
groups with pulse numbers of (B) 1,000, (C) 2,000 and (D) 3,000;
and (E) histogram plotted according to the data in A-D. Data are
presented as the mean ± standard deviation of three separate
experiments. *P<0.05 and #P<0.01;
comparison between two neighboring groups. |
As shown in Fig. 5,
the apoptosis rates of the 200, 400 and 600 kV/cm groups were
elevated significantly (P<0.05), in comparison with the control
group. The apoptosis rate in the 200 kV/cm group (11.43±1.60%) was
significantly higher (P<0.01) than that in the untreated group
(3.71±0.56%). The difference between the 200 and 400 kV/cm groups
(19.94±2.99%) was also significant (P<0.05). When the electric
field amplitude was increased to 600 kV/cm, the cell apoptosis rate
reached 29.77±4.47%, which was a notably higher result (P<0.05)
than that in the 400 kV/cm group.
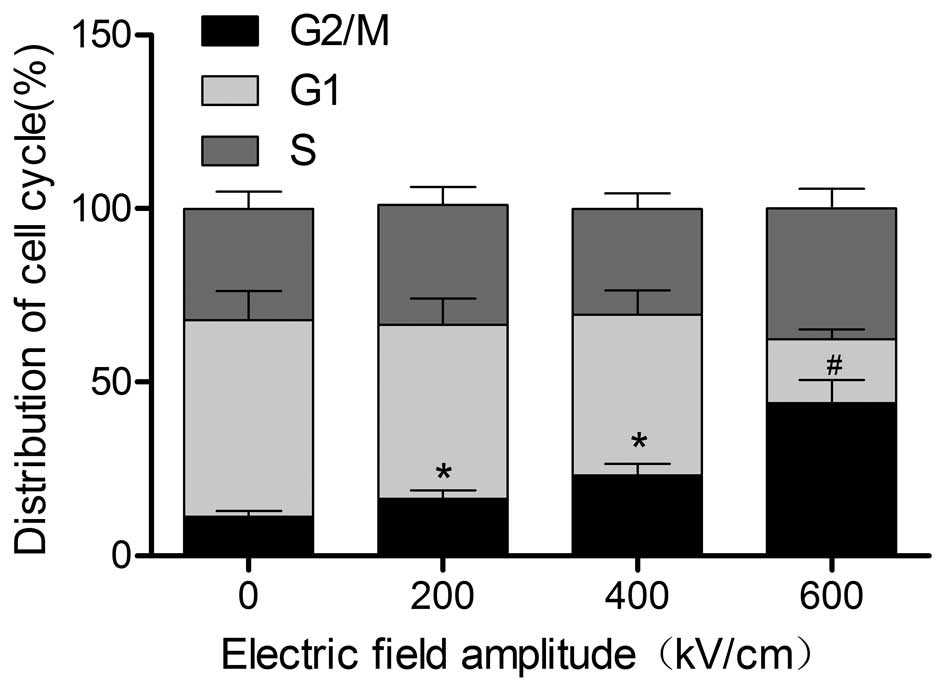
Effects of different electric field
amplitudes on the cell cycle of HeLa cells
To determine the role of psPEF in the cell cycle
progression of HeLa cells, cell cycle distribution was assessed by
monitoring the intensity of PI fluorescence. As shown in Fig. 6, the ratios of G2/M phase of the
cell cycle in the three experimental groups were greater than those
in the untreated group (P<0.05). A significant difference was
also observed between the 200-kV/cm group (16.36±2.45%) and the
400-kV/cm group (23.33±3.03%, P<0.05). In addition, the
proportion of cells in the G2/M phase of cell cycle in the
600-kV/cm group (44.04±6.61%) was significantly greater than in the
400-kV/cm group (P<0.01). As the electric field amplitude
increased, the number of G2/M phase cells increased significantly,
while the number of cells in the G1 phase decreased gradually. This
indicated that the psPEF blocked the cell cycle of HeLa cells in
the G2/M phase.
Discussion
Cervical cancer is a serious threat to female health
and quality of life (22).
Although the late stages of cervical cancer development carry a
poor prognosis, the early stages may be effectively cured with
surgery. However, this results in a loss of fertility in young
patients. Therefore, a treatment capable of, not only curing
cervical cancer, but also preserving the fertility of young
patients is necessary. As a result of widespread conventional
screening, and the fact that the cervix may be fully exposed using
simple methods and instruments, a number of cervical cancer
patients may be diagnosed at an early stage of development,
providing a suitable time for treatment with PEF.
Numerous experiments have demonstrated that PEF
(6–17), from msPEF to nsPEF, is capable of
producing corresponding biological effects on cells. These effects
have been validified by clinical experiments, although limitations
for the clinical application of PEF continue to exist, as mentioned
previously. It has been suggested that cell responses to different
types of PEF vary due to their different spectral distributions.
For example, msPEF has a long pulse duration, and therefore a lower
equivalent frequency; thus, it predominantly affects cell
membranes, while intracellular organelles are shielded. Its
influence on the cell membrane may result in the formation of
electroporation. However, the frequency of nsPEF or PEF with a
shorter pulse duration is higher, which facilitates penetration of
the cell membrane. This enables the nsPEF to have an impact on
intracellular organelles, such as mitochondria and endoplasmic
reticulum, and to induce electroporation in intracellular
membranes, as well as intracellular electromanipulation (23,24).
Other studies have demonstrated that when pulse duration was taken
as a reference point, the biological medical effects elicited
differed in accordance with the changing parameters of PEF
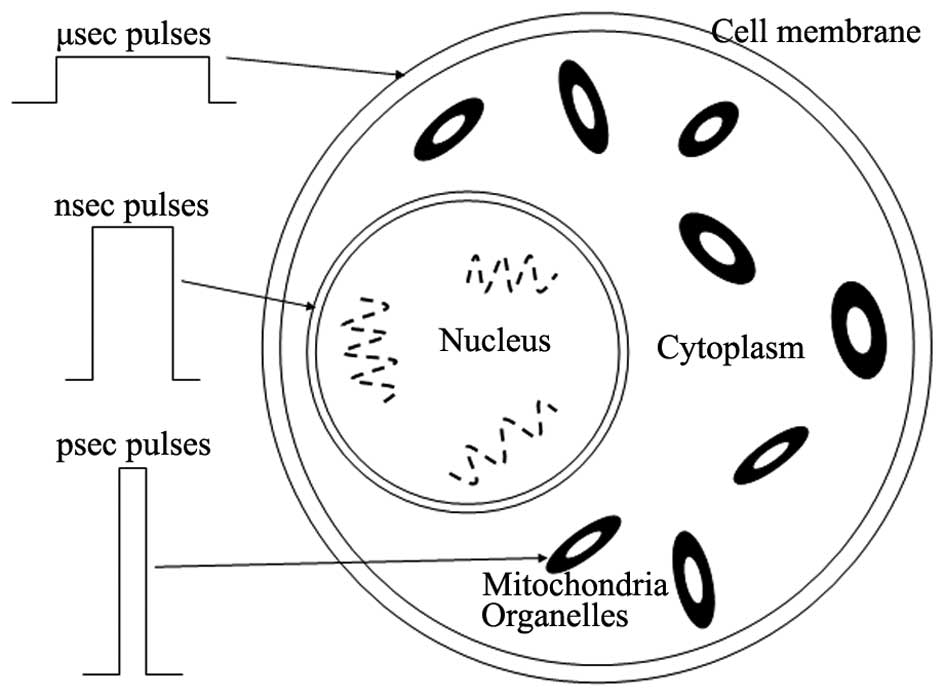
(25,26). As Fig.
7 shows, when the pulse duration reduces from a millisecond to
a microsecond, and further to a nanosecond, the target of the PEF
transfers from the cell membrane (the intracellular membrane) to
the nucleus and the cytoplasm (the cytosol). A further decrease in
the pulse duration, to a subnanosecond or a picosecond, changes the
target action point to the nucleus and organelles, such as the
mitochondria, thereby killing the cells directly. Therefore, this
study focused on psPEF, with the aim of providing a foundation for
the development of a non-invasive, safe and effective
treatment.
In a previous study, platelets were exposed to
150-psec pulses with an electric field amplitude of 150 kV/cm. The
results indicated that the uptake of calcium was in a pulse
number-dependent manner. Further investigation demonstrated that
when B16 melanoma cells were treated with 800-psec pulses with an
electric field amplitude of 950 kV/cm, phosphatidylserine valgus
was exhibited and caspase-3 was activated. When the electric field
amplitudes of 800-psec pulses were 550 and 490 kV/cm, trypan blue
staining and the determination of caspase activity indicated that
there was an interrelation between the dose (pulse number) and the
effect elicited (27–29). The preliminary study of this
experimental team indicated that psPEF, with an electric field
amplitude of 250 kV/cm and various pulse numbers (0–50,000),
induced apoptosis in cultured human cervical cancer cells, and that
the apoptotic effect was possibly through the
mitochondrial-mediated pathway (30).
At present, the pulse generator at Chongqing
University is capable of generating a maximum electric field
amplitude of 600 kV/cm. Thus, the aim of this study was to
establish the effects of psPEF with a range of electric field
amplitudes (100–600 kV/cm) and a range of pulse numbers
(1,000–3,000), and to further observe the dose-effect correlation.
Initially, investigations were undertaken to determine the culture
duration required, following the pulse treatment, for psPEF to
achieve a plateau of maximum cell inhibition under different
electric field amplitudes. The first MTT assay result demonstrated
that at a given electric field amplitude, electrical pulses
achieved a plateau of maximum cell inhibition 12 h following the
pulse treatment. Based on the experimental results, an MTT assay
was then used to test whether psPEF was capable of inhibiting cell
proliferation. The results demonstrated that psPEF had an
inhibitory effect on the proliferation of the HeLa cell line in a
dose (electric field amplitude and pulse number)-dependent manner.
As the amplitude increased, cell growth inhibition also increased
significantly, particularly at electric field amplitudes of 200,
400 and 600 kV/cm, and at a pulse number of 2,000. At these
specifications, the cell growth inhibition rates were markedly
higher than those in the lower electric field amplitudes or pulse
number groups. From the results of our preliminary experiments,
200, 400 and 600 kV/cm were chosen as the standard values for low-,
middle- and high amplitudes, respectively. To investigate the
biological effect induced by psPEF at these parameters of electric
field amplitude, and at three typical parameters of pulse number
(1,000, 2,000 and 3,000), the flow cytometric analysis was used to
determine the cell apoptosis level of each group. Under different
psPEF conditions, the apoptosis of HeLa cells varied. It was
apparent that the apoptosis rates in the 200-, 400- and 600-kV/cm
groups were significantly higher than the rate in the untreated
group. In the treatment groups, the apoptosis rate in the higher
amplitude group was greater than that in the lower amplitude group.
Similar results were obtained when the variable was pulse number.
In addition, typical characteristics of apoptosis in the HeLa cells
were observed under TEM. This indicated that intense psPEF induced
apoptosis of the HeLa cells significantly in a dose-dependent
manner. Flow cytometric evaluation of the cell cycle demonstrated
that psPEF with electric field amplitudes of 200, 400 and 600 kV/cm
affected the cell cycle, thereby inhibiting tumor cell growth.
This study was limited to experiments in
vitro, although the next progression will be to examine
experiments in vivo. This technology is currently in its
infancy in gynecological diseases, and further investigation is
required.
Acknowledgements
This study was supported by two grants from the
National Natural Science Foundation of China (project no. 81172123)
and the Health Bureau of Chongqing (project no. 2012-2-068). The
authors would like to thank the members of their laboratory for
their support.
References
|
1
|
Bray F, Loos AH, McCarron P, et al: Trends
in cervical squamous cell carcinoma incidence in 13 European
countries: changing risk and the effects of screening. Cancer
Epidemiol Biomarkers Prev. 14:677–686. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pecorelli S, Pasinetti B, Angioli R,
Favalli G and Odicino F: Systemic therapy for gynecological
neoplasms: ovary, cervix, and endometrium. Cancer Chemother Biol
Response Modif. 22:515–544. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Green J, Kirwan J, Tierney J, et al:
Concomitant chemotherapy and radiation therapy for cancer of the
uterine cervix. Cochrane Database Syst Rev. 3:CD0022252005.
|
|
4
|
Barnes FS and Greenebaum B: Biological and
medical aspects of electromagnetic fields. Handbook of Biological
Effects of Electromagnetic Fields. 3rd edition. CRC Press; Florida:
2006
|
|
5
|
Joshi RP and Schoenbach KH: Bioelectric
effects of intense ultrashort pulses. Crit Rev Biomed Eng.
38:255–304. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weaver JC: Electroporation: a general
phenomenon for manipulating cells and tissues. J Cell Biochem.
51:426–435. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mir LM: Nucleic acids
electrotransfer-based gene therapy (electrogenetherapy): past,
current and future. Mol Biotechnol. 43:167–176. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okino M, Tomie H, Kanesada H, Marumoto M,
Esato K and Suzuki H: Optimal electric conditions in electrical
impulse chemotherapy. Jpn J Cancer Res. 83:1095–1101. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hofmann GA, Dev SB, Dimmer S and Nanda GS:
Electroporation therapy: a new approach for the treatment of head
and neck cancer. IEEE Trans Biomed Eng. 46:752–759. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dev SB, Rabussay DP, Widera G and Hofmann
GA: Medical applications of electroporation. IEEE Trans Plasma Sci.
28:206–223. 2000. View Article : Google Scholar
|
|
11
|
Tien HT and Ottova A: The bilayer lipid
membrane (BLM) under electrical fields. IEEE Trans Dielectr Electr
Insul. 10:717–727. 2003. View Article : Google Scholar
|
|
12
|
Lee EW, Chen C, Prieto VE, Dry SM, Loh CT
and Kee ST: Advanced hepatic ablation technique for creating
complete cell death: irreversible electroporation. Radiology.
255:426–433. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou W, Xiong Z, Liu Y, Yao C and Li C:
Low voltage irreversible electroporation induced apoptosis in HeLa
cells. J Cancer Res Ther. 8:80–85. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stacey M, Stickley J, Fox P, Statler V,
Schoenbach K, Beebe SJ and Buescher S: Differential effects in
cells exposed to ultra-short, high intensity electric fields: cell
survival, DNA damage, and cell cycle analysis. Mutat Res.
542:65–75. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katsuki S, Nomura N, Koga H, et al:
Biological effects of narrow band pulsed electric fields. IEEE
Trans Dielectr Electr Insul. 14:663–668. 2007. View Article : Google Scholar
|
|
16
|
Chen N, Garner AL, Chen G, et al:
Nanosecond electric pulses penetrate the nucleus and enhance
speckle formation. Biochem Biophys Res Commun. 364:220–225. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Craviso GL, Chatterjee P, Maalouf G, et
al: Nanosecond electric pulse-induced increase in intracellular
calcium in adrenal chromaffin cells triggers calcium-dependent
catecholamine release. IEEE Trans Dielectr Electr Insul.
16:1294–1301. 2009. View Article : Google Scholar
|
|
18
|
Baum CE, Stone AP and Tyo JS:
Ultra-Wideband, Short-Pulse Electromagnetics. 8. Springer Press;
New York: 2007
|
|
19
|
Bajracharya C, Shu X, Baum CE and
Schoenbach KH: Target detection with impulse radiating antenna.
IEEE Antennas Wireless Propag Lett. 10:496–499. 2011. View Article : Google Scholar
|
|
20
|
Long Z, Yao C, Li C, Mi Y and Sun C:
Focusing properties of picosecond electric pulses in non-invasive
cancer treatment. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi.
27:1128–1132. 2010.(In Chinese).
|
|
21
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics. CA Cancer J Clin. 59:225–249. 2009.
|
|
23
|
Beebe SJ and Schoenbach KH: Nanosecond
pulsed electric fields: a new stimulus to activate intracellular
signaling. J Biomed Biotechnol. 2005:297–300. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schoenbach KH, Joshi RP, Kolb JF, et al:
Ultrashort electrical pulses open a new gateway into biological
cells. Proc IEEE. 92:1122–1137. 2004. View Article : Google Scholar
|
|
25
|
Yao C, Mo D, Li C, Sun C and Mi Y: Study
of transmembrane potentials of inner and outer membranes induced by
pulsed-electric-field model and simulation. IEEE Trans Plasma Sci.
35:1541–1549. 2007. View Article : Google Scholar
|
|
26
|
Yao C, Mi Y, Li C, et al: Study of
transmembrane potentials on cellular inner and outer membrane -
frequency response model and its filter characteristic simulation.
IEEE Trans Biomed Eng. 55:1792–1799. 2008. View Article : Google Scholar
|
|
27
|
Camp JT, Shu X, Beebe SJ, Blackmore PF and
Schoenbach KH: Bioelectric studies with subnanosecond pulsed
electric fields. In: 2009 IEEE Pulsed Power Conference; June
28-July 2; pp. 876–879. 2009
|
|
28
|
Schoenbach KH, Shu X, Joshi RP, Camp JT,
Heeren T, Kolb JF and Beebe SJ: The effect of intense subnanosecond
electrical pulses on biological cells. IEEE Trans Plasma Sci.
36:414–422. 2008. View Article : Google Scholar
|
|
29
|
Schoenbach KH, Katsuki S, Akiyama H, et
al: Biological effects of intense subnanosecond electrical pulses.
In: Proceedings of the Power Modulator Symposium, 2006. Conference
Record of the 2006 Twenty-Seventh International; May 14–18; pp.
573–576. 2006
|
|
30
|
Hua YY, Wang XS, Zhang Y, Yao CG, Zhang XM
and Xiong ZA: Intense picosecond pulsed electric fields induce
apoptosis through a mitochondrial-mediated pathway in HeLa cells.
Mol Med Rep. 5:981–987. 2012.PubMed/NCBI
|