Introduction
Leukemia accounts for the largest number of cases of
childhood cancer (1). The
predominant therapeutic approaches for human leukemia are
radiotherapy, hyperthermia and chemotherapy; however, cure rates
remain unsatisfactory (2). Hence,
effective drugs, including novel naturally occurring or chemically
synthesized compounds, are required to control the malignant
progression of leukemia.
Pseudolaric acid B (PAB), a natural diterpene acid
present in the traditional Chinese medicinal herb Tu-Jin-Pi, has
exerted anticancer effects on various types of cancer cells
(3). It was demonstrated that PAB
significantly delayed the growth of a taxol-resistant liver cancer
without obvious toxicity in vivo and may possess a selective
anti-proliferative effect in human cancer cells but not in normal
cells in vitro(1). It was
also demonstrated that PAB induced apoptosis via
proteasome-mediated Bcl-2 degradation in hormone-refractory DU145
prostate cancer cells (3).
Although PAB exhibits cytotoxic activity and induces apoptosis in
several cancer cell types, its action in leukemia is unclear.
Apoptosis is a selective process of physiological
cell deletion that regulates the balance between cell proliferation
and cell death (4). The failure of
apoptosis is considered to contribute to the development of human
malignancies (5). Therefore,
targeting key apoptotic regulators has become a strategy for the
development of novel treatments for human cancer (6,7).
Changes in the apoptotic signaling pathway
contribute to the development of multiple human diseases, including
cancer, autoimmunity, diabetes and neurodegenerative disorders, and
may provide rational targets for novel anticancer drugs (8). Apoptosis occurs via the mitochondria-
and death receptor-mediated pathways. The two apoptotic pathways
converge at caspase-3 and subsequently at other proteases and
nucleases that drive the terminal events of apoptosis (9). The mitochondrial pathway is termed
the intrinsic pathway of apoptosis (10). Mitochondria are the predominant
energy producers in the cell and thus are essential for maintaining
cell survival. Moreover, mitochondria are also involved in cell
death and contain the majority of the important mediators of
apoptosis (11). However, the
involvement of the mitochondrial pathway in the anti-leukemia
effects of PAB remains to be fully elucidated. Therefore, the
principal aim of this study was to investigate the mechanisms
underlying PAB-induced apoptosis in U937 human leukemia cells by
determining the involvement of the mitochondrial pathway. In the
present study, it was demonstrated that PAB activated a
caspase-dependent apoptotic pathway in U937 cells through the
regulation of the Bcl-2 family protein-mediated mitochondrial
pathway. Moreover, the initiation of caspase-3 and -9 activation
mediated apoptotic induction. To the best of our knowledge, this is
the first study to investigate mitochondrial signaling in U937
cells in response to PAB treatment.
Materials and methods
Reagents
PAB was purchased from the Liaoning Institute for
Food and Drug Control (Shenyang, Liaoning, China). Monoclonal
anti-β-actin antibodies were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). Anti-cytochrome
c, apoptosis inducing factor (AIF), anti-Bcl-2, -Mcl-1,
-tBid, -Noxa, -Bax and -Bcl-xl were purchased from New England
Biolabs (Santa Cruz, CA, USA). RPMI-1640 medium and fetal bovine
serum were purchased from Gibco-BRL (Grand Island, NY, USA). An
Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kit
was purchased from R&D Systems (Abingdon, UK). Cell isolation
and tissue culture reagents were obtained from Invitrogen Life
Technologies (Lindingö, Sweden). All other reagents were obtained
from Sigma-Aldrich (St. Louis, MO, USA).
U937 cell culture
U937 cells, obtained from Sun Yat-sen University
(Guangzhou, China), were used in all experiments. The cells were
cultured in RPMI-1640 medium supplemented with 10% fetal calf
serum, 2 mM L-glutamine, 100 U/ml penicillin and 100 g/ml
streptomycin, and incubated at 37ºC in humidified air containing 5%
CO2. The exponentially growing U937 cells were seeded
onto 24-well flat-bottomed plates at a density of 5×105
cells/ml. The cells were collected at 0, 24, 48, 72 and 96 h. Prior
to the addition of PAB, the cells were washed once with
phosphate-buffered saline (PBS) to remove dead cells, and incubated
in tissue-culture plates for 30 min at 37ºC. U937 cells in
multiwell tissue culture plates were incubated with PAB at
different concentrations (0, 1, 2, 4 and 8 μmol/l) for 48 h or at a
concentration of 2 μmol/l for 0, 24, 48, 72 and 96 h.
Intracellular reactive oxygen species
(ROS) detection
Intracellular production of ROS was measured by the
Reactive Oxygen Species Assay kit (Enzo Life Sciences, Shanghai,
China), which determines the oxidation of
2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) (12). Following treatment with PAB (2
μmol/l) for 48 h, the U937 cells were washed three times with
serum-free RPMI-1640 and incubated with 10 μM DCFH-DA in the dark
for 20 min at 37ºC. Cells were then washed twice with cold PBS. The
qualitative analysis of ROS generation was conducted using a
fluorescence microscope (Carl Zeiss, Hamburg, Germany). The
fluorescence was measured in a Tecan Infinite 200 plate reader
(Eastwin Life Sciences, Inc., Haidian, China) with excitation at
485 nm and emission at 520 nm. Values were expressed as the mean
absorbance normalized to the percentage of the vehicle control.
Cell viability
To assess the overall viability of the U937 cells
following PAB treatment, the cells were treated as described
previously. At particular time points (0, 24, 48, 72 and 96 h), the
U937 cells were washed twice with PBS and treated with a 0.4%
solution of trypan blue. Viable cells were visualized as clear
under the microscope; however, U937 cells that were no longer
viable exhibited damaged membranes and thus allowed entry of the
dye, staining blue. Assays were performed in triplicate and
repeated at least three times. The number of intact viable cells
was expressed as a percentage of the total number of cells and was
assessed at different time points.
Hoechst 33258 staining
The cells were stained with Hoechst 33258 (Molecular
Probes, Inc., Eugene, OR, USA) in a dilution of 1:600 (stock
solution, 1 mg/ml) for 5 min in the dark. The samples were observed
under a fluorescence microscope. Two-hundred cells were counted
from each coverslip and the results were confirmed by visualizing
the apoptotic nuclei. There were five coverslips per group, unless
otherwise noted in the figure legends.
Flow cytometry analysis
U937 cell apoptosis was quantified by flow cytometry
using FITC-conjugated Annexin V and propidium iodide (PI). Specific
binding of Annexin V was achieved by incubating 106
cells in 60 μl binding buffer saturated with Annexin V for 15 min
at 4ºC in the dark. To discriminate between early apoptosis and
necrosis, the cells were simultaneously stained with Annexin V and
PI prior to analysis. The binding of Annexin V-FITC and PI to the
cells was measured by flow cytometry (FACSCalibur, BD Biosciences,
Franklin Lakes, NJ, USA) using CellQuest software (BD Biosciences).
A minimum of 10,000 cells were counted in each sample. Experiments
were performed and interpreted as follows: Annexin
V−/PI− cells (lower left quadrant) were
considered to be living cells; Annexin V+/PI−
cells (lower right quadrant) were considered to be apoptotic cells;
Annexin V+/PI+ cells (upper right quadrant)
were considered to be necrotic or advanced apoptotic cells; and
Annexin V−/PI+ staining (upper left quadrant)
was considered to indicate bare nuclei, cells in late necrosis or
cellular debris.
Measurement of cytochrome c release from
mitochondria
Cells were treated with 0.1% dimethylsulfoxide
(DMSO) for 24 h. The mitochondria and the cytosol were separated
using a Cytochrome c-Releasing Apoptosis Assay kit (Enzo
Life Sciences). Cells were suspended in cytosol extraction buffer
and, following 10 min on ice, were homogenized using a Dounce
homogenizer (GlobalSpec, East Greenbush, NY, USA) and centrifuged
(at 700 × g for 10 min). Subsequent to this, the collected
supernatant was re-centrifuged (at 10,000 × g and 4ºC for 30 min).
The resulting supernatant (cytosolic fraction) and the pellet
(mitochondrial fraction) were processed for western blot analysis
(13).
Western blot analysis
Briefly, following treatment, cells were washed once
with ice-cold PBS containing 1 mM Na2VO4 and
extracted with lysis buffer (50 mM Tris, pH 8.0; 150 mM NaCl; 5 mM
EDTA; 5% glycerol; 1% Triton X-100; 25 mM NaF; 2 mM
Na2VO4; and 10 μg/ml aprotinin, leupeptin and
pepstatin). The preparation of the cytoplasm was conducted using
the NE-PER Nuclear and Cytoplasmic Extraction reagents (Pierce
Biotechnology, Inc., Rockford, IL, USA). The cell lysates were
frozen and thawed three times, and were further centrifuged at
14,000 × g for 10 min at 4ºC to pellet insoluble material. The
supernatant of the cell extracts was analyzed for the concentration
of proteins by a DC Protein Assay kit based on the Lowry method
(Bio-Rad, Hercules, CA, USA). Equal quantities of protein (50 μg)
from each sample were separated on 10% sodium dodecyl
sulfate-polyacrylamide gels and transferred onto polyvinylidene
fluoride membranes (Micron Separations, Inc., Westborough, MA,
USA). Membranes were blocked in 5% non-fat dry milk in
Tris-buffered saline containing 0.05% Tween-20 (TBST), and then
incubated with rabbit anti-human cytochrome c, AIF,
anti-Bcl-2, Mcl-1, Bcl-xl, Noxa, Bax and tBid primary antibodies
for 60 min. β-actin (dilution, 1:2,000) was used as a control for
equal protein loading. The immunoblots were then washed three times
with TBST buffer, incubated with a horseradish
peroxidase-conjugated secondary antibody (goat anti-rabbit IgM;
Santa Cruz Biotechnology, Inc.) and developed using a
chemiluminescent substrate (Pierce Biotechnology, Inc.). To
quantify and compare levels of proteins, the density of each band
was measured with a densitometer (GE Healthcare, Little Chalfont,
UK).
Measurement of caspase-3 and -9
activity
U937 cells were harvested and centrifuged at 400 × g
for 10 min. Cells were washed twice with PBS (pH 7.4), resuspended
with 50 μl lysis buffer at 4ºC and incubated on ice for 10 min. All
subsequent steps were performed on ice. Following centrifugation,
cell extracts were transferred to fresh tubes, and protein
concentrations were measured. Each 50-μl cell extract containing
100 μg protein was combined with equal volumes of 2X reaction
buffer in a microplate followed by the addition of 5 μl peptide
substrates of caspase-3 and -9. Subsequent to overnight incubation
in the dark at 37ºC, samples were analyzed in a microplate reader
(Tecan Austria, Salzburg, Austria) at 405 nm. Caspase-3 and -9
activity was evaluated by the absorbance ratio of treated to
control samples. In certain experiments, the caspase-3 inhibitor
(Z-DEVD-FMK) and the caspase-9 inhibitor (Z-LEHD-FMK) were added to
the fresh medium of U937 cells 1 h prior to the addition of
PAB.
Statistical analysis
Each experiment was conducted in duplicate or
triplicate, and three or four independent experiments were
performed. Results are presented as the mean ± standard deviation
and analyzed with SPSS 11.5 software (SPSS, Inc., Chicago, IL,
USA). Results were compared using analysis of variance (ANOVA).
When the ANOVA showed a statistically significant difference, a
group-by-group comparison was performed using a t-test with Tukey’s
correction for multiple comparison. P<0.05 was considered to
indicate a statistically significant difference.
Results
PAB induces apoptosis in U937 cells
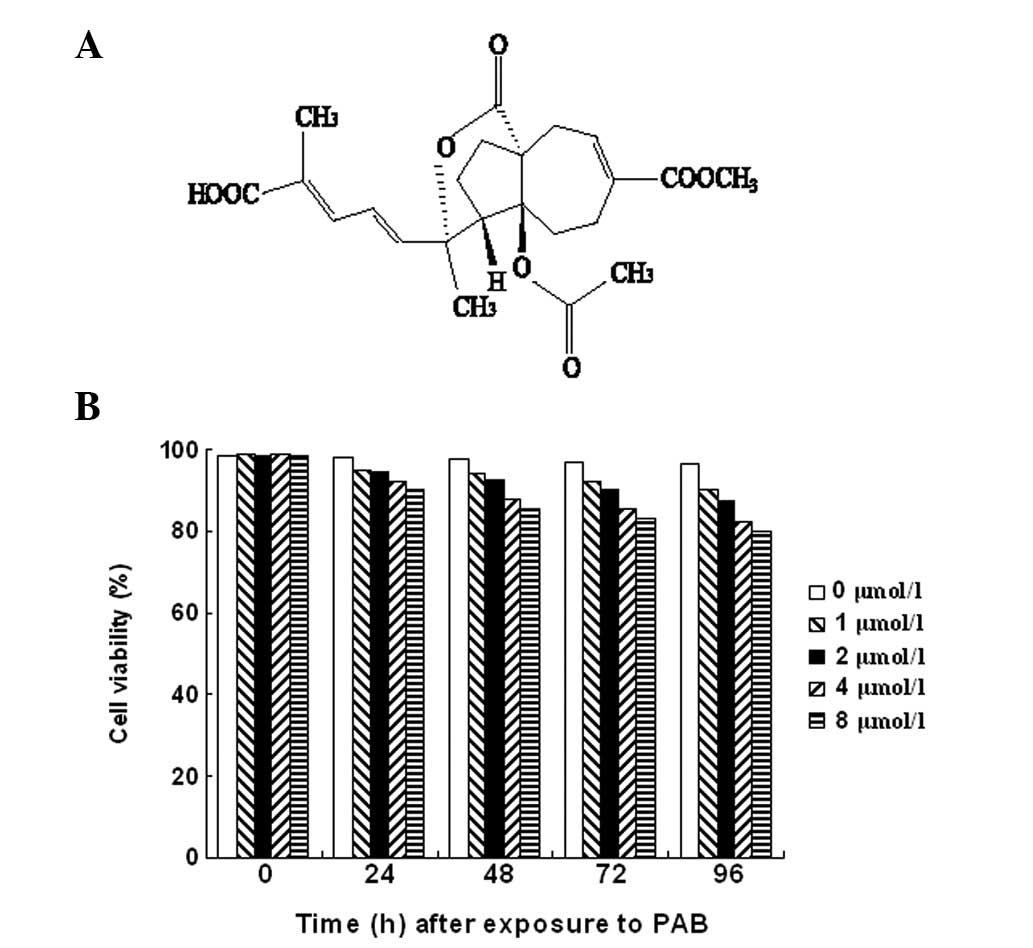
The chemical structure of PAB (14) is shown in Fig. 1A. In order to distinguish apoptosis
from necrosis in the U937 cells, a trypan blue exclusion assay was
conducted. Trypan blue staining demonstrated that 98.12±1.96% (n=5)
of the cells incubated with medium alone retained an integrated
cell membrane (i.e., resisted trypan blue staining). The percentage
of necrotic cells increased with time and with an increase in the
concentration of PAB (Fig.
1B).
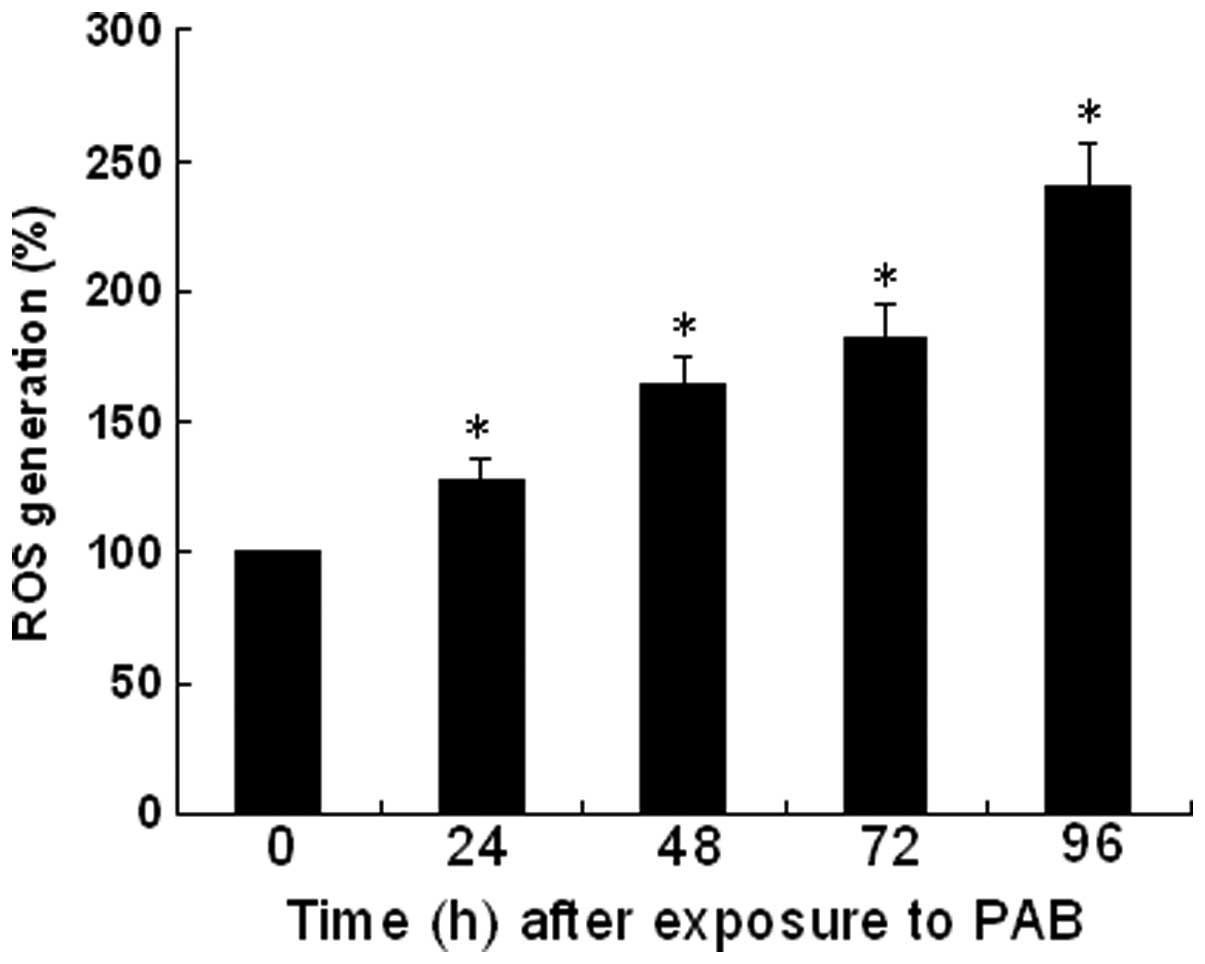
ROS is involved in PAB-induced apoptosis
of U937 cells
Intracellular ROS generation and mitochondrial
dysfunction are important in the signal transduction of apoptotic
stimuli (15). Therefore, the
fraction of cells with high intracellular ROS levels was
investigated following treatment with PAB. To determine the
mechanism involved in PAB-induced apoptosis, ROS production in the
U937 cells was investigated. The rate of ROS generation at 0 h was
considered to be 100%, and a qualitative and quantitative
concentration-dependent increase in the rate of ROS generation was
observed as increased levels of fluorescence following treatment
with PAB in U937 cells (Fig.
2).
PAB induces apoptosis in human U937
cells
Microscopy of PAB-induced U937 cells revealed
morphological changes that were consistent with the induction of
apoptosis and necrosis (Fig. 3A).
Apoptotic cells were characterized by membrane blebbing and nuclear
condensation, while necrotic cells were typically larger and
lighter with plasma membrane lesions and mitochondrial
abnormalities. The percentage of apoptotic cells was calculated
from the observation of 200 cells.
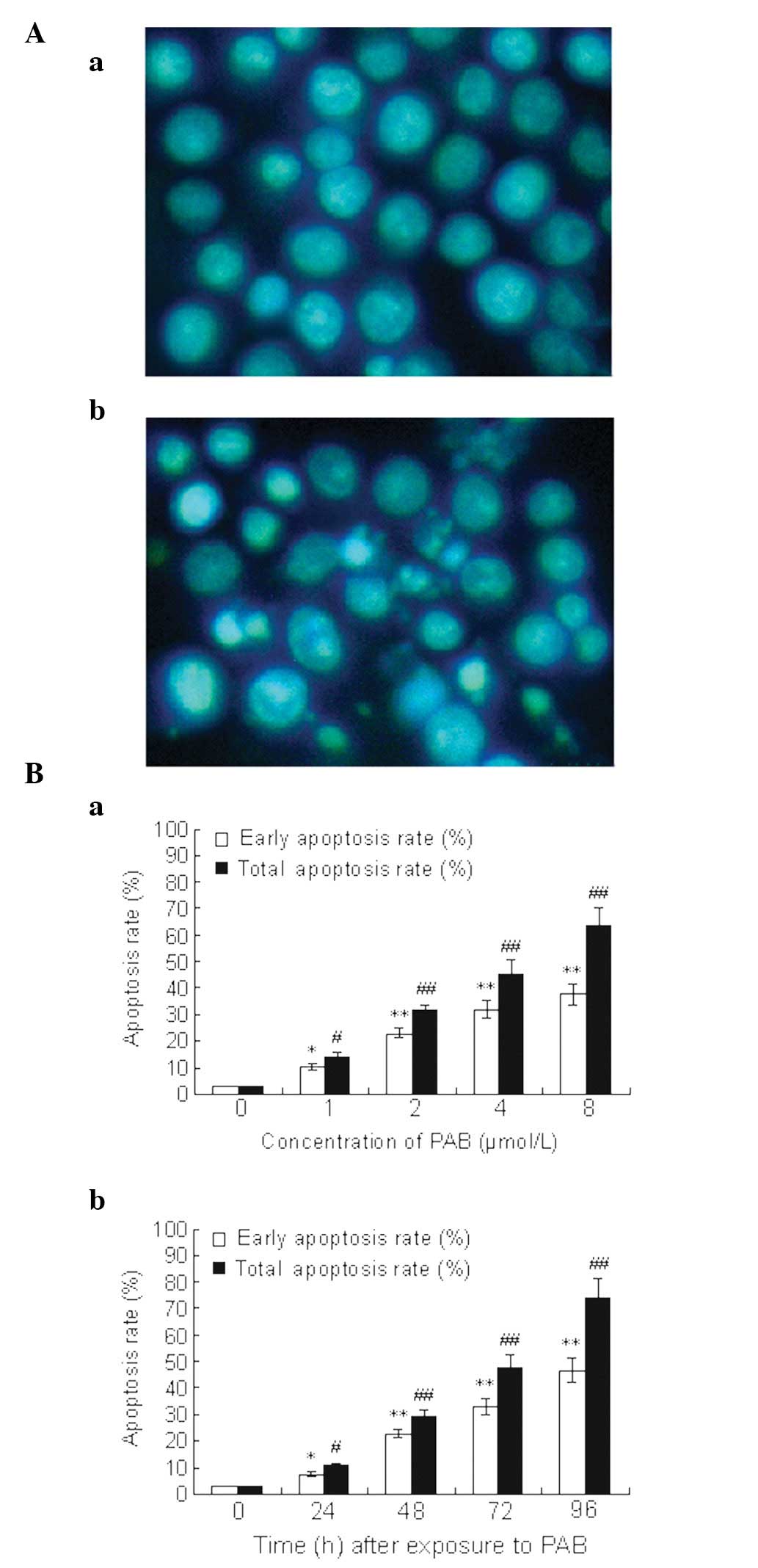 | Figure 3PAB-induced apoptosis in U937 cells.
(A) Morphology of U937 cells. U937 cells were cultured (Aa) with or
(Ab) without PAB. Control cells and cells treated with PAB were
stained with Hoechst 33258 dye (magnification, ×400). (B) Apoptosis
rate of U937 cells detected by flow cytometric analysis. U937 cells
were harvested (Ba) after 48 h of treatment with different
concentrations of PAB or (Bb) following different times of exposure
to 2 μmol/l PAB. The cells were then incubated with FITC-conjugated
Annexin V and PI double staining. Flow cytometric analysis was
performed. Data shown are representative of three separate
experiments. The lower right quadrants represent early apoptotic
cells that were stained by Annexin V but not by PI, and upper right
quadrants represent late apoptotic cells that were stained by
Annexin V and PI.’(Ba) *P<0.05,
#P<0.05, **P<0.001 and
##P<0.001, compared with cells treated with 0 μmol/l
PAB; (Bb) *P<0.05, #P<0.05,
**P<0.001 and ##P<0.001, compared with
cells treated with PAB at 0 h. PAB, pseudolaric acid B; FITC,
fluorescein isothiocyanate; PI, propidium iodide. |
Flow cytometry using FITC-conjugated Annexin V
demonstrated that U937 cells exposed to PAB underwent rapid
apoptosis. This effect was positively correlated with the exposure
time and the concentration of PAB (Fig. 3B). In addition, excessive apoptosis
was associated with the loss of membrane integrity in an increased
number of U937 cells, which indicated that necrosis or late
apoptosis had occurred.
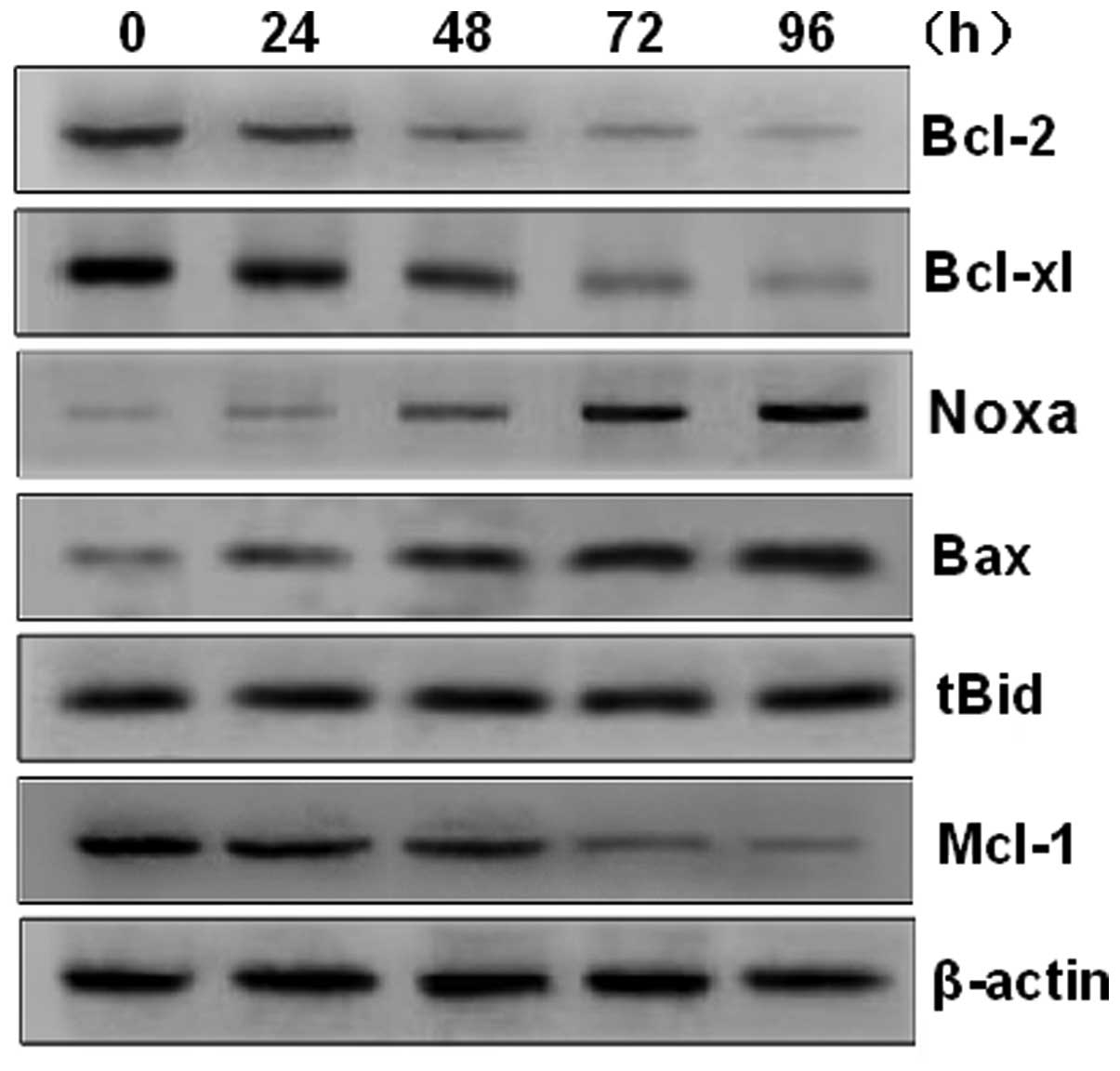
PAB induces apoptosis of U937 cells
through the Bax initiated mitochondrial pathway
Members of the Bcl-2 family are divided into
pro-apoptotic and anti-apoptotic proteins. The Bcl-2 protein family
is responsible for the release of cytochrome c from the
mitochondria, a pivotal event in the induction of apoptosis. Mcl-1,
a member of the anti-apoptotic Bcl-2 protein family, has been
demonstrated to be overexpressed in numerous types of cancer
(16). Notably, in the present
study, PAB significantly increased the expression of Bax and Noxa,
and decreased the expression of Mcl-1, Bcl-xl and Bcl-2 (Fig. 4). However, the expression of tBid
was not significantly affected by treatment with PAB. These results
suggest that the apoptotic effect may be mediated by raising the
ratio of Bax to Bcl-xL, to induce the release of cytochrome
c into the cytoplasm.
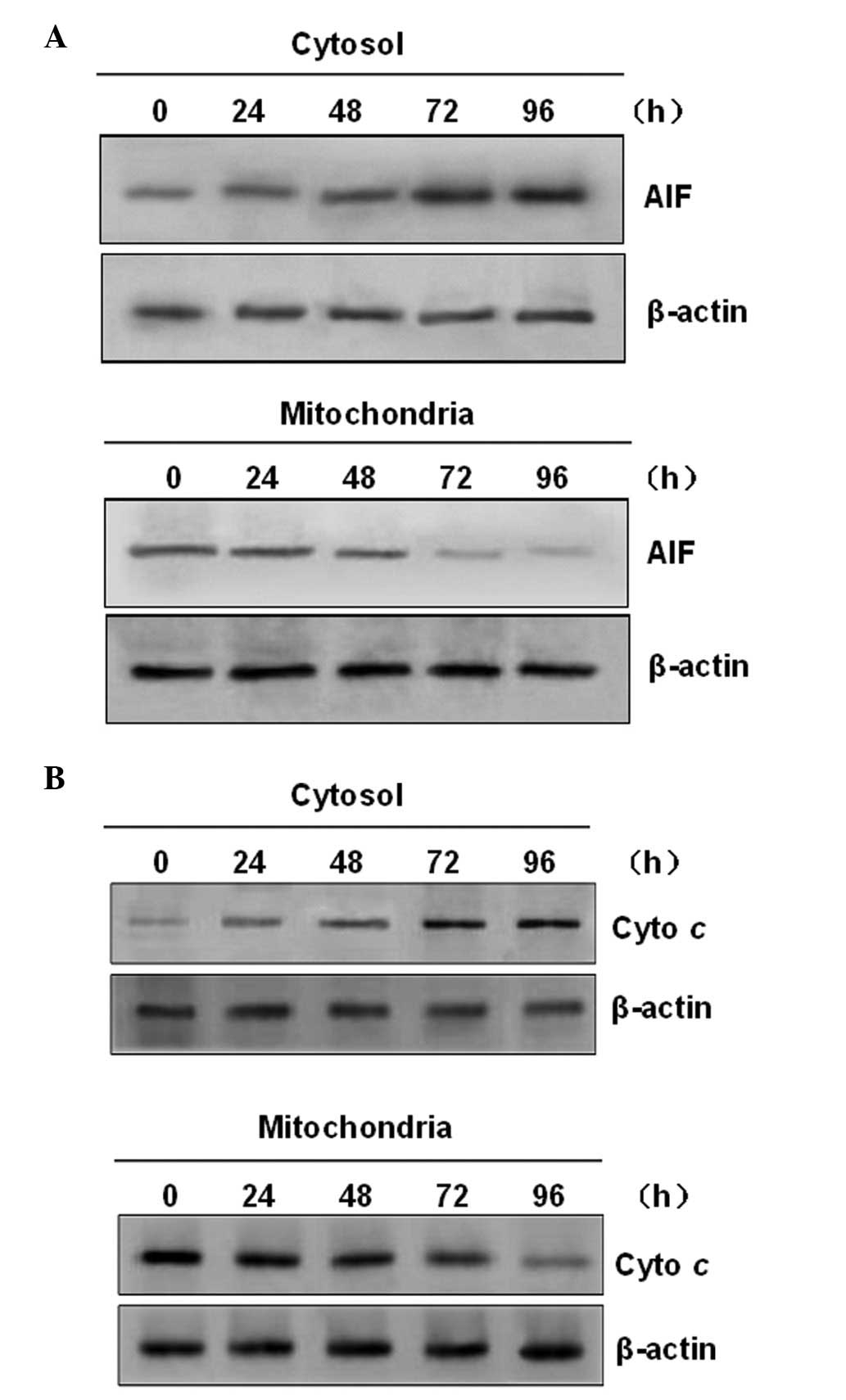
PAB induces translocation of
mitochondrial apoptotic factors
The cells were analyzed for alterations in the
mitochondria, which have been shown to be important in numerous
forms of apoptotic cell death. Mitochondrial damage occurring as a
result of drug-induced apoptosis is often accompanied by the
release of mitochondrial apoptotic factors into the cytosol
(17). Therefore, it was
investigated whether AIF was involved in PAB-induced apoptosis. The
results of the western blot analysis demonstrated that PAB
treatment induced the release of AIF from the mitochondria into the
cytosol in a time-dependent manner (Fig. 5A). These characteristics are
typically due to apoptosis. In addition, it was determined whether
treatment of U937 cells with PAB induced the release of cytochrome
c (a mitochondrial protein localized in the mitochondrial
intermembrane space) into the cytosol. As shown in Fig. 5B, PAB treatment mobilized
cytochrome c from the mitochondria into the cytosol.
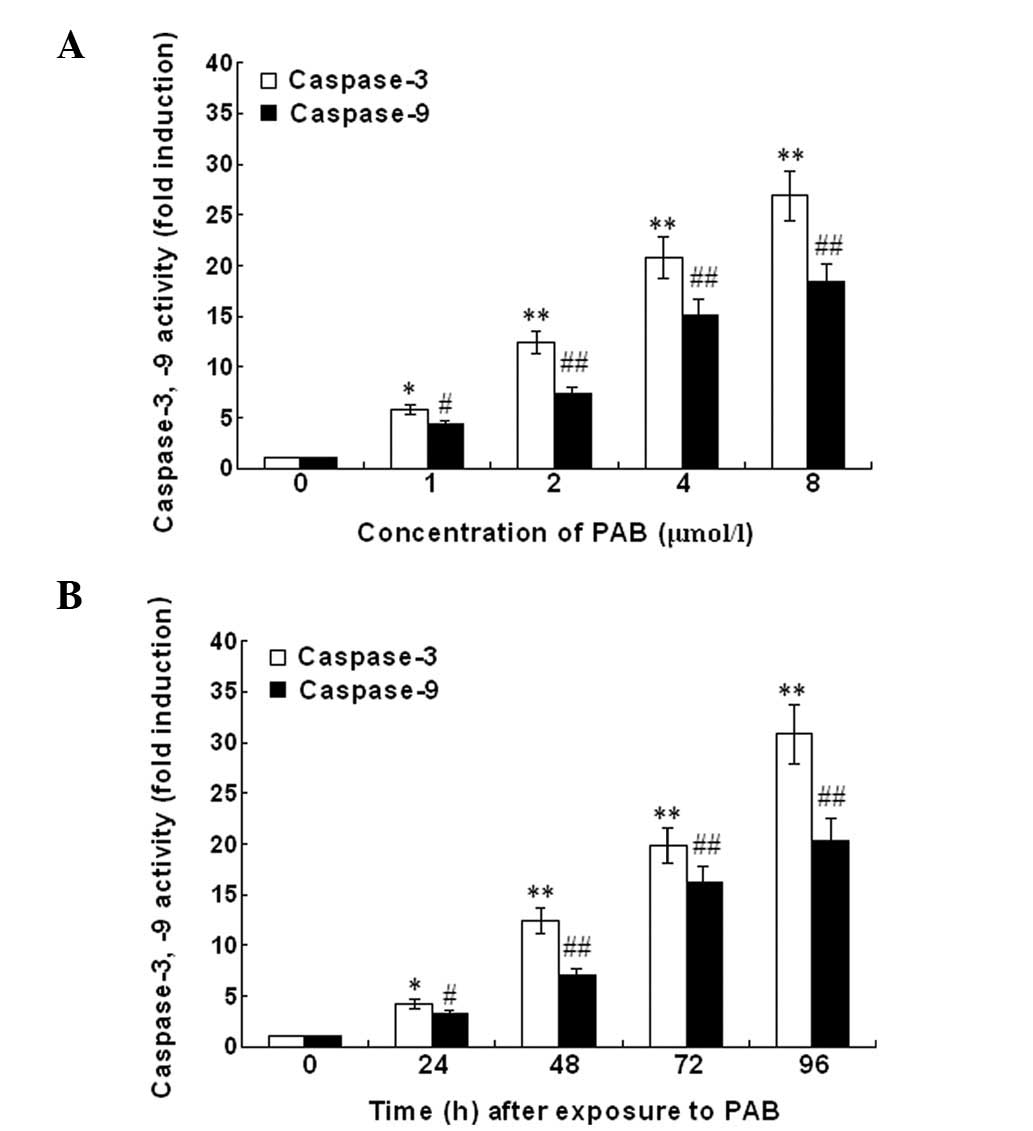
Caspase-3 and -9 activity
The caspase-3 and -9 activity in U937 cells
incubated with PAB is shown in Fig.
6. The treatment of U937 cells with PAB for different times at
a concentration of 2 mg/ml or for 48 h at concentrations of 0, 1,
2, 4 and 8 μM demonstrated a significant increase of caspase-3 and
-9 activation. The activity of caspase-3 and -9 in U937 cells
following PAB treatment was upregulated in a dose- and
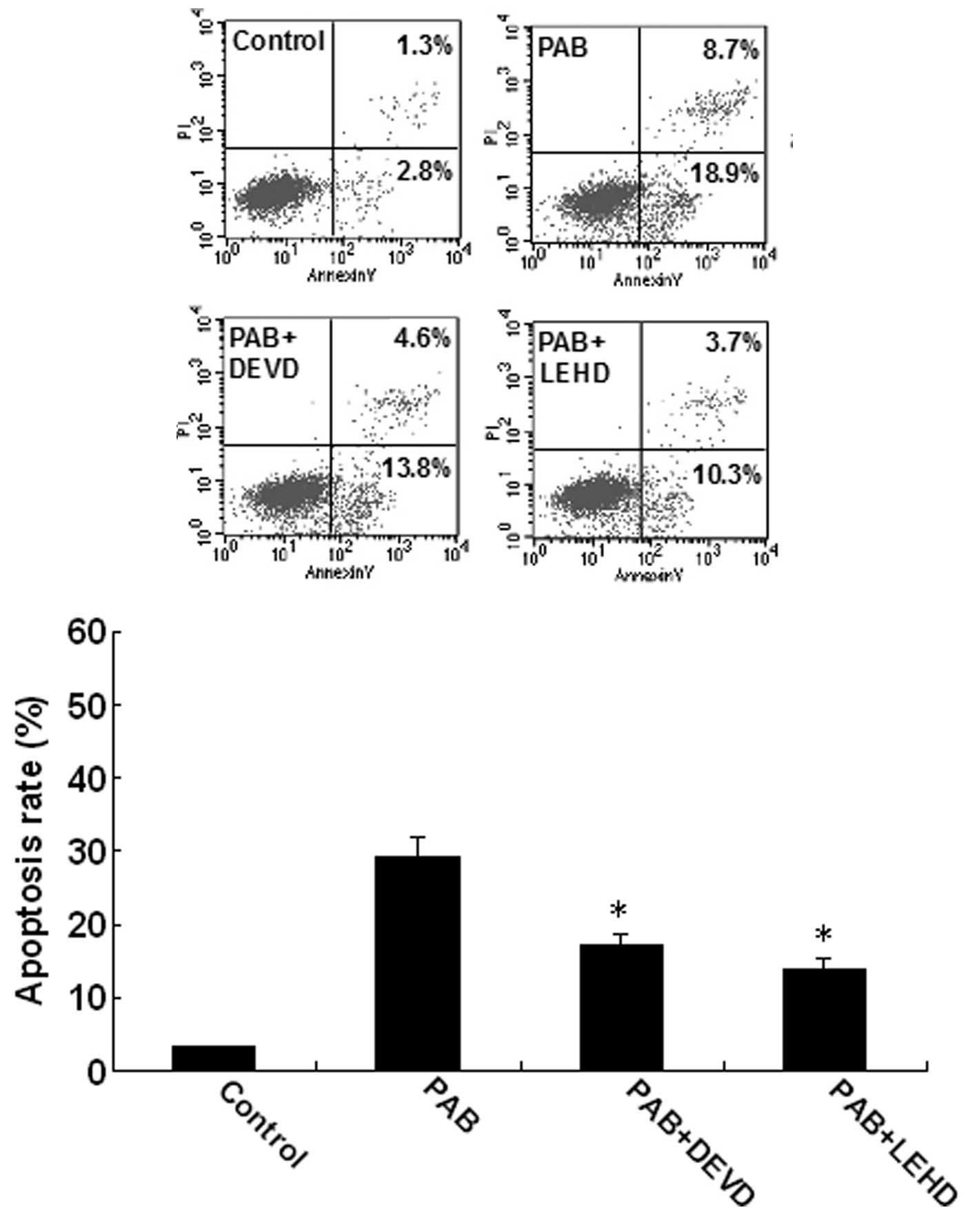
time-dependent manner. To characterize the pathway of apoptosis
execution, experiments were conducted using Z-DEVD-FMK and
Z-LEHD-FMK. The rate of apoptosis was significantly reduced by
Z-DEVD-FMK and Z-LEHD-FMK (Fig.
7). In conclusion, the data demonstrate that caspase-3 and -9
mediated PAB-induced U937 cell apoptosis.
Discussion
Previous studies have demonstrated that PAB induces
growth inhibition, cell cycle arrest and apoptosis in a variety of
cancer cell lines, including breast cancer, colon cancer,
hepatocellular carcinoma and melanoma cells (18–22).
In the present study, the effects of PAB on cell growth and death
in U937 cells (a human acute monocytic leukemia cell line) and the
underlying mechanisms were investigated. It was demonstrated that
PAB induced U937 cell apoptosis in a time- and dose-dependent
manner.
Apoptosis, or programmed cell death, is important in
a variety of physiological processes during fetal development and
in adult life (23–25). Cell shrinkage, chromatin
condensation and nuclear fragmentation are the morphological
hallmarks of apoptosis (26). The
inhibition of apoptosis, a universal and efficient cellular suicide
pathway, is one of the predominant characteristics of cancer
(27). Therefore, the induction of
apoptosis is an aim of various antitumor therapies (28). Numerous studies have demonstrated
that the majority of anticancer agents induce cell death through
apoptotic pathways. Previously, several unrelated natural products
were observed to elicit their cytotoxic effects by targeting the
mitochondria, thereby acting as mitocans (11). The mitochondrial membrane potential
and the release of cytochrome c are fine-tuned by the
balance between Bax and Bcl-xL (13). An increase in the ratio of Bax to
Bcl-xL stimulates the release of cytochrome c from the
mitochondria into the cytosol, where it binds to Apaf-1, leading to
the activation of caspase-9. The present study showed that PAB
increased the ratio of Bax to Bcl-xL and induced the release of
cytochrome c into the cytosol in U937 cells. This suggested
that the mitochondria may function in the mediation of the
apoptotic signal in PAB-induced apoptosis in U937 cells.
The Bcl-2 family of proteins are essential in
apoptotsis. They are regulators of the mitochondrial membrane
permeability and intermembrane space protein efflux, according to
the opposing fractions of the anti-apoptotic and pro-apoptotic
members (16). The Bcl-2 family of
proteins are divided into three subclasses: Bcl-2-like survival
members, such as Bcl-xl, Bcl-2 and Mcl-1; Bax-like death members,
such as Bax, Bak and Bcl-xs; and BH3-only death members, including
Bim, Bid and Bik (16). Noxa
upregulation/Mcl-1 downregulation upstream of subsequent
mitochondrial outer membrane permeabilization and cytochrome
c release has recently emerged as a key mechanism underlying
potent apoptogenicity of various experimental drugs targeting
metastatic melanoma (29). The
present study demonstrated that PAB significantly increased the
expression of Bax and Noxa, and decreased the expression of Mcl-1,
Bcl-xl and Bcl-2. However, the expression of tBid was not
significantly affected by treatment with PAB.
As downstream products of cytochrome c,
caspases are critical mediators of the principle factors in
apoptotic cells. Caspase-3 is a frequently activated death
protease, catalyzing the specific cleavage of numerous important
cellular proteins (30). Caspases
are classified by their mode of activation as either initiator or
effector caspases. Initiators, such as caspase-8 and -9, are
referred to as apical caspases, which are activated by a variety of
apoptotic signals. In the present study, the activation of
caspase-9 was observed to occur in a dose- and time-dependent
manner. Previous studies have suggested that caspase-3 is involved
in several events in the apoptosis pathway, including cytochrome
c release, DNA fragmentation and nuclear collapse. Caspase-3
is downstream of caspase-9; thus, it is plausible that applying
caspase-3 or -9 inhibitors would reduce PAB-induced apoptosis, as
was demonstrated in the present study. These data suggest that
apoptosis by PAB was mediated through the mitochondrial pathway
involving caspase-9 and -3.
In conclusion, the results revealed the importance
of the mitochondrial signaling pathways in response to PAB, and
extended the understanding of the molecular mechanisms mediating
these responses. Targeting PAB may be a potential novel therapeutic
strategy for leukemia treatment, and these results may provide a
foundation for the development of targeted anti-leukemia therapies
and drug screening. PAB-induced U937 cell apoptosis is a complex
process. Moreover, the in vivo relevance of these results
obtained from in vitro cell culture require verification in
animal models.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81101224), the
National Natural Science Foundation of Liaoning Province (grant no.
201202270) and the Outstanding Scientific Fund of Shengjing
Hospital (grant no. 201206).
References
|
1
|
Ma G, Chong L, Li XC, Khan IA, Walker LA
and Khan SI: Selective inhibition of human leukemia cell growth and
induction of cell cycle arrest and apoptosis by pseudolaric acid B.
J Cancer Res Clin Oncol. 136:1333–1340. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li YC, Tyan YS, Kuo HM, Chang WC, Hsia TC
and Chung JG: Baicalein induced in vitro apoptosis undergo caspases
activity in human promyelocytic leukemia HL-60 cells. Food Chem
Toxicol. 42:37–43. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao D, Lin F, Wu X, Zhao Q, Zhao B, Lin
P, Zang Y and Yu X: Pseudolaric acid B induces apoptosis via
proteasome-mediated Bcl-2 degradation in hormone-refractory
prostate cancer DU145 cells. Toxicol In Vitro. 26:595–602. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim EJ, Park SY, Lee JY and Park JH:
Fucoidan present in brown algae induces apoptosis of human colon
cancer cells. BMC Gastroenterol. 10:962010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shinkai K, Akedo H, Mukai M, Imamura F,
Isoai A, Kobayashi M and Kitagawa I: Inhibition of in vitro tumor
cell invasion by ginsenoside Rg3. Jpn J Cancer Res. 87:357–362.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thompson CB: Apoptosis in the pathogenesis
and treatment of disease. Science. 267:1456–1462. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nicholson DW: From bench to clinic with
apoptosis-based therapeutic agents. Nature. 407:810–816. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Danial NN and Korsmeyer SJ: Cell death:
critical control points. Cell. 116:205–219. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim KN, Ham YM, Moon JY, Kim MJ, Jung YH,
Jeon YJ, Lee NH, Kang N, Yang HM, Kim D and Hyun CG: Acanthoic acid
induces cell apoptosis through activation of the p38 MAPK pathway
in HL-60 human promyelocytic leukaemia. Food Chem. 135:2112–2117.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Du RH, Cui JT, Wang T, Zhang AH and Tan
RX: Trichothecin induces apoptosis of HepG2 cells via caspase-9
mediated activation of the mitochondrial death pathway. Toxicon.
59:143–150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guizzunti G, Theodorakis EA, Yu AL,
Zurzolo C and Batova A: Cluvenone induces apoptosis via a direct
target in mitochondria: a possible mechanism to circumvent
chemo-resistance? Invest New Drugs. 30:1841–1848. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rastogi RP, Singh SP, Häder DP and Sinha
RP: Detection of reactive oxygen species (ROS) by the
oxidant-sensing probe 2′,7′-dichlorodihydrofluorescein diacetate in
the cyanobacterium Anabaena variabilis PCC 7937. Biochem Biophys
Res Commun. 397:603–607. 2010.PubMed/NCBI
|
|
13
|
An J, Gao Y, Wang J, Zhu Q, Ma Y, Wu J,
Sun J and Tang Y: Flavokawain B induces apoptosis of non-small cell
lung cancer H460 cells via Bax-initiated mitochondrial and JNK
pathway. Biotechnol Lett. 34:1781–1788. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu JH, Cui Q, Jiang YY, Yang W, Tashiro S,
Onodera S and Ikejima T: Pseudolaric acid B induces apoptosis,
senescence, and mitotic arrest in human breast cancer MCF-7. Acta
Pharmacol Sin. 28:1975–1983. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Circu M and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang C, Chen X, Guo B, Huang W, Shen T,
Sun X, Xiao P and Zhou Q: Induction of apoptosis by Icariside II
through extrinsic and intrinsic signaling pathways in human breast
cancer MCF7 cells. Biosci Biotechnol Biochem. 76:1322–1328. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kroemer G and Reed JC: Mitochondrial
control of cell death. Nat Med. 6:513–519. 2000. View Article : Google Scholar
|
|
18
|
Yu JH, Wang HJ, Li XR, Tashiro S, Onodera
S and Ikejima T: Protein tyrosine kinase, JNK, and ERK involvement
in pseudolaric acid B-induced apoptosis of human breast cancer
MCF-7 cells. Acta Pharmacol Sin. 29:1069–1076. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ko JK, Leung WC, Ho WK and Chiu P: Herbal
diterpenoids induce growth arrest and apoptosis in colon cancer
cells with increased expression of the nonsteroidal
anti-inflammatory drug-activated gene. Eur J Pharmacol. 559:1–13.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong X, Wang M, Wu Z, Tashiro S, Onodera S
and Ikejima T: Pseudolaric acid B induces apoptosis via activation
of c-Jun N-terminal kinase and caspase-3 in HeLa cells. Exp Mol
Med. 36:551–556. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gong XF, Wang MW, Tashiro S, Onodera S and
Ikejima T: Pseudolaric acid B induces apoptosis through p53 and
Bax/Bcl-2 pathways in human melanoma A375-S2 cells. Arch Pharm Res.
28:68–72. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu J, Li X, Tashiro S, Onodera S and
Ikejima T: Bcl-2 family proteins were involved in pseudolaric acid
B-induced autophagy in murine fibrosarcoma L929 Cells. J Pharmacol
Sci. 107:295–302. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yim SY, Lee YJ, Lee YK, Jung SE, Kim JH,
Kim HJ, Son BG, Park YH, Lee YG, Choi YW and Hwang DY: Gomisin N
isolated from Schisandra chinensis significantly induces
anti-proliferative and pro-apoptotic effects in hepatic carcinoma.
Mol Med Rep. 2:725–732. 2009.PubMed/NCBI
|
|
24
|
Tanaka T, Bai T, Utsunomiya H, Fukumoto T
and Yukawa K: STAT3 enhances intracellular Fas-mediated apoptotic
signals in HHUA human endometrial epithelial cells. Mol Med Rep.
4:307–312. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jeong JK, Moon MH, Seo JS, Seol JW, Lee YJ
and Park SY: Sulforaphane blocks hypoxia-mediated resistance to
TRAIL-induced tumor cell death. Mol Med Rep. 4:325–330.
2011.PubMed/NCBI
|
|
26
|
Saraste A and Pulkki K: Morphologic and
biochemical hallmarks of apoptosis. Cardiovasc Res. 45:528–537.
2000. View Article : Google Scholar
|
|
27
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar
|
|
28
|
Schneider-Jakob S, Corazza N, Badmann A,
Sidler D, Stuber-Roos R, Keogh A, Frese S, Tschan M and Brunner T:
Synergistic induction of cell death in liver tumor cells by TRAIL
and chemotherapeutic drugs via the BH3-only proteins Bim and Bid.
Cell Death Dis. 1:e862010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Qiao SX, Cabello CM, Lamore SD, Lesson JL
and Wondrak GT: D-Penicillamine targets metastatic melanoma cells
with induction of the unfolded protein response (UPR) and Noxa
(PMAIP1)-dependent mitochondrial apoptosis. Apoptosis.
17:1079–1094. 2012. View Article : Google Scholar
|
|
30
|
Yuan Z, Long C, Junming T, Qihuan L,
Youshun Z and Chan Z: Quercetin-induced apoptosis of HL-60 cells by
reducing PI3K/Akt. Mol Biol Rep. 39:7785–7793. 2012. View Article : Google Scholar : PubMed/NCBI
|