Introduction
Lung cancer is the leading cause of
cancer-associated mortality worldwide, and non-small cell lung
cancer (NSCLC), which consists predominantly of squamous cell
carcinoma (SCC) and adenocarcinoma (AD) (1), accounts for ~80% of all lung cancer
cases. Thus, understanding of the mechanisms of lung carcinogenesis
in NSCLC subtypes is urgently required.
The Silent information regulator 2 (Sir2)
family of genes, encoding a group of nicotinamide adenine
dinucleotide (NAD+)-dependent histone deacetylases, is
highly conserved from bacteria to humans (2). Mammalian cells possess seven homologs of
yeast Sir2, Silent mating type information regulation 2
homologs (sirtuins); these homologs, SIRTs 1–7, have a common
catalytic domain with Sir2 (3).
SIRT1, which localizes predominantly in the nucleus, is the closest
homolog of yeast Sir2 and has been extensively studied. Numerous
studies have demonstrated that SIRT1 is overexpressed in various
types of cancer, including breast (4), liver (5),
prostate (6) and lung cancers
(7,8),
and that SIRT1 inhibitors are able to suppress tumor growth. SIRT1
is responsible for the deacetylation of various transcription
factors that are involved in stress responses and apoptosis,
including p53 (9), Ku70 (10), Nuclear factor kB (8), Forkhead box protein O (11) and Hypermethylated in cancer 1 (HIC1)
(12,13).
The High-mobility group A (HMGA) family comprises
three proteins (HMGA1a, HMGA1b and HMGA2), which are encoded by two
distinct genes; HMGA1a and HMGA1b proteins are generated through
alternative splicing of a single gene (14). During embryogenesis, HMGA protein
expression is high (15,16), whilst normal adult tissues exhibit low
or undetectable HMGA expression. However, high HMGA expression
levels have been observed in human malignant neoplasias, including
carcinomas of the thyroid (17),
colon (18), prostate (19), pancreas (20), cervix (21), ovary (22) and breast (23). Although HMGA proteins alone do not
exert transcriptional activity, they are able to alter chromatin
structure, and thus regulate the expression of a number of genes,
by interacting with the transcription machinery (24,25).
Overexpression of HMGA proteins is associated with a highly
malignant phenotype and a poor prognostic index, as their
overexpression correlates with metastasis and reduced survival time
(26). P53 is a
well-established tumor suppressor gene (27), the product of which mediates tumor
development (28). Recently,
HMGA1-p53 interactions have been demonstrated to affect
carcinogenesis. HMGA1 binds with p53, thus interfering with the
p53-mediated transcription of BCL2-associated X protein (BAX),
P21Waf1, MDM2 and B-Cell CLL/Lymphoma 2 (BCL2); this results in
reduced p53-dependent apoptosis (29–31). HMGA1
also inhibits p53 apoptotic function by relocalizing the nuclear
p53 proapoptotic activator Homeodomain-interacting protein kinase 2
(HIPK2) to the cytoplasm (32).
Based on these reports, we hypothesized that SIRT1
may affect the progression of lung cancer in association with
HMGA1. To test this, the expression levels of SIRT1 and HMGA1 and
the prognostic value of these proteins in NSCLC were investigated.
The association between SIRT1 and HMGA1 expression was also
evaluated.
Materials and methods
Patients and tissue samples
Paired tumor and normal lung specimens were obtained
from 260 patients with NSCLC who were surgically treated at
Zhejiang Hospital (Hangzhou, China) between 2002 and 2010. Informed
consent was obtained from the patients, in compliance with the
Declaration of Helsinki, and the study was approved by the ethics
committee of Zhejiang Hospital (no. 2013-k-2). Clinicopathological
data, including gender, age and presence of distant metastasis,
were collected from medical records. All cases were reviewed and
classified according to the criteria of the World Health
Organization (WHO) (33).
Pathological staging was based on the Tumor-Node-Metastasis (TNM)
staging system of the American Joint Committee on Cancer (34). The patients were grouped according to
age, gender, tumor size, TNM stage, histological type, histological
grade, tumor invasion (vascular and pleural invasion), presence of
lymph node metastasis and smoking history. The mean follow-up
period was 37.1 months (range, 0–128 months). All patients were
followed up through March 2012. The 260 lung tumors comprised 127
cases of AD and 133 cases of SCC. For the normal lung tissue, 100
of these cases were selected and tissue was taken at a distance of
>3 cm from the tumor. All specimens were fixed in 10% neutral
formalin.
Immunohistochemical analysis
All tissues were neutral formalin-fixed and
paraffin-embedded. Tissues were sectioned into 4-µm slices and then
processed using standard deparaffinization and rehydration
techniques. Immunohistochemical staining was performed using the
enhanced labeled polymer method (35). Briefly, tissue sections were subjected
to a boiling antigen retrieval procedure in sodium citrate buffer
(pH 6.0; Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing,
China) for 3 min. After blocking endogenous peroxidase activity
with peroxidase quenching solution (Zhongshan Golden Bridge
Biotechnology Co., Ltd.), the sections were incubated overnight at
4°C with a monoclonal rabbit anti-human SIRT1 antibody (clone E104;
ab32441; 1:200; Abcam, Cambridge, UK) and a polyclonal rabbit
anti-human HMGA1 antibody (ab4078; 1:200; Abcam). Negative control
sections were treated in the same manner except they were incubated
in phosphate-buffered saline (Zhongshan Golden Bridge Biotechnology
Co., Ltd.) without primary antibody. After applying the EnVision
System-HRP (Invitrogen) for 20 min, the reaction products were
visualized by immersing in a mixture which contained 1 drop of
DAB-SuperPicture GenIHC Detection Kit and 1 ml DAB substrate
buffer, and the samples were counterstained with hematoxylin
(Invitrogen). Immunohistochemical scoring was determined by
consensus by two pathologists who were blinded to the
clinicopathological information associated with the specimens.
SIRT1 and HMGA1 expression levels were semi-quantitatively scored
by assessing the intensity of staining (0, no staining; 1, mild
staining; 2, moderate staining; and 3, strong staining) and the
percentage of positively stained cells (0, <30%; 1, 30–49%; 2,
50–69%; and 3, ≥70%). The sum index was obtained by totaling the
staining intensity and percentage scores. A final score of ≥4 was
considered to indicate positive expression in a specimen;
otherwise, the tumor was considered negative, based on the findings
of previous reports (36,37).
Statistical analysis
Statistical analysis was performed using SPSS
software version 17.0 (SPSS, Inc., Chicago, IL, USA). P-values for
differences between groups were determined by χ2 tests.
Overall survival (OS) time was calculated from the day of surgery
to the date of mortality or the last follow-up. Univariate Cox
regression analyses were performed to estimate the effect of
clinicopathological factors and expression of each marker on OS.
Survival curves were calculated according to the Kaplan-Meier
method; comparison was performed using the log-rank test. P<0.05
was considered to indicate statistically significant
differences.
Results
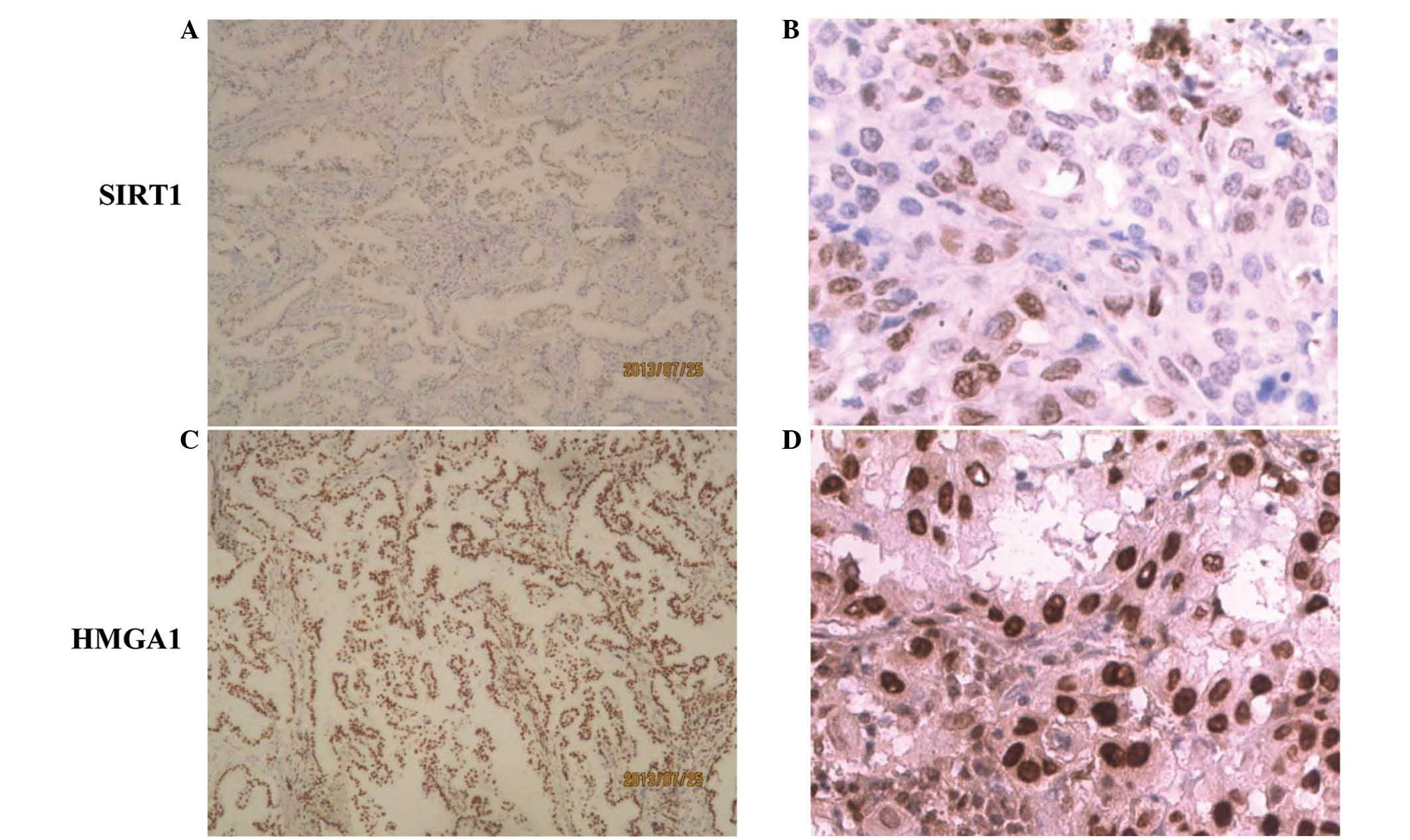
SIRT1 and HMGA1 expression was evaluated
immunohistochemically in 260 NSCLC and 100 normal lung specimens
(Fig. 1). The median age of the
patients was 63 years (range, 35–90 years; Table I). Although immunoreactivity for SIRT1
has been reported in the cytoplasm and the nucleus (38), SIRT1 exhibited only nuclear staining
the current study; its score in normal tissues was 0–1. HMGA1
expression was also found to be nuclear, and was weak in normal
tissues, with overall scores of 2–3. Among the NSCLC samples, 12 of
133 SCC cases (9.0%) were SIRT1+, all exhibiting
positive expression in the nucleus. A significantly higher
percentage of the AD specimens were SIRT1+ (18.9%;
P=0.021 vs. SCC specimens). HMGA1+ expression, which was
also localized to the cell nuclei, was observed in 62.6% of NSCLC
patients (163 of 260 cases), and was significantly associated with
male gender (P=0.041), histological type (P=0.015) and degree of
differentiation (P=0.028), and also with positive SIRT1 expression
in NSCLC (P<0.001) (Table I).
 | Table I.Association between
clinicopathological characteristics and HMGA1 and SIRT1 expression,
and their effect on survival in non-small cell lung carcinoma
assessed by univariate Cox proportional hazards regression
analysis. |
Table I.
Association between
clinicopathological characteristics and HMGA1 and SIRT1 expression,
and their effect on survival in non-small cell lung carcinoma
assessed by univariate Cox proportional hazards regression
analysis.
|
|
| HMGA1
expression | SIRT1
expression | Univariate Cox
regression |
|---|
|
|
|
|
|
|
|---|
| Characteristic | n | Positive, n(%) | P-value | Positive, n(%) | P-value | Hazard ratio | P-value |
|---|
| All | 260 | 163 (62.7) |
| 36 (13.8) |
|
|
|
| Gender |
|
|
0.041 |
|
0.111 |
|
|
|
Male | 188 | 125 (66.5) |
| 30 (16.0) |
| 1.000 |
|
|
Female | 72 | 38
(52.8) |
| 6 (8.3) |
| 1.116 | 0.526 |
| Age, years |
|
|
0.310 |
|
0.768 |
|
|
|
≤69 | 175 | 106 (60.6) |
| 25 (14.3) |
| 1.083 |
|
|
>69 | 85 | 57
(67.1) |
| 11 (12.9) |
| 1.000 | 0.727 |
| Histological
type |
|
|
0.015 |
|
0.021 |
|
|
|
Squamous cell carcinoma | 133 | 93
(69.9) |
| 12 (9.0) |
| 1.000 |
|
|
Adenocarcinoma | 127 | 70
(55.1) |
| 24 (18.9) |
| 0.796 | 0.286 |
| Tumor size, cm |
|
|
0.898 |
|
0.683 |
|
|
|
<4 | 129 | 80
(62.0) |
| 19 (14.7) |
| 1.000 |
|
| ≥4 | 131 | 83
(63.4) |
| 17 (13.0) |
| 1.532 | 0.049 |
| T stage |
|
|
0.300 |
|
0.491 |
|
|
| T1 | 87 | 53
(60.9) |
| 10 (11.5) |
|
|
|
| T2 | 87 | 51
(58.6) |
| 13 (14.9) |
| 2.213 | 0.006 |
|
T3/T4 | 86 | 59
(68.6) |
| 13 (15.1) |
| 3.130 | 0.002 |
| Lymph node
metastasis |
|
|
0.602 |
|
0.608 |
|
|
|
Present | 156 | 100 (64.1) |
| 23 (14.7) |
| 1.000 |
|
|
Absent | 104 | 63
(60.6) |
| 13 (12.5) |
| 0.419 | <0.001 |
| Tumor invasion |
|
|
0.859 |
|
0.818 |
|
|
|
Absent | 213 | 133 (62.4) |
| 29 (13.6) |
| 1.000 |
|
|
Present | 47 | 30
(63.8) |
| 7 (14.9) |
| 1.659 | 0.047 |
| Degree of
differentiation |
|
|
0.028 |
|
0.300 |
|
|
|
Well | 61 | 31
(19.0) |
| 6 (16.7) |
| 1.000 |
|
|
Moderate/poor | 199 | 132 (81.0) |
| 30 (83.3) |
| 1.195 | 0.477 |
| Smoking |
|
|
0.123 |
|
0.100 |
|
|
|
Present | 163 | 108 (66.3) |
| 27 (16.6) |
| 1.000 |
|
|
Absent | 97 | 55
(56.7) |
| 9 (9.3) |
| 0.927 | 0.734 |
| HMGA1
expression |
|
|
|
| <0.001 |
|
|
|
Negative | 97 |
|
| 4 (4.1) |
| 1.165 |
|
|
Positive | 163 |
|
| 32 (19.6) |
| 1.000 | 0.477 |
| SIRT1
expression |
|
| <0.001 |
|
|
|
|
|
Negative | 224 | 131 (58.5) |
|
|
| 1.000 |
|
|
Positive | 36 | 32
(88.9) |
|
|
| 1.024 | 0.939 |
To clarify the clinicopathological significance of
the combined SIRT1/HMGA1 expression status, the patients were
divided into four groups based on SIRT1 and HMGA1 status
(SIRT1−/HMGA1−,
SIRT1−/HMGA1+,
SIRT1+/HMGA1− or
SIRT1+/HMGA1+), and the association with
clinicopathological parameters was assessed. As shown in table II,
SIRT1+/HMGA1− was associated with male gender
(P=0.016) and poorly and moderately differentiated tumors
(P=0.025). Univariate Cox regression analysis of OS time was
conducted in the 260 patients with NSCLC for whom complete
information for all variables was available. No significant
difference in OS time was observed between the four groups
(P>0.05). By contrast, tumor size (P=0.049), T1 stage (P=0.005),
T2 stage (P=0.006), T3/T4 stage (P=0.002), positive lymph node
metastasis (P<0.001) and tumor invasion (P=0.047) were
significantly associated with a shorter OS time on univariate Cox
regression analysis (Table I).
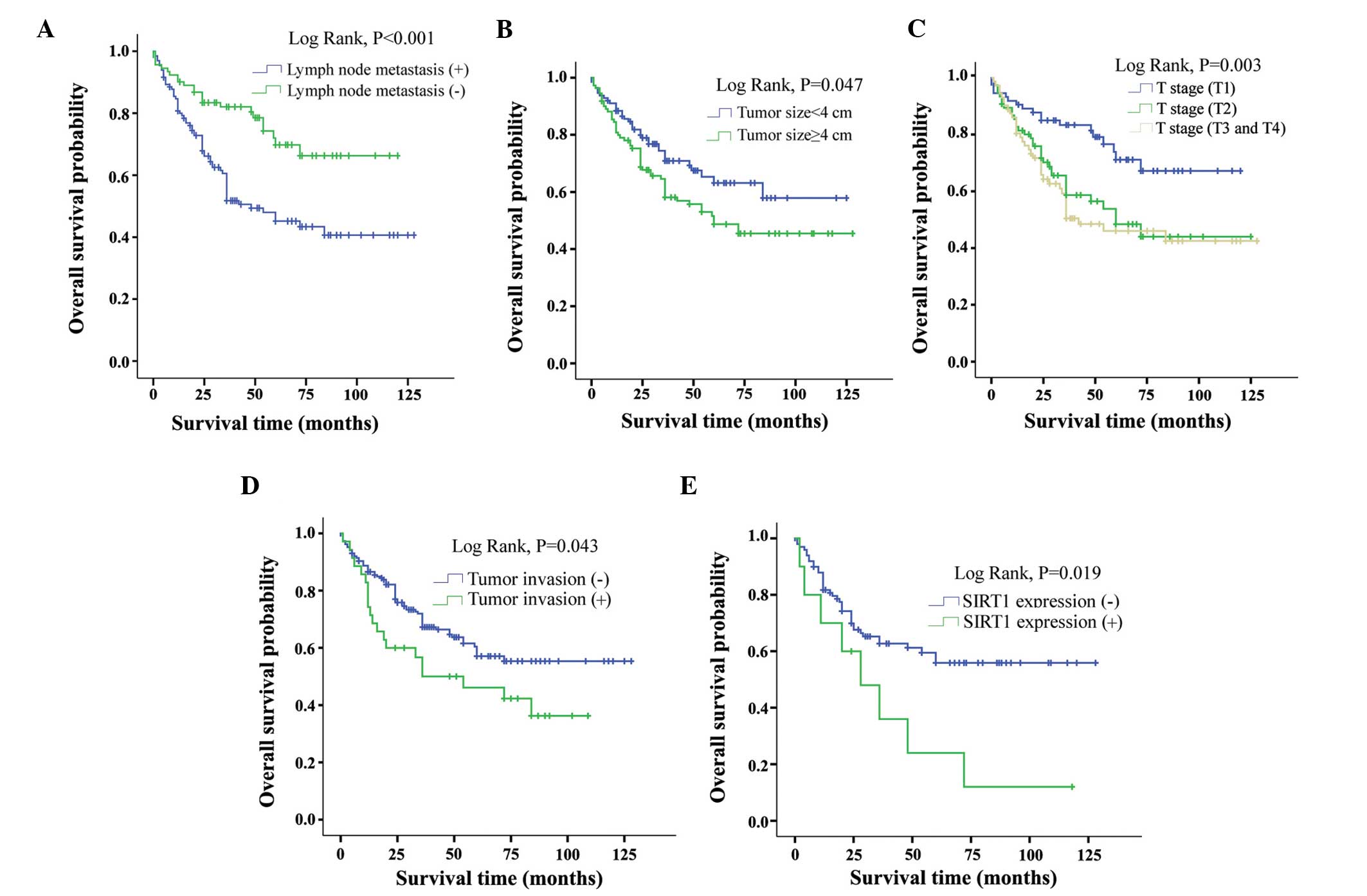
Kaplan-Meier survival curves for the effect of various factors on
OS are shown in Fig. 2. Tumor
invasion (P=0.043), lymph node metastasis (P<0.001), larger (≥4
cm)tumor size (P=0.047), higher pathological T stage (P=0.003) and
SIRT1+ SCC (P=0.019) predicted shorter OS time in NSCLC
patients (Fig. 2).
| Table II.Association between
clinicopathological characteristics and combined expression of
HMGA1/SIRT1. |
Table II.
Association between
clinicopathological characteristics and combined expression of
HMGA1/SIRT1.
|
|
| HMGA1/SIRT1
expression, n |
|
|---|
|
|
|
|
|
|---|
| Characteristic | n | −/− | −/+ | +/− | +/+ | P-value |
|---|
| All | 260 | 93 | 4 | 131 | 32 |
|
| Gender |
|
|
|
|
| 0.016 |
|
Male | 188 | 61 | 2 | 97 | 28 |
|
|
Female | 72 | 32 | 2 | 34 | 4 |
|
| Age, years |
|
|
|
|
| 0.427 |
|
>69 | 85 | 28 | 0 | 46 | 11 |
|
|
≤69 | 175 | 65 | 4 | 85 | 21 |
|
| Histological
type |
|
|
|
|
| 0.281 |
|
Squamous cell carcinoma | 133 | 38 | 2 | 83 | 10 |
|
|
Adenocarcinoma | 127 | 55 | 2 | 48 | 22 |
|
| Degree of
differentiation |
|
|
|
|
| 0.025 |
|
Well | 61 | 29 | 1 | 26 | 5 |
|
|
Moderate and poor | 199 | 64 | 3 | 105 | 27 |
|
| Tumor size, cm |
|
|
|
|
| 0.124 |
|
<4 | 129 | 46 | 3 | 64 | 16 |
|
| ≥4 | 131 | 47 | 1 | 67 | 16 |
|
| T stage |
|
|
|
|
| 0.255 |
| T1 | 87 | 33 | 1 | 44 | 9 |
|
| T2 | 87 | 34 | 2 | 40 | 11 |
|
|
T3/T4 | 86 | 26 | 1 | 47 | 12 |
|
| Lymph node
metastasis |
|
|
|
|
| 0.518 |
|
Present | 156 | 53 | 3 | 80 | 20 |
|
|
Absent | 104 | 40 | 1 | 51 | 12 |
|
| Tumor invasion |
|
|
|
|
| 0.767 |
|
Absent | 213 | 76 | 4 | 108 | 25 |
|
|
Present | 47 | 17 | 0 | 23 | 7 |
|
| Smoking |
|
|
|
|
| 0.064 |
|
Present | 163 | 51 | 4 | 85 | 23 |
|
|
Absent | 97 | 42 | 0 | 46 | 9 |
|
Discussion
The present study evaluated SIRT1 and HMGA1
expression immunohistochemically in human NSCLC. Previous studies
revealed that SIRT1 can act as a tumor suppressor by repressing a
number of oncogenes (39–41). By contrast, SIRT1 expression has been
observed to be increased in various human malignancies (6,42), and
changes in SIRT1-mediated signaling allows mammalian cells to
survive under oxidative stress and DNA damage, which are closely
associated with tumorigenesis (43,44),
cancer progression and poor prognosis in cancer patients (3,4,36). The present study identified an
association of SIRT1 overexpression with shorter OS time and poor
prognostic indicators in SCC, which was consistent with the results
of previous studies (4,36,45). By
contrast, certain reports have found a significant association
between SIRT1 overexpression and more favorable prognosis in serous
carcinoma of the ovary (46) and
colorectal cancer (47,48). In the present study, Kaplan-Meier
survival analysis suggested that patients with tumor invasion
(P=0.043), lymph node metastasis (P<0.001), larger tumor size
(P=0.047) and higher pathological T stage (P=0.001) have poorer
prognoses (Fig. 2).
In the current study, SIRT1+ SCC was
significantly associated with shorter OS time compared with
SIRT1– SCC (P=0.019). However, no association was
identified between SIRT1 expression in AD and shorter OS. A
previous study reported that patients with SCC had significantly
higher low-acetylated p53 status compared with patients with AD
(P=0.012), and that low-acetylated p53 was associated with poorer
survival relative to that of patients with acetylated p53 (13). These findings indicate that p53
deacetylation may play a role in lung tumorigenesis, particularly
for SCC patients. The different status of p53
acetylation/deacetylation in patients with lung AD and SCC may be
caused by the distinct deregulation of SIRT1 epigenetic control
(13). In addition, AD is now
classified according to its predominant pattern following
comprehensive histological subtyping into lepidic, acinar,
papillary, solid, mucinous, mixed muciniusand nonmucinous
adenocarcinoma, and other subtype AD, based on the WHO
classification criteria (33). In the
present study, there were 71 cases of lepidic adenocarcinoma, 6
cases of acinar adenocarcinoma, 7 cases of papillary
adenocarcinoma, 4 cases of solid adenocarcinoma, 25 cases of
mucinous adenocarcinoma, 14 cases of mixed mucinius and nonmucinous
subtype AD, however, no association was identified between SIRT1
expression in AD subtypes and shorter OS. This may be due to the
heterogeneity of the subtype of adenocarcinoma, and the
pathogenesis of these subtypes is different. Taken together, these
results suggest that SIRT1 expression is associated with tumor
progression and poor prognosis in NSCLC, particularly for patients
with SCC.
HMGA protein overexpression and gene rearrangements
are frequent in various types of human cancer (22). HMGA localization appears to be almost
exclusively nuclear (30), and was so
in the present study. HMGA1 overexpression is associated with a
highly malignant phenotype and a poor prognostic index, due to its
association with metastasis and reduced survival time (26). Sarhadi et al (49) observed that HMGA1 overexpression was
present in all types of lung cancer, and was an independent
indicator of poor prognosis, particularly in patients with AD. By
contrast, the present study found that neither univariate Cox
regression analysis nor Kaplan-Meier survival analysis associated
HMGA1+ expression with poor OS (P>0.05 for both
methods). This discrepancy may be due to a high rate of patients
lost to follow-up. However, the present study did identify a clear
association between HMGA1+ expression and poor cellular
differentiation (P=0.028). In addition, SCC patients were more
likely to express HMGA1 than AD patients (P=0.015), and
HMGA1+ expression was significantly associated with male
gender (P=0.041).
To the best of our knowledge, the current study is
the first to report on the association between SIRT1 and HMGA1
expression in lung cancer, although their respective roles in
tumorigenesis have been widely studied previously (12–15,26). HMGA1
proteins are reported to be important in the process of
carcinogenesis, based on the HMGA1-p53 interaction. As p53
functions as a tumor suppressor, P53 mutations commonly lead
to the development of cancer. By binding p53, HMGA1 interferes with
p53-mediated transcription of BAX, P21Waf1, MDM2 and BCL2, leading
to a reduction in p53-dependent apoptosis (29–31). HMGA1
also counteracts p53 transcriptional activity by relocalizing the
nuclear p53 proapoptotic activator HIPK2 to the cytoplasm, thereby
inhibiting the apoptotic function of p53 (32). SIRT1 is able to promote cell survival
or inhibit apoptosis by deacetylating p53 (50,51).
Deacetylation of the p53 protein promotes its accumulation during
the stress response, and is required for p53-induced apoptosis and
arrest of cell growth (52,53). It has been reported that p53 is
responsible for transcriptional repression of SIRT1 in
humans and mice, which depends upon p53 response elements in the
proximal promoter. The region of the SIRT1 promoter
containing the p53-binding sequence also contains a binding site
for the transcriptional repressor HIC1 (12). In addition, dysregulation of the
HIC1-SIRT1-p53 loop may be involved in lung tumorigenesis and
disease outcome (13). Taken
together, these results indicate there is a mechanism connecting
the expression of SIRT1 and HMGA1. We hypothesized that SIRT1 and
HMGA1 may interact with each other by inactivating or activating
p53-mediated transcription. Therefore, the current study
investigated the possible associations between SIRT1 and HMGA1
expression and the clinicopathological significance of combined
expression of these factors in NSCLC. HMGA1 expression was found to
be significantly correlated with SIRT1 expression (P<0.001).
SIRT1+/HMGA1− expression was significantly
associated with male gender (P=0.016) and and poorly and moderately
differentiated tumors (P=0.025; Table
II). Thus, SIRT1 and HMGA1 may cooperate during tumor
progression in NSCLC, particularly in patients with SCC.
In conclusion, the present study demonstrated the
expression of SIRT1 and HMGA1 in lung cancer and their association
with clinicopathological factors and patient survival. NSCLC
specimens in this study frequently expressed SIRT1 and HMGA1, and
their expression was significantly associated with unfavorable
NSCLC characteristics. Furthermore, SIRT1 and HMGA1 expression were
found to be significantly correlated in NSCLC patients. SIRT1 and
HMGA1 may interact with each other through p53; this mechanism
merits further study, as do the SIRT1- and HMGA1-associated
pathways that are involved in NSCLC progression.
Acknowledgements
This work was supported by the Zhejiang Provincial
Health Bureau Foundation (grant nos. 2011KYA024 and
2013KYA004).
References
|
1
|
Niklinski J, Niklinska W, Chyczewski L,
Becker HD and Pluygers E: Molecular genetic abnormalities in
premalignant lung lesions: Biological and clinical implications.
Eur J Cancer Prev. 10:213–226. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Imai S, Armstrong CM, Kaeberlein M and
Guarente L: Transcriptional silencing and longevity protein
Sir2 is an NAD-dependent histone deacetylase. Nature.
403:795–800. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Blander G and Guarente L: The Sir2 family
of protein deacetylases. Annu Rev Biochem. 73:417–435. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee H, Kim KR, Noh SJ, Park HS, Kwon KS,
Park BH, Jung SH, Youn HJ, Lee BK, Chung MJ, et al: Expression of
DBC1 and SIRT1 is associated with poor prognosis for breast
carcinoma. Hum Pathol. 42:204–213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bae HJ, Chang YG, Noh JH, Kim JK, Eun JW,
Jung KH, Kim MG, Shen Q, Ahn YM, Kwon SH, et al: DBC1 does not
function as a negative regulator of SIRT1 in liver cancer. Oncol
Lett. 4:873–877. 2012.PubMed/NCBI
|
|
6
|
Kuzmichev A, Margueron R, Vaquero A,
Preissner TS, Scher M, Kirmizis A, Ouyang X, Brockdorff N,
Abate-Shen C, Farnham P and Reinberg D: Composition and histone
substrates of polycomb repressive group complexes change during
cellular differentiation. Proc Natl Acad Sci USA. 102:1859–1864.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Solomon JM, Pasupuleti R, Xu L, McDonagh
T, Curtis R, DiStefano PS and Huber LJ: Inhibition of SIRT1
catalytic activity increases p53 acetylation but does not alter
cell survival following DNA damage. Mol Cell Biol. 26:28–38. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yeung F, Hoberg JE, Ramsey CS, Keller MD,
Jones DR, Frye RA and Mayo MW: Modulation of NF-kappaB-dependent
transcription and cell survival by the SIRT1 deacetylase. EMBO J.
23:2369–2380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng HL, Mostoslavsky R, Saito S, Manis
JP, Gu Y, Patel P, Bronson R, Appella E, Alt FW and Chua KF:
Developmental defects and p53 hyperacetylation in Sir2 homolog
(SIRT1)-deficient mice. Proc Natl Acad Sci USA. 100:10794–10799.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cohen HY, Miller C, Bitterman KJ, Wall NR,
Hekking B, Kessler B, Howitz KT, Gorospe M, de Cabo R and Sinclair
DA: Calorie restriction promotes mammalian cell survival by
inducing the SIRT1 deacetylase. Science. 305:390–392. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brunet A, Sweeney LB, Sturgill JF, Chua
KF, Greer PL, Lin Y, Tran H, Ross SE, Mostoslavsky R, Cohen HY, et
al: Stress-dependent regulation of FOXO transcription factors by
the SIRT1 deacetylase. Science. 303:2011–2015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Naqvi A, Hoffman TA, DeRicco J, Kumar A,
Kim CS, Jung SB, Yamamori T, Kim YR, Mehdi F, Kumar S, et al: A
single-nucleotide variation in a p53-binding site affects
nutrient-sensitive human SIRT1 expression. Hum Mol Genet.
19:4123–4133. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tseng RC, Lee CC, Hsu HS, Tzao C and Wang
YC: Distinct HIC1-SIRT1-p53 loop deregulation in lung squamous
carcinoma and adenocarcinoma patients. Neoplasia. 11:763–770. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Johnson KR, Lehn DA and Reeves R:
Alternative processing of mRNAs encoding mammalian chromosomal
high-mobility-group proteins HMG-I and HMG-Y. Mol Cell Biol.
9:2114–2123. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou X, Benson KF, Ashar HR and Chada K:
Mutation responsible for the mouse pygmy phenotype in the
developmentally regulated factor HMGI-C. Nature. 376:771–774. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chiappetta G, Avantaggiato V, Visconti R,
Fedele M, Battista S, Trapasso F, Merciai BM, Fidanza V, Giancotti
V, Santoro M, et al: High level expression of the HMGI (Y) gene
during embryonic development. Oncogene. 13:2439–2446.
1996.PubMed/NCBI
|
|
17
|
Chiappetta G, Tallini G, De Biasio MC,
Manfioletti G, Martinez-Tello FJ, Pentimalli F, de Nigris F, Mastro
A, Botti G, Fedele M, et al: Detection of high mobility group I
HMGI(Y) protein in the diagnosis of thyroid tumors: HMGI(Y)
expression represents a potential diagnostic indicator of
carcinoma. Cancer Res. 58:4193–4198. 1998.PubMed/NCBI
|
|
18
|
Fedele M, Bandiera A, Chiappetta G,
Battista S, Viglietto G, Manfioletti G, Casamassimi A, Santoro M,
Giancotti V and Fusco A: Human colorectal carcinomas express high
levels of high mobility group HMGI(Y) proteins. Cancer Res.
56:1896–1901. 1996.PubMed/NCBI
|
|
19
|
Tamimi Y, van der Poel HG, Denyn MM, Umbas
R, Karthaus HF, Debruyne FM and Schalken JA: Increased expression
of high mobility group protein I(Y) in high grade prostatic cancer
determined by in situ hybridization. Cancer Res. 53:5512–5516.
1993.PubMed/NCBI
|
|
20
|
Abe N, Watanabe T, Izumisato Y, Masaki T,
Mori T, Sugiyama M, Chiappetta G, Fusco A, Fujioka Y and Atomi Y:
Diagnostic significance of high mobility group I(Y) protein
expression in intraductal papillary mucinous tumors of the
pancreas. Pancreas. 25:198–204. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bandiera A, Bonifacio D, Manfioletti G,
Mantovani F, Rustighi A, Zanconati F, Fusco A, Di Bonito L and
Giancotti V: Expression of HMGI(Y) proteins in squamous
intraepithelial and invasive lesions of the uterine cervix. Cancer
Res. 58:426–431. 1998.PubMed/NCBI
|
|
22
|
Masciullo V, Baldassarre G, Pentimalli F,
Berlingieri MT, Boccia A, Chiappetta G, Palazzo J, Manfioletti G,
Giancotti V, Viglietto G, et al: HMGA1 protein over-expression is a
frequent feature of epithelial ovarian carcinomas. Carcinogenesis.
24:1191–1198. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chiappetta G, Botti G, Monaco M,
Pasquinelli R, Pentimalli F, Di Bonito M, D'Aiuto G, Fedele M,
Iuliano R, Palmieri EA, et al: HMGA1 protein overexpression in
human breast carcinomas: Correlation with ErbB2 expression. Clin
Cancer Res. 10:7637–7644. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Grosschedl R, Giese K and Pagel J: HMG
domain proteins: Architectural elements in the assembly of
nucleoprotein structures. Trends Genet. 10:94–100. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thanos D and Maniatis T: The high mobility
group protein HMG I(Y) is required for NF-kappa B-dependent virus
induction of the human IFN-beta gene. Cell. 71:777–789. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fusco A and Fedele M: Roles of HMGA
proteins in cancer. Nat Rev Cancer. 7:899–910. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sionov RV and Haupt Y: The cellular
response to p53: The decision between life and death. Oncogene.
18:6145–6157. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kruse JP and Gu W: Modes of p53
regulation. Cell. 137:609–622. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Esposito F, Tornincasa M, Federico A,
Chiappetta G, Pierantoni GM and Fusco A: High-mobility group A1
protein inhibits p53-mediated intrinsic apoptosis by interacting
with Bcl-2 at mitochondria. Cell Death Dis. 3:e3832012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pierantoni GM, Rinaldo C, Esposito F,
Mottolese M, Soddu S and Fusco A: High Mobility Group A1 (HMGA1)
proteins interact with p53 and inhibit its apoptotic activity. Cell
Death Differ. 13:1554–1563. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Esposito F, Tornincasa M, Chieffi P, De
Martino I, Pierantoni GM and Fusco A: High-mobility group A1
proteins regulate p53-mediated transcription of Bcl-2 gene. Cancer
Res. 70:5379–5388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pierantoni GM, Rinaldo C, Mottolese M, Di
Benedetto A, Esposito F, Soddu S and Fusco A: High-mobility group
A1 inhibits p53 by cytoplasmic relocalization of its proapoptotic
activator HIPK2. J Clin Invest. 117:693–702. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sobin LH: The World Health Organization's
Histological Classification of Lung Tumors: A comparison of the
first and second editions. Cancer Detect Prev. 5:391–406.
1982.PubMed/NCBI
|
|
34
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Grbesa I, Pajares MJ, Martínez-Terroba E,
Agorreta J, Mikecin AM, Larráyoz M, Idoate MA, Gall-Troselj K, Pio
R and Montuenga LM: Expression of sirtuin 1 and 2 is associated
with poor prognosis in non-small cell lung cancer patients. PLoS
One. 10:e01246702015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cha EJ, Noh SJ, Kwon KS, Kim CY, Park BH,
Park HS, Lee H, Chung MJ, Kang MJ, Lee DG, et al: Expression of
DBC1 and SIRT1 is associated with poor prognosis of gastric
carcinoma. Clin Cancer Res. 15:4453–4459. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
You TK, Kim KM, Noh SJ, Bae JS, Jang KY,
Chung MJ, Moon WS, Kang MJ, Lee DG and Park HS: Expressions of
E-cadherin, Cortactin and MMP-9 in pseudoepitheliomatous
hyperplasia and squamous cell carcinoma of the head and neck: Their
relationships with clinicopathologic factors and prognostic
implication. Korean J Pathol. 46:331–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Noh SJ, Baek HA, Park HS, Jang KY, Moon
WS, Kang MJ, Lee DG, Kim MH, Lee JH and Chung MJ: Expression of
SIRT1 and cortactin is associated with progression of non-small
cell lung cancer. Pathol Res Pract. 209:365–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Luo J, Nikolaev AY, Imai S, Chen D, Su F,
Shiloh A, Guarente L and Gu W: Negative control of p53 by Sir2alpha
promotes cell survival under stress. Cell. 107:137–148. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Firestein R, Blander G, Michan S,
Oberdoerffer P, Ogino S, Campbell J, Bhimavarapu A, Luikenhuis S,
de Cabo R, Fuchs C, et al: The SIRT1 deacetylase suppresses
intestinal tumorigenesis and colon cancer growth. PLoS One.
3:e20202008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang RH, Zheng Y, Kim HS, Xu X, Cao L,
Luhasen T, Lee MH, Xiao C, Vassilopoulos A, Chen W, et al:
Interplay among BRCA1, SIRT1 and Survivin during BRCA1-associated
tumorigenesis. Mol Cell. 32:11–20. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Stunkel W, Peh BK, Tan YC, Nayagam VM,
Wang X, Salto-Tellez M, Ni B, Entzeroth M and Wood J: Function of
the SIRT1 protein deacetylase in cancer. Biotechnol J. 2:1360–1368.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen WY, Wang DH, Yen RC, Luo J, Gu W and
Baylin SB: Tumor suppressor HIC1 directly regulates SIRT1 to
modulate p53-dependent DNA-damage responses. Cell. 123:437–448.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jang KY, Noh SJ, Lehwald N, Tao GZ,
Bellovin DI, Park HS, Moon WS, Felsher DW and Sylvester KG: SIRT1
and c-Myc promote liver tumor cell survival and predict poor
survival of human hepatocellular carcinomas. PLoS One.
7:e451192012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang T, Rong N, Chen J, Zou C, Jing H,
Zhu X and Zhang W: SIRT1 expression is associated with the
chemotherapy response and prognosis of patients with advanced
NSCLC. PLoS One. 8:e791622013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jang KY, Kim KS, Hwang SH, Kwon KS, Kim
KR, Park HS, Park BH, Chung MJ, Kang MJ, Lee DG and Moon WS:
Expression and prognostic significance of SIRT1 in ovarian
epithelial tumours. Pathology. 41:366–371. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jang SH, Min KW, Paik SS and Jang KS: Loss
of SIRT1 histone deacetylase expression associates with tumour
progression in colorectal adenocarcinoma. J Clin Pathol.
65:735–739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jung W, Hong KD, Jung WY, Lee E, Shin BK,
Kim HK, Kim A and Kim BH: SIRT1 Expression is associated with good
prognosis in colorectal cancer. Korean J Pathol. 47:332–339. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sarhadi VK, Wikman H, Salmenkivi K, Kuosma
E, Sioris T, Salo J, Karjalainen A, Knuutila S and Anttila S:
Increased expression of high mobility group A proteins in lung
cancer. J Pathol. 209:206–212. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dixit D, Sharma V, Ghosh S, Mehta VS and
Sen E: Inhibition of Casein kinase-2 induces p53-dependent cell
cycle arrest and sensitizes glioblastoma cells to tumor necrosis
factor (TNFα)-induced apoptosis through SIRT1 inhibition. Cell
Death Dis. 3:e2712012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Peck B, Chen CY, Ho KK, Di Fruscia P,
Myatt SS, Coombes RC, Fuchter MJ, Hsiao CD and Lam EW: SIRT
inhibitors induce cell death and p53 acetylation through targeting
both SIRT1 and SIRT2. Mol Cancer Ther. 9:844–855. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kracikova M, Akiri G, George A,
Sachidanandam R and Aaronson SA: A threshold mechanism mediates p53
cell fate decision between growth arrest and apoptosis. Cell Death
Differ. 20:576–588. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zou T, Yang Y, Xia F, Huang A, Gao X, Fang
D, Xiong S and Zhang J: Resveratrol Inhibits CD4(+) T Cell
Activation by Enhancing the Expression and Activity of Sirt1. PLoS
One. 8:e751392013. View Article : Google Scholar : PubMed/NCBI
|