Introduction
Pancreatic cancer is one of the most common types of
human cancer, with a 5-year overall survival rate of only 5%
(1). Even among those patients who
undergo resection and have tumor-free margins, the 5-year survival
rate is only 10–25% (1). Despite the
fact that combined surgery, radiotherapy and chemotherapy have been
applied for >30 years, no significant improvement has been
obtained in the treatment of metastatic pancreatic cancer (2–4). As the
dysregulation of oncogenes or tumor suppressor genes has been
observed to play key roles in pancreatic cancer, the development of
potential molecular targets is a promising approach to achieve
effective therapies for pancreatic cancer (5).
The phosphoinositide 3-kinase (PI3K) signaling
pathway has been demonstrated to be frequently dysregulated in
various types of human cancer, including pancreatic cancer
(6–8).
Aberrant activation of PI3K signaling promotes cancer cell
proliferation, invasion and migration (9–11).
Phosphatase and tensin homolog (PTEN) is able to antagonize PI3K
signaling, thus acting as an inhibitor of the PI3K signaling
pathway (12). However, PTEN is
frequently mutated or its activity is downregulated in human cancer
(13,14).
Phosphatidylinositol-3,4,5-trisphosphate-dependent Rac exchanger
factor 2 (PREX2) is a regulator of the small guanosine
triphosphatase Rac, and has recently been reported to act as an
inhibitor of PTEN (15,16). PREX2 is able to inhibit the activity
of PTEN by directly binding to PTEN through its guanine nucleotide
exchange factor domains. Through inhibition of PTEN activity, PREX2
is capable of activating the downstream PI3K signaling pathway
(15,16). Indeed, it has been reported that
knockdown of PREX2 inhibits the invasion and clonogenicity of
gastric cancer cells via inhibition of PI3K signaling (17). In addition, silencing of PREX2 could
also induce cell cycle arrest at the G1-S phase and promote
apoptosis in gastric cancer cells (17). Since the dysregulation of the PI3K
signaling pathway has been demonstrated to be involved in multiple
types of human cancer, PREX2 may generally act as an oncogene in
tumorigenesis (16).
However, the detailed role of PREX2 in pancreatic
cancer has never been previously reported. Therefore, the present
study aimed to explore the role of PREX2 in the regulation of
pancreatic cancer cell invasion and migration, and to identify the
underlying molecular mechanisms.
Materials and methods
Reagents
Dulbecco's modified Eagle's medium (DMEM), fetal
bovine serum (FBS) and Lipofectamine 2000 were purchased from
Thermo Fisher Scientific, Inc. (Waltham, MA, USA). RNAeasy Mini kit
was purchased from Qiagen, Inc. (Valencia, CA, USA). PrimeScript™
RT reagent kit was purchased from Takara Bio, Inc. (Otsu, Japan).
iQ™ SYBR® Green supermix was purchased from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA). All antibodies were
purchased from Abcam (Cambridge, MA, USA). Enhanced
chemiluminescence (ECL) kit was purchased from Pierce (Thermo
Fisher Scientific, Inc.). Transwell chambers were purchased from
Corning Incorporated (Corning, NY, USA). PREX2 small interfering
(si)RNA was purchased from Santa Cruz Biotechnology, Inc. (Dallas,
TX, USA). PREX2 plasmid was purchased from DNASU Plasmid Repository
(Tempe, AZ, USA).
Tissue specimens
The present study was approved by the Ethics
Committee of Central South University (Changsha, China). Written
informed consent was obtained from each patient. A total of 10
primary pancreatic cancer tissues and their matched normal adjacent
specimens were collected between March and September 2013. The
histomorphology of all samples was confirmed at the Department of
Pathology of Xiangya Hospital of Central South University
(Changsha, China). Tissues were immediately snap frozen in liquid
nitrogen following surgical removal.
Cell culture
The human pancreatic cancer cell lines AsPC-1,
BxPC-3, PANC-1 and CFAPC-1, and the normal pancreatic epithelial
cell line HPC-Y5, were obtained from the Cell Bank of Central South
University (Changsha, China). Cells were cultured in DMEM with 10%
FBS at 37°C in a humidified incubator containing 5%
CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated using RNAeasy Mini kit,
according to the manufacturer's protocol. First-strand
complementary (c)DNA was generated using PrimeScrip™ RT reagent
kit, according to the manufacturer's protocol. PCR was conducted
using 2 µl cDNA, 0.2 µl sense and antisense primers, and iQ™
SYBR® Green supermix on a 7500 Fast Real Time PCR system
(Applied Biosystems, Thermo Fisher Scientific, Inc.). The PCR
parameters were as follows: 50°C for 2 min, 95°C for 2 min, and 40
cycles of 95°C for 15 sec and 60°C for 1 min. Glyceraldehyde
3-phosphate dehydrogenase (GAPDH) was used as an internal control.
Gene expression was determined by the 2−ΔΔCq method,
where ΔCq = (Cqgene-CqGAPDH) and Cq is the
threshold cycle. The specific primer pairs used were as follows:
PREX2 sense, 5′-TGGGAGGGGTCCAACATCA-3′ and anti-sense,
5′-TCTTCAACCGTCTGTGTTTTCTT-3′; and GAPDH sense,
5′-CTCCTCCTGTTCGACAGTCAGC-3′ and anti-sense,
5′-CCCAATACGACCAAATCCGTT-3′.
Transfection
Lipofectamine 2000 was used to perform cell
transfection following the manufacturer's protocol. Briefly, PREX2
siRNA, PREX2 plasmid and Lipofectamine 2000 were diluted with
serum-free medium. Diluted Lipofectamine 2000 was added into the
diluted PREX2 siRNA or PREX2 plasmid, and incubated for 20 min at
room temperature, prior to be added to the cell suspension. Then,
cells were incubated at 37°C with 5% CO2 for 6 h.
Subsequently, the medium in each well was replaced by DMEM with 10%
FBS, and cells were cultured for additional 24 h prior to be
subjected to the corresponding assays.
Western blotting
Tissues and cells were solubilized in cold
radioimmunoprecipitation assay lysis buffer. Proteins were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis, and then transferred onto a polyvinylidene
difluoride membrane. The membrane was incubated overnight at 4°C
with phosphate-buffered saline containing 5% milk. Subsequently,
the membrane was incubated with Tris-buffered saline containing 5%
milk at room temperature for 3 h, and then incubated with rabbit
anti-PREX2 monoclonal antibody (ab169027; 1:100), rabbit
anti-phosphorylated PTEN monoclonal antibody (ab109454; 1:50),
rabbit anti-PTEN monoclonal antibody (ab32199; 1:50), rabbit
anti-phosphorylated-AKT monoclonal antibody (ab81283; 1:50), rabbit
anti-AKT monoclonal antibody (ab32505; 1:50) or rabbit anti-GAPDH
monoclonal antibody (EPR16891; 1:100) at room temperature for 3 h,
followed by incubation with rabbit anti-mouse secondary antibody
(1:5,000; ab99697) at room temperature for 1 h, all from Abcam
(Cambridge, UK). An ECL kit was then used for chemiluminent
detection. Relative protein expression was analyzed by Image-Pro
Plus software version 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA), and represented as the density ratio vs. GAPDH.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
MTT assay was used to measure cell proliferation.
Cells in each group were cultured in 96-well plates, and 100 µl
fresh serum-free medium with 0.5 g/l MTT was added to each well.
Following incubation at 37°C for 6, 24, 48 and 72 h, the medium was
removed by aspiration, and 50 µl dimethyl sulfoxide was added to
each well. Upon incubation at 37°C for additional 10 min, the
absorbance at 492 nm of each sample was measured using a plate
reader.
Cell invasion assay
The invasive ability of PANC-1 cells was determined
in 24-well Transwell chambers, which had a layer of Matrigel. A
cell suspension was added in the upper chamber, while DMEM
containing 10% FBS was added into the lower chamber. Following
incubation for 24 h, non-invading cells and the matrix gel on the
interior of the inserts was removed using a cotton-tipped swab.
Invasive cells on the lower surface of the membrane were stained
with gentian violet, rinsed with water and air-dried. Five fields
were randomly selected, and the number of cells in those fields was
counted under a microscope.
Wound healing assay
Wound healing assay was performed to evaluate cell
migration. In brief, PANC-1 cells were cultured to full confluence.
Wounds of ~1-mm width were created with a plastic scriber, and the
cells were next washed and incubated in a serum-free medium. Upon
wounding for 24 h, cells were incubated in a medium containing 10%
FBS. Cultures at 0, 24 and 48 h were fixed and observed under a
microscope.
Statistical analysis
All data are presented as the mean ± standard
deviation. One-way analysis of variance was used to analyze
statistically the data by SPSS version 17 software (SPSS, Inc.,
Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
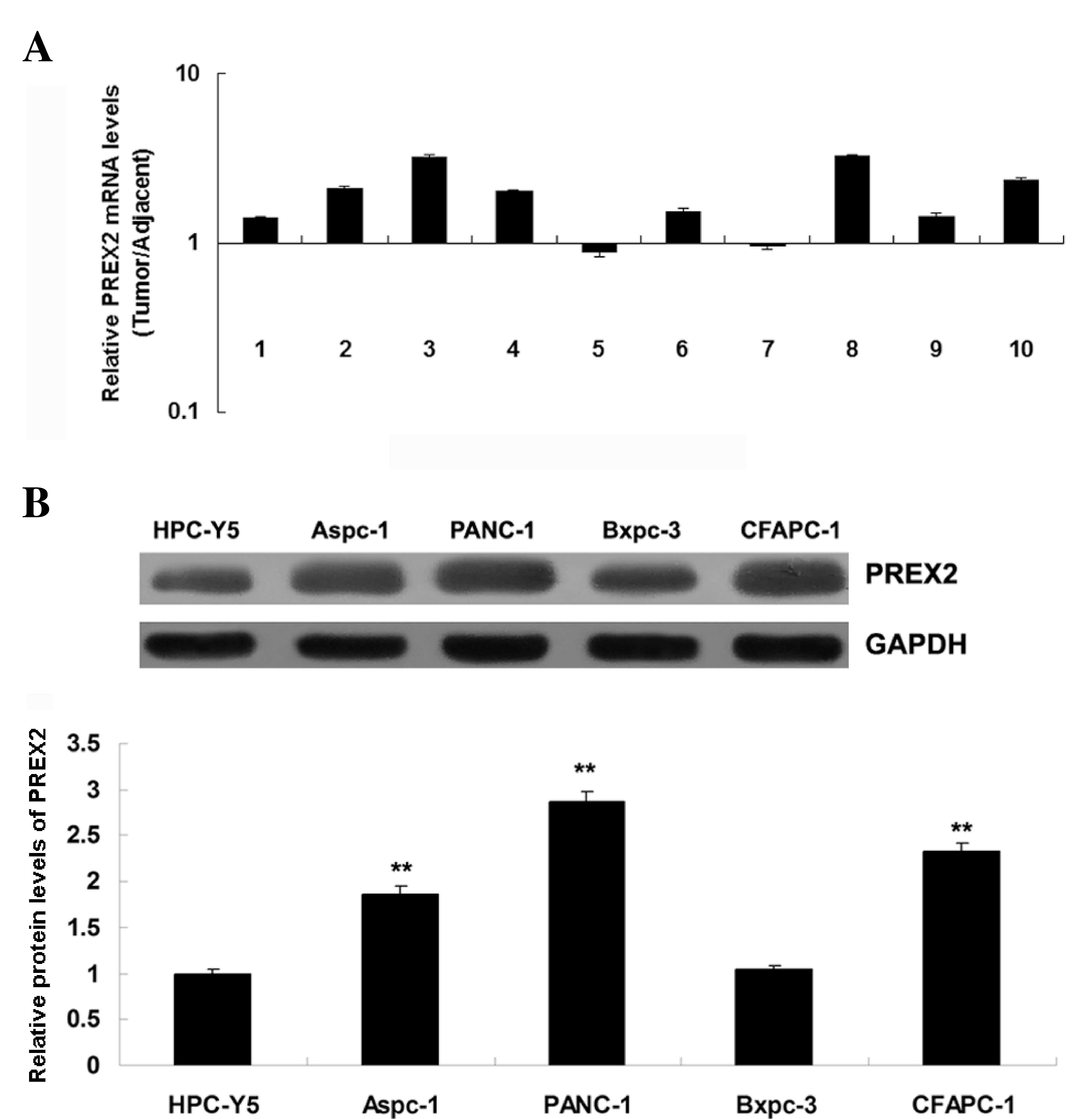
PREX2 was frequently upregulated in
pancreatic cancer
To study the role of PREX2 in pancreatic cancer, the
expression of PREX2 in pancreatic cancer tissues and their matched
adjacent normal tissues was determined by RT-qPCR. The data
revealed that PREX2 was frequently upregulated in pancreatic cancer
tissues, compared with the matched normal adjacent tissues
(Fig. 1A). These findings suggest
that dysregulation of PREX2 may play a role in pancreatic cancer.
Subsequently, the expression of PREX2 in several human pancreatic
cancer cell lines, including AsPC-1, BxPC-3, PANC-1 and CFAPC-1,
was examined. The normal pancreatic epithelial cell line HPC-Y5 was
used as control. Consistent with the tissue data, the expression of
PREX2 was also frequently upregulated in pancreatic cancer cell
lines, compared with normal pancreatic epithelial HPC-Y5 cells
(Fig. 1B). As PANC-1 cells exhibited
the most significant upregulation of PREX2 expression, this cell
line was used in the following in-vitro studies.
Role of PREX2 in promoting pancreatic
cancer cell proliferation
The role of PREX2 in the regulation of the
proliferation of pancreatic cancer cells was further investigated.
Firstly, pancreatic cancer PANC-1 cells were transfected with
pcDNA3.1-PREX2 plasmid (Addgene, Inc., Cambridge, MA, USA) or
PREX2-specifc siRNA to modulate the expression levels of PREX2.
Upon transfection, western blot assay was performed to determine
the expression of PREX2 in PANC-1 cells. As indicated in Fig. 2A, the protein levels of PREX2 were
significantly increased in PANC-1 cells following transfection with
pcDNA3.1-PREX2 plasmid, compared with the control group. By
contrast, the protein levels of PREX2 were notably decreased in
PANC-1 cells following transfection with PREX2-specific siRNA,
compared with the control group. Subsequently, MTT assay was
performed to determine the proliferation capacity of pancreatic
cancer PANC-1 cells following upregulation or downregulation of
PREX2. As represented in Fig. 2B,
PANC-1 cell proliferation was significantly upregulated following
overexpression of PREX2, while it significantly decreased upon
inhibition of PREX2, compared with the control group. Based on
these findings, it is possible to suggest that PREX2 plays a
promoting role in the regulation of pancreatic cancer cell
invasion.
Promoting role of PREX2 in pancreatic
cancer cell invasion
Transwell assay was performed to determine the
invasive capacity of pancreatic cancer PANC-1 cells following
upregulation or downregulation of PREX2. As indicated in Fig. 3, the invasive capacity of PANC-1 cells
was significantly upregulated following overexpression of PREX2,
while it significantly decreased upon inhibition of PREX2, compared
with the control group. These findings suggest that PREX2 plays a
promoting role in the regulation of pancreatic cancer cell
invasion.
Promoting role of PREX2 in pancreatic
cancer cell migration
Wound healing assay was performed to determine the
cell migratory capacity of PANC-1 cells following upregulation or
downregulation of PREX2. As represented in Fig. 4, the migratory capacity of PANC-1
cells was downregulated upon knockdown of PREX2, but it was
significantly upregulated following PREX2 upregulation, compared
with the control group. Therefore, PREX2 is able to promote
pancreatic cancer cell migration.
Effect of PREX2 on the activity of the
PI3K signaling pathway in pancreatic cancer cells
PREX2 has been previously reported to act as an
inhibitor of PTEN and an activator of the PI3K signaling pathway
(15). However, the effect of PREX2
on PTEN and PI3K signaling has never been reported in pancreatic
cancer. Therefore, the activity of PTEN and the PI3K signaling
pathway in pancreatic cancer cells following upregulation or
downregulation of PREX2 was further investigated in the present
study. AKT is a downstream effector of PI3K, and its
phosphorylation levels represent the activity of the PI3K signaling
pathway. (15) The present data
revealed that the phosphorylation levels of PTEN increased in
PANC-1 cells that overexpressed PREX2, indicating that the activity
of PTEN was reduced in these cells. Furthermore, the
phosphorylation levels of AKT were increased, indicating that the
activity of the PI3K signaling pathway was upregulated (Fig. 5). On the contrary, the inhibition of
PREX2 led to reduced phosphorylation levels of PTEN, indicating
that the activity of PTEN was upregulated. In addition, the
phosphorylation levels of AKT were reduced, which indicated that
the activity of the PI3K signaling pathway was decreased (Fig. 5). Based on these findings, it is
possible to suggest that PREX2 could activate the PI3K signaling
pathway via inhibition of PTEN activity in pancreatic cancer
cells.
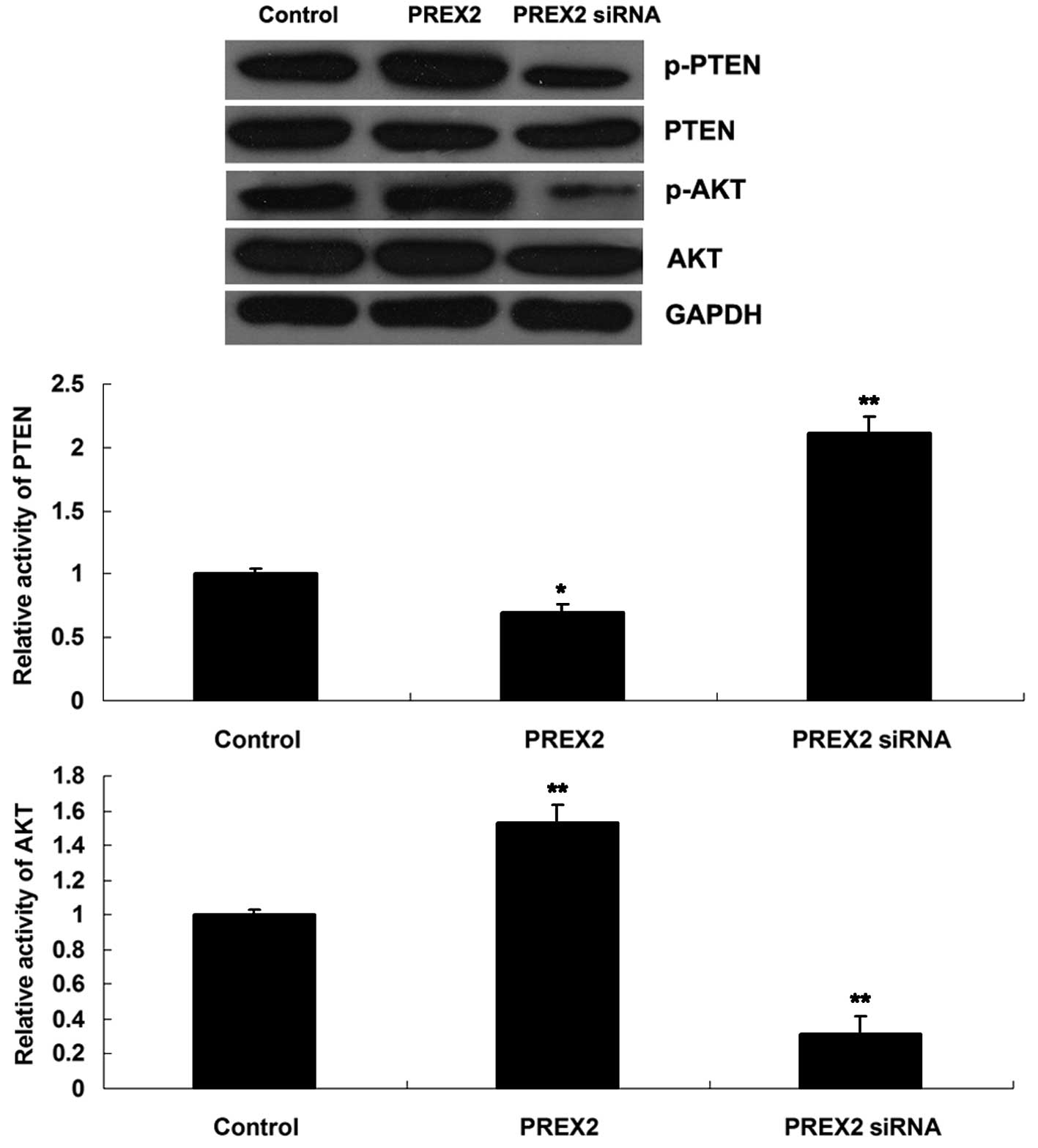 | Figure 5.Western blot assay was performed to
determine the expression of p-PTEN, PTEN, p-AKT and AKT in PANC-1
cells transfected with pcDNA3.1-PREX2 plasmid or PREX2-specific
small interfering RNA. Glyceraldehyde 3-phosphate dehydrogenase was
used as reference, and non-transfected PANC-1 cells were used as
control. *P<0.05 vs. control; **P<0.01 vs. control. PREX2,
phosphatidylinositol-3,4,5-trisphosphate-dependent Rac exchanger
factor 2; si, small interfering; GAPDH, glyceraldehyde 3-phosphate
dehydrogenase; PTEN, phosphatase and tensin homolog; p-,
phosphorylated. |
Discussion
PREX2 has been reported to be involved in several
types of human cancer, including melanoma, gastric cancer,
hepatocellular carcinoma and neuroblastoma (17–20).
However, the detailed role of PREX2 in pancreatic cancer has not
been previously reported. In the present study, the expression of
PREX2 was demonstrated to be significantly increased in pancreatic
cancer tissues compared with that in normal adjacent tissues. In
addition, PREX2 expression was also frequently downregulated in
pancreatic cancer cell lines compared with that in normal
pancreatic epithelial cells. Furthermore, PREX2 was demonstrated to
play a promoting role in the regulation of the proliferation,
invasion and migration of pancreatic cancer cells, probably at
least by suppressing the activity of PTEN and thus upregulating the
activity of the PI3K signaling pathway.
Fine et al (21) revealed for the first time the
inhibitory effect of PREX2 on PTEN. The authors noticed that PREX2
was more abundant in human cancer cells compared with normal cells,
and was significantly upregulated in tumors with wild-type PTEN
that expressed an activated mutant of
phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit
alpha, which encoded the p110 subunit of PI3Kα. In the present
study, the expression of PREX2 was demonstrated to be frequently
upregulated in pancreatic cancer tissues and cell lines, suggesting
that PREX2 may be implicated in the tumorigenesis of pancreatic
cancer. Fine et al (21)
further demonstrated that only in the presence of PTEN, PREX2 could
inhibit the lipid phosphatase activity of PTEN and stimulate the
PI3K signaling pathway, thus suggesting that PREX2 is a component
of the PI3K signaling pathway able to antagonize PTEN in cancer
cells. In addition, P-REX2 has been observed to stimulate cell
growth and growth factor-independent cell proliferation and
transformation, and knockdown of PREX2 led to reduced growth of
human cells with intact PTEN and reduced levels of phosphorylated
AKT (21). In the present study, the
downregulation of PREX2 notably suppressed pancreatic cancer cell
proliferation.
Recently, the overexpression of P-Rex proteins has
been linked to poor patient outcome in breast cancer, since it may
facilitate metastatic dissemination of prostate cancer cells
(16). Whole-genome sequencing
identified PREX2 as a significantly mutated gene in melanoma,
suggesting that PREX2 may also be involved in the development and
progression of melanoma (18).
Additionally, Chen et al (20)
demonstrated that microRNA (miR)-338-3p could inhibit neuroblastoma
cell proliferation, migration and invasion, and PREX2 was
identified as a direct target of miR-338-3p. Knockdown of PREX2
inhibits cell proliferation, migration and invasion through the
PTEN/AKT signaling pathway. Similarly, Guo et al (17) reported that miR-338-3p enhanced
gastric cancer progression through PTEN-AKT signaling by targeting
PREX2 in gastric cancer cells. However, the detailed role of PREX2
in the regulation of pancreatic cancer metastasis has not been
previously reported. The present study demonstrated that PREX2
played a promoting role in the regulation of pancreatic cell
proliferation and enhanced pancreatic cell migration and invasion,
suggesting that the dysregulation of PREX2 may be involved in the
metastasis of pancreatic cancer. Indeed, silencing of the PI3K/AKT
signaling pathway has been observed to effectively inhibit the
metastasis of pancreatic cancer (22,23).
In conclusion, the results of the present study
revealed an oncogenic role of PREX2 in pancreatic cancer cells, and
suggested that PREX2 may be a potential therapeutic target for
pancreatic cancer.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Páez D, Labonte MJ and Lenz HJ: Pancreatic
cancer: Medical management (novel chemotherapeutics). Gastroenterol
Clin North Am. 41:189–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hidalgo M, Cascinu S, Kleeff J, Labianca
R, Löhr JM, Neoptolemos J, Real FX, Van Laethem JL and Heinemann V:
Addressing the challenges of pancreatic cancer: Future directions
for improving outcomes. Pancreatology. 15:8–18. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Poruk KE, Firpo MA and Mulvihill SJ:
Screening for pancreatic cancer. Adv Surg. 48:115–136. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eser S, Schnieke A, Schneider G and Saur
D: Oncogenic KRAS signalling in pancreatic cancer. Br J Cancer.
111:817–822. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun C, Rosendahl AH, Andersson R, Wu D and
Wang X: The role of phosphatidylinositol 3-kinase signaling
pathways in pancreatic cancer. Pancreatology. 11:252–260. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dimitrova V and Arcaro A: Targeting the
PI3K/AKT/mTOR signaling pathway in medulloblastoma. Curr Mol Med.
15:82–93. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou SL, Zhou ZJ, Hu ZQ, Li X, Huang XW,
Wang Z, Fan J, Dai Z and Zhou J: CXCR2/CXCL5 axis contributes to
epithelial-mesenchymal transition of HCC cells through activating
PI3K/AKT/GSK-3β/Snail signaling. Cancer Lett. 358:124–135. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Falasca M, Selvaggi F, Buus R, Sulpizio S
and Edling CE: Targeting phosphoinositide 3-kinase pathways in
pancreatic cancer-from molecular signalling to clinical trials.
Anticancer Agents Med Chem. 11:455–463. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji BC, Hsiao YP, Tsai CH, Chang SJ, Hsu
SC, Liu HC, Huang YP, Lien JC and Chung JG: Cantharidin impairs
cell migration and invasion of A375.S2 human melanoma cells by
suppressing MMP-2 and −9 through PI3K/NF-κB signaling pathways.
Anticancer Res. 35:729–738. 2015.PubMed/NCBI
|
|
11
|
Massimiani M, Vecchione L, Piccirilli D,
Spitalieri P, Amati F, Salvi S, Ferrazzani S, Stuhlmann H and
Campagnolo L: Epidermal growth factor-like domain 7 promotes
migration and invasion of human trophoblast cells through
activation of MAPK, PI3K and NOTCH signaling pathways. Mol Hum
Reprod. 21:435–451. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Carnero A and Paramio JM: The
PTEN/PI3K/AKT pathway in vivo, cancer mouse models. Front Oncol.
4:2522014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu J, Li Z, Wang J, Chen H and Fang JY:
Combined PTEN mutation and protein expression associate with
overall and disease-free survival of glioblastoma patients. Transl
Oncol. 7:196–205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hühns M, Salem T, Schneider B, Krohn M,
Linnebacher M and Prall F: PTEN mutation, loss of heterozygosity,
promoter methylation and expression in colorectal carcinoma: Two
hits on the gene? Oncol Rep. 31:2236–2244. 2014.PubMed/NCBI
|
|
15
|
Rosenfeldt H, Vazquez-Prado J and Gutkind
JS: P-REX2, a novel PI-3-kinase sensitive Rac exchange factor. FEBS
Lett. 572:167–171. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pandiella A and Montero JC: Molecular
pathways: P-Rex in cancer. Clin Cancer Res. 19:4564–4569. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guo B, Liu L, Yao J, Ma R, Chang D, Li Z,
Song T and Huang C: miR-338-3p suppresses gastric cancer
progression through a PTEN-AKT axis by targeting P-REX2a. Mol
Cancer Res. 12:313–321. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Berger MF, Hodis E, Heffernan TP, Deribe
YL, Lawrence MS, Protopopov A, Ivanova E, Watson IR, Nickerson E,
Ghosh P, et al: Melanoma genome sequencing reveals frequent PREX2
mutations. Nature. 485:502–506. 2012.PubMed/NCBI
|
|
19
|
Lan X, Xiao F, Ding Q, Liu J, Liu J, Li J,
Zhang J and Tian DA: The effect of CXCL9 on the invasion ability of
hepatocellular carcinoma through up-regulation of PREX2. J Mol
Histol. 45:689–696. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen X, Pan M, Han L, Lu H, Hao X and Dong
Q: miR-338-3p suppresses neuroblastoma proliferation, invasion and
migration through targeting PREX2a. FEBS Lett. 587:3729–3737. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fine B, Hodakoski C, Koujak S, Su T, Saal
LH, Maurer M, Hopkins B, Keniry M, Sulis ML, Mense S, et al:
Activation of the PI3K pathway in cancer through inhibition of PTEN
by exchange factor P-REX2a. Science. 325:1261–1265. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao H, Wang L, Wei R, Xiu D, Tao M, Ke J,
Liu Y, Yang J and Hong T: Activation of glucagon-like peptide-1
receptor inhibits tumourigenicity and metastasis of human
pancreatic cancer cells via PI3K/AKT pathway. Diabetes Obes Metab.
16:850–860. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Missiaglia E, Dalai I, Barbi S, Beghelli
S, Falconi M, della Peruta M, Piemonti L, Capurso G, Di Florio A,
delle Fave G, et al: Pancreatic endocrine tumors: Expression
profiling evidences a role for AKT-mTOR pathway. J Clin Oncol.
28:245–255. 2010. View Article : Google Scholar : PubMed/NCBI
|