Introduction
Transcriptional targeting in gene therapy is an
approach based on the use of tissue-specific or tumor-specific
promoters (TSPs) to direct the expression of therapeutic genes
specifically to a tumor (1). It is
assumed that it must meet the requirements of transgene expression
in tumor tissues but not in normal tissues. This combination of
‘tumor on’ and ‘normal tissue off’ profile can result in an
increase in the therapeutic index and limit the toxicity of vectors
and transgenes in vivo. Numerous promoters have already been
evaluated for transcriptional targeting in cancer gene therapy [for
reviews, see (2–4)], such as the α-fetoprotein promoter for
hepatic carcinoma, tyrosinase gene promoter for melanoma,
prostate-specific antigen promoter for prostate cancer,
cyclooxygenase-2 promoter for gastrointestinal cancer, midkine
promoter for Wilms' tumor or neuroblastoma, chemokine (C-X-C motif)
receptor 4 promoter for breast cancer and melanoma,
hypoxia-inducible promoter for hypoxic tumor-targeting gene
therapy, promoter of the carcinoembryonic antigen gene (5), and numerous others. However, many of the
gene promoters used for this goal have been shown to be abundantly
expressed in a variety of normal tissues (4), such that their usage also affects normal
host cells, thus increasing the risk of unwanted side effects. TSPs
that are expressed in a wide variety of tumors and have low
expression in normal tissue are highly desirable to meet gene
therapy demands. Two gene promoters frequently used for the gene
therapeutic purposes appear to most completely satisfy the
requirements: The human telomerase reverse transcriptase
(hTERT) promoter, and the promoter driving the expression of
BIRC5, encoding the apoptosis inhibitor survivin.
Survivin is one of the central players of cancer
development (1,6–8), and the
BIRC5 gene (9) promoter is
also active in wide spectrum of cancer cells (10–13). In
normal tissues, survivin expression is found during embryonic and
fetal development but is largely undetectable in terminally
differentiated adult tissues (14).
Although growing evidence indicates that survivin is expressed in
primitive hematopoietic cells, T lymphocytes, polymorphonuclear
neutrophils and vascular endothelial cells, and that it may
regulate their proliferation or survival, targeted anti-survivin
therapies have exhibited efficacy without overt toxicity in
numerous preclinical animal models (14). It has been reported that the
functional promoter region of the BIRC5 gene spans 1,456 bp
upstream of the transcription start site (15) and continues to ~40 nucleotides (nt)
downstream of the transcription start point. The BIRC5 gene
promoter is highly tumor-specific and works in a great majority
(80–85%) of tumors (9,14,16), thus
presenting the possibility of its general utilization in cancer
treatment (17). However, in common
with the majority of TSPs (17–19), it is
rather weak in comparison with such promoters as constitutive
cytomegalovirus (CMV) and Simian vacuolating virus 40 (SV40)
promoters with their enhancers (when used as isolated promoters in
the context of a vector to drive transgene expression).
BIRC5 promoter activity in different cell lines comprises
0.3–16% of that of a strong constitutive CMV promoter (4,13,18,20).
Therefore, it is highly desirable to reconstruct the promoter so
that it acquires a higher activity level while remaining strictly
cancer-specific.
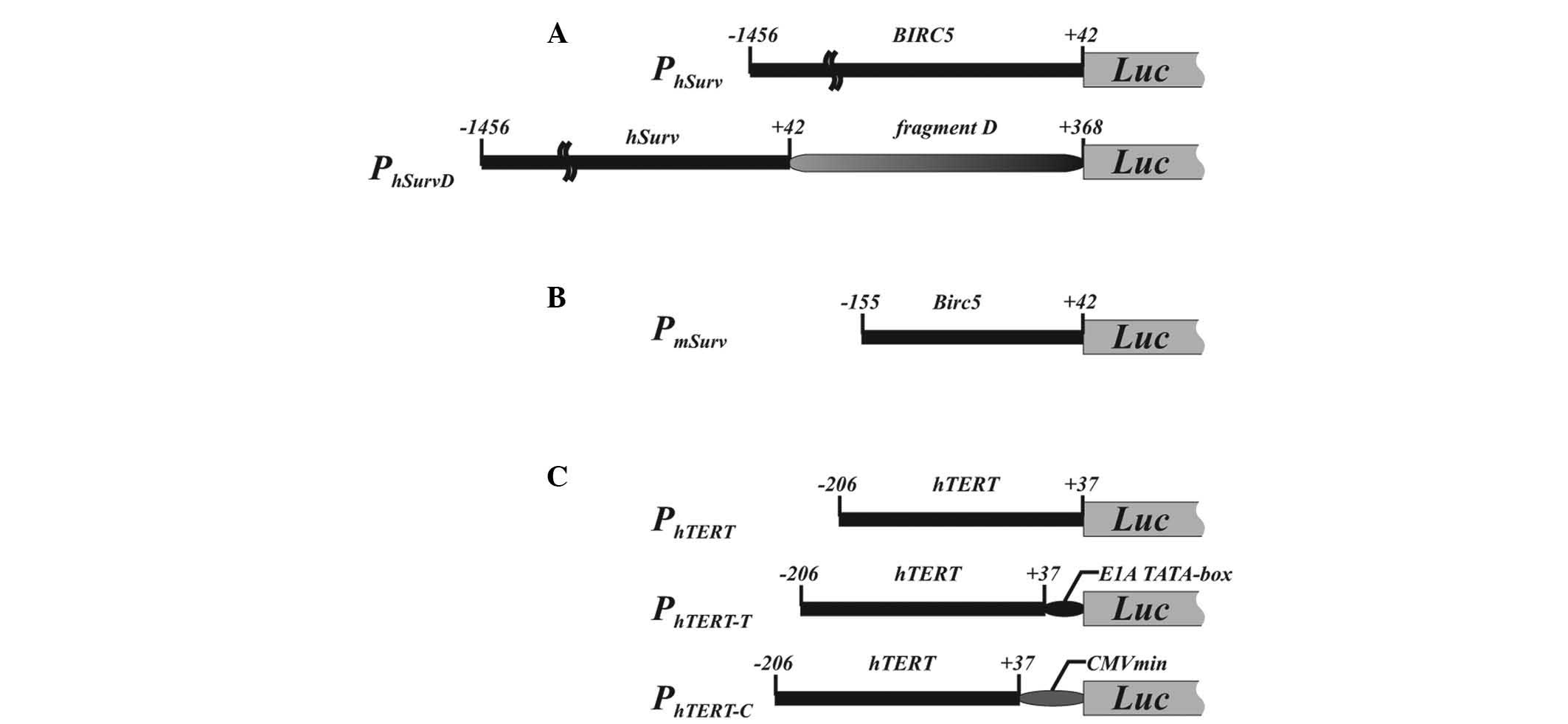
Recently, we constructed a modified version of the
human BIRC5 promoter (PhSurv) by inserting
fragment D (326 nt), consisting of the first exon and a part of the
first intron of the human BIRC5 gene, which contains a
CpG-island and possesses enhancer-like activity (Fig. 1A). This modified promoter, referred to
as PhSurvD, exhibited enhanced transcriptional activity
in the majority of p53-negative lung cancer cell lines (21). It must be noted that shortening of the
human BIRC5 promoter fragment beyond 1.4 kb upstream of the
transcription start site resulted in decreased promoter activity
(15). At the same time, the overall
expression cassette length could be a concern for gene therapy
application due to the limited capacity of certain vectors used for
delivery, such as that of adenoviral vectors (22). Therefore, short promoters that are
able to retain tumor specificity and possess maximal activity are
preferred. A short fragment of the murine Birc5 gene
promoter spanning nt −155 to +42 relative to the transcription
start site [corresponding to nt −195 to +2 as numbered in (23,24)]
(Fig. 1B) was reported to be highly
active in an in vitro reporter gene analysis (23) and demonstrated higher transgene
expression in murine cells than that observed in human cells with
the human BIRC5 promoter (20), thus providing another option for
improved survivin promoter-driven expression in tumor cells.
Telomerase activity is critical for the acquisition
of immortality by cancer cells through maintaining telomere length.
In humans, telomerase activation in cancer cells is achieved due to
restoring the telomerase reverse transcriptase subunit of
telomerase; this relies on transcriptional activation of the
hTERT gene, which is silent in somatic human tissues.
hTERT expression has been observed in various types of human
cancer (25–27), with human telomerase being highly
active in >85% of primary cancers, regardless of their tissue
origins, but not in normal differentiated human cells (26,28–31). It
has been revealed and later confirmed in numerous studies that a
~200-bp fragment of the hTERT 5′-flanking region is enough
to function as a promoter for transcriptional activation in cancer
cells while maintaining cancer cell specificity (29). The hTERT promoter lacks TATA-
and CAAT-boxes (29,32,33). In an
attempt to increase the promoter activity critical for improved
transgene expression, the promoter was modified to increase its
activity without appreciable loss of cancer cell specificity. One
of the reported modifications consisted of the joining of
PhTERT with a minimal CMV promoter (Fig. 1C) (34).
In another modification, a TATA-box derived from the adenoviral E1A
promoter was added to hTERT promoter (Fig. 1C) (35).
These two modifications were reported to improve the promoter
performance without compromising its cancer cell specificity.
However, no direct comparison of these two modifications has been
previously conducted to identify which one is preferable to drive
transgene expression in cancer cells.
Therefore, the BIRC5 and hTERT
promoters and their modified versions may be good candidates for
use as a TSPs in gene therapy approaches to treat cancers, with a
low potential toxicity for normal host tissue in vivo.
However, despite the widespread use of the two promoters in
experimental studies of potential gene therapy schemes, to the best
of our knowledge there is no comprehensive comparative analysis
that would allow a comparison of the advantages and disadvantages
of each promoter and determine a rational basis for selecting the
optimal promoter for gene therapy of a particular tumor. Such a
comparison would also be of use in the creation of more universal
synthetic promoters bearing combinations of elements from these two
promoters. In the present study, such a comparison was conducted,
revealing that activities of hTERT- and BIRC5-based
promoters are, in many cases, complementary to one another, such
that if one of them fails to support efficient therapeutic gene
expression in a tumor, the other may be used instead.
Materials and methods
Cell lines
Human HT1080 fibrosarcoma (#CCL-121; ATCC, Manassas,
VA, USA), Calu-I epidermoid carcinoma of lung (#93120818; ECACC,
Porton Down, UK), NCI-H1299 non-small cell lung carcinoma
(#CRL-5803; ATCC), AsPC-1 adenocarcinoma of the pancreas
(#CRL-1682; ATCC), Panc-1 ductal carcinoma of the pancreas
(#CRL-1469; ATCC) and HCT116 colorectal carcinoma (#CCL-247; ATCC)
cell lines were cultured in HyClone Dulbecco's modified Eagle's
medium (DMEM)/F12 (1:1) (GE Healthcare Life Sciences, Logan, UT,
USA) supplemented with 10% fetal bovine serum (FBS) (HyClone; GE
Healthcare Life Sciences), penicillin (100 U/ml) and streptomycin
(100 µg/ml). T47D breast ductal carcinoma (#HTB-133; ATCC) and A431
epidermal squamous carcinoma (#CRL-1555; ATCC) cells were cultured
in HyClone DMEM supplemented with 10% FBS, penicillin (100 U/ml)
and streptomycin (100 µg/ml). The human melanoma cell lines MelCher
and MelKor (36) were cultured in
HyClone RPMI-1640 medium (GE Healthcare Life Sciences) supplemented
with 10% FBS, penicillin (100 U/ml) and streptomycin (100 µg/ml).
Cells were transfected using Unifectin-56 transfection reagent
(Rusbiolink, Moscow, Russia) (HCT116, HT1080, Panc-1, Calu-I,
NCI-H1299 and MelKor cell lines), or Lipofectamine 2000 (Thermo
Fisher Scientific, Inc., Eugene, OR, USA) (AsPC1, MelCher, A431 and
T47D cell lines).
Plasmids
The human BIRC5 promoter fragment nt −1,456 to +42
(PhSurv; nt 76,208,870 to 76,210,367; GenBank accession no.
NC_000017.10) was amplified on a template of human genomic DNA with
primers 5′-aga tct aaa tct ggg tga agg gta tat gagt-3′ and 5′-aag
ctt cgc gat tca aat ctg gcggt-3′ and cloned into the pGL3-Basic
vector (Promega Corporation, Madison, WI, USA) to control firefly
luciferase reporter gene expression (Fig.
1A).
Fragment D (nt +43 to +368 of the human BIRC5
gene) was amplified on a template of human genomic DNA with primers
5′-cccgggacccgttggcagaggtggcggcggcggc(a>t)t ggg-3′ [with the
A>T substitution (indicated) in order to replace the ATG
translation initiation codon of the BIRC5 gene by a TTG
triplet] and 5′-taa gct tcc tcg atg ggg aca aag cag-3′, and cloned
at the 3′-end of PhSurv (promoter PhSurvD).
The orientation of fragment D in the PhSurvD coincided
with that in human genomic DNA (Fig.
1A).
The mouse Birc5 gene promoter fragment nt
−155 to +42 (PmSurv; nt 117,710,470 to 117,710,664;
GenBank accession no. NC_000077.5) was amplified on the template of
mouse genomic DNA with primers 5′-aga tct cca cgc cca caa ggc cag
gc-3′ and 5′-aag ctt atg atg gcg tca cca caacc-3′, and cloned into
the pGL3-Basic vector (Fig. 1B).
The hTERT promoter fragment nt −206 to +37
(PhTERT; nt 110,006 to 11,248; GenBank accession no.
AF128893) was amplified on the template of human peripheral blood
lymphocyte genomic DNA and cloned into pGL3-Basic vector to place
the firefly luciferase reporter gene under the control of
PhTERT (Fig. 1C). To
construct a hybrid PhTERT promoter with a synthetic
TATA-box (PhTERT-T promoter), complementary
oligonucleotides with an adenoviral E1A TATA-box sequence (5′-aat
tcg tgt agt gta ttt ata ccc ggt gag tag atc tg-3′ and 5′-gat cca
gat cta ctc acc ggg tat aaa tac act aca cg-3′) (35) were annealed and linked to the 3′-end
of PhTERT. The resulting PhTERT-T fragment
was cloned into pGL3-Basic vector to drive firefly luciferase
reporter gene expression (Fig. 1C).
Hybrid telomerase reverse transcriptase promoter
(PhTERT-C) with a minimal CMV promoter derived from the
pTRE-Tight plasmid (Clontech Laboratories, Inc., Mountain View, CA,
USA) was constructed in a similar way with the 69-nt minimal major
immediate early CMV promoter fragment amplified with primers 5′-atg
aat tcg gta ggc gtg tac ggt ggg ag-3′ and 5′-atg gat cca gat ctc
cag gcg atc tga cgg ttc-3′ on the template of the pTRE-Tight vector
(Fig. 1C).
Luciferase reporter assay
Cells were transfected with a mixture of firefly
reporter plasmid and a pRL-CMV (Promega Corporation) plasmid
(encoding a Renilla luciferase reporter gene under the
control of CMV immediate early enhancer/promoter) in a ratio
selected for each cell line to result in optimal signals for each
reporter. Cells in three wells were transfected in parallel for
each plasmid combination. Along with plasmids with firefly reporter
gene under the control of the studied promoters, a pGL3-Control
plasmid (Promega Corporation) with a firefly luciferase reporter
gene under the control of the SV40 promoter and enhancer sequences,
and a promoter-less pGL3-Basic plasmid (Promega Corporation) were
used. Luciferase activities were quantified 2 days after
transfection either with Dual-Glo™ Luciferase Assay System or with
Dual-Luciferase® Reporter Assay System (both from
Promega Corporation) depending on the cell line transfection
efficiency. Firefly luciferase activity was normalized to
Renilla luciferase activity and then the mean ± standard
deviation (SD) values were calculated from the values for three
analyzed wells for each experimental point. The data presented are
the mean± SD following the subtraction of basal activity of the
promoter-less reporter (plasmid pGL3-Basic). Values were considered
statistically significant if the two-tailed P-value (unpaired
t-test) was <0.05.
Relative transcript level
determination
Total RNA was isolated from cells with RNeasy Mini
RNA Kit (Qiagen GmbH, Hilden, Germany). Reverse transcription (RT)
was performed on a template of 1 µg of RNA with random hexamer
primers and SuperScript III reverse transcriptase (Invitrogen;
Thermo Fisher Scientific, Carlsbad, CA, USA). RT reaction products
were diluted to give approximately equal amplification of
GAPDH transcript across the samples. Polymerase chain
reaction (PCR) was conducted in quadruplicates for each transcript
analyzed in each cell line. The 20-µl reaction mixture contained 2
µl of 10X amplification buffer (100 mM Tris-HCl, pH 8.3, and 500 mM
KCl), 0.8 µl of 25 mM magnesium chloride, 0.4 µl of 10 mM dNTP mix,
0.2 µl of each sense and antisense primer at 10 optical units/ml,
0.2 µl of Taq DNA polymerase (5 U/ml) and template. The primer
pairs used were 5′-gaa ggt gaa ggt cgg agtca-3′ and 5′-ttc aca ccc
atg acg aacat-3′ for GAPDH; 5′-cgg aag agt gtc tgg agcaa-3′
and 5′-gga tga agc gga gtc tgga-3′ for hTERT; and 5′-acc gca
tct cta cat tcaag-3′ and 5′-gga ata aac cct gga agtgg-3′ for
BIRC5. Cycling conditions following initial heating at 94°C
for 2 min were 94°C for 30 sec; 61°C (GAPDH) or 63°C
(hTERT) or 60°C (BIRC5) for 1 min; and 72°C for 1
min. The number of cycles was 15, 18, 21 and 24 for GAPDH;
25, 29, 33 and 37 for hTERT; and 21, 25, 29 and 33 for
BIRC5. Amplification products (402 bp for GAPDH, 145
bp for hTERT and 438 nt for major BIRC5 transcript isoform)
were resolved on an agarose gel containing ethidium bromide. An
O'GeneRuler 50 bp DNA Ladder (Fermentas; Thermo Fisher Scientific,
Inc.) was used to monitor amplification product lengths. Gel images
were acquired with ChemiDoc XRS Documentation system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) and band intensities were
quantified with Quantity One 1-D Analysis Software (version 4.6.2;
Bio-Rad Laboratories, Inc.). Relative hTERT and BIRC5
transcript levels were determined using the band intensities at
cycle numbers before reactions reached saturation by the
amplification products (typically at 29 and 33 cycles for
BIRC5 and hTERT, respectively).
Statistical analysis
To determine significant differences between two
values, an unpaired t-test was used to calculate two-tailed
P-values using GraphPad QuickCalcs online tool (www.graphpad.com/quickcalcs/ttest1/).
P<0.05 indicated a statistically significant difference. To
evaluate correlation between two datasets, r2
coefficient of correlation was calculated using the least squares
method in Microsoft Excel 2011 for Mac (Microsoft Corporation,
Redmond, WA, USA).
Results
Classification of cell lines according
to the efficiency and pattern of various promoter activities
PhSurv and PhTERT promoter activities were assessed
in 10 cancer cell lines of various origins using firefly luciferase
reporter gene and Renilla luciferase expressed under the
control of a CMV promoter as a reference for normalization. For an
appropriate comparison of activities of the promoters in different
tumor cell lines, a proper reference control is necessary. We
observed that the activity of the SV40 promoter and enhancer
sequences, which are supposed to be constitutive, varied relative
to the CMV promoter in a range of up to ~100 times for 5 cell lines
investigated (data not shown). The observed difference indicates
that SV40 promoter and enhancer sequences or/and CMV promoter
activity cannot be used as a reference control. The possibility
that the observed variations were due to the variability in
activity of the CMV promoter driving Renilla luciferase
expression used for firefly luciferase activity normalization could
not be excluded. Thus an adequate comparative analysis was possible
only for different promoters for a particular cell line, and not
across different cells.
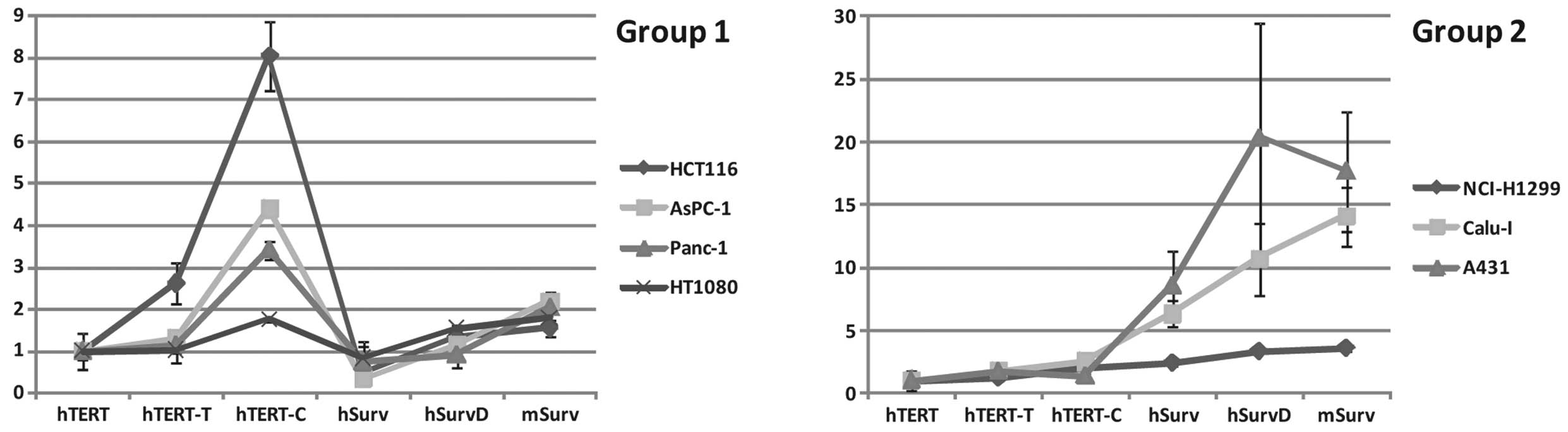
The studied tumor cell lines could be classified
based on PhTERT- or PhSurv-driven promoter
expression preference. According to this classification, there are
PhTERT expressors (MelKor, HCT116, AsPC-1),
PhSurv expressors (Calu-I, NCI-H1299, A431, MelCher),
and cells with similar promoter activities (HT1080, Panc-1, T47D)
(Table I). Among the cell lines
analyzed, PhTERT activity relative to the activity of
PhSurv ranged from 0.17 in A431 cells to 8.56 in MelKor
cells, giving a 50-fold difference between PhTERT or
PhSurv activities in different tumor cell lines. This
observation clearly indicates the necessity for choosing between
the two promoters for efficient transgene expression in a given
tumor type.
 | Table I.Promoter activities in various cell
lines. |
Table I.
Promoter activities in various cell
lines.
|
| Promoter
activity |
|---|
|
|
|
|---|
| Cell line |
PhTERT |
PhTERT-T |
PhTERT-C |
PhSurv |
PhSurvD |
PmSurv |
|---|
| HCT116 |
1.00±0.17 |
2.63±0.48a |
8.01±0.82a,b |
0.48±0.26a |
1.34±0.23c |
1.56±0.19c |
| AsPC-1 |
1.00±0.54 |
1.32±0.15 |
4.40±0.49a,b |
0.33±0.04a |
1.16±0.11c |
2.19±0.11c,d |
| Panc-1 |
1.00±0.43 |
1.15±0.17 |
3.41±0.21a,b |
0.73±0.49 |
0.92±0.31 |
2.03±0.39c,d |
| HT1080 |
1.00±0.10 |
1.02±0.28 |
1.74±0.04a,b |
0.83±0.31 |
1.55±0.08c |
1.80±0.22c |
| Calu-I |
1.00±0.81 |
1.76±0.31 |
2.51±0.57a |
6.31±1.06a |
10.67±2.82 |
14.06±2.32c |
| NCI-H1299 |
1.00±0.16 |
1.15±0.09 |
1.97±0.07a,b |
2.40±0.24a |
3.34±0.07c |
3.60±0.26c |
| A431 |
1.00±0.22 |
1.68±0.34a |
1.33±0.14 |
8.56±2.75a |
20.34±9.08 |
17.66±4.78c |
| MelCher |
1.00±0.61 |
0.95±0.56 |
11.38±3.73a,b |
3.85±2.67 |
7.99±2.51 |
3.97±1.39 |
| MelKor |
1.00±0.07 |
2.20±0.06a |
1.47±0.04a |
0.17±0.01a |
0.73±0.08c |
1.17±0.10c,d |
| T47D |
1.00±0.08 |
0.74±0.04a |
1.42±0.06a,b |
0.72±0.09a |
2.24±0.18c |
2.79±0.13c,d |
With regard to relative activities of various
promoter variants used in the study, two different patterns can be
distinguished. Panc-1, AsPC-1, HCT116 and HT1080 cell lines (‘Group
1’) had similar patterns of expression, with approximately the same
(with 3-fold difference in range) activities of PhTERT
or PhSurv which were significantly enhanced by
modification of the PhTERT promoter (particularly
PhTERT-C) and when using other variants of the survivin
promoter series, particularly PmSurv (Fig. 2). NCI-H1299, Calu-I and A431 (‘Group
2’) also had similar patterns of expression, with PhSurv
promoter activity prevailing over the activities of
PhTERT and its derivatives, and further potentiation
observed for PhSurvD and PmSurv (Group 2 in
Fig. 2). The rest of the cell lines
assayed (MelCher, MelKor and T47D) showed distinct and individual
patterns of promoter variant expression. Clustering of cell lines
on the basis of activities of PhTERT, PhSurv,
PmSurv promoters and their variants may reflect common
transcription factor repertoires for clustered cell lines and
different repertoires between different clusters.
Comparison of modified and wild type
promoter efficiencies
The data obtained demonstrate that modification of
the PhTERT promoter with the synthetic TATA-box or with the minimal
CMV promoter improves its performance in tumor cells. The
CMV-modified promoter was more active in 9 out of 10 cell lines,
while the TATA-modified promoter was more active in 3 out of 10
cell lines; in 1 cell line, the TATA-modified promoter showed
slightly lower activity. Importantly, the CMV-modified promoter was
more potent compared to the TATA-modified promoter in 7 out of 10
cell lines. Therefore, it is possible to conclude that modification
with the minimal CMV promoter is generally preferable to promoters
modified with a TATA-box. Modification of PhTERT with the minimal
CMV promoter preserves the intrinsic tumor specificity of the
hTERT promoter (34);
therefore, PhTERT-C appears to be more preferable than the other
promoters in this series for the goals of transcriptional targeting
of tumor cells.
The human PhSurv promoter was also
compared with its modified version, PhSurvD, and mouse
counterpart, PmSurv. The PhSurvD promoter had
higher activity in 6 out of 10 cell lines. The mouse promoter was
generally more active in human tumor cell lines compared to the
human promoter (in 9 out of 10 cell lines). Finally, the
PmSurv promoter was more active than the
PhSurvD promoter in 4 cell lines. In none of the cell
lines did the mouse promoter perform worse than human promoter or
its modified version. However the specificity of the mouse promoter
toward human tumor cells must be investigated further before its
practical utilization to drive therapeutic transgene
expression.
Transgene expression levels directed
by the studied promoters are not associated with levels of
endogenous hTERT and BIRC5 transcripts
According to the present results, the relative
activities of hTERT and survivin gene promoters may vary
broadly between tumor cell lines. Thus, we investigated whether
relative levels of endogenous hTERT or BIRC5
transcripts could be used to predict the relative activities of the
corresponding out-of-gene promoters in gene therapeutic
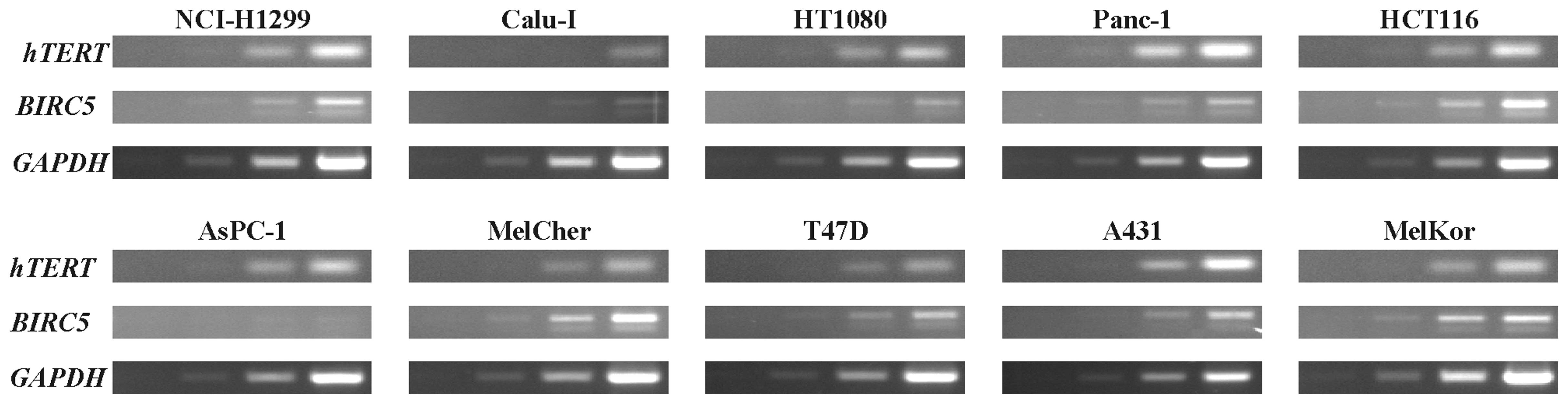
constructions. The relative levels of hTERT and BIRC5
transcripts in cells were determined by semi-quantitative RT-PCR
(Fig. 3; Table II). Juxtaposition of the relative
levels of BIRC5 and hTERT transcript levels (Table II) with the relative activities of
PhTERT and PhSurv (Table I) did not
reveal a correlation between them (r2=0.0696; least
squares method). Similarly, no correlation was observed when the
relative activities of the most potent modified hTERT
promoter and survivin promoter variant were taken into
consideration (r2=0.0762; least squares method). Thus,
the relative levels of hTERT and BIRC5 transcripts
cannot be used to predict better performance of hTERT or
survivin promoters (or their derivatives and variations). In
agreement with our data, no association was observed between
endogenous survivin expression and the level of survivin
promoter-driven transgene activity in the previous study (20).
| Table II.Semi-quantified results of reverse
transcription-polymerase chain reaction (Fig. 3). |
Table II.
Semi-quantified results of reverse
transcription-polymerase chain reaction (Fig. 3).
| Transcript | NCI-H1299 | Calu-I | HT1080 | Panc-1 | HCT116 | AsPC-1 | MelCher | T47D | A431 | MelKor |
|---|
| hTERT | 0.92 | 0.14 | 0.54 | 1.00 | 0.59 | 0.73 | 0.31 | 0.16 | 0.75 | 0.49 |
| BIRC5 | 0.31 | 0.15 | 0.22 | 0.11 | 0.73 | 0.03 | 0.67 | 0.21 | 0.21 | 1.00 |
| Ratio | 0.34 | 1.10 | 0.40 | 0.11 | 1.24 | 0.04 | 2.17 | 1.38 | 0.29 | 2.04 |
Discussion
The choice of promoter for the expression of
therapeutic genes in tumor gene therapy is of critical importance.
Specificity of expression in cancer cells determines the degree of
safety of the gene therapeutic approach, whereas promoter strength
determines the efficiency of gene expression and, consequently, the
effectiveness of treatment. Despite the obvious importance of these
parameters, there is virtually no comparison of different promoters
used for gene-therapeutic purposes. This renders it difficult to
select the promoter that would provide the best outcome when
utilized for a certain tumor treatment. In the present study, such
a comparison was undertaken to assess the two promoters that are
known for their activity in a wide variety of tumors. The results
obtained provide several important inferences.
The first inference relates to selection of promoter
modification to achieve the most pronounced activity. Taking into
account that modification of PhTERT with a minimal CMV
promoter generally results in the most potent promoter among the
studied hTERT-based promoters, and that this modification
preserves the intrinsic tumor specificity of the hTERT
promoter (34), PhTERT-C
is the promoter of choice within the analyzed hTERT promoter
series to be used in transcriptional targeting of tumor cells. The
modified human PhSurvD and mouse PmSurv
survivin gene promoters are more active than the human
PhSurv promoter; however, their tumor specificity
requires further investigation before practical utilization.
An important conclusion is associated with the
observation that the relative levels of hTERT and
BIRC5 transcripts cannot be used to predict better
performance of hTERT or survivin promoters (or their
derivatives and variations). It is likely that the promoters used
for the ectopic expression of the reporter gene do not contain the
distal or proximal regulatory elements used for endogenous
regulation of the corresponding gene expression.
Finally, and most importantly, the two promoters
have strikingly different cell specificities. In some cells,
hTERT-based promoters have predominant activity, whereas
survivin-based promoters have their own cells of preference. Only
40% of the cells analyzed maintain activity of both of the promoter
series. The levels of promoter activity are strongly different
among the cell lines, such that PhTERT activity
expressed relative to the activity of PhSurv ranged from
0.17 in A431 cells to 8.56 in MelKor cells, giving 50-fold
difference between PhTERT or PhSurv
activities among the tumor cell lines. In many cases, promoter
activities are complementary to one another such that, if one of
them fails to support efficient transgene expression in a tumor,
the other can be used instead. However, the necessity of selecting
the most active promoter between PhTERT and
PhSurv complicates the use of the separate promoters and
makes highly desirable the creation of a combined promoter composed
from elements derived from hTERT- and survivin-based
promoters so that such a hybrid promoter would be equally active in
different cells. Otherwise, owing to the complementary nature of
survivin and hTERT promoter activities observed in the
present study, the simultaneous use of survivin- and
hTERT-driven therapeutic transgene vectors emerges as a
feasible option to ensure efficient transgene expression in a
variety of cancer cells.
Acknowledgements
This research was supported by the Russian
Foundation for Basic Research (grant nos. 13-04-40173-H and
13-04-40170-H), the Russian Presidential Program ‘Leading
Scientific Schools’ (grant nos. 5638.2010.4 and 1674.2012.4),
Molecular and Cell Biology Program of the Presidium of Russian
Academy of Sciences, and the Russian Federation State programs
contract nos. 16.512.12.2002 and 11411.1008700.13.084.
Glossary
Abbreviations
Abbreviations:
|
TSP
|
tumor-specific promoter
|
|
hTERT
|
human telomerase reverse
transcriptase
|
References
|
1
|
Saukkonen K and Hemminki A:
Tissue-specific promoters for cancer gene therapy. Expert Opin Biol
Ther. 4:683–696. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dorer DE and Nettelbeck DM: Targeting
cancer by transcriptional control in cancer gene therapy and viral
oncolysis. Adv Drug Deliv Rev. 61:554–571. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee M: Hypoxia targeting gene expression
for breast cancer gene therapy. Adv Drug Deliv Rev. 61:842–849.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhu ZB, Makhija SK, Lu B, Wang M,
Kaliberova L, Liu B, Rivera AA, Nettelbeck DM, Mahasreshti PJ,
Leath CA, et al: Transcriptional targeting of tumors with a novel
tumor-specific survivin promoter. Cancer Gene Ther. 11:256–262.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qiao J, Doubrovin M, Sauter BV, Huang Y,
Guo ZS, Balatoni J, Akhurst T, Blasberg RG, Tjuvajev JG, Chen SH
and Woo SL: Tumor-specific transcriptional targeting of suicide
gene therapy. Gene Ther. 9:168–175. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hardcastle J, Kurozumi K, Chiocca EA and
Kaur B: Oncolytic viruses driven by tumor-specific promoters. Curr
Cancer Drug Targets. 7:181–189. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Glinka EM, Edelweiss EF and Deyev SM:
Eukaryotic expression vectors and immunoconjugates for cancer
therapy. Biochemistry (Mosc). 71:597–606. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sadeghi H and Hitt MM: Transcriptionally
targeted adenovirus vectors. Curr Gene Ther. 5:411–427. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Altieri DC: Survivin, cancer networks and
pathway-directed drug discovery. Nat Rev Cancer. 8:61–70. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Andersen MH, Svane IM, Becker JC and
Straten PT: The universal character of the tumor-associated antigen
survivin. Clin Cancer Res. 13:5991–5994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bao R, Connolly DC, Murphy M, Green J,
Weinstein JK, Pisarcik DA and Hamilton TC: Activation of
cancer-specific gene expression by the survivin promoter. J Natl
Cancer Inst. 94:522–528. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen JS, Liu JC, Shen L, Rau KM, Kuo HP,
Li YM, Shi D, Lee YC, Chang KJ and Hung MC: Cancer-specific
activation of the survivin promoter and its potential use in gene
therapy. Cancer Gene Ther. 11:740–747. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fukuda S and Pelus LM: Survivin, a cancer
target with an emerging role in normal adult tissues. Mol Cancer
Ther. 5:1087–1098. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li F and Altieri DC: Transcriptional
analysis of human survivin gene expression. Biochem J. 344:305–311.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Vaĭshlia NA, Zinov'eva MV, Sass AV,
Kopantsev EP, Vinogradova TV and Sverdlov ED: Increase of BIRC5
gene expression in non-small cell lung cancer and esophageal
squamous cell carcinoma does not correlate with expression of genes
SMAC/DIABLO and PML encoding its inhibitors. Mol Biol (Mosk).
42:652–661. 2008.PubMed/NCBI
|
|
17
|
Van Houdt WJ, Haviv YS, Lu B, Wang M,
Rivera AA, Ulasov IV, Lamfers ML, Rein D, Lesniak MS, Siegal GP, et
al: The human survivin promoter: A novel transcriptional targeting
strategy for treatment of glioma. J Neurosurg. 104:583–592. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu B, Makhija SK, Nettelbeck DM, Rivera
AA, Wang M, Komarova S, Zhou F, Yamamoto M, Haisma HJ, Alvarez RD,
et al: Evaluation of tumor-specific promoter activities in
melanoma. Gene Ther. 12:330–338. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rein DT, Breidenbach M, Nettelbeck DM,
Kawakami Y, Siegal GP, Huh WK, Wang M, Hemminki A, Bauerschmitz GJ,
Yamamoto M, et al: Evaluation of tissue-specific promoters in
carcinomas of the cervix uteri. J Gene Med. 6:1281–1289. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Konopka K, Spain C, Yen A, Overlid N,
Gebremedhin S and Düzgüneş N: Correlation between the levels of
survivin and survivin promoter-driven gene expression in cancer and
non-cancer cells. Cell Mol Biol Lett. 14:70–89. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mityaev MV, Kopantzev EP, Buzdin AA,
Vinogradova TV and Sverdlov ED: Enhancer element potentially
involved in human survivin gene promoter regulation in lung cancer
cell lines. Biochemistry (Mosc). 75:182–191. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bett AJ, Prevec L and Graham FL: Packaging
capacity and stability of human adenovirus type 5 vectors. J Virol.
67:5911–5921. 1993.PubMed/NCBI
|
|
23
|
Li F and Altieri DC: The cancer
antiapoptosis mouse survivin gene: Characterization of locus and
transcriptional requirements of basal and cell cycle-dependent
expression. Cancer Res. 59:3143–3151. 1999.PubMed/NCBI
|
|
24
|
Xia F and Altieri DC: Mitosis-independent
survivin gene expression in vivo and regulation by p53.
Cancer Res. 66:3392–3395. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kyo S, Takakura M, Fujiwara T and Inoue M:
Understanding and exploiting hTERT promoter regulation for
diagnosis and treatment of human cancers. Cancer Sci. 99:1528–1538.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gu J and Fang B: Telomerase
promoter-driven cancer gene therapy. Cancer Biol Ther. 2(4 Suppl
1): S64–S70. 2003.PubMed/NCBI
|
|
27
|
Janknecht R: On the road to immortality:
HTERT upregulation in cancer cells. FEBS Lett. 564:9–13. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Horikawa I, Cable PL, Afshari C and
Barrett JC: Cloning and characterization of the promoter region of
human telomerase reverse transcriptase gene. Cancer Res.
59:826–830. 1999.PubMed/NCBI
|
|
29
|
Takakura M, Kyo S, Kanaya T, Hirano H,
Takeda J, Yutsudo M and Inoue M: Cloning of human telomerase
catalytic subunit (hTERT) gene promoter and identification of
proximal core promoter sequences essential for transcriptional
activation in immortalized and cancer cells. Cancer Res.
59:551–557. 1999.PubMed/NCBI
|
|
30
|
Horikawa I and Barrett JC: Transcriptional
regulation of the telomerase hTERT gene as a target for cellular
and viral oncogenic mechanisms. Carcinogenesis. 24:1167–1176. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Poole JC, Andrews LG and Tollefsbol TO:
Activity, function, and gene regulation of the catalytic subunit of
telomerase (hTERT). Gene. 269:1–12. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wick M, Zubov D and Hagen G: Genomic
organization and promoter characterization of the gene encoding the
human telomerase reverse transcriptase (hTERT). Gene. 232:97–106.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cong YS, Wen J and Bacchetti S: The human
telomerase catalytic subunit hTERT: Organization of the gene and
characterization of the promoter. Hum Mol Genet. 8:137–142. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Davis JJ, Wang L, Dong F, Zhang L, Guo W,
Teraishi F, Xu K, Ji L and Fang B: Oncolysis and suppression of
tumor growth by a GFP-expressing oncolytic adenovirus controlled by
an hTERT and CMV hybrid promoter. Cancer Gene Ther. 13:720–723.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wirth T, Zender L, Schulte B, Mundt B,
Plentz R, Rudolph KL, Manns M, Kubicka S and Kühnel F: A
telomerase-dependent conditionally replicating adenovirus for
selective treatment of cancer. Cancer Res. 63:3181–3188.
2003.PubMed/NCBI
|
|
36
|
Mikhaĭlova IN, Lukashina MI, Baryshnikov
AIu, Morozova LF, Burova OS, Palkina TN, Kozlov AM, Golubeva VA,
Cheremushkin EA, Doroshenko MB, et al: Melanoma cell lines as the
basis for antitumor vaccine preparation. Vestn Ross Akad Med Nauk.
37–40. 2005.
|