Introduction
Lung cancer is the leading cause of
cancer-associated mortality worldwide (1,2). The
majority of lung cancers belong to the non-small cell lung cancer
(NSCLC) type, which accounts for >80% of all cases of lung
cancer (3). The main characteristic
of NSCLC is its high aggressive invasion ability and metastatic
properties (4). Therefore, the
majority of advanced NSCLC cases require to receive combination of
chemotherapy, and/or radiation treatment (5). The survival rates of patients with NSCLC
are poor, and >85% of patients succumb to disease within 5 years
of diagnosis (6). Due to the low
therapeutic efficacy and serious side effects of current
chemotherapeutic drugs (7), a
paradigm shift is obviously required for the management of this
disease. In this context, recent cancer research has focused on the
possibility of preventing or controlling cancer using dietary
agents and phytochemicals, which are less toxic than
chemotherapeutic drugs (8).
Natural substances exert their anti-cancer activity
by modulating cell cycle progression and inducing
apoptosis-regulatory proteins (8,9). Apoptosis
is a type I programmed cell death that is characterized by distinct
phenotypes from necrosis, including membrane blebbing, cell
shrinkage, nuclear fragmentation, chromatin condensation and
apoptotic body formation (10,11). The
apoptotic process happens in either a caspase-dependent or
-independent manner (12). In
addition, B-cell lymphoma (Bcl)-2 family proteins are important for
apoptosis regulation, which could be either proapoptotic, such as
Bcl-2-associated X protein (Bax) or Bcl-2 homologous
antagonist/killer, or antiapoptotic, such as Bcl-2 and Bcl-extra
large (Bcl-xL) (13).
The cell cycle is controlled by a complex series of
signaling pathways by which a cell grows, replicates its DNA and
divides (14). This process includes
mechanisms to ensure errors are corrected while cell replication
occurs, and if the correction cannot be performed, cells enter into
the apoptotic process (15). However,
in cancer cells, this regulatory process malfunctions and results
in uncontrolled cell proliferation (16). Therefore, induction of apoptosis and
modulation of cell cycle is very important in cancer treatment with
anti-cancer agents.
Citrus platymamma hort. ex Tanaka (also known
as Byungkyul in Korea) belongs to the Rutaceae family, and has been
used in Korean traditional medicine for the treatment of
inflammatory disorders and cancer (17). Flavonoids, which are abundantly
present in fresh fruits and vegetables, are known to safely
modulate physiological functions and enhance anti-cancer activity
(18,19). Citrus species, including
Citrus platymamma, contain a wide range of active components
such as flavonoids, which exhibit anti-proliferative, anti-cancer,
anti-oxidant, anti-inflammatory and anti-diabetic activities
(20). Numerous studies have been
reported on Citrus flavonoids that inhibit the growth of
various cancer cells and exhibit anti-inflammatory effects in
vivo and in vitro (21–23).
However, the cellular mechanism of the anti-cancer properties of
flavonoids from Citrus platymamma (FCP) remains to be
elucidated.
Based on the above evidences, the present authors
hypothesized that FCP may exert anti-cancer effects, since they
have been used as a traditional medicine for cancer treatment.
Therefore, in the present study, FCP were isolated and
characterized, and the mechanisms of their anti-cancer effects were
investigated on A549 cells. These flavonoids induced G2/M cell
cycle arrest and apoptosis in A549 cells. To the best of our
knowledge, the present study is the first report that elucidates
the molecular mechanism of FCP in inducing apoptosis and G2/M cell
cycle arrest on the A549 human lung cancer cell line.
Materials and methods
Materials and reagents
A549 human lung cancer cells were obtained from the
Korean Cell Line Bank (Seoul, Korea). RPMI-1640 medium, fetal
bovine serum (FBS) and antibiotics (penicillin/streptomycin) were
purchased from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Hoechst 33342 and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
were obtained from Sigma-Aldrich (St. Louis, MO, USA). Materials
and chemicals used for electrophoresis were obtained from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA). Antibodies against Bcl-xL
(#2762), Bax (#2772), caspase-3 (#9662), caspase-6 (#9762),
caspase-8 (#9746) and caspase-9 (#9502), cleaved caspase-3 (#9661),
poly (adenosine diphosphate-ribose) polymerase (PARP; #9542) and
cleaved PARP (#9541), were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Anti-cyclin B1 (#05-373),
anti-cyclin-dependent kinase 1 (CDK1; #06-923), anti-cell division
cycle 25c (cdc25c; #05-507) and anti-β-actin antibodies (#MABT825)
were obtained from EMD Millipore (Billerica, MA, USA). Horseradish
peroxidase (HRP)-coupled goat anti-mouse IgG ALX-211-205TS-C100 and
anti-rabbit IgG ADI-SAB-301-J were purchased from Enzo Life
Sciences, Inc. (Farmingdale, NY, USA). Muse™ Annexin V & Dead
Cell kit was purchased from EMD Millipore.
Isolation of flavonoids from Korean
Citrus platymamma hort. ex Tanaka
The fruit of Korean Citrus platymamma hort.
ex Tanaka (called Byungkyul in Korea) was obtained from the Animal
Bio Resources Bank (Jinju, Korea). The flavonoids were isolated at
the Department of Chemistry, Gyeongsang National University (Jinju,
Korea) by Professor Sung Chul Shin. The sample was prepared
according to a previously described method (24). Samples were stored at −20°C until used
for further experiments.
Cell culture and treatment
A549 cells were grown in RPMI-1640 medium
supplemented with 10% heat-inactivated FBS and 1%
penicillin/streptomycin in a humidified incubator with 5%
CO2 in air at 37°C. The stock solution of flavonoids was
prepared in dimethyl sulfoxide (DMSO), and subsequent dilutions
were prepared for treatment. Cells grown to 70–80% confluence were
untreated (control) or treated with FCP at various concentrations
(100, 200, 300, 400 and 500 µg/ml) for 24 h in complete growth
medium.
Cell viability assay and morphological
studies
A549 cells were seeded at 10×104 cells/ml
in a 12-well plate and incubated for 24 h. The cytotoxicity was
measured by a standard MTT assay following treatment with FCP at
the specified concentrations for 24 h at 37°C. After 24-h
incubation, 100 µl MTT reagent (5 mg/ml) was added to each well,
and incubated at 37°C for 3 h to form formazan crystals. Then, the
supernatant was discarded, and 500 µl DMSO was added to each well
to dissolve the crystals. The optical density of the cells at 540
nm was measured using an enzyme-linked immunosorbent assay plate
reader. The morphology of flavonoids-treated A549 cells was
observed under a phase contrast microscope (Olympus Corporation,
Tokyo, Japan).
Flow cytometry analysis
Cells were seeded at a density of 5×105
cells/well into a 6-well plate and treated with FCP at various
concentrations (0, 91, 182 and 364 µg/ml) for 24 h at 37°C. The
floating and adherent cells were collected, washed twice with cold
phosphate-buffered saline (PBS) and centrifuged at 300 × g for 5
min at room temperature. The cell pellet was fixed using cold 70%
ethanol (v/v) for 3 h at −20°C. The cells were washed once with
PBS, and 200 µl of the cell suspension was transferred to a fresh
tube. Next, 200 µl Muse™ Cell Cycle kit reagent (EMD Millipore) was
added to each tube and incubated for 30 min at room temperature in
the dark. For the apoptosis assay, the floating and trypsinized
cells were collected, washed once with cold PBS and centrifuged at
300 × g for 5 min at room temperature. The pellet was resuspended
in 1 ml medium, and 100 µl of this suspension was transferred to a
new tube. Then, 100 µl Muse™ Annexin V & Dead Cell kit reagent
was added to each tube and incubated for 20 min at room temperature
in the dark. Then stained samples were analyzed with a Mini Flow
Cytometry Muse™ Cell Analyzer (EMD Millipore).
4′,6-diamidino-2-phenylindole (DAPI)
fluorescent staining
A549 cells were grown to 80% confluence on 10-mm
diameter glass slides placed in 24-well plates and treated with FCP
at the indicated concentrations for 24 h at 37°C. Cells were washed
with cold PBS and fixed with 37% formaldehyde (1:4 dilution with
95% ethanol) for 10 min at room temperature. The fixed cells were
washed with PBS and stained with DAPI and VECTASHIELD®
(catalog number H-1500; Vector Laboratories, Inc., Burlingame, CA,
USA). Cells were mounted in Fluorescence Mounting Medium (Dako,
Glostrup, Denmark), and the nuclear morphology of the cells was
examined by fluorescence microscopy (magnification, ×400; Leica
Microsystems GmbH, Wetzlar, Germany).
Western blot analysis
The protein content was quantified using a Bradford
assay (Bio-Rad Laboratories, Inc.). Proteins were separated by 12%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred to a polyvinylidene fluoride membrane (Immunobilon-P,
0.45 mm; EMD Millipore) using the TE 77 semi-dry transfer unit (GE
Healthcare Life Sciences, Chalfont, UK). The membranes were blocked
with 5% non-fat milk in Tris-buffered saline containing 0.1% Tween
20 (pH 7.4) at room temperature for 1 h. Membranes were incubated
overnight at 4°C with the primary antibodies (dilution, 1:1,000),
and a second incubation was conducted with HRP-conjugated secondary
antibodies (dilution, 1:1,000) for 3 h at room temperature. The
bound antibodies were visualized using an enhanced
chemiluminescence kit (GE Healthcare Life Sciences), and images
were acquired using a ChemiDoc™ XRS+ system with Image Lab™
software version 4.1 (Bio-Rad Laboratories, Inc.).
Statistical analysis
Data are expressed as the mean ± standard deviation
of ≥3 independent experiments. Statistical analysis was performed
with Student's t-test and one-way analysis of variance
(ANOVA), using SPSS version 10.0 (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate as statistical significant
difference. Significant differences were determined using ANOVA
with post-test Neuman-Keuls method for comparisons of ≥3 treatment
groups, while Student's t test was used for comparisons of
two groups.
Results
High-performance liquid chromatography
(HPLC) chromatogram
The HPLC chromatogram of the fruit of Korean
Citrus platymamma hort. ex Tanaka, which was recorded at 280
nm, is shown in Fig. 1, where peaks
corresponding to 11 different flavonoids were clearly
distinguished. These peaks were identified based on their HPLC
retention times, molecular ion masses and spectroscopic data. In
total, 11 flavonoids were successfully identified from Korean
Citrus platymamma hort. ex Tanaka, including neoeriocitrin,
naringin, hesperidin, isosinensetin, sinensetin,
tetramethyl-O-isoscutellarein, nobiletin, tetramethoxyflavone,
heptamethoxyflavone, tangeretin and hydroxypentamethoxyflavone. The
quantification is shown in Table
I.
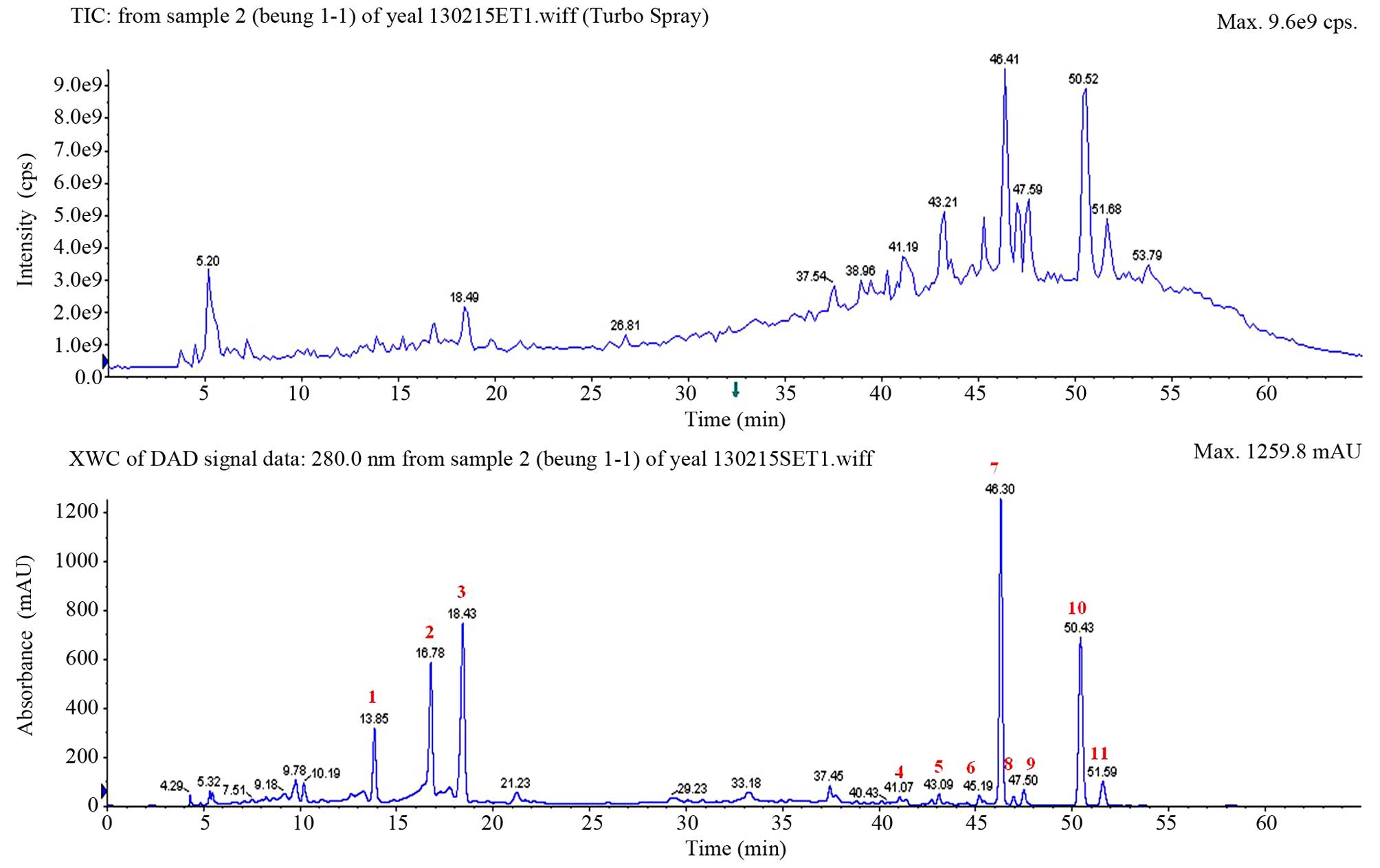 | Figure 1.High-performance liquid
chromatography chromatogram patterns of the fruit of Korean
Citrus platymamma hort. ex Tanaka recorded at 280 nm. The
identified flavonoids from the peaks marked in red were as follows:
1, neoeriocitrin; 2, naringin; 3, hesperidin; 4, isosinensetin; 5,
sinensetin; 6, tetramethyl-O-isoscutellarein; 7, nobiletin; 8,
tetramethoxyflavone; 9, heptamethoxyflavone; 10, tangeretin; and
11, hydroxypentamethoxyflavone. TIC, total ion chromatogram; XWC,
extracted wavelength chromatogram; DAD, diode array detector; cps,
counts per second; mAU, milli-absorption units. |
 | Table I.Spectral data and retention time of
the identified flavonoids isolated from Korean Citrus
platymamma hort. ex Tanaka. |
Table I.
Spectral data and retention time of
the identified flavonoids isolated from Korean Citrus
platymamma hort. ex Tanaka.
| No. | Compound | Retention time
(min) |
[M+H]+/[M-H]−
(m/z) | MS/MS (m/z) |
|---|
| 1 | Neoeriocitrin | 13.85 | −/595 | 459, 329, 311, 287,
151, 135, 107 |
| 2 | Naringin | 16.78 | −/579 | 459, 313, 271, 193,
151 |
| 3 | Hesperidin | 18.43 | −/609 | 608, 325, 301 |
| 4 | Isosinensetin | 41.07 | 373 | 358, 357, 343, 329,
181, 165 |
| 5 | Sinensetin | 43.09 | 373 | 358, 357, 343, 340,
329, 312, 162 |
| 6 |
Tetramethyl-O-isoscutellarein | 45.19 | 343 | 328, 313, 299, 285,
240, 152, 133 |
| 7 | Nobiletin | 46.30 | 403 | 388, 373, 355, 327,
241, 211, 165 |
| 8 |
Tetramethoxyflavone | 46.95 | 343 | 327, 313, 282,
150 |
| 9 |
Heptamethoxyflavone | 47.50 | 433 | 418, 403, 385, 211,
165 |
| 10 | Tangeretin | 50.43 | 373 | 358, 343, 325,
297 |
| 11 |
Hydroxypentamethoxyflavone | 51.59 | 389 | 374, 359, 341,
165 |
Effect of FCP on A549 cells
proliferation
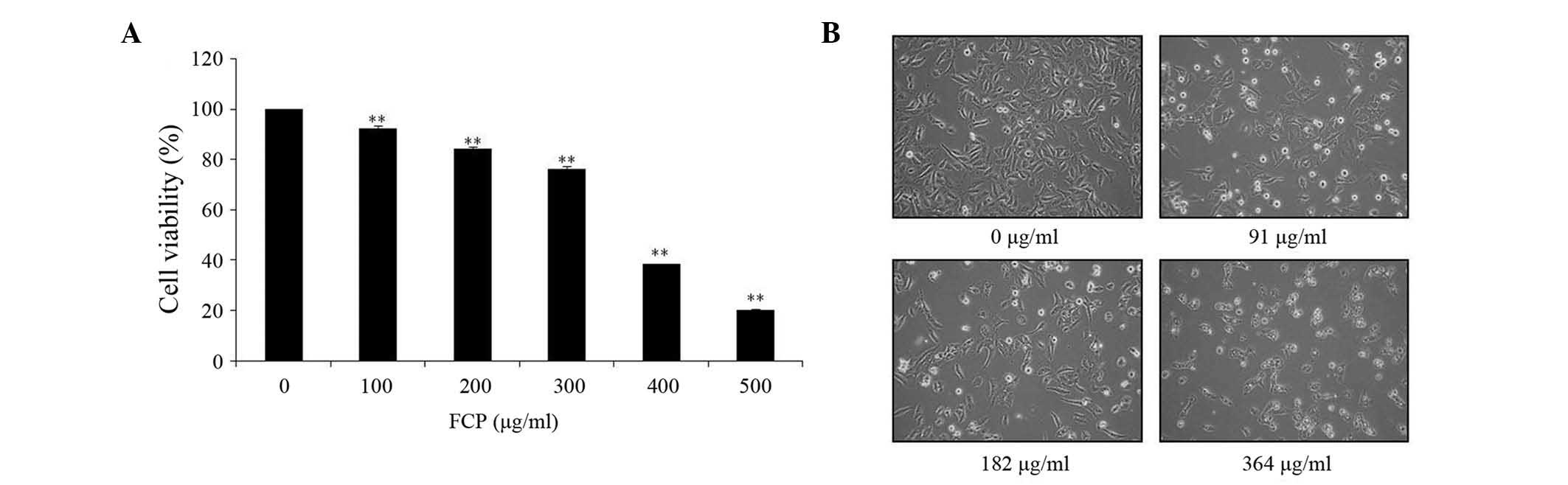
The anti-proliferative effect of FCP was
investigated by MTT assay in A549 cells incubated for 24 h with
different doses of FCP (0, 100, 200, 300, 400 and 500 µg/ml). As
indicated in Fig. 2A, FCP
significantly inhibited the viability of A549 cells in a
dose-dependent manner. The results revealed that the half maximal
inhibitory concentration (IC50) of FCP was 364 µg/ml.
According to their IC50 value, the concentrations of
flavonoids used in the following experiments were 0, 91, 182 and
364 µg/ml. In addition, microscopic examination revealed that
morphological changes in cell shape such as cell shrinkage, and a
gradual decrease in the living cell population, were observed with
increasing concentrations of FCP (Fig.
2B). These results suggest that FCP have anti-proliferative
effects on A549 cells.
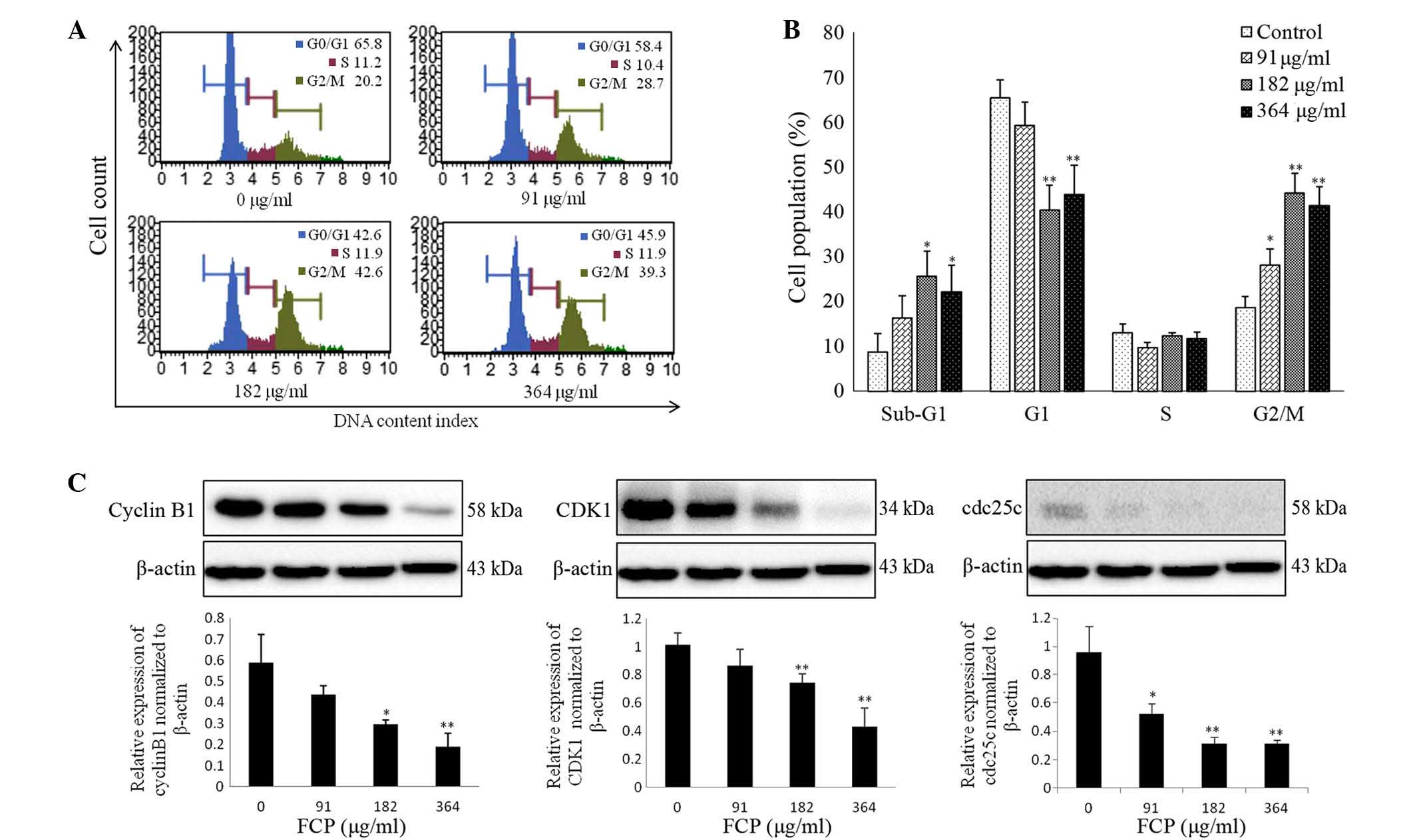
FCP induced G2/M arrest by
downregulation of cyclin B1, CDK1 and cdc25c proteins
To investigate the mechanisms responsible for the
anti-proliferative effects of FCP, the cell cycle progression of
A549 cells following FCP treatment at the specified concentrations
was examined by flow cytometry. The sub-G1 apoptotic cell
population and G2/M phase population were significantly increased
in a dose-dependent manner, whereas the number of cells in the
G0/G1 and S phases were remarkably decreased (Fig. 3A). Fig.
3B shows the quantitative representations of cell cycle
progression in A549 cells treated with FCP, which were observed to
be dose-dependent. These results indicate that FCP are able to
induce G2/M arrest and apoptosis in A549 cells.
Cyclin B1, cdc2 and cdc25c are
important in the cell cycle during the G2/M phase (14)
This phase is controlled by formation of a complex
of cyclin B1 and CDK1, which is regulated by cdc25c (25). In the present study, the effect of FCP
on the regulatory proteins involved in G2/M arrest was examined by
immunoblotting, which revealed that FCP significantly decreased the
expression levels of cyclin B1, CDK1 and cdc25c proteins in a
dose-dependent manner (Fig. 3C).
These data suggest that FCP induce G2/M arrest by downregulation of
cyclin B1, CDK1 and cdc25c proteins in A549 cells.
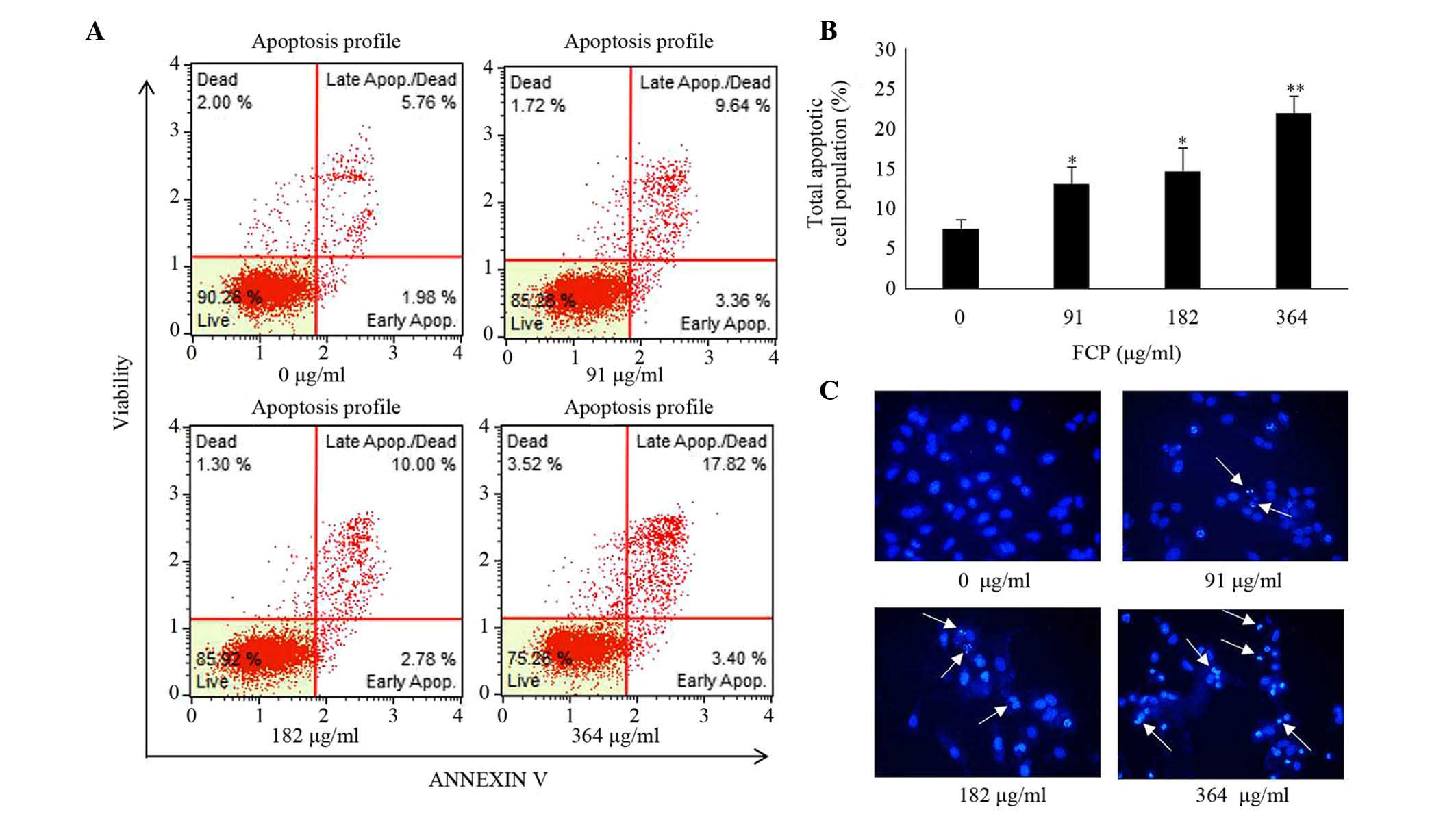
FCP induced apoptosis in A549
cells
Induction of apoptosis was examined in A549 cells
following FCP treatment by Annexin V -fluorescein isothiocyanate
(FITC)/propidium iodide (PI) double-labelled flow cytometry and
Hoechst 33342 staining. In FCP-treated A549 cells, the total
apoptotic cell proportion was significantly increased in a
dose-dependent manner (Fig. 4A and
B), and the late apoptotic cell proportion was higher than the
early apoptotic cell proportion, compared with the control.
Furthermore, Hoechst 33342 staining of FCP-treated A549 cells
revealed apoptotic changes in these cells, including condensed and
fragmented nuclei (Fig. 4C). These
data indicate that FCP are able to induce apoptosis in A549
cells.
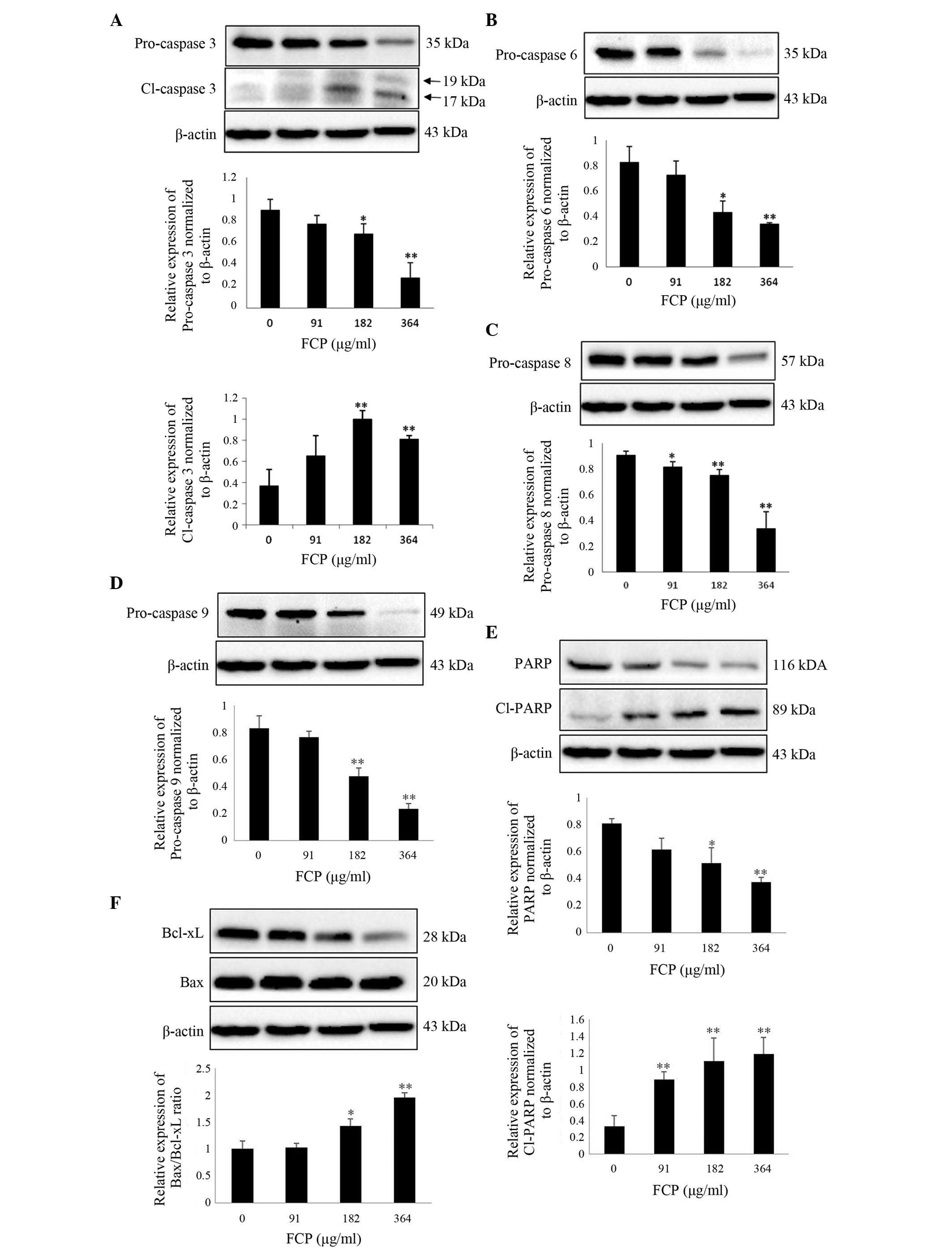
FCP induce caspase-3 activation and
subsequent cleavage of PARP in A549 cells
It is well known that caspases play a key role in
modulating apoptosis (26). Among
them, caspase-3 is the major effector protein of apoptosis, and is
activated by an initiator caspase such as caspase-9 (27). The activation of caspases and
subsequent cleavage of PARP from the native 116-kDa form to the
89-kDa protein is one of the hallmarks of apoptosis (28). To clarify the mechanism of FCP-induced
apoptosis, the expression of apoptosis-related proteins in A549
cells following FCP treatment was examined. Upon treatment with
FCP, the expression levels of PARP and pro-caspases −3, −6, −8 and
−9 were significantly decreased in a dose-dependent manner, which
indicates caspase activation (Fig.
5A-D). FCP significantly increased the levels of cleaved PARP,
a substrate of caspase-3, in A549 cells (Fig. 5E). The ratio of proapoptotic and
antiapoptotic proteins is the determinant factor of apoptosis. In
the present study, the Bax/Bcl-xL ratio was significantly increased
by FCP in a dose-dependent manner (Fig.
5F). These results suggest that FCP induce apoptosis in A549
cells by the activation of caspases and subsequent PARP cleavage,
at least in part through an increase in the ratio of
Bax/Bcl-xL.
Discussion
To investigate the anti-cancer effects of FCP and
their anti-cancer mechanisms in A549 human lung cancer cells, a
mixture of 11 flavonoids was isolated from Korean Citrus
platymamma hort. ex Tanaka and characterized by HPLC-tandem
mass spectrometry. The flavonoids identified in the present study
are similar to the previously reported flavonoids in other
Citrus spp. (21,29). The present authors previously
demonstrated that the flavonoids isolated from Citrus
aurantium exhibited anti-cancer and anti-inflammatory effects
against various cancer cell lines (24,30,31). In
addition, monomers such as naringin, nobiletin and hesperidin have
demonstrated anti-cancer activity in vitro and in
vivo (22,32). However, no reports are available
regarding the anti-cancer activities of FCP. Therefore, the present
study attempted to elucidate the mechanism responsible for the
anti-cancer effects of FCP on A549 cells.
Firstly, the viability of A549 cells following
treatment with FCP was assessed. FCP strongly inhibited A549 cell
growth, and dose-dependent accumulations of sub-G1 population
(apoptotic cell population) and G2/M phase populations were
observed. Immunoblotting indicated that FCP induced G2/M arrest by
regulation of cyclin B1, CDK1 and cdc25c proteins in A549 cells.
Previous studies demonstrated that the accumulation of G2/M phase
population is prominently associated with apoptosis (33). Similar to the present results, recent
studies have reported that flavonoids induce G2/M arrest and
apoptosis in a wide range of human cancer cell lines (34–36).
Furthermore, FCP-induced apoptosis was confirmed by FITC-Annexin V
and PI double staining in the present study, which also observed
nuclear condensation, fragmentation and apoptotic bodies in
FCP-treated A549 cells. These observations are consistent with
biochemical and morphological changes observed during the apoptosis
process (37), and thus suggest that
FCP effectively induce apoptosis in A549 cells.
Caspases are a family of cysteine proteases that
generally constitute key components of the apoptotic pathways (an
extrinsic death receptor-mediated and an intrinsic
mitochondria-mediated apoptotic pathway) (38). The intrinsic apoptotic pathway is
triggered by activation of the initiator caspase-9, which directly
initiates downstream events by activating caspase-3, a crucial
executioner that is associated with the subsequent cleavage of its
target substrate protein PARP (39–41). The
present results indicated that FCP significantly decreased the
levels of pro-caspases −3, −6, −8 and −9, and increased the
expression of cleaved caspase-3 and cleaved PARP. In the
mitochondrial apoptotic pathway, the ratio of expression of the
proapoptotic and antiapoptotic proteins determines the death or
survival of a cell (42). The present
study demonstrated that the expression of the antiapoptotic Bcl-xL
was decreased, whereas the expression of the proapoptotic Bax
protein was unchanged, and the ratio of Bax/Bcl-xL was
dose-dependently increased in FCP-treated A549 cells. As a result,
cytochrome c is released from the mitochondria into the
cytosol and is able to bind apoptotic protease activating factor 1,
thus leading to the activation of caspase-3 and subsequent cell
death (43,44). In addition, caspase-8 activation was
not prominent in the present study. In the intrinsic
pathway-related apoptosis, caspase-8 can be activated by a feedback
mechanism (45). Therefore, the
present findings suggest that FCP induce apoptosis in A549 cells at
least in part through intrinsic apoptotic pathways by activation of
caspase-3 and PARP cleavage.
In summary, the present results indicate that FCP
induce cell cycle arrest at the G2/M phase by regulating the
protein expression levels of cyclin B1, CDK1 and cdc25c, and induce
caspase-dependent cell death by upregulation of the Bax/Bcl-xL
ratio. Altogether, the present study provides the first report on
the molecular mechanism of FCP-induced apoptosis in A549 cells, and
thus reveals that Citrus platymamma may have a therapeutic
potential for the treatment of human lung cancer.
Acknowledgements
The present study was supported by a grant from the
National Research Foundation of Korea, which is funded by the
Ministry of Science, Information and Communications Technology
& Future Planning of Korea (Seoul, Korea; grant numbers
2012M3A9B8019303 and 2012R1A2A2A06045015) and the National Research
& Development Program for Cancer Control of the Ministry for
Health, Welfare and Family Affairs of Korea (Seoul, Korea; grant
number 0820050).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jung KW, Won YJ, Kong HJ, Oh CM, Seo HG
and Lee JS: Prediction of cancer incidence and mortality in Korea,
2013. Cancer Res Treat. 45:15–21. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meoni G, Cecere FL, Lucherini E and Di
Costanzo F: Medical treatment of advanced non-small cell lung
cancer in elderly patients: A review of the role of chemotherapy
and targeted agents. J Geriatr Oncol. 4:282–290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park KI, Park HS, Kang SR, Nagappan A, Lee
DH, Kim JA, Han DY and Kim GS: Korean Scutellaria baicalensis water
extract inhibits cell cycle G1/S transition by suppressing cyclin
D1 expression and matrix-metalloproteinase-2 activity in human lung
cancer cells. J Ethnopharmacol. 133:634–641. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: epidemiology, risk
factors, treatment, and survivorship. Mayo Clin Proc. 83:584–594.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liang XJ, Chen C, Zhao Y and Wang PC:
Circumventing tumor resistance to chemotherapy by nanotechnology.
Methods Mol Biol. 596:467–488. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hyun HB, Lee WS, Go SI, Nagappan A, Park
C, Han MH, Hong SH, Kim G, Kim GY, Cheong J, et al: The flavonoid
morin from Moraceae induces apoptosis by modulation of Bcl-2 family
members and Fas receptor in HCT 116 cells. Int J Oncol.
46:2670–2678. 2015.PubMed/NCBI
|
|
9
|
Jeong JW, Lee WS, Go SI, Nagappan A, Baek
JY, Lee JD, Lee SJ, Park C, Kim GY, Kim HJ, et al: Pachymic Acid
Induces Apoptosis of EJ Bladder Cancer Cells by DR5 Up-Regulation,
ROS Generation, Modulation of Bcl-2 and IAP Family Members.
Phytother Res. 29:1516–1524. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zamzami N, Susin SA, Marchetti P, Hirsch
T, Gómez-Monterrey I, Castedo M and Kroemer G: Mitochondrial
control of nuclear apoptosis. J Exp Med. 183:1533–1544. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cregan SP, Dawson VL and Slack RS: Role of
AIF in caspase-dependent and caspase-independent cell death.
Oncogene. 23:2785–2796. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Findley HW, Gu L, Yeager AM and Zhou M:
Expression and regulation of Bcl-2, Bcl-xl, and Bax correlate with
p53 status and sensitivity to apoptosis in childhood acute
lymphoblastic leukemia. Blood. 89:2986–2993. 1997.PubMed/NCBI
|
|
14
|
DiPaola RS: To arrest or not to G(2)-M
Cell-cycle arrest: Commentary re: A. K. Tyagi et al., Silibinin
strongly synergizes human prostate carcinoma DU145 cells to
doxorubicin-induced growth inhibition, G(2)-M arrest, and
apoptosis. Clin. cancer res., 8: 3512–3519, 2002. Clin Cancer Res.
8:3311–3314. 2002.PubMed/NCBI
|
|
15
|
Adams JM: Ways of dying: multiple pathways
to apoptosis. Genes Dev. 17:2481–2495. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gibbs WW: Untangling the roots of cancer.
Sci Am. 289:56–65. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee HJ, Nagappan A, Park HS, Hong GE,
Yumnam S, Raha S, Saralamma VV, Lee WS, Kim EH and Kim GS:
Flavonoids isolated from Citrus platymamma induce
mitochondrial-dependent apoptosis in AGS cells by modulation of the
PI3K/AKT and MAPK pathways. Oncol Rep. 34:1517–1525.
2015.PubMed/NCBI
|
|
18
|
Liu BL, Zhang X, Zhang W and Zhen HN: New
enlightenment of French Paradox: Resveratrol's potential for cancer
chemoprevention and anti-cancer therapy. Cancer Biol Ther.
6:1833–1836. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hatcher H, Planalp R, Cho J, Torti FM and
Torti SV: Curcumin: From ancient medicine to current clinical
trials. Cell Mol Life Sci. 65:1631–1652. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nogata Y, Sakamoto K, Shiratsuchi H, Ishii
T, Yano M and Ohta H: Flavonoid composition of fruit tissues of
citrus species. Biosci Biotechnol Biochem. 70:178–192. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Benavente-Garcia O and Castillo J: Update
on uses and properties of citrus flavonoids: New findings in
anticancer, cardiovascular and anti-inflammatory activity. J Agric
Food Chem. 56:6185–6205. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Luo G, Guan X and Zhou L: Apoptotic effect
of citrus fruit extract nobiletin on lung cancer cell line A549 in
vitro and in vivo. Cancer Biol Ther. 7:966–973. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lai CS, Li S, Miyauchi Y, Suzawa M, Ho CT
and Pan MH: Potent anti-cancer effects of citrus peel flavonoids in
human prostate xenograft tumors. Food Funct. 4:944–949. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park KI, Park HS, Nagappan A, Hong GE, Lee
do H, Kang SR, Kim JA, Zhang J, Kim EH, Lee WS, et al: Induction of
the cell cycle arrest and apoptosis by flavonoids isolated from
Korean Citrus aurantium L. Food Chem. 135:2728–2735. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Donzelli M and Draetta GF: Regulating
mammalian checkpoints through Cdc25 inactivation. EMBO Rep.
4:671–677. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Parrish AB, Freel CD and Kornbluth S:
Cellular mechanisms controlling caspase activation and function.
Cold Spring Harb Perspect Biol. 5:a0086722013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Slee EA, Adrain C and Martin SJ: Serial
killers: ordering caspase activation events in apoptosis. Cell
Death Differ. 6:1067–1074. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang GH, Mitsui K, Kotliarova S, Yamashita
A, Nagao Y, Tokuhiro S, Iwatsubo T, Kanazawa I and Nukina N:
Caspase activation during apoptotic cell death induced by expanded
polyglutamine in N2a cells. Neuroreport. 10:2435–2438. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Harapu CD, Miron A, Cuciureanu M and
Cuciureanu R: Flavonoids-bioactive compounds in fruits juice. Rev
Med Chir Soc Med Nat Iasi. 114:1209–1214. 2010.(In Romanian).
PubMed/NCBI
|
|
30
|
Lee DH, Park KI, Park HS, Kang SR,
Nagappan A, Kim JA, Kim EH, Lee WS, Hah YS, Chung HJ, et al:
Flavonoids isolated from Korea Citrus aurantium L. Induce
G2/M phase arrest and apoptosis in human gastric cancer AGS Cells.
Evid Based Complement Alternat Med. 2012:5159012012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim JA, Park HS, Park KI, Hong GE,
Nagappan A, Zhang J, Han DY, Shin SC, Won CG, Kim EH and Kim GS:
Proteome analysis of the anti-inflammatory response of flavonoids
isolated from Korean Citrus aurantium L. In
lipopolysaccharide-induced L6 rat skeletal muscle cells. Am J Chin
Med. 41:901–912. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim DI, Lee SJ, Lee SB, Park K, Kim WJ and
Moon SK: Requirement for Ras/Raf/ERK pathway in naringin-induced
G1-cell-cycle arrest via p21WAF1 expression. Carcinogenesis.
29:1701–1709. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boonstra J and Post JA: Molecular events
associated with reactive oxygen species and cell cycle progression
in mammalian cells. Gene. 337:1–13. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nicolini F, Burmistrova O, Marrero MT,
Torres F, Hernández C, Quintana J and Estévez F: Induction of G2/M
phase arrest and apoptosis by the flavonoid tamarixetin on human
leukemia cells. Mol Carcinog. 53:939–950. 2014.PubMed/NCBI
|
|
35
|
Li Y, Duan S, Jia H, Bai C, Zhang L and
Wang Z: Flavonoids from tartary buckwheat induce G2/M cell cycle
arrest and apoptosis in human hepatoma HepG2 cells. Acta Biochim
Biophys Sin (Shanghai). 46:460–470. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tao L, Fu R, Wang X, Yao J, Zhou Y, Dai Q,
Li Z, Lu N and Wang W: LL-202, a newly synthesized flavonoid,
inhibits tumor growth via inducing G(2)/M phase arrest and cell
apoptosis in MCF-7 human breast cancer cells in vitro and
in vivo. Toxicol Lett. 228:1–12. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Majno G and Joris I: Apoptosis, oncosis
and necrosis. An overview of cell death. Am J Pathol. 146:3–15.
1995.PubMed/NCBI
|
|
38
|
Mao WP, Ye JL, Guan ZB, Zhao JM, Zhang C,
Zhang NN, Jiang P and Tian T: Cadmium induces apoptosis in human
embryonic kidney (HEK) 293 cells by caspase-dependent
and-independent pathways acting on mitochondria. Toxicol In Vitro.
21:343–354. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lazebnik YA, Kaufmann SH, Desnoyers S,
Poirier GG and Earnshaw WC: Cleavage of poly (ADP-ribose)
polymerase by a proteinase with properties like ICE. Nature.
371:346–347. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Debatin KM: Apoptosis pathways in cancer
and cancer therapy. Cancer Immunol Immunother. 53:153–159. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
MacKenzie SH and Clark AC: Targeting cell
death in tumors by activating caspases. Curr Cancer Drug Targets.
8:98–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wong WW and Puthalakath H: Bcl-2 family
proteins: The sentinels of the mitochondrial apoptosis pathway.
IUBMB Life. 60:390–397. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cai J, Yang J and Jones DP: Mitochondrial
control of apoptosis: the role of cytochrome c. Biochim Biophys
Acta. 1366:139–149. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu X, Kim CN, Yang J, Jemmerson R and
Wang X: Induction of apoptotic program in cell-free extracts:
requirement for dATP and cytochrome c. Cell. 86:147–157. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shin DY, Ryu CH, Lee WS, Kim DC, Kim SH,
Hah YS, Lee SJ, Shin SC, Kang HS and Choi YH: Induction of
apoptosis and inhibition of invasion in human hepatoma cells by
anthocyanins from meoru. Ann N Y Acad Sci. 1171:137–148. 2009.
View Article : Google Scholar : PubMed/NCBI
|