Introduction
Gastric carcinoma is a common human malignancy with
a high incidence, including in countries such as China (1). The morbidity and mortality associated
with gastric carcinoma is the leading cause of all malignancies
worldwide (2). A recent
epidemiological study revealed that the 5-year survival rate of
gastric carcinoma was approximately 90% (3). Thus, it is important to study and
improve its prevention and treatment alternatives.
Immunotherapy is a relatively new treatment method
based on the manipulation of various immune cells for cancer
treatment. This treatment can improve the ability of recognition
and presentation of tumor antigens by the immune system, resulting
in a reduction in the incidence of tumor recurrence and metastasis
(4–6).
However, to achieve this, immunotherapy should be able to direct
the innate immunity and adaptive response that exerts an antitumor
effect (7,8). Μononuclear cells can be differentiated
into classically activated macrophages (CAMs/Mϕ1) by granulocyte
macrophage colony-stimulating factor (GM-CSF) and play a key role
in the induction of the specific immune response and immunological
regulation (9,10). The γδT cells are an important subset
of the T-cell population of the innate immune system in
vivo, and have an important role in tumor immune surveillance
(11). Currently, studies concerning
the interactions of Mϕ1 and γδT cells are mostly limited to
inflamed tissues (12,13). To the best of our knowledge, the
influence of Mϕ1 on the antitumor effect of γδT cells has yet to be
reported.
We hypothesized that Mϕ1 can signal γδT cells to
achieve a comprehensive cellular immunotherapeutic effect that may
improve the elimination of cancer cells. To test this hypothesis,
we used the supernatant of in vitro cultured Mϕ1 to signal
γδT cells and documented the effects on their proliferation, cell
surface marker expression and cytotoxicity against gastric cancer
cells. In addition, we examined the possible mechanisms involved in
the findings and the opportunity for more direct tests for an
immunotherapeutic approach.
Materials and methods
Materials
The human SGC-7901 gastric cancer cell line was
obtained from the Shanghai Institute of Biochemistry and Cell
Biology (Shanghai, China). rhGM-CSF was purchased from Promega
Corp. (Madison, WI, USA), and rhIL-2 and rhIFN-γ were purchased
from Xiamen Amoytop Biotech Co., Ltd. (Xiamen, China). RPMI-1640
medium, calf serum and tryptase were purchased from Gibco (Grand
Island, NY, USA). Methyl thiazolyl tetrazolium (MTT) and dimethyl
sulfoxide (DMSO) were purchased from Sigma (St. Louis, MO, USA).
Isopentenyl pyrophosphate (IPP), PE-labeled mouse anti-human
monoclonal antibody CD3, FICT-labeled mouse anti-human monoclonal
antibodies CD68, CD44, and γδTCR were purchased from MultiSciences
(Lianke) Biotech Co., Ltd. (Hangzhou, China). Interleukin (IL)-10,
IL-12 were analyzed using the commercially available kit from
Gibco. Lactate dehydrogenase (LDH) was assayed using the
commercially available kit by Shino-Test Corp. (Tokyo, Japan).
Culture and identification of γδT
cells
Approximately 10 ml of peripheral blood with
heparin/EDTA as anticoagulant were drawn aseptically from healthy
donors and added to lymphocyte separation medium. Centrifugation
was performed at 1,500 × g for 15 min and the peripheral blood
mononuclear cells (PBMCs) were separated. The PBMCs were washed
three times with normal saline (each wash consisted of
centrifugation at 1,000 × g/for 10 min) and then added to RPMI-1640
medium supplemented with 10% calf serum, 5% human AB serum, IL-2
150 kU/l and IPP 2 µg/l. The γδT cells were cultured according to
the method described by Chen et al and Liu et al
(14,15). The γδT cells that had been cultured
for 10 days were then collected to detect the cell surface markers
γδTcR and CD44, and to determine the growth and cytotoxicity of γδT
cells.
Macrophage culture
Six healthy volunteers were chosen and 200 ml blood
were drawn from each volunteer. PBMCs were separated with
lymphocyte separation medium. RPMI-1640 complete medium was used to
dilute PBMCs to 2×109/l, and the diluted PBMCs were
seeded in 6-well plates with 5 ml for each well. The cells were
then cultured at 37°C for 2 h with 5% CO2. The
unattached cells were washed with phosphate-buffered saline (PBS)
and the cells that adhered were washed with warm saline only once.
The attached cells were then cultured with RPMI-1640 medium that
contained 700 kU/l GM-CSF and 10% FBS at 37°C with 5%
CO2. Half of the medium was changed once every 2 days,
interferon (IFN)-γ of 166 kU/l was added on the sixth day,
incubated for 24 h and Mϕ1 was retrieved.
Detection of the expression of
macrophage surface marker CD68 using flow cytometry
Mϕ1 cultured for 7, 10 and 13 days were digested
with trypsin and the cell concentrations were adjusted to
5×109/l with PBS. Centrifugation, washing and
resuspension were performed with PBS. Approximately 100 µl of the
cells were then resuspended in centrifuge tubes. CD68 was labeled
with a fluorescent marker (FITC-labeled) to a final concentration
of 5 mg/l and incubated in the dark at 4°C for 20 min. The
unattached label was washed off with PBS and the cell phenotype was
detected using a flow cytometer (Amnis Corp., Seattle, USA).
Detection of the effect of Mϕ1 culture
supernatant on γδT cell proliferation using MTT
γδT cells cultured for 10 days were diluted to
obtain a final concentration of 1×109/l. Subsequently,
0.2 ml of the cell suspension were added to each of the wells in a
96-well plate. Each group had 5 wells in replicates. The plates
were incubated at 37°C in the presence of 5% CO2 for 24
h. This was followed by addition of the culture supernatants of
Mϕ1, which were cultured for 10 days in each well. No Mϕ1
supernatant was added to the control group. The cells were then
cultured for another 72 h under the same conditions and 20 µl of
MTT was added into each well and incubated for 4 h. The supernatant
was removed and 100 µl of DMSO was added per well, and mixed for 10
min. When the precipitate was completely dissolved, the absorbance
(A) of each well was detected at 570 nm wavelength using ELISA. The
cell proliferation rate was calculated using the formula: cell
proliferation = value for test group/value for control group - 1) ×
100%.
Detection of the effect of Mϕ1 culture
supernatant on γδT cell phenotype using a flow cytometer
γδT cells, cultured for 10 days with Mϕ1 supernatant
(test group) or without Mϕ1 supernatant (control group) were
collected. In the test group, the volume ratio of Mϕ1 culture
supernatant to RPMI-1640 medium was 1:8. The concentration of cells
was adjusted to 2×109/l with PBS and 100 µl of cell
suspension was taken in centrifuge tubes. FITC-labeled γδTCR and
anti-human CD44-FITC antibodies were added to the centrifuge tubes
at a final concentration of 5 mg/l, incubated in the dark at 4°C
for 20 min, washed with PBS and detected using a flow
cytometer.
Measurement of tumor cytotoxic effect
of γδT cells following treatment with Mϕ1 culture supernatant
Cell cytotoxicity was detected using the LDH-release
method. SGC-7901 gastric cancer cells were cultured to the
logarithmic phase using high-glucose Dulbecco's modified Eagle's
medium (DMEM). The cells were collected and washed with Hank's
balanced salt solution twice and then prepared at a concentration
of 2×108/l. The γδT cells that were induced for 72 h at
different concentrations (either 1:4, 1:8, 1:32 or 1:64) of Mϕ1
supernatant to RPMI-1640 were used as effector cells at a
concentration of 2×109/l. The SGC-7901 gastric cancer
and effector γδT cells were mixed at a ratio of 10:1. After
centrifugation at 500 rpm for 5 min, the cells were incubated at
37°C for 6 h in the presence of 5% CO2. The cells were
mixed gently and centrifuged at 1,500 rpm for 10 min. Each group of
cells had replicates. The control group consisted of γδT cells that
had no Mϕ1 supernatant added to them. After the incubation period,
the culture supernatants of each group of cells were collected and
used to detect the activity of LDH (U/l). The cytotoxicity of γδT
cells was calculated using the formula: cytotoxicity of γδT cells =
(LDH units of test tubes - LDH units released from effector
cells)/(LDH units of maximum release tubes - LDH units released
from target cells) × 100%.
Statistical analysis
SPSS 19.0 (Chicago, IL, USA)was used to analyze the
results. The values were presented as mean ± standard deviation
(SD). The t-test was used to compare the two groups, while one-way
analysis of variance was used for comparison between groups.
P<0.05 was considered statistically significant.
Results
Cultures, γδT cell phenotype and
assessment of cell purity
PBMCs had adherent growth after culturing for 24 h,
in the γδT cell induction system. The cell colony size markedly
increased after 48 h of cultivation. A single adherent cell layer
was apparent after 10 days of cultivation. The individual cells
presented a fusiform shape and few suspended cells were found. The
cells collected before and after cultivation were labeled with mAb
fluorescence, and then detected and analyzed by flow cytometry.
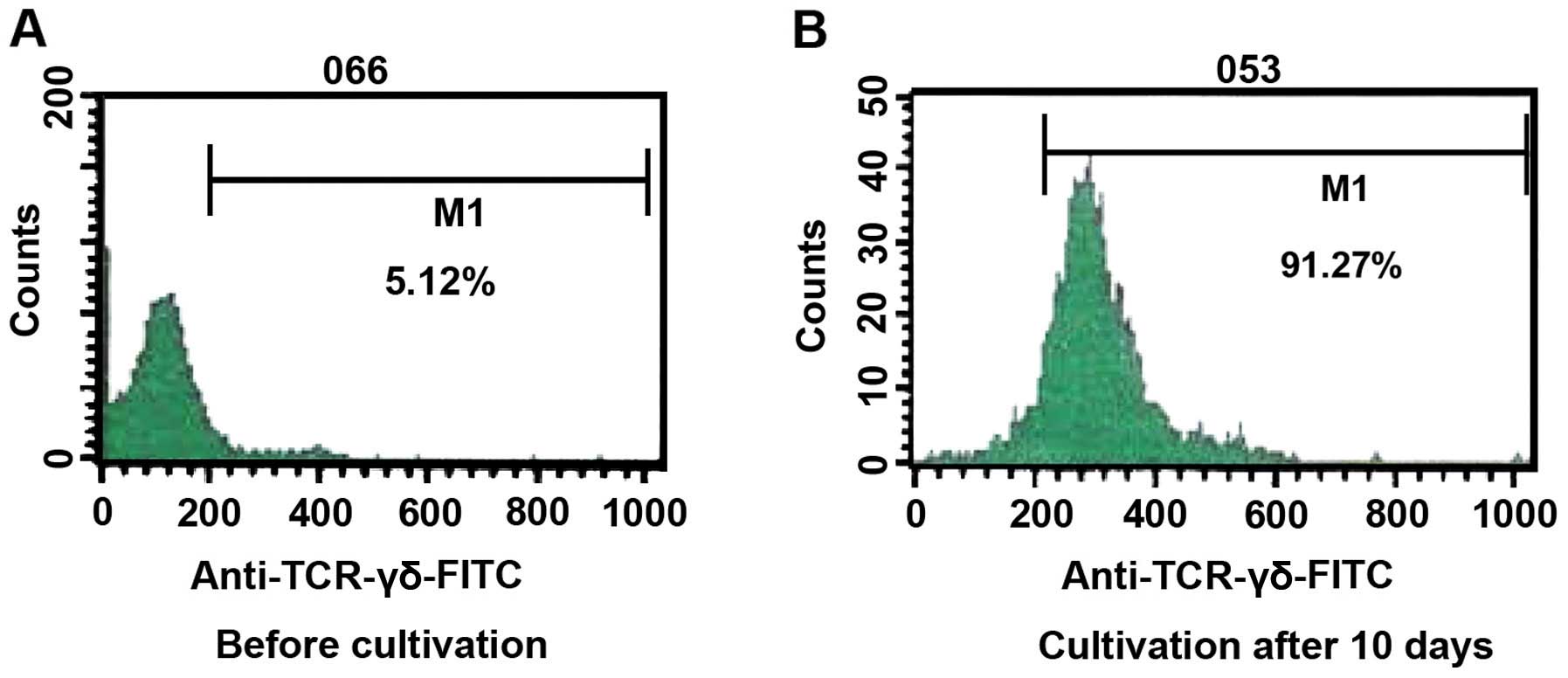
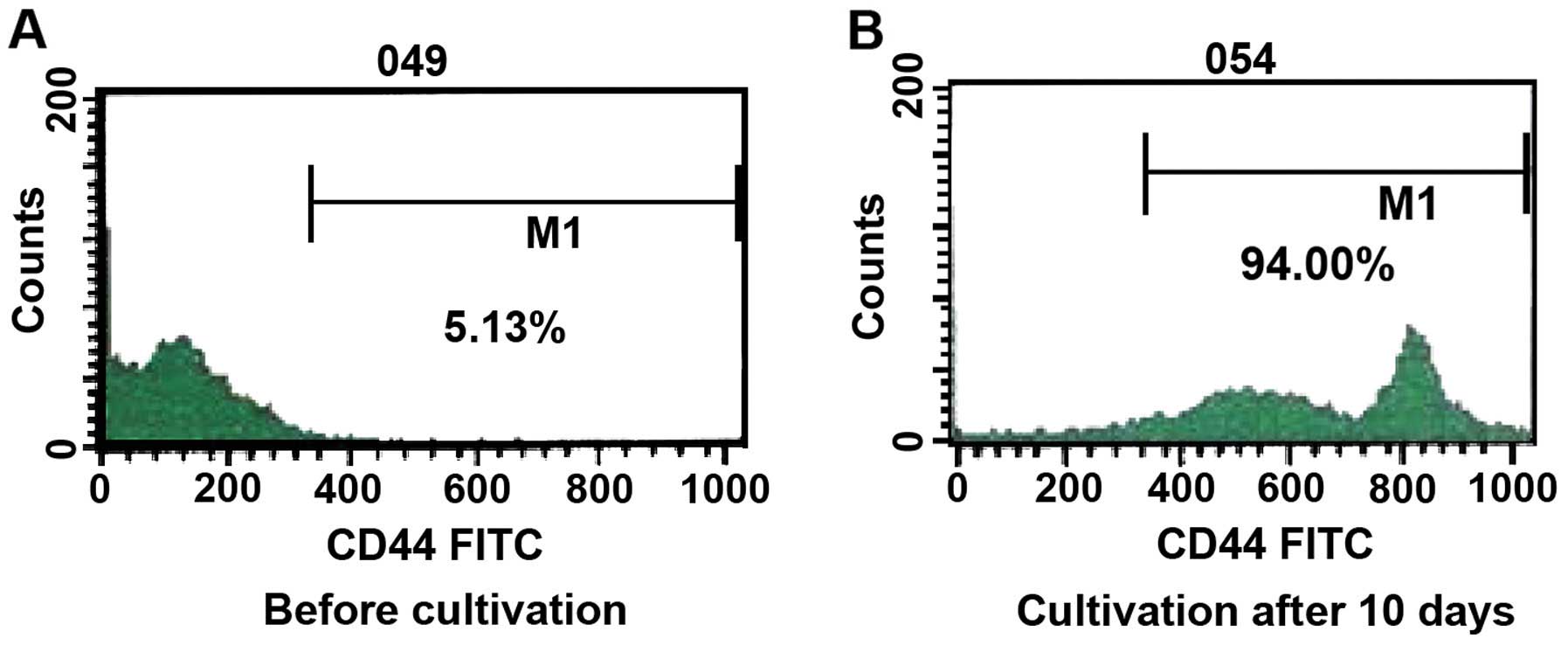
Prior to cultivation, the ratio of γδT cells was 5.12% of the
total, and the expression of CD44 was 5.13%. After cultivation for
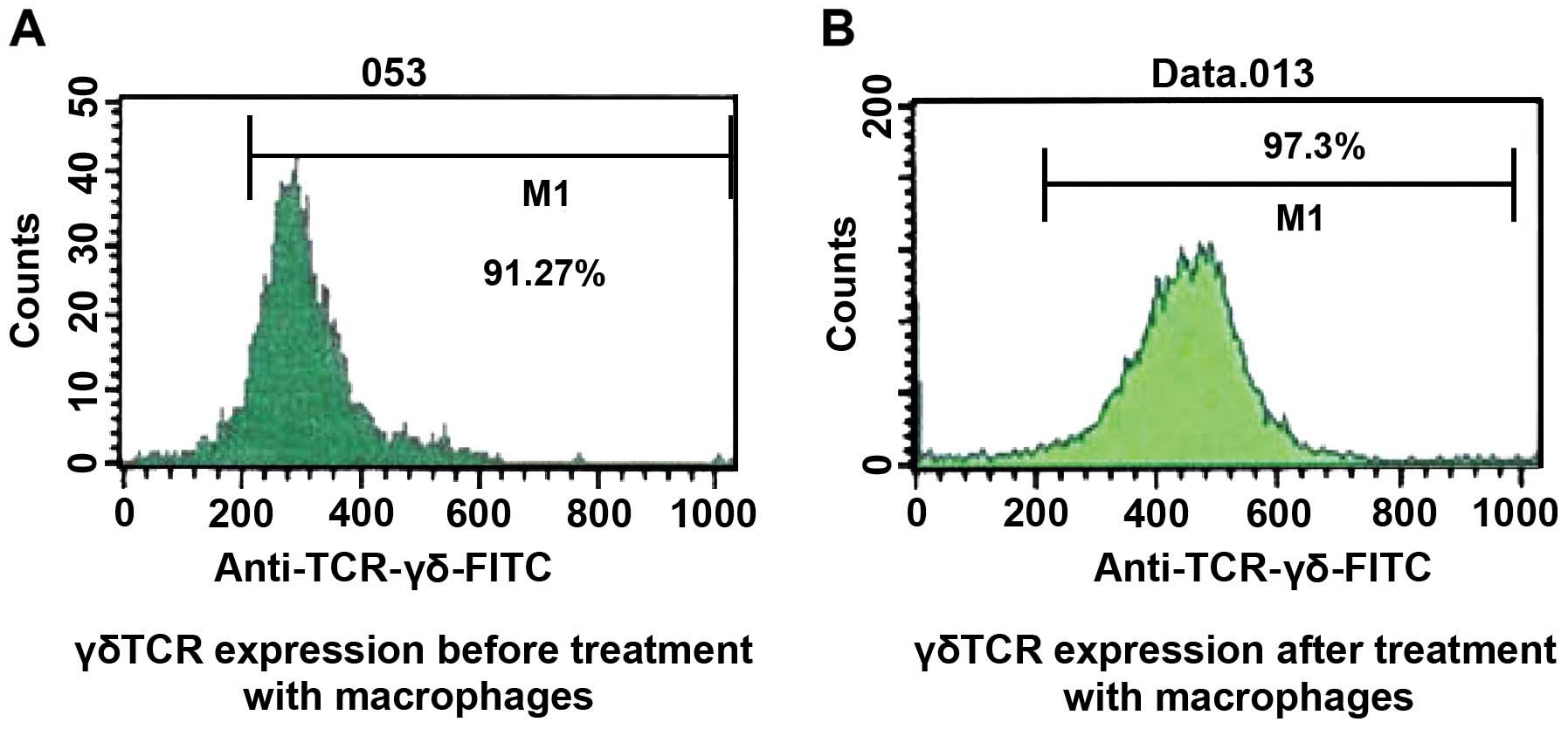
10 days, the percentage of γδT cells was 91.27% and the percentage
of expression of CD44 was 94.00% a reliable indicator of the purity
of the γδT cells (Figs. 1 and
2).
Cultures, Mϕ1 phenotype and assessment
of cell purity
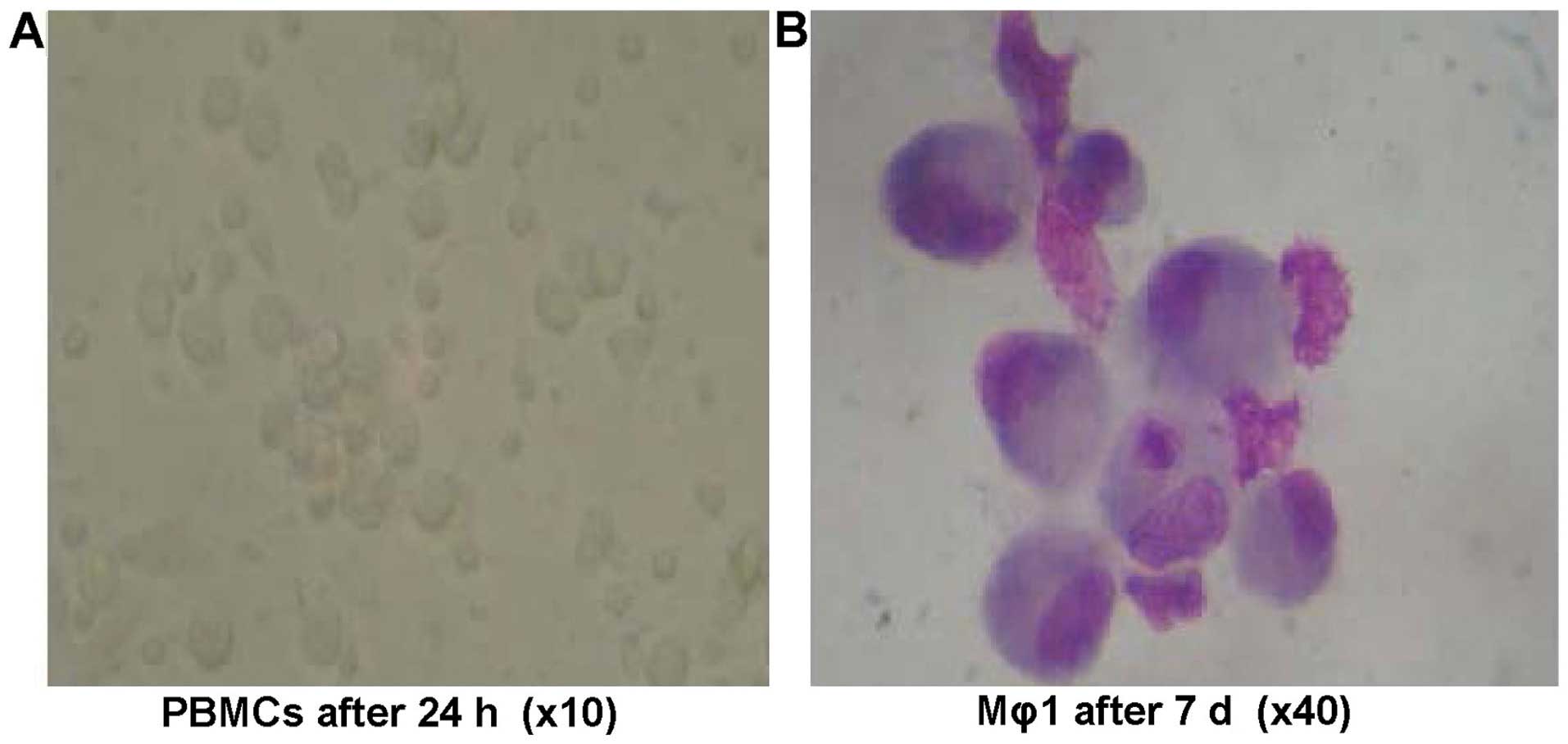
In the Mϕ1 induction system, PBMCs were shown to
have adherent growth after culturing for 24 h. Cell colonies of 3–6
cells became apparent after 48 h of cultivation. A bigger colony
and few single adherent cells were observed after 7 days of culture
and each individual cell appeared irregular in shape. Morphological
characteristics of macrophages were apparent after 7 days. The
cells appeared large in volume and oval or irregular in shape. The
nuclei of the cells were oval or irregular in shape. Loose
chromatin was present inside irregular nuclear membranes with
protrusions. The cytoplasms and pseudopodia were abundant and
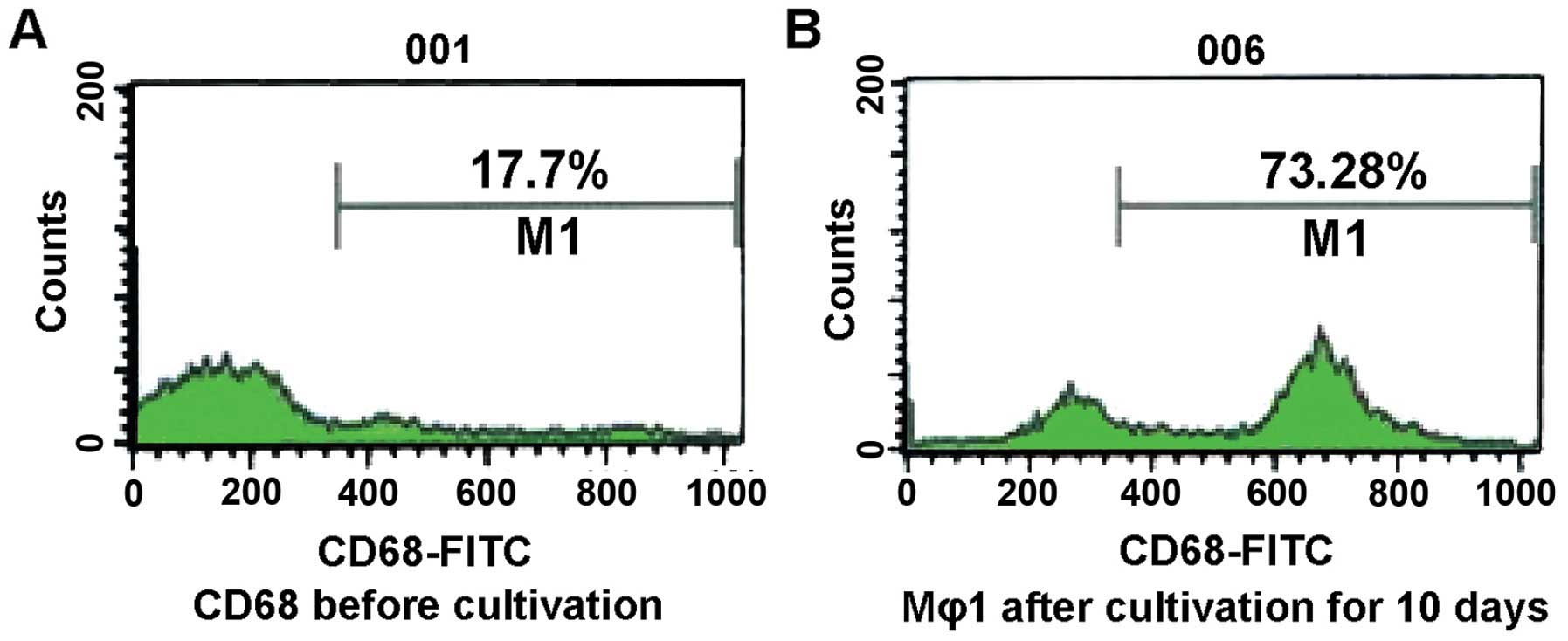
stained with gray blue. The percentage of expression on the cell
surfaces of CD68 among the cells was 17.7% before cultivation, and
73.2% after cultivation (Figs. 3 and
4).
Effect of Mϕ1 culture supernatant on
the proliferation of γδT cells
The proliferation of γδT cells in the test groups
that received different concentrations of Mϕ1 supernatant was
significantly higher than that in the control group which did not
receive any supernatant (P<0.01). Within the supernatant:medium
1:1 to 1:8 ratio groups, the proliferation of γδT cells increased
with decreasing concentrations of Mϕ1 supernatant, reaching a peak
(33.8% of cells) at the 1:8 ratio. The proliferation decreased by
small degrees when the volume ratios used were <1:16 (Table I).
 | Table I.Effect of Mϕ1 culture supernatant on
the proliferation of γδT cells. |
Table I.
Effect of Mϕ1 culture supernatant on
the proliferation of γδT cells.
| M:1640 (volume
ratio) | γδT cell
proliferation (A value) of the Mϕ1 culture group | γδT cell
proliferation percentage (%) of the Mϕ1 culture group |
|---|
| Control | 0.430±0.026 | 0.00 |
| 1:1 |
1.647±0.097a | 283.32a |
| 1:2 |
1.829±0.036a | 325.68a |
| 1:4 |
1.871±0.021a | 335.53a |
| 1:8 |
1.882±0.029a | 338.09a |
| 1:16 |
1.554±0.044a | 261.60a |
| 1:32 |
1.569±0.011a | 265.09a |
| 1:64 |
1.040±0.069a | 142.12a |
| 1:128 |
0.914±0.050a | 112.08a |
Effect of Mϕ1 culture supernatant on
the expression of the γδT cell surface marker γδTCR
The percentage of expression of the γδT cell surface
marker γδTCR was 97.3% after treatment with Mϕ1 culture
supernatant, while that of the control group was only 91.27%. This
difference was found to be statistically significant (P<0.05)
(Fig. 5).
Effect of Mϕ1 culture supernatant on
the tumor cytotoxic effect of γδT cells
The tumor cytotoxic effect of γδT cells in the test
groups treated with different concentrations of Mϕ1 supernatant was
always higher than the effect in the control groups (P<0.01).
Within the groups treated with 1:4 to 1:16 supernatant:medium
ratios the tumor-cytotoxic effect of γδT cells was higher in the
group treated with a 1:16 solution, reaching a peak (70.18%) at the
volume ratio of 1:16, while the proliferation ratio was decreased
slightly when the volume ratio used was <1:16 (Table II).
| Table II.Effect of Mϕ1 culture supernatant on
the tumor cytotoxic effect of γδT cells. |
Table II.
Effect of Mϕ1 culture supernatant on
the tumor cytotoxic effect of γδT cells.
| M:1640 | Mϕ1 culture
group | Control group |
|---|
| 1:4 |
61.16±2.11a | 46.12±1.23 |
| 1:8 |
65.23±1.23a | 45.55±0.96 |
| 1:16 |
70.18±0.94a | 47.25±1.02 |
| 1:32 |
59.94±1.36a | 46.21±0.79 |
Discussion
Macrophages have many functions including
phagocytosis, antigen presentation, and secretion of cytokines.
They occupy a central position within the cellular and molecular
networks composed of immune and non-immune cells and various
cytokines. Macrophages are also key players in the induction and
regulation of specific immune responses (16).
Baron-Bodo et al cultured Mϕ1 treating them
with GM-CSF and IFN-γ and proved that IFN-γ activates Mϕ1 and
enhances their antitumor effects (17). GM-CSF induces macrophage proliferation
and enhances their cancer cytotoxicity on cells in vitro. In
addition, GM-CSF induces the macrophages to secrete inflammatory
cytokines (18,19). CD68 is a transmembrane glycoprotein
with a molecular weight of 110 kDa that is distributed on the cell
surface of macrophages. Although there are only scarce CD68
glycoproteins on the surface of monocytes, their expression is
increased significantly as monocytes differentiate into
macrophages, allowing the marker to be used for the detection of
human macrophages (20). In the
present study, it was found that the expression of CD68 on the
surface of the cells had increased after cultivation. The
percentage of the cells carrying the CD68 marker increased from
17.7% to 73.2%. This finding suggests that our experiment
successfully transformed human PBMCs into macrophages.
γδT cells are an important subset of the innate
immunity population of T cells in vivo, and account for
5–15% of T cells in peripheral blood. γδT recognize various
antigens in an MHC unrestricted manner and are widely distributed
in the epithelial tissues of the digestive and respiratory tracts.
Nevertheless, there are some γδT cells circulating in peripheral
blood (21). Previous studies have
shown that γδT cells play an important role in tumor immune
surveillance and immunotherapy (22–26).
γδTCR, expressed on the surface of γδT cells, is a
receptor mainly associated with inflammation, tumor and other
immune responses. Dalton et al showed that the interaction
of macrophages and γδT cells is characteristic of the Vδ1 subtype
with induction of the TCR (27,28).
γδTCRs affect the antigen-presenting function, cytokine secretion
profile, inflammatory reaction and tumor immunity action of
macrophages. In addition, the cytotoxicity of γδT cells correlates
with the presence of ligands on tumor cells, such as MICA/MICB and
ULBPI-4. In the present study, treating γδT cells with Mϕ1 culture
supernatant resulted in an increased expression of the cell surface
marker γδTCR, suggesting that Mϕ1 can upregulate the expression of
γδTCR.
The MTT experiment showed that when γδT cell
cultures were treated with a Mϕ1 culture supernatant mixture, the
proliferation of the γδT cells was higher than the proliferation of
the control cells grown without Mϕ1 culture supernatant. The
proliferation of γδT cells reached a peak, becoming 33.8% of the
cells in the repertoire, at the supernatant:media volume ratio of
1:8. It was concluded that, the Mϕ1 supernatant promotes the growth
of γδT cells and provides a new approach for the improvement of
immunotherapy methods. Furthermore, the present findings showed
that Mϕ1 culture supernatant adds strength to the cell-killing
effect of γδT cells on human SGC-7901 gastric adenocarcinoma cells
in vitro. We suggest that the effect may be due to the
hyper-secretion by the Mϕ1 of Th1-promoting cytokines, such as
IL-12, which promote T-cell proliferation. IL-12 was first
described as a maturation factor of cytotoxic T lymphocytes and an
activator of the natural killer (NK) cells alone. However, the
variety of functions of IL-12 increased to include, inducing
peripheral blood lymphocytes to produce IFN-γ, enhancing the
cytotoxicity of NK cells, promoting the T-cell proliferation and
releasing the cytokines IFN-γ and TNF-α. Thus, IL-12 plays a role
in the immune killing of tumor cells. It is possible that IL-12,
which is present in the Mϕ1 supernatant, activates the γδTCR of γδT
cells, which in turn upregulates the expression of FasL, which
induces the apoptosis of gastric cancer cells. The present study
excluded the direct cell-cell contact effects by using only the
supernatant of activated macrophages and γδT cells.
The findings of the present study have shown that
activated macrophages can induce human γδT cells to kill SGC-7901
gastric cancer cells. Thus, the immunotherapeutic treatment of
gastric carcinoma combining activated macrophage supernatant and
γδT cells is a potentially successful anticancer strategy. Such
treatment may kill tumor cells immediately, bypassing the
restrictions imposed by MHC-restricted killing of cancer cells.
Additionally, it may improve the functions of the antitumor immune
network by ensuring the interaction between various immune cells
and enhancing the effect of cytokines. Nevertheless, the exact
mechanisms of action and the cell pathways involved remain to be
investigated prior to considering the implementation of such a
method in anticancer therapy.
References
|
1
|
Sun XD, Mu R, Zhou YS, Dai XD, Zhang SW,
Huangfu XM, Sun J, Li LD, Lu FZ and Qiao YL: Analysis of mortality
rate of stomach cancer and its trend in twenty years in China.
Zhonghua Zhong Liu Za Zhi. 26:4–9. 2004.(In Chinese). PubMed/NCBI
|
|
2
|
Patru CL, Surlin V, Georgescu I and Patru
E: Current issues in gastric cancer epidemiology. Rev Med Chir Soc
Med Nat Iasi. 117:199–204. 2013.PubMed/NCBI
|
|
3
|
Khan MI, Baqai MT, Bukhari M and Hashmi
RI: Gastric carcinoma: 5 years survival after gastric surgery. J
Pak Med Assoc. 55:158–160. 2005.PubMed/NCBI
|
|
4
|
Dunn GP, Old LJ and Schreiber RD: The
three Es of cancer immunoediting. Annu Rev Immunol. 22:329–360.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verreck FA, de Boer T, Langenberg DM,
Hoeve MA, Kramer M, Vaisberg E, Kastelein R, Kolk A, de
Waal-Malefyt R and Ottenhoff TH: Human IL-23-producing type 1
macrophages promote but IL-10-producing type 2 macrophages subvert
immunity to (myco)bacteria. Proc Natl Acad Sci USA. 101:4560–4565.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kabelitz D, Wesch D and He W: Perspectives
of gammadelta T cells in tumor immunology. Cancer Res. 67:5–8.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Casetti R and Martino A: The plasticity of
gamma delta T cells: innate immunity, antigen presentation and new
immunotherapy. Cell Mol Immunol. 5:161–170. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Born WK, Reardon CL and O'Brien RL: The
function of gammadelta T cells in innate immunity. Curr Opin
Immunol. 18:31–38. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Harris KM: Monocytes differentiated with
GM-CSF and IL-15 initiate Th17 and Th1 responses that are
contact-dependent and mediated by IL-15. J Leukoc Biol. 90:727–734.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barilli A, Rotoli BM, Visigalli R,
Bussolati O, Gazzola GC, Kadija Z, Rodi G, Mariani F, Ruzza ML,
Luisetti M and Dall'Asta V: In lysinuric protein intolerance system
y+l activity is defective in monocytes and in GM-CSF-differentiated
macrophages. Orphanet J Rare Dis. 5:322010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Moser B and Brandes M: Gammadelta T cells:
an alternative type of professional APC. Trends Immunol.
27:112–118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kobayashi H, Tanaka Y, Yagi J, Osaka Y,
Nakazawa H, Uchiyama T, Minato N and Toma H: Safety profile and
anti-tumor effects of adoptive immunotherapy using gamma-delta T
cells against advanced renal cell carcinoma: a pilot study. Cancer
Immunol Immunother. 56:469–476. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bennouna J, Bompas E, Neidhardt EM,
Rolland F, Philip I, Galéa C, Salot S, Saiagh S, Audrain M, Rimbert
M, et al: Phase-I study of Innacell gammadelta, an autologous
cell-therapy product highly enriched in gamma9delta2 T lymphocytes,
in combination with IL-2, in patients with metastatic renal cell
carcinoma. Cancer Immunol Immunother. 57:1599–1609. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen FX, Liu JQ, Xia F, Zhang J and Zhang
S: A new method for the amplification of human γδT cells. Chin Cell
Mol Immunol. 23:662–664. 2007.(In Chinese).
|
|
15
|
Liu JQ: The detection of LAK cell activity
by LDH kit. Chin J Clin Lab Sci. 13:831995.(In Chinese).
|
|
16
|
Smith PD, Ochsenbauer-Jambor C and
Smythies LE: Intestinal macrophages: Unique effector cells of the
innate immune system. Immunol Rev. 206:149–159. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baron-Bodo V, Doceur P, Lefebvre ML,
Labroquère K, Defaye C, Cambouris C, Prigent D, Salcedo M, Boyer A
and Nardin A: Anti-tumor properties of human-activated macrophages
produced in large scale for clinical application. Immunobiology.
210:267–277. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wallace PK, Romet-Lemonne JL, Chokri M,
Kasper LH, Fanger MW and Fadul CE: Production of
macrophage-activated killer cells for targeting of glioblastoma
cells with bispecific antibody to FcgammaRI and the epidermal
growth factor receptor. Cancer Immunol Immunother. 49:493–503.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Solinas G, Germano G, Mantovani A and
Allavena P: Tumor-associated macrophages (TAM) as major players of
the cancer-related inflammation. J Leukoc Biol. 86:1065–1073. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gordon S: Alternative activation of
macrophages. Nat Rev Immunol. 3:23–35. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kondo M, Izumi T, Fujieda N, Kondo A,
Morishita T, Matsushita H and Kakimi K: Expansion of human
peripheral blood gammadelta T cells using zoledronate. J Vis Exp.
Sep 9–2011.(Epub ahead of print). View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Beck BH, Kim HG, Kim H, Samuel S, Liu Z,
Shrestha R, Haines H, Zinn K and Lopez RD: Adoptively transferred
ex vivo expanded gammadelta-T cells mediate in vivo antitumor
activity in preclinical mouse models of breast cancer. Breast
Cancer Res Treat. 122:135–144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kondo M, Sakuta K, Noguchi A, Ariyoshi N,
Sato K, Sato S, Sato K, Hosoi A, Nakajima J, Yoshida Y, et al:
Zoledronate facilitates large-scale ex vivo expansion of functional
gammadelta T cells from cancer patients for use in adoptive
immunotherapy. Cytotherapy. 10:842–856. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuasa T, Sato K, Ashihara E, Takeuchi M,
Maita S, Tsuchiya N, Habuchi T, Maekawa T and Kimura S:
Intravesical administration of gammadelta T cells successfully
prevents the growth of bladder cancer in the murine model. Cancer
Immunol Immunother. 58:493–502. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Viey E, Fromont G, Escudier B, Morel Y, Da
Rocha S, Chouaib S and Caignard A: Phosphostim-activated gamma
delta T cells kill autologous metastatic renal cell carcinoma. J
Immunol. 174:1338–1347. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han LY, Fei SJ, Chen FX, Liu JQ and Chen
GL: Effect of zoledronate on human γδT cells killing gastric cancer
cell lines SGC-7901. World Chin Digestol. 17:181–185. 2009.(In
Chinese). View Article : Google Scholar
|
|
27
|
Dalton JE, Pearson J, Scott P and Carding
SR: The interaction of gamma delta T cells with activated
macrophages is a property of the V gamma 1 subset. J Immunol.
171:6488–6494. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dalton JE, Howell G, Pearson J, Scott P
and Carding SR: Fas-Fas ligand interactions are essential for the
binding to and killing of activated macrophages by gamma delta T
cells. J Immunol. 173:3660–3667. 2004. View Article : Google Scholar : PubMed/NCBI
|