Introduction
Osteosarcoma is the most common primary malignant
bone tumor, and usually occurs in adolescents and children (peak of
incidence, ~18 years old) (1). The
cancer causes tremendous disfiguration as a result of amputation,
and has a high morbidity and mortality rate (2). For these reasons, it is of great
importance to clarify the mechanisms that underlie the cause,
occurrence and development of osteosarcoma in order to identify
more effective approaches for its treatment. Systemic chemotherapy
following surgical removal of the tumor has been an effective
therapeutic method for the treatment of osteosarcoma (3). However, major problems, including
cytotoxic side effects and drug resistance, are associated with
chemotherapy (4,5). Thus, safe and more effective anti-cancer
treatments are required for patients with osteosarcoma.
Ezrin is currently considered one of the reasonable
and effective targets for cancer gene therapy (6). Ezrin, a membrane cytoskeletal
cross-linker, belongs to the ezrin/radixin/moesin protein family,
and is involved in the regulation of the cell cycle, cell
proliferation, cell differentiation and apoptosis (6,7). Ezrin
protein expression was reported to be significantly increased in
osteosarcoma tumors, and its levels are negatively correlated with
patient 5-year survival rates (8–10). As a
potential effective target, the silencing or downregulation of
ezrin expression may be an effective approach to suppress tumor
cell proliferation and to improve patient survival rate (11,12).
It is difficult to completely knockout or inhibit
the expression of a single gene to kill tumor cells by current
transgenic technologies, which explains why incomplete tumor
removal, recurrence and metastasis remain a challenge in tumor
treatment (13). Therefore, extensive
efforts have been paid to the selection of a combination of target
genes to achieve a better curative effect (14,15). Heat
shock protein (HSP)70, which is an adenosine triphosphate-dependent
molecular chaperone, regulates protein conformation, stability and
interactions (16). The majority of
HSP70 ligands are proteins essential for cell survival and growth,
including protein kinases, steroid receptors and transcription
factors (16,17). In addition, when tissue damage or
tumorigenesis occurs, HSPs are abundantly expressed, and form
complexes with peptides (18). If the
peptides are generated in normal tissues, the HSP70-peptide
complexes do not induce an immune response; however, if the
peptides are tumor-derived mutated antigens, the HSP70-peptide
complexes can be presented effectively to immune cells, thus
breaking the immune tolerance and inducing the tumor killing
effects of T lymphocytes and natural killer (NK) cells (18,19).
Therefore, it was hypothesized that inhibiting
proliferation and promoting apoptosis of tumor cells by
simultaneously providing exogenous danger signals such as
overexpressing HSPs would promote the recognition and presentation
of tumor antigenic peptides by antigen-presenting cells (APCs)
(20), enhance the immunogenicity of
tumor-associated antigens (21) and
tumor-specific antigens (22) derived
from membrane molecules of apoptotic tumor cells, and induce the
active anti-tumor immune response mediated by specific T
lymphocytes, thus enhancing their killing effects on tumor cells
and removing the remaining tumor cells in patients. Recently, the
present authors observed that simultaneously knocking down ezrin
and overexpressing HSP70 promoted the apoptosis and inhibited the
proliferation of human osteosarcoma cells (23). Based on the unique features of ezrin
and HSP70, a specific vector was designed and constructed in the
present study to simultaneously knock down ezrin expression and
upregulate HSP70 expression. Stably transfected LM8 osteosarcoma
cell lines with this vector were established and used to analyze
the influence of ezrin-small hairpin (sh)RNA in combination with
HSP70 on cell growth, proliferation, apoptosis and HSP70-induced
cytotoxic T lymphocyte (CTL) activity in vitro. In addition,
the suppression of proliferation and the tumor killing effects on
LM8 cells were assessed. Furthermore, tumor-bearing mice were
prepared by injection with the stably transfected cells, and the
inhibitory effects of ezrin knock-down in combination with
HSP70-induced immune response on tumor growth in vivo were
analyzed. The results obtained in the present study provide the
basis for a novel method of gene therapy for osteosarcoma based on
suppressing the proliferation and promoting the apoptosis of tumor
cells, in addition to inducing dual effects of specific immune
response.
Materials and methods
Cell culture
The murine osteosarcoma cell line LM8 was purchased
from the China Center for Type Culture Collection (Wuhan, China),
and was cultured in Dulbecco's modified Eagle medium (DMEM;
HyClone; GE Healthcare Life Sciences, Logan, UT, USA) supplemented
with 10% fetal bovine serum (HyClone; GE Healthcare Life Sciences)
at 37°C in a 5% CO2 incubator.
Vector construction and transient
transfection
Ezrin-shRNA containing a hairpin loop was designed
according to the ezrin messenger (m)RNA complementary (c)DNA
sequence (GenBank, BC013903.2; http://www.ncbi.nlm.nih.gov/gene/22350). The sequences
of the primers used were
5′-TGTATGAGCCTGTGAATT-TTCAAGAGA-AATTCACAGGCTCATACATT-3′ and
5′-TGGAGGCCAAAGTACCACAC-3′, in which there was a recognition site
for BamHI at the 5′ end and a recognition site for HindIII at the
3′ end. The sequences were synthesized and cloned into the
pGFP-V-RS vector (OriGene Technologies, Inc., Rockville, MD, USA)
to generate the pGFP-V-RS-shRNA vector. Then, the vector was
transformed into JM10 cells (obtained from China Center for Type
Culture Collection, Wuhan, China), which were amplified and
selected by puromycin resistance. Sequence identification of the
ezrin gene cloned in the vector was performed by Invitrogen (Thermo
Fisher Scientific, Inc., Waltham, MA, USA).
The HSP70 DNA sequence was synthesized according to
its mRNA sequence (GenBank, NM_010478.2). Then, the green
fluorescent protein (GFP) coding sequence in the pGFP-V-RS vector
was substituted by the GFP-HSP70 coding sequence by enzymatic
digestion and ligation in order to generate the pGFP-V-RS-HSP70
vector, which was transformed into DH5α cells (Gene Company Ltd.,
Shanghai, China), amplified and selected. Sequence identification
of the HSP70 gene cloned in the vector was performed by Invitrogen
(Thermo Fisher Scientific, Inc.). A similar method was used to
construct the pGFP-V-RS-shRNA-HSP70 vector.
Vectors, including empty vector pGFP-V-RS (control
group), pGFP-V-RS-shRNA (shRNA group) and pGFP-V-RS-shRNA-HSP70
(dual group), were transfected into LM8 cells using Lipofectamine
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The average transfection efficiency was
50–70%. Cells were allowed to recover in medium for 24 h after
transfection. All experiments were performed in 6-well tissue
culture plates with cells plated to reach 50–60% confluence on the
day of transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from cultured cells was isolated using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
260/280 absorbance ratio was measured for verification of the
purity of RNA. RNA samples were reverse transcribed into cDNA using
M-MLV Reverse Transcriptase (Promega Corporation, Madison, WI,
USA). The sequences of the ezrin, HSP70 and 18S ribosomal (r)RNA
genes were obtained from the GenBank database, and specific primers
for them were designed by Primer Premier 5.0 software (Premier
Biosoft International, Palo Alto, CA, USA). The following human
primers were used: Ezrin, forward 5′-ACTCACCAGAAACCGAAAATG-3′ and
reverse 5′-TGGAGGCCAAAGTACCACAC-3′; HSP70, forward
5′-AAGAGCAACAGCAGCAGACA-3′ and reverse 5′-CGATTGGCAGGTCCACAGTA-3′;
and 18S rRNA, forward 5′-CCTGGATACCGCAGCTAGGA-3′ and reverse
5′-GCGGCGCAATACGAATGCCCC-3′. RT-qPCR was performed using SYBR Green
qPCR SuperMix (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol, and glyceraldehyde
3-phosphate dehydrogenase (GAPDH) served as an internal control.
The thermal cycling conditions were as follows: 95°C for 3 min, 25
cycles of 95°C for 30 sec, 55°C for 2 min and 72°C for 30 sec, and
a final step of 72°C for 5 min. qPCRs for all samples were repeated
three times.
Western blot analysis
The cells were washed twice in ice-cold
phosphate-buffered saline (PBS) and resuspended in 5 v/v of
ice-cold lysis buffer [20 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid-KOH, 1.5 mM
MgCl2, 1 mM ethylenediaminetetraacetic acid, 1 mM
ethylene glycol-bis(β-aminoethyl ether)-N,N,N',N'-tetraacetic acid,
1 mM dithiothreitol and 0.1 mM phenylmethanesulfonyl fluoride (pH
7.5)]. The resuspended cells were homogenized with 10 pulses in a
Teflon® homogenizer to extract the total protein.
Protein samples (10 µg) were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis in 12 and 8%
polyacrylamide gels. The separated proteins were then
electrotransferred to a polyvinylidene fluoride membrane. Upon
blocking in 5% non-fat milk for 1 h, the membrane was incubated at
room temperature for 1 h with primary antibodies against ezrin
(1:1,000; #3145), HSP70 (1:1,000; #4872) (CST Biological Reagents
Company Limited, Shanghai, China), GAPDH (1:2,000; #TA309157;
Shanghai KangChen Bio-tech Inc., Shanghai, China), B-cell lymphoma
(Bcl)-2 (1:1,000; #2872), Bcl-2 associated X protein (Bax; 1:1,000;
#2772) and cyclin D1 (1:1,000; #2922) (CST Biological Reagents
Company Limited). The membrane was washed with Tris-buffered saline
and Tween 20 (TBST) three times for 5 min each, and incubated at
room temperature for 2 h in TBST containing horseradish peroxidase
(HRP)-conjugated goat anti-rabbit immunoglobulin G antibody
(1:20,000; SouthernBiotech, Birmingham, AL, USA). The membrane was
next washed with TBST three times for 10 min each, and then
incubated for 30 sec with Pierce ECL Western Blotting Substrate
(Thermo Fisher Scientific, Inc.) reagent for the development of the
HRP signals, followed by exposure to autoradiography film for
visualization of the bands. GAPDH was used as a loading
control.
Analysis of apoptosis
Cellular apoptosis was determined using an annexin
V-fluorescein isothiocyanate (FITC) apoptosis detection kit
(Clontech Laboratories Inc., Mountainview, CA, USA). Brieflly,
cells were cultured at a density of 4×106 cells/ml and
seeded in 6-well plates. Cells were harvested by trypsinization,
washed twice with cold PBS and centrifuged at 100 × g. Cells
(1×105-1×106) were then resuspended in 300 µl
1X binding buffer and centrifuged again at 100 × g for 5 min. The
supernatant was next removed, and 10 µl annexin V-FITC was added to
the cells, which were incubated in the dark for 30 min at room
temperature. Subsequently, cells were incubated in the dark with 5
µl propidium iodide, and analyzed by flow cytometry (BD
FACSCalibur™; BD Biosciences, Franklin Lakes, NJ, USA). The test
for each sample was repeated three times, and data were represented
as the mean value.
Analysis of cell proliferation
Cell proliferation was determined by
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
(MTT) assay. The transfected cells were plated in 96-well plates at
a density of 0.1–0.2×104 cells/well. Next, 20 µl
MTS/phenazine methosulfate mixture was added to each well, and
cells were incubated for 3–4 h. The absorbance was then determined
at an optical density (OD) of 570 nm. The cell proliferation was
measured over 7 days. The experiment was repeated three times, and
data were represented as the mean absorbance value. The cell growth
curve was represented to compare the growth rates upon
transfection, and the proliferation rates and proliferation
inhibitory rates were calculated.
Preparation of mouse spleen
lymphocytes
BALB/c mouse (Yunnan Animal Center, Kunming, China)
spleens were removed under non-sterile conditions (25°C, cycle of
day/night of 12 h/12 h, fed twice per day), cut into small pieces
with sterile scissors and pushed through a stainless steel screen
(100-mesh) in Hank's solution (pH 7.2–7.6). The spleen cell
suspension was prepared and used to isolate mononuclear cells
(lymphocytes and monocytes) with a lymphocyte isolation solution
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany). The isolated
cells were suspended at a concentration of 1×106
cells/ml in RPMI 1640 supplemented with 15% newborn calf serum
(Thermo Fisher Scientific, Inc.). Spleen lymphocytes
(1×106) were resuspended in the culture supernatant of
LM8 cells that had been stably transfected with
pGFP-V-RS-shRNA-HSP70. Recombinant interleukin-2 (rIL-2) was added
to the mixture at 2,000 U/ml. After culture at 37°C for 7 days, the
cells were harvested and used as sensitized mouse spleen CTL.
Control spleen lymphocytes were isolated as described above and
treated with rIL-2 (2,000 U/ml) but without the culture
supernatant.
CTL killing assays
LM8 osteosarcoma cells in logarithmic growth phase
were harvested and suspended at a concentration of 2×106
cells/ml in RPMI 1640 medium, serving as target (T) cells. Upon
amplification, the induced specific CTLs, including the sensitized
and non-sensitized spleen lymphocytes, served as effector (E)
cells. The E and T cells were mixed at 100:1, 50:1 and 25:1 (E:T)
ratios, and seeded in 96-well plates (200 µl/well). For controls,
100 µl/well E lymphocytes or T cells were seeded, while the blank
control contained medium without cells. Sample evaluation was
repeated four times. Briefly, the plate was incubated at 37°C in a
5% CO2 incubator for 20 h, and then 150 µl
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
solution (0.5 mg/ml in serum-free medium) was added to each well.
After incubation at 37°C for 4 h, the MTT solution was discarded,
and 100 µl dimethyl sulfoxide was added to each well. The plate was
placed on a horizontal agitator for 10 min, and the absorbance was
determined at OD 570 nm using a microplate reader. The mean value
of the four repeated tests was used, and the killing effect was
calculated using the following equation:
Killingeffect=(1–ODexp–ODctrEODctrT)×100%,
where ODexp is the absorbance value of the
experimental wells, ODctrE represents the signal from the control
wells of E cells and ODctrT represents the signal from the control
wells of T cells.
Tumor formation in BALB/c nude
mice
The LM8 murine osteosarcoma cells, which were stably
transfected with the pGFP-V-RS, pGFP-V-RS-shRNA or
pGFP-V-RS-shRNA-HSP70 vectors, were harvested in the logarithmic
growth phase. The cells were centrifuged in a 10-ml tube at 90 × g
for 10 min, and the supernatants were removed. Then, the cells were
resuspended in serum-free DMEM (5×107 cells/ml). A total
of 18 BALB/c mice (4–6 week-old, males and females) were divided
into three groups (n=6/group). The cell suspension (0.1 ml) was
injected subcutaneously to the right sides of the back of the mice
in the three groups, and tumor formation was observed every other
day, with the longest (D) and shortest (d) diameter of the tumor
being measured. The tumor volume was calculated using the following
equation: Volume = Dxd2/2.
After 40 days, the tumors were removed, weighed and
fixed in 10% neutral formalin, followed by hematoxylin and eosin
(H&E) staining.
Analysis of the splenic T cell
population in tumor-bearing mice
Blood samples from tumor-bearing mice were obtained
from the orbital sinus. Tumor-bearing mice were sacrificed by
cervical dislocation, and their spleens were removed with tweezers,
placed on a steel mesh, cut into small pieces with scissors and
ground with a mortar using 2–3 ml 1X PBS. The liquid was passed
through the mesh into a culture dish and transferred to a 15-ml
tube. After natural sedimentation for 3–5 min, the supernatant was
transferred to a new 15-ml tube, and the sediment was discarded.
Upon centrifugation at 200 × g for 6 min, the supernatants were
removed, and 1 ml double distilled H2O was added to the
sediment and mixed. After 30 sec, 1 ml 2X PBS was added, and the
tube was centrifuged at 200 × g for 6 min. The supernatant was
removed, and the sediment was resuspended in 1.5 ml PBS and then
divided into 250 µl aliquots in 6 centrifuge tubes
(~1×105 cells/tube). After centrifugation at 200 × g for
6 min, the supernatants were removed, and 100 µl of the
corresponding fluorescent-labeled antibody was added to each tube.
For the blank control, 100 µl 1X PBS was added instead. Upon
mixing, the cells were placed on ice away from light for 30 min and
centrifuged at 2,000 rpm for 6 min. The supernatants were removed,
and 300 µl 1X PBS was added to each tube. Then, the cells were
analyzed by flow cytometry.
Enzyme-linked immunosorbent assay
(ELISA) analysis of IL-4 and interferon (IFN)-γ
The serum samples obtained from tumor-bearing mice
in the control, shRNA and the dual groups were analyzed using an
IL-4 or IFN-γ ELISA kit (Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. The experiment was repeated three
times.
Ethics statement
The present study was undertaken according to the
protocol approved by the constituted Ethics Committee of Xiamen
University (Xiamen, China), which conforms to the principles of the
Declaration of Helsinki.
Statistical analysis
Statistical analysis was performed using SPSS 17.0
software (SPSS, Inc., Chicago, IL, USA). Data were expressed as the
mean ± standard deviation. Multiple comparisons were evaluated by
analysis of variance. For normally distributed data, the t
test was used for comparisons between groups; for non-normally
distributed data, the Dunnett's t test was used for
comparisons between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of ezrin and HSP70 in
transfected LM8 cells
In the shRNA and dual groups, the ezrin mRNA level
was significantly lower than that in the control group (P<0.01).
The RNA expression of ezrin was knocked down by >99%. Western
blot analysis also revealed that ezrin protein expression was
markedly decreased by ezrin-shRNA (Fig.
1A). The HSP70 mRNA level was significantly increased in the
ezrin-shRNA/HSP70 group compared with the ezrin-shRNA and control
groups (P=0.005). HSP70 protein overexpression was also confirmed
by western blot analysis (Fig.
1B).
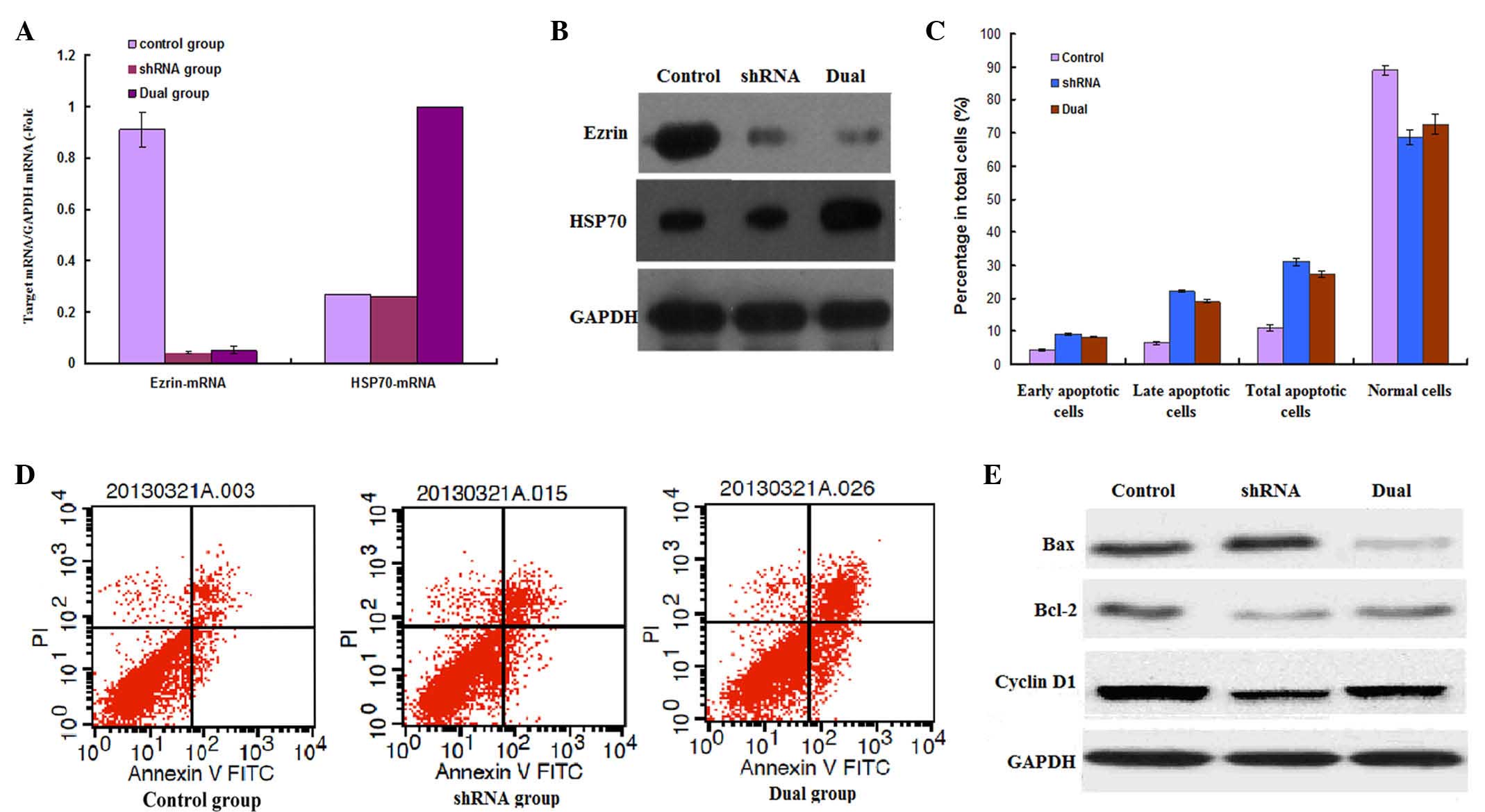 | Figure 1.mRNA and protein expression of ezrin
and HSP70 in transfected LM8 cells were detected by (A) reverse
transcription-quantitative polymerase chain reaction and (B)
western blotting, respectively. (C) Cellular apoptosis was analyzed
by detecting the number of early and late apoptotic cells, as well
as normal cells, in the NC, ezrin-shRNA and ezrin-shRNA/HSP70
groups. (D) Apoptosis analysis of stably transfected LM8 cells was
performed by flow cytometry. (E) Apoptosis-associated proteins were
detected by western blotting. NC, negative control; mRNA, messenger
RNA; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; shRNA, small
hairpin RNA; HSP70, heat shock protein 70; Bcl-2, B-cell
lymphoma-2; Bax, Bcl-2 associated X protein; PI, propidium iodide;
FITC, fluorescein isothiocyanate. |
Ezrin-shRNA/HSP70 transfection
promotes the apoptosis of LM8 cells
As indicated in Fig. 1C
and D, the apoptosis rate of LM8 cells was significantly
increased in the shRNA group (31.56±1.10%) compared with the
control group (11.01±0.80%) (P=0.023), particularly the rate of
late apoptotic cells. When simultaneously overexpressing HSP70, the
apoptosis rate of LM8 cells was slightly decreased compared with
that of the shRNA group; however, the apoptosis rate (27.31±0.95%)
was still significantly higher than that in the control group
(11.01±0.80%) (P=0.002).
In addition, western blot analysis demonstrated that
ezrin-shRNA transfection promoted the expression of pro-apoptotic
Bax, whereas it suppressed the expression of anti-apoptotic Bcl-2
and cyclin D1. The ezrin-shRNA/HSP70 group had overall weaker but
similar effects regarding the expression of these proteins compared
with the shRNA group. There were obvious differences compared with
the control group (Fig. 1E).
Thus, stable transfection of ezrin-shRNA/HSP70
promoted the apoptosis of LM8 osteosarcoma cells.
Ezrin-shRNA/HSP70 transfection
suppresses cell proliferation of LM8 cells
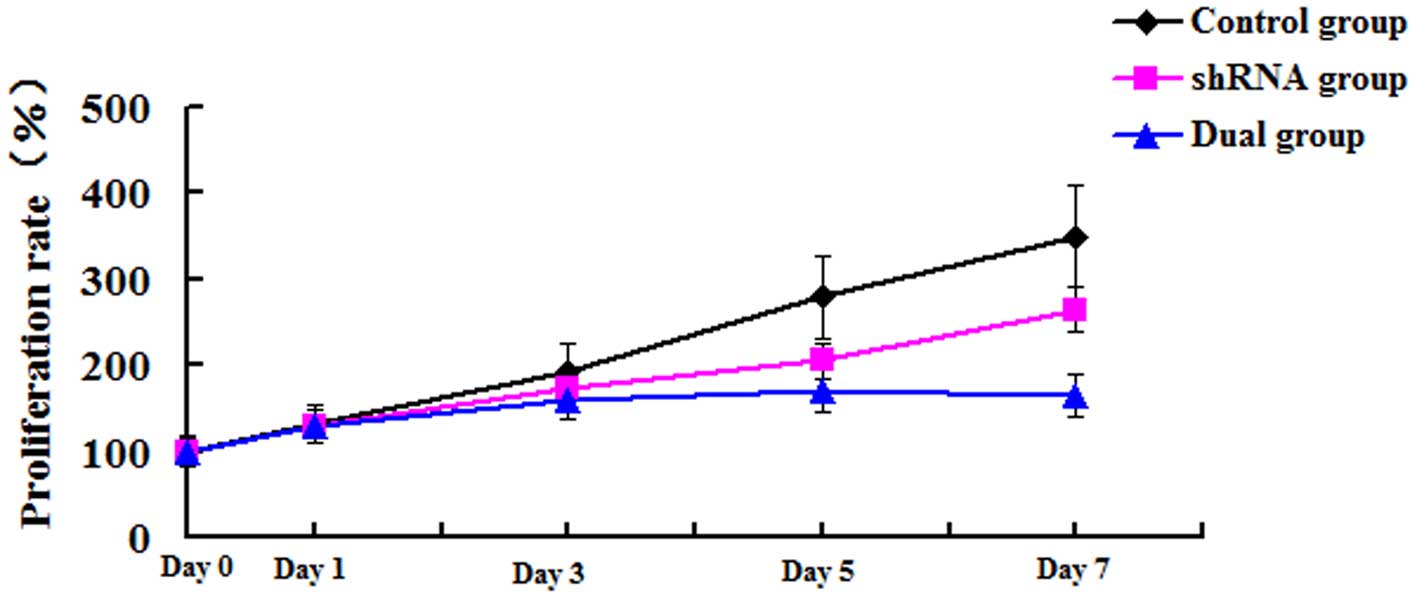
As represented in Fig.
2, the proliferation of LM8 cells in the three groups was
analyzed by MTT assay. The absorbance value at day 0 was set to be
100%. When compared with the control group, the proliferation rate
in the shRNA group was significantly decreased from day 3, and
reached a peak at day 7 (350.28±3.56 vs. 190.76±4.71%) (P=0.001).
The simultaneous reduction in ezrin expression and overexpression
of HSP70 slightly recovered the cell proliferation decreased by
ezrin knock-down. However, the proliferation rate was still
significantly decreased at day 7 (350.28±3.56 vs. 280.61±3.23%)
(P=0.003). Thus, stable transfection of ezrin-shRNA/HSP70
suppressed the proliferation of LM8 osteosarcoma cells.
Cytotoxic effects of HSP70-induced CTL
on LM8 cells
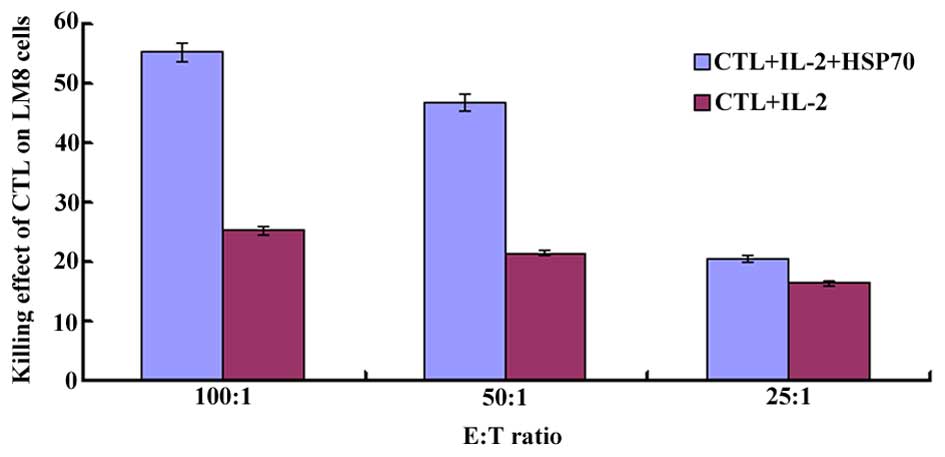
As indicated in Fig.
3, HSP70-induced CTL had a greater cytotoxic effect on LM8
cells, with a killing effect as high as 55.56±2.10%, in the dual
group. There were significant differences between the two
experimental groups (shRNA and dual groups) and the control group
(P=0.001). When comparing the CTL activity in different E:T ratio
groups, HSP70-induced CTL had the highest killing effect on LM8
cells in the 100:1 (E:T) group.
Ezrin-shRNA/HSP70 transfection
suppresses tumor formation in mice
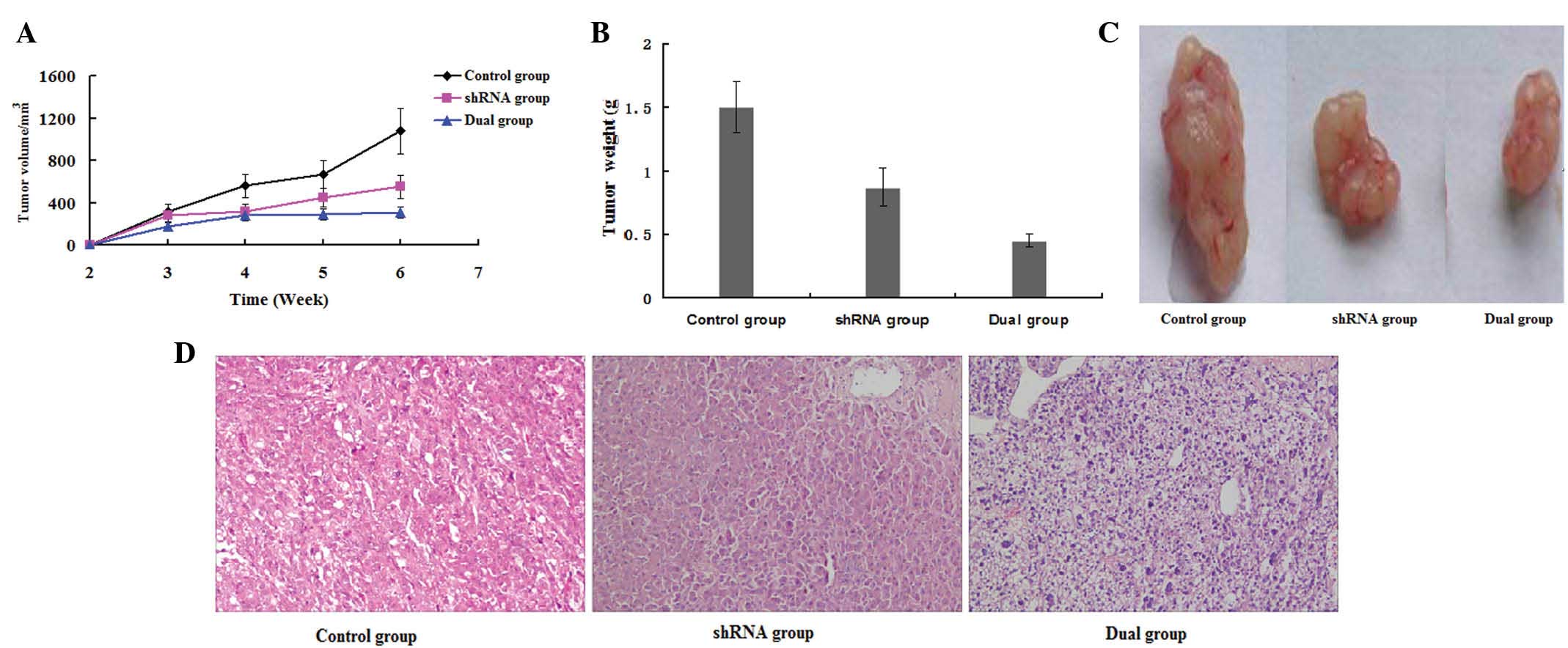
Upon injection of stably transfected LM8 cells into
nude mice, the tumor size and growth rate were analyzed. It was
observed that the tumor growth was suppressed in the dual group 2–3
weeks after injection. There were significant differences in tumor
growth rate compared with the shRNA and control groups (P=0.004)
(Fig. 4A). After 6 weeks, the mice
were sacrificed; the tumors were removed and weighed. The tumor
weights in the dual group were significantly lower than those in
the shRNA and control groups (P=0.029) (Fig. 4B and C).
Furthermore, histological examination of tumor
sections by H&E staining revealed stroma-rich tumors with
significantly smaller size and suppressed growth in the
ezrin-shRNA/HSP70 group, compared with the other two groups
(Fig. 4D). Thus, stable transfection
of ezrin-shRNA/HSP70 in LM8 osteosarcoma cells suppressed tumor
formation in nude mice.
Transfection of ezrin-shRNA/HSP70
increases cluster of differentiation (CD)8+ T cells in
tumor-bearing mice
As indicated in Table
I and Fig. 5, the percentage of
CD8+ T lymphocytes was increased, whereas the percentage
of CD4+ T lymphocytes was decreased, in the
ezrin-shRNA/HSP70 group, and the ratio of
CD4+/CD8+ T lymphocytes was significantly
smaller than that in the shRNA and control groups (P=0.006). There
were no significant differences between the shRNA group and the
control group (P=0.102).
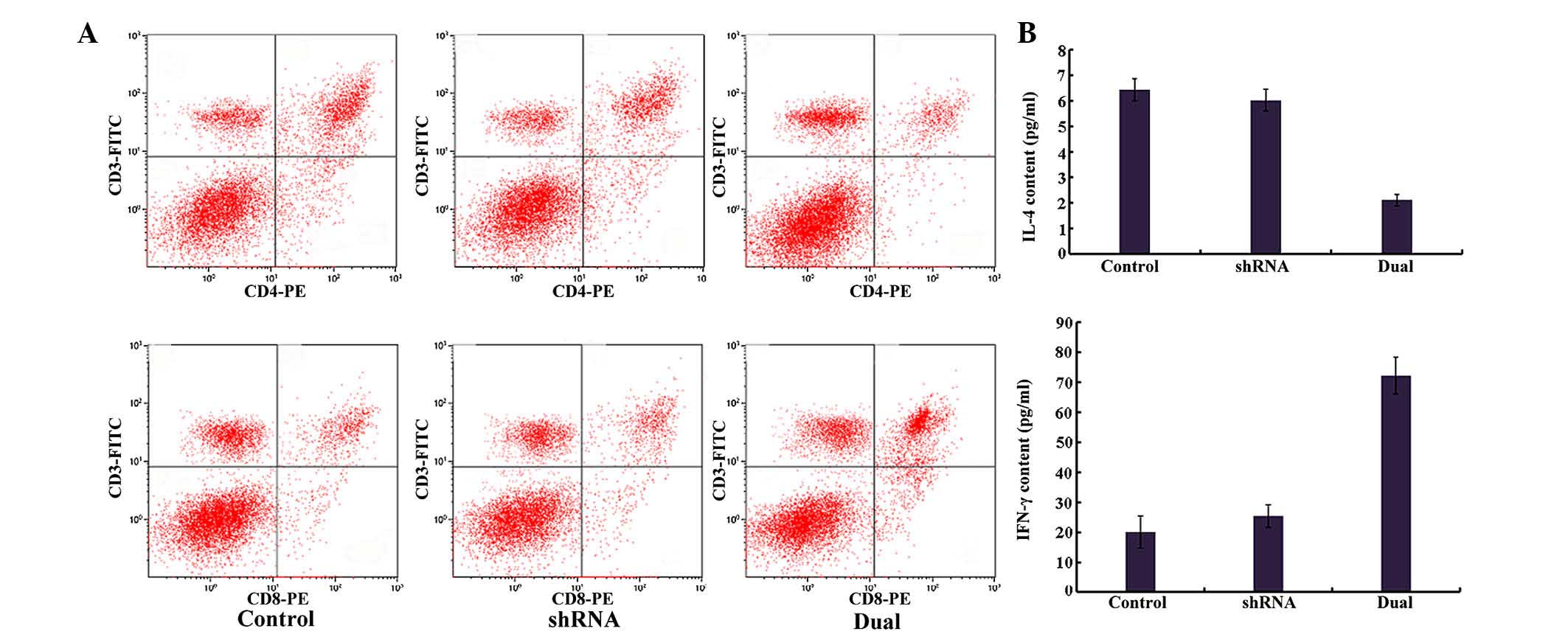 | Figure 5.Immunological analysis of
tumor-bearing mice. (A) Analysis of the splenic T cell population
from tumor-bearing mice. At 40 days after injection, spleens were
removed from the mice, and the splenic T cell population was
analyzed by flow cytometry. (B) Analysis of serum IL-4 and IFN-γ
levels in tumor-bearing mice. At 40 days after injection, the serum
samples were collected from the tumor-bearing mice, and the levels
of IL-4 and IFN-γ were determined by enzyme-linked immunosorbent
assay. CD, cluster of differentiation; IL, interleukin; IFN,
interferon; shRNA, small hairpin RNA; PE, phycoerythrin; FITC,
fluorescein isothiocyanate. |
 | Table I.Analysis of the T cell population of
the tumor-bearing mice. |
Table I.
Analysis of the T cell population of
the tumor-bearing mice.
| Group | CD4+/CD3+(%) | CD8+/CD3+(%) | CD4+/CD8+(ratio) |
|---|
| Control group | 23.56±2.30 | 18.29±2.10 | 1.279±0.036 |
| shRNA group | 22.49±1.60 | 19.14±2.30 | 1.182±0.052 |
| Dual group | 17.01±1.40 | 27.91±2.60 |
0.623±0.042a |
Transfection of ezrin-shRNA/HSP70
promotes immunological killing of tumors
As represented in Fig.
5B, in the dual group, the serum IL-4 level was significantly
decreased, and the serum IFN-γ level was significantly increased,
compared with the ezrin-shRNA and negative control (NC) groups
(P<0.01), whereas there were no significant differences in IL-4
or IFN-γ levels between the ezrin-shRNA group and the NC group
(P>0.05).
Discussion
It has been reported that downregulation of ezrin
expression by small interfering (si)RNA (12) or alteration of its phosphorylation
(10) resulted in the apoptosis of
osteosarcoma cells and a reduced survival rate of tumor cells.
However, the siRNA technique has certain disadvantages, including
low effectiveness and instability, whereas the shRNA technique can
establish stable, long-term gene silencing cell lines with high
success rates (24). In the present
study, shRNA eukaryotic expression vectors were used to
continuously downregulate ezrin expression. After 1 month of
selection, the LM8 cell lines that silenced ezrin gene expression
were established and confirmed by fluorescence microscopy. The
efficiency of gene silencing ezrin was >99%, indicating that the
shRNA expression vector is an efficient method to downregulate
ezrin gene expression. When comparing cells with or without gene
silencing, it was noticed that the apoptosis rate was significantly
increased, while the cell proliferation rate was significantly
reduced, upon reducing ezrin expression, and thus, the tumor cell
growth was effectively inhibited.
However, even though the ezrin gene was almost
completely silenced, the growth rate of LM8 cells was inhibited
only by 45.7%. In addition, the transgenic technology is unable to
transfect all tumor cells. Therefore, it is difficult to completely
remove all tumor cells, which is an important reason for incomplete
treatment, and ultimately leads to tumor recurrence and metastasis
(25). Wang et al reported
that immunization of mice with HSP70 extracted from tumor tissues
could induce specific CTL activity and tumor killing effects
(26). That study was performed in a
mouse model of colon cancer. In the current study, the pGFP-V-RS
vector, which contains a cytomegalovirus promoter and a U6
promoter, was constructed to simultaneously silence ezrin gene
expression while producing HSP70 overexpression, and to establish
stably transfected cell lines. HSP70 overexpression partially
recovered the promoted cellular apoptosis and proliferation
suppression by knocking down ezrin. However, when compared with the
normal control, HSP70 overexpression significantly inhibited the
tumor cell growth and induced the apoptosis of tumor cells. At the
same time, the overexpressed HSP70 protein could be released by the
apoptotic tumor cells and act as a danger signal, further inducing
the specific immune response against osteosarcoma cells, and thus,
was able to remove tumor cells to a greater extent.
In the present study, tumor-bearing mice models were
established by injecting the stably transfected murine LM8
osteosarcoma cells with pGFP-V-RS, pGFP-V-RS-shRNA and
pGFP-V-RS-shRNA-HSP70. The results demonstrated that mice in the
ezrin-shRNA/HSP70 group experienced higher anti-tumor effects than
those in the control group, since histopathological examination of
their tumor sections revealed more necrotic tissues and smaller
volumes of tumor cells compared with the control mice. Flow
cytometric analysis demonstrated that, in the ezrin-shRNA/HSP70
group, the percentage of CD8+ T lymphocytes in spleen
was higher, and the ratio of CD4+/CD8+ T
lymphocytes was lower, compared with the control group; this
suggests that HSP70 is important in tumor-antigen presentation and
CD8+ T lymphocyte activation. Serum levels of IL-4 and
IFN-γ in tumor-bearing mice were measured by ELISA, which revealed
that, in the ezrin-shRNA/HSP70 group, the IFN-γ level was
significantly increased, while the IL-4 level was significantly
decreased. This suggests that, once the antigen was presented, T
cells and NK cells were activated, thus secreting IFN-γ and
consequently activating macrophages, dendritic cells and NK cells,
and triggering cell-mediated immunity to inhibit tumor growth.
Whereas the humoral immunity was suppressed, the IL-4 level was
decreased. These results were consistent with our recent findings
in human osteosarcoma cells (23).
Overall, it was speculated that, in the tumor
tissues of mice in the ezrin-shRNA/HSP70 group, ezrin-shRNA induced
the release of overexpressed HSP70 protein from tumor cells; then,
the HSP70-bound tumor-derived mutated antigens and the
HSP70-peptide complexes were recognized by the pattern recognition
receptors on the surface of APCs and subsequently phagocytosed by
APCs. Thus, the immune tolerance was broken, and the tumor killing
effects of T lymphocytes and NK cells were induced.
In summary, the results of the present study
suggested that stable transfection of the specific
pGFP-V-RS-shRNA-HSP70 vector induced apoptosis and reduced the
proliferation rate of osteosarcoma cells, and in combination with
HSP70-induced cellular immune response, it also induced the
amplification of tumor killing effects. The present study has
assessed the effects of the ezrin-shRNA/HSP70 transfection method
on tumor treatment in vitro and in vivo, and provides
the theoretical and experimental basis for clinical application of
gene therapy for osteosarcoma.
References
|
1
|
Janeway KA, Barkauskas DA, Krailo MD,
Meyers PA, Schwartz CL, Ebb DH, Seibel NL, Grier HE, Gorlick R and
Marina N: Outcome for adolescent and young adult patients with
osteosarcoma: A report from the children's oncology group. Cancer.
118:4597–4605. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bacci G, Ferrari S, Longhi A, Donati D,
Manfrini M, Giacomini S, Briccoli A, Forni C and Galletti S:
Nonmetastatic osteosarcoma of the extremity with pathologic
fracture at presentation: Local and systemic control by amputation
or limb salvage after preoperative chemotherapy. Acta Orthop Scand.
74:449–454. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gobin B, Moriceau G, Ory B, Charrier C,
Brion R, Blanchard F, Redini F and Heymann D: Imatinib mesylate
exerts anti-proliferative effects on osteosarcoma cells and
inhibits the tumour growth in immunocompetent murine models. PLoS
One. 9:e907952014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arias JL: Drug targeting strategies in
cancer treatment: An overview. Mini Rev Med Chem. 11:1–17. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
De Santi C, Pietrabissa A, Mosca F and
Pacifici GM: Methylation of quercetin and fisetin, flavonoids
widely distributed in edible vegetables, fruits and wine, by human
liver. Int J Clin Pharmacol Ther. 40:207–212. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khanna C, Wan X, Bose S, Cassaday R, Olomu
O, Mendoza A, Yeung C, Gorlick R, Hewitt SM and Helman LJ: The
membrane-cytoskeleton linker ezrin is necessary for osteosarcoma
metastasis. Nat Med. 10:182–186. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ren L, Hong SH, Cassavaugh J, Osborne T,
Chou AJ, Kim SY, Gorlick R, Hewitt SM and Khanna C: The
actin-cytoskeleton linker protein ezrin is regulated during
osteosarcoma metastasis by PKC. Oncogene. 28:792–802. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y, Zhang L, Zhang G, Li S, Duan J,
Cheng J, Ding G, Zhou C, Zhang J, Luo P, et al: Osteosarcoma
metastasis: Prospective role of ezrin. Tumour Biol. 35:5055–5059.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin LJ and Chen LT: Association between
ezrin protein expression and the prognosis of colorectal
adenocarcinoma. Mol Med Rep. 8:61–66. 2013.PubMed/NCBI
|
|
10
|
Li L, Wang YY, Zhao ZS and Ma J: Ezrin is
associated with gastric cancer progression and prognosis. Pathol
Oncol Res. 17:909–915. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lo Vasco VR, Leopizzi M, Puggioni C and
Rocca C Della: Ezrin silencing remodulates the expression of
Phosphoinositide-specific Phospholipase C enzymes in human
osteosarcoma cell lines. J Cell Commun Signal. 8:219–229. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shang X, Wang Y, Zhao Q, Wu K, Li X, Ji X,
He R and Zhang W: siRNAs target sites selection of ezrin and the
influence of RNA interference on ezrin expression and biological
characters of osteosarcoma cells. Mol Cell Biochem. 364:363–371.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang T, Zhang H, Cai SY, Shen YN, Yuan SX,
Yang GS, Wu MC, Lu JH and Shen F: Elevated SHOX2 expression is
associated with tumor recurrence of hepatocellular carcinoma. Ann
Surg Oncol. 20(Suppl 3): S644–S649. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Murugesan SR, Akiyama M, Einfeld DA,
Wickham TJ and King CR: Experimental treatment of ovarian cancers
by adenovirus vectors combining receptor targeting and selective
expression of tumor necrosis factor. Int J Oncol. 31:813–822.
2007.PubMed/NCBI
|
|
15
|
Cai G, Zhu X, Xu Y, Du X, Zhang Z, Zhang
Y, Chen T, Zhou X, Guan Z and Cai S: Case report of extrarenal
rhabdoid tumor of pelvic retroperitoneum molecular profile of
angiogenesis and its implication in new treatment strategy. Cancer
Biol Ther. 8:417–421. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goloudina AR, Demidov ON and Garrido C:
Inhibition of HSP70: A challenging anti-cancer strategy. Cancer
Lett. 325:117–124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sliutz G, Karlseder J, Tempfer C, Orel L,
Holzer G and Simon MM: Drug resistance against gemcitabine and
topotecan mediated by constitutive hsp70 overexpression in vitro:
Implication of quercetin as sensitiser in chemotherapy. Br J
Cancer. 74:172–177. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sashchenko LP, Dukhanina EA, Shatalov YV,
Yashin DV, Lukyanova TI, Kabanova OD, Romanova EA, Khaidukov SV,
Galkin AV, Gnuchev NV and Georgiev GP: Cytotoxic T lymphocytes
carrying a pattern recognition protein Tag7 can detect evasive,
HLA-negative but Hsp70-exposing tumor cells, thereby ensuring
FasL/Fas-mediated contact killing. Blood. 110:1997–2004. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stangl S, Gross C, Pockley AG, Asea AA and
Multhoff G: Influence of Hsp70 and HLA-E on the killing of leukemic
blasts by cytokine/Hsp70 peptide-activated human natural killer
(NK) cells. Cell Stress Chaperones. 13:221–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zitzler S, Hellwig A, Hartl FU, Wieland F
and Diestelkötter-Bachert P: Distinct binding sites for the ATPase
and substrate-binding domain of human Hsp70 on the cell surface of
antigen presenting cells. Mol Immunol. 45:3974–3983. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nishikawa M, Otsuki T, Ota A, Guan X,
Takemoto S, Takahashi Y and Takakura Y: Induction of tumor-specific
immune response by gene transfer of Hsp70-cell-penetrating peptide
fusion protein to tumors in mice. Mol Ther. 18:421–428. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamaoka A, Guan X, Takemoto S, Nishikawa M
and Takakura Y: Development of a novel Hsp70-based DNA vaccine as a
multifunctional antigen delivery system. J Control Release.
142:411–415. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yao Q, Zhao HY and Xie BZ: effects of
ezrin and heat shock protein 70 on apoptosis and proliferation of
human osteosarcoma cells. Orthop Surg. 7:273–280. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rácz Z and Hamar P: SiRNA technology, the
gene therapy of the future? Orv Hetil. 149:153–159. 2008.(In
Hungarian). View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wirtzfeld LA, Wu G, Bygrave M, Yamasaki Y,
Sakai H, Moussa M, Izawa JI, Downey DB, Greenberg NM, Fenster A, et
al: A new three-dimensional ultrasound microimaging technology for
preclinical studies using a transgenic prostate cancer mouse model.
Cancer Res. 65:6337–6345. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang ZH, Ye Q, Hu ZQ, Ye ZQ, Yu X and Shen
GX: In vitro anti-tumor effect of CTL induced by HSP70-Id
complex-modified dendritic cells. Zhonghua Zhong Liu Za Zhi.
28:481–485. 2006.(In Chinese). PubMed/NCBI
|