Introduction
Cervical cancer is the third most common cancer in
women globally, with an estimated global incidence of >500,000
novel cases and a tremendously high number of mortalities
(~274,000), annually (1). Although
radiotherapy, chemotherapy and surgery have been recently used as
standard treatment to improve overall survival, progression free
survival and recurrence rates for patients with cervical cancer
(2,3),
the 5-year survival rates for advance stage cervical cancer (stage
III and IV patients) remains at <40% (4,5). In
addition, ~30% of patients with this disease experience lymph node
recurrence and distant metastasis after primary treatment (6). Therefore, uncovering the molecular
mechanisms responsible for the development of cervical cancer is
crucial for identifying potential therapeutic targets for cervical
cancer.
MicroRNAs (miRNAs/miRs) are non-coding RNA molecules
of 19–25 nucleotides in length that modulate the translational
efficiency or stability of target messenger RNAs (mRNAs),
predominantly through targeting of the 3′ untranslated regions
(3′UTRs) of mRNAs (7). Over the past
decade, emerging evidences have indicated that miRNAs are critical
regulators of cancer occurrence and progression that are involved
in cancer cell proliferation, differentiation, apoptosis and
metastasis (8,9). miRNAs are abnormally expressed in human
cancers and can act as either oncogenes by repressing tumor
suppressors or tumor suppressors by negatively regulating oncogenes
(10). In recent years, miRNAs have
been used as novel targets for anticancer therapies and as
molecular diagnosis marker or prognostic markers in various
cancers, including cervical cancer (11,12).
There is particularly a growing interest toward
miR-138, in the context of numerous cancers. miR-138, a family of
microRNA precursors, has been reported to function as a tumor
suppressor in a variety of human cancers, including renal carcinoma
(13), non-small lung cancer
(14), colorectal cancer (15), neuroblastoma (16), esophageal squamous cell carcinoma
(17), nasopharyngeal carcinoma
(18) and hepatocellular carcinoma
(19). However, the role and the
molecular mechanisms of miR-138 in cervical remain unclear.
Therefore, the aim of the present study was to
investigate the clinical significance of miR-138 expression in
cervical cancer, and to evaluate its role and underlying mechanisms
in cervical cancer. The results of the study showed that miR-138
expression was downregulated in cervical cancer tissues and cell
lines. Overexpression of miR-138 in cervical cancer cells inhibited
cell growth in vitro and tumor growth in nude mice.
Furthermore, the present study showed that human telomerase reverse
transcriptase (hTERT) was a direct target of miR-138 in cervical
cancer. These findings provide a novel therapeutic strategy for the
treatment of cervical cancer.
Materials and methods
Ethics statement
Written informed consent was obtained from patients
according to a protocol approved by the Ethics Committee of the
First of Hospital of Jilin University (Changchun, China). Animal
experiments were performed in strict accordance with the protocols
approved by the Animal Care Committee of the Jilin University.
Patients and tissue samples
In total, 36 pairs of cervical cancer samples and
matched normal cervical tissues were collected from 36 patients,
who underwent surgery between October 2010 and July 2014 at the
First Hospital of Jilin University (Changchun, China). The
specimens were collected immediately after surgery and stored in
liquid nitrogen prior to further use. The category of cervical
samples was confirmed by pathological analysis, as previously
described (20). The patient clinical
information was listed in Table
I.
 | Table I.Association between miR-138 expression
and the clinicopathological features of human cervical cancer. |
Table I.
Association between miR-138 expression
and the clinicopathological features of human cervical cancer.
| Feature | No. of patients | Relative miR-138
level | P-value |
|---|
| Age, years |
|
|
0.743 |
|
<55 | 16 | 0.42±0.11 |
|
|
≥55 | 20 | 0.42±0.09 |
|
| Tumor size |
|
|
0.718 |
| <5
cm | 19 | 0.41±0.10 |
|
| ≥5
cm | 17 | 0.43±0.08 |
|
| Histological
grades |
|
|
0.152 |
|
Well/moderate | 24 | 0.44±0.12 |
|
|
Poor | 12 | 0.38±0.09 |
|
| FIGO stage |
|
| <0.010 |
|
Ib-IIa | 22 | 0.53±0.11 |
|
|
IIb-IIIa | 14 | 0.25±0.06 |
|
| Lymph node
metastasis |
|
| <0.010 |
| No | 28 | 0.48±0.08 |
|
|
Yes | 8 | 0.21±0.06 |
|
Cell lines and transfections
HaCaT cells (an immortalized HPV-negative skin
keratinocyteline) and two cervical cancer cell lines, HeLa and SiHa
cells, were obtained from the Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China), and were maintained
in Dulbecco's modified Eagle's medium (DMEM; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing 10% fetal
bovine serum (FBS; HyClone; GE Healthcare Life Sciences, Logan, UT,
USA). All cells were cultured at 37°C in a humidified atmosphere
containing 5% CO2.
miR-138 mimic and a corresponding negative control
miRNA (miR-NC) were obtained from Shanghai GenePharma Co., Ltd.,
(Shanghai, China), and were transiently transfected into HeLa cells
at a concentration of 50 nM using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions.
Quantitative reverse
transcription-polymerase chain reaction (qRT-PCR)
Total RNA from cells (SiHa, HaCaT and HeLa) or
tissue was isolated using Trizol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Total RNA was reverse-transcribed using the Takara PrimeScript™
First Strand cDNA Synthesis kit (Takara Bio, Inc., Otsu, Japan)
according to the manufacturer's instructions. The levels of miR-138
were determined using a TaqMan MicroRNA Assays kit (TaqMan assay
ID, 002284; measuring mature miR-138; Applied Biosystems; Thermo
Fisher Scientific Inc.) as previously described (21), on an ABI Applied Biosystems 7900 Real
Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The U6 small RNA was used as an internal control. The
primers used were as follows: miR-138 forward,
5′-AGCTGGTGTTGTGAATCAGGCCG-3′ and reverse, 5′-TGGTGTCGTGGAGTCG-3′;
and U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. The PCR cycling conditions were 95°C
for 10 min, and 40 cycles of 15 sec at 95°C and 30 sec at 60°C,
followed by an annealing/elongation step at 72°C for 5 min. The
comparative 2−∆∆Cq method (22) was used for relative quantification and
statistical analysis.
Assessment of cell viability and
proliferative capacity
To determine the effect of miR-138 on cell viability
and proliferative capacity in cervical cancer cells,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and colony formation assays were performed in HeLa cells following
transfection with miR-138 mimic or miR-NC. For cell viability,
5×103 cells were seeded into 96-well plates with 100 µl
of DMEM and incubated for 24 h. Cells were then transfected with
miR-138 or miR-NC, and cultured for 24, 48 or 72 h. Subsequently,
20 µl MTT (5 mg/ml; Sigma-Aldrich, St. Louis, MO, USA) was added to
the samples, and cultured for 4 h at 37°C. Following incubation,
200 µl of dimethyl sulfoxide (DMSO; Sigma-Aldrich) was added to
solubilize the crystals for 20 min at room temperature. Cell
viability was assessed at an absorbance of 490 nm using a biorad
iMark Microplate Absorbance Reader (BD Biosciences, Franklin Lakes,
NJ, USA).
For the colony formation assay, the number of viable
HeLa cell colonies was determined after 14 days, following the
inoculation of 1,000 cells/well in triplicate in 6-well plates. The
cells were fixed with 4% paraformaldehyde (Sigma-Aldrich), and
stained with 1% crystal violet (Sigma-Aldrich). The percentage
colony formation was calculated by adjusting control cells to
100%.
Cell apoptosis assay
The percentage of apoptotic cells was assessed by
flow cytometry. In brief, HeLa cells were transfected with miR-138
mimic or miR-NC for 48 h, and then apoptotic cells were determined
using the AnnexinV/propidium iodide detection kit (Nanjing KeyGen
Biotech. Co. Ltd., Nanjing, China), according to the manufacturer's
instructions. The apoptotic rate were measured by using a FACS
Calibur flow cytometer (BD Biosciences), and the data were analyzed
using CellQuest software (version 5.1; BD Biosciences).
Cell migration and invasion
assays
For the Transwell migration assay, 1×105
transfected HeLa cells in 200 µl of serum-free DMEM were seeded
into the upper part of each Transwell chamber (pore size, 8-µm;
Costar; Corning Incorporated, Corning, NY, USA) without Matrigel.
For the invasion assay, 1×105 transfected cells were
placed on the upper chamber of each insert coated with Matrigel (BD
Biosciences). Subsequently, 500 µl DMEM with 20% FBS was added to
the lower part of the chamber. After incubating for 24 h at 37°C
with 5% CO2, cells that had migrated or invaded to the
lower surface of the filter were fixed in 70% ethanol
(Sigma-Aldrich) for 30 min and stained with 2% crystal violet for
10 min on a glass slide. The invaded or migrated cells were
photographed under an IX51 inverted microscope (Olympus
Corporation, Tokyo, Japan) and counted in 5 randomly selected
fields.
Plasmid construction and luciferase
reporter assay
The 3′UTR of hTERT containing the putative miR-138
binding site (wild-type) was cloned into the pGL3-control vector
(Ambion; Thermo Fisher Scientific, Inc.). A mutant 3′UTR of
phosphatase and tensin homolog (mutant type) was synthesized by PCR
and cloned into the pGL3-control vector. Subsequently,
5×103 HeLa cells were seeded in a 24-well plate and
transiently co-transfected with wild-hTERT-UTR-pGL3 or
mutant-hTERT-UTR-pGL3, a Renilla luciferase control vector
(20 ng) and miR-138 mimic or miR-NC. Luciferase activity was
measured using the Dual Luciferase Reporter Assay System (Promega
Corporation, Madison, WI, USA) at 48 h post-transfection. The
specific activity is expressed as the fold changes of the
experimental group vs. the miR-NC group.
Western blot analysis
Tissue or HeLa cell samples were harvested and lysed
in ice-cold radioimmunoprecipitation assay buffer (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) to extract proteins,
according to the manufacturer's instructions. The concentrations of
the final protein samples were determined with a bicinchoninic acid
protein assay kit (Pierce Biotechnology, Inc., Rockford, IL, USA)
Protein sample (30 µg each lane) was separated on 10% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis and
electroblotted to a polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). Membranes were blocked with 5%
non-fat dried milk in 1X Tris-buffered saline containing 0.1% Tween
20 (TBST; Sigma-Aldrich) buffer for 2 h and then incubated with
primary antibodies separately overnight at 4°C. The primary
antibodies included mouse monoclonal anti-human β-actin (dilution,
1:5,000; catalog no., sc-47778; Santa Cruz Biotechnology, Inc.) and
rabbit polyclonal anti-human hTERT (dilution, 1:1,000; catalog no.,
sc-7212; Santa Cruz Biotechnology, Inc.). β-actin was used as the
internal control. Membranes were washed with TBST three times and
then further incubated with horseradish peroxidase (HRP)-conjugated
polyclonal goat anti-rabbit immunoglobulin G (dilution, 1:5,000;
catalog no., sc-2004; Santa Cruz Biotechnology, Inc.) or
HRP-conjugated polyclonal goat-anti-mouse IgG (1:5,000; catalog no.
sc-2005; Santa Cruz Biotechnology) for 2 h at room temperature.
Signal intensities were detected using the Odyssey Infrared Imaging
System (Li-COR Biosciences, Lincoln, NE, USA) using Odyssey v1.2
software (Li-COR Biosciences).
Xenograft experiments
Five week-old female BALB/c nu/nu mice (18–20 g)
were purchased from Jilin Institute of Experimental Animals
(Changchun, China), and bred at the Animal Laboratory Center, Jilin
University (Changchun, China).
Equal numbers of HeLa cells (2×106) with
forced expression of miR-138 mimic or scramble were suspended in
100 µl serum-free DMEM and injected subcutaneously into the right
rear flank of each mouse (n=10). Tumor volume was periodically
blindly measured by caliper every 5 days until mice were sacrificed
under anesthesia. The formula for tumor volume was as follows: V
(volume) = 1/2 × A × B2, where A and B are the longest
and shortest diameters, respectively. The mice were sacrificed 25
days subsequent to injection. The tumor tissues were dissected and
weighed. A part of tumor tissues was used to measure the hTERT
level by western blot analysis, using the aforementioned
method.
Statistical analysis
Experimental data are presented as the mean ±
standard deviation based on the results of at least 3 repeats.
Statistical analysis between two samples was performed using a
two-tailed Student's t-test, and analysis between more than
two groups was performed using one-way analysis of variance,
followed by Tukey's post hoc test. GraphPad Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA) was used for
statistical analyses. A value of P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-138 is downregulated in human
cervical cancer cell lines and tissue specimens
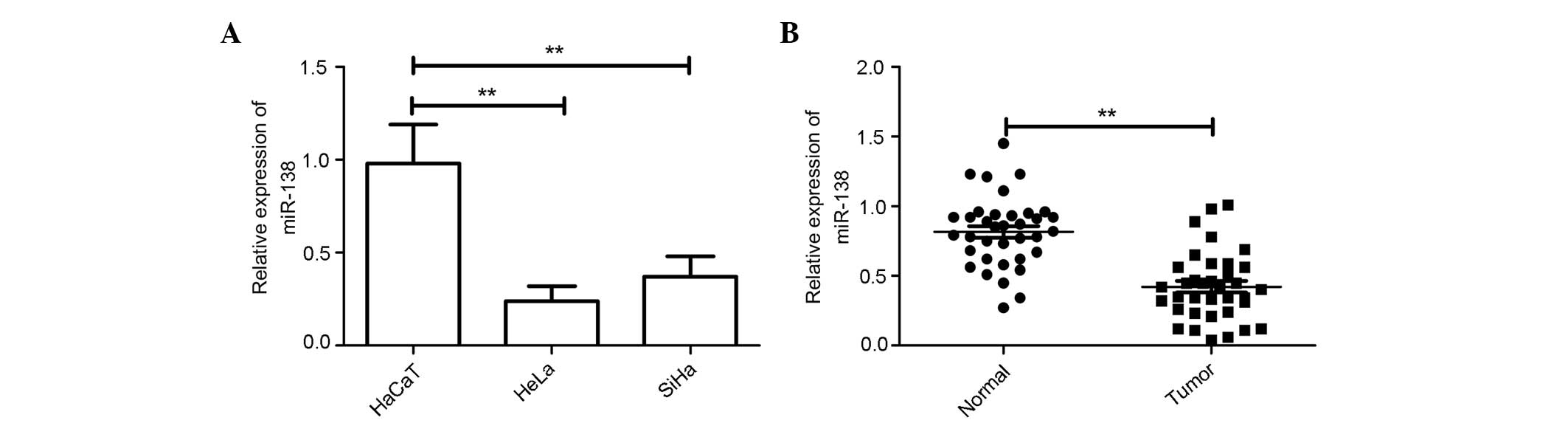
A panel of human cervical cancer cell lines was
first analyzed to quantitate the expression level of miR-138. The
result showed that the expression level of miR-138 was
downregulated in cervical cell lines compared with the normal
cervical HaCaT cells (Fig. 1A).
Additionally, the expression level of miR-138 in the HeLa cell line
was decreased compared with the SiHa cell line; thus, HeLa cells
were selected for the rest of the study.
The expression levels of miR-138 were examined in
the 36 cervical cancer specimens and the patient-matched normal
specimens. RT-PCR analysis showed that the expression of miR-138
was downregulated in 83.3% of the cervical tissues compared with
the adjacent normal tissues; the average magnitude of this decrease
was 2.1-fold (Fig. 1B).
In addition, the association between miR-138
expression and the clinical factors was assessed. As shown in
Table I, the aberrant expression of
miR-138 was associated with lymph node metastasis (P<0.01) and
International Federation of Gynecology and Obstetrics (FIGO) stage
(P<0.01). There was no association between miR-138 expression
and age, histological grade or tumor size. These findings suggest
that miR-138 could be involved in the metastasis and progression of
cervical cancer.
Overexpression of miR-138 inhibits
cell viability and colony formation, but induces apoptosis in
cervical cancer cells
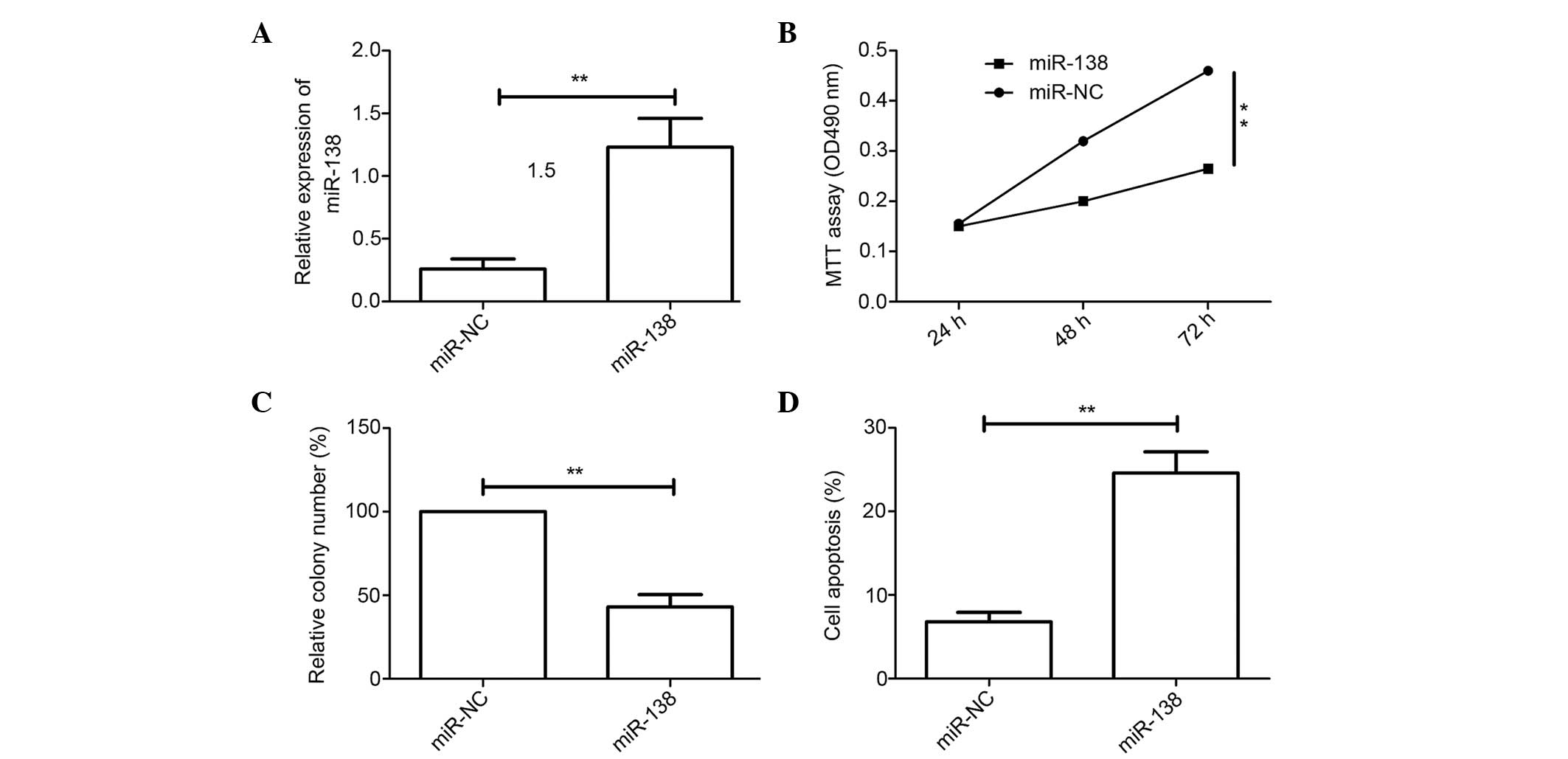
To investigate the biological effects of miR-138 in
cervical cancer cells, the cervical cancer HeLa cell line was
transfected with miR-138 mimic and then the function of miR-138 in
the cervical cancer cells was evaluated. qRT-PCR analysis confirmed
that the transfected miR-138 mimic resulted in the upregulation of
miR-138 expression in cervical cancer cells compared with cells
transfected with miR-NC (Fig. 2A).
The MTT assay showed that transfection with miR-138 mimic inhibited
cell proliferation compared with transfection with miR-NC (Fig. 2B). The colony formation assay
demonstrated that the overexpression of miR-138 inhibited colony
formation in cervical cancer cells (Fig.
2C). Flow cytometry as then used to test the role of miR-138 in
apoptosis. The results showed that upregulated miR-138 induced
apoptosis (Fig. 2D). These results
suggest that miR-138 may function as a tumor suppressor in cervical
cancer cells.
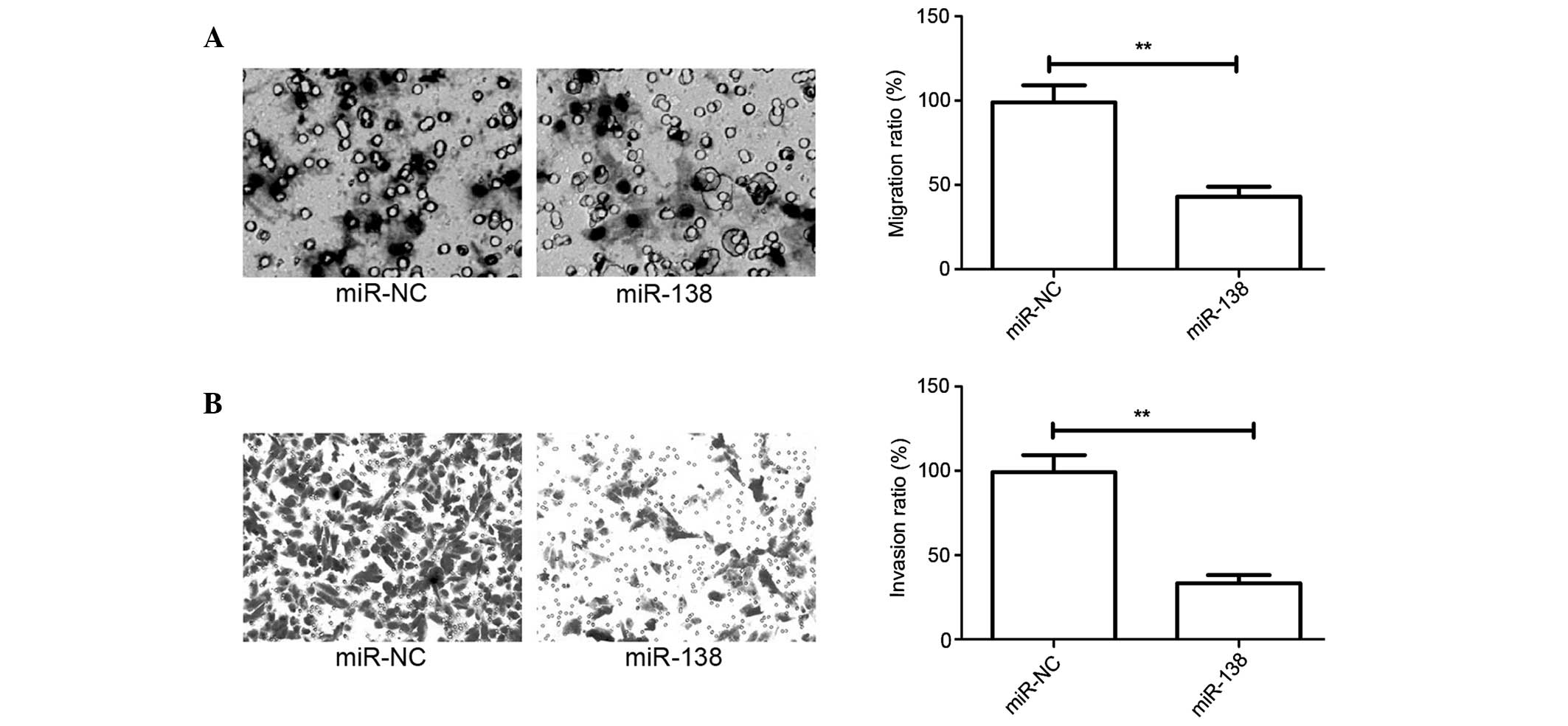
Overexpression of miR-138 inhibits
cell migration and invasion in cervical cancer cells
The present study has showed that miR-138 expression
is associated with lymph node metastasis in patients with cervical
cancer. However, this finding does not determine whether miR-138
effects the migration and invasion of cervical cancer cells.
Migration and invasion assays in cervical cancer cells were
performed using Transwell chambers, after HeLa cells were
transfected with miR-138 mimic or miR-NC. Consistent with the
clinical data, the overexpression of miR-138 significantly
decreased the migration and invasion capacities of HeLa cells
(P<0.05; Fig. 3A and B).
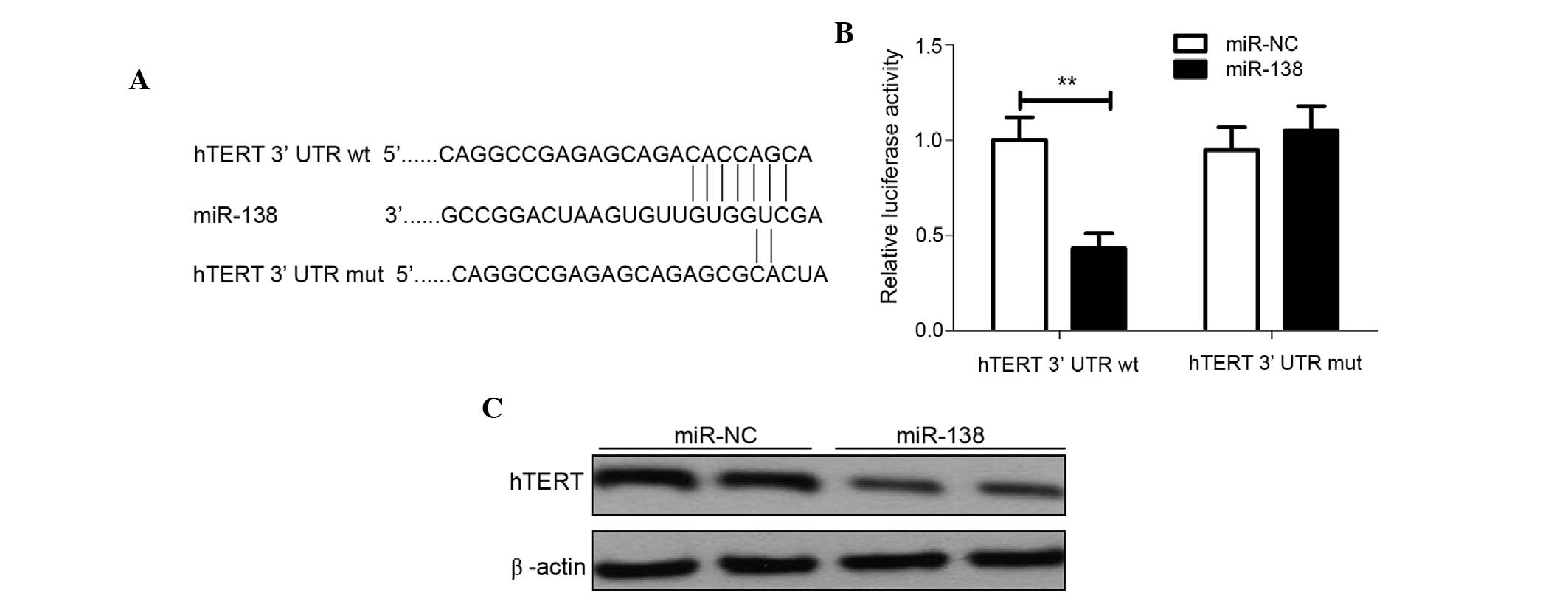
hTERT is a direct target of
miR-138
hTERT has been reported to be an important molecule
for promoting cell proliferation and invasion in various cancers,
including cervical cancer (23).
Using predictive tools (TargetScanHuman 7.1; www.targetscan.org/vert_71/), hTERT was hypothesized
to be a target of miR-138 (Fig. 4A).
To additionally confirm whether miR-138 directly targets the hTERT
oncogene, luciferase reporter assays were performed. The luciferase
assay showed that HeLa cells transfected with miR-138 significantly
decreased wild-type hTERT-3′UTR reporter activity compared with the
cells co-transfected with miR-NC (P<0.01), while miR-138 had no
inhibitory effect on mutant hTERT-3′UTR reporter activity (Fig. 4B), indicting the direct regulation of
miR-138 in the 3′UTR of hTERT mRNA. To further validate the
association between miR-138 and hTERT, endogenous hTERT protein
expression was detected in the HeLa cells transfected with miR-138
mimic or miR-NC. Western blot analysis revealed that miR-138 mimics
dramatically decreased the expression of endogenous hTERT
expression (Fig. 4C). Overall, these
results suggest that hTERT is a direct target gene of miR-138.
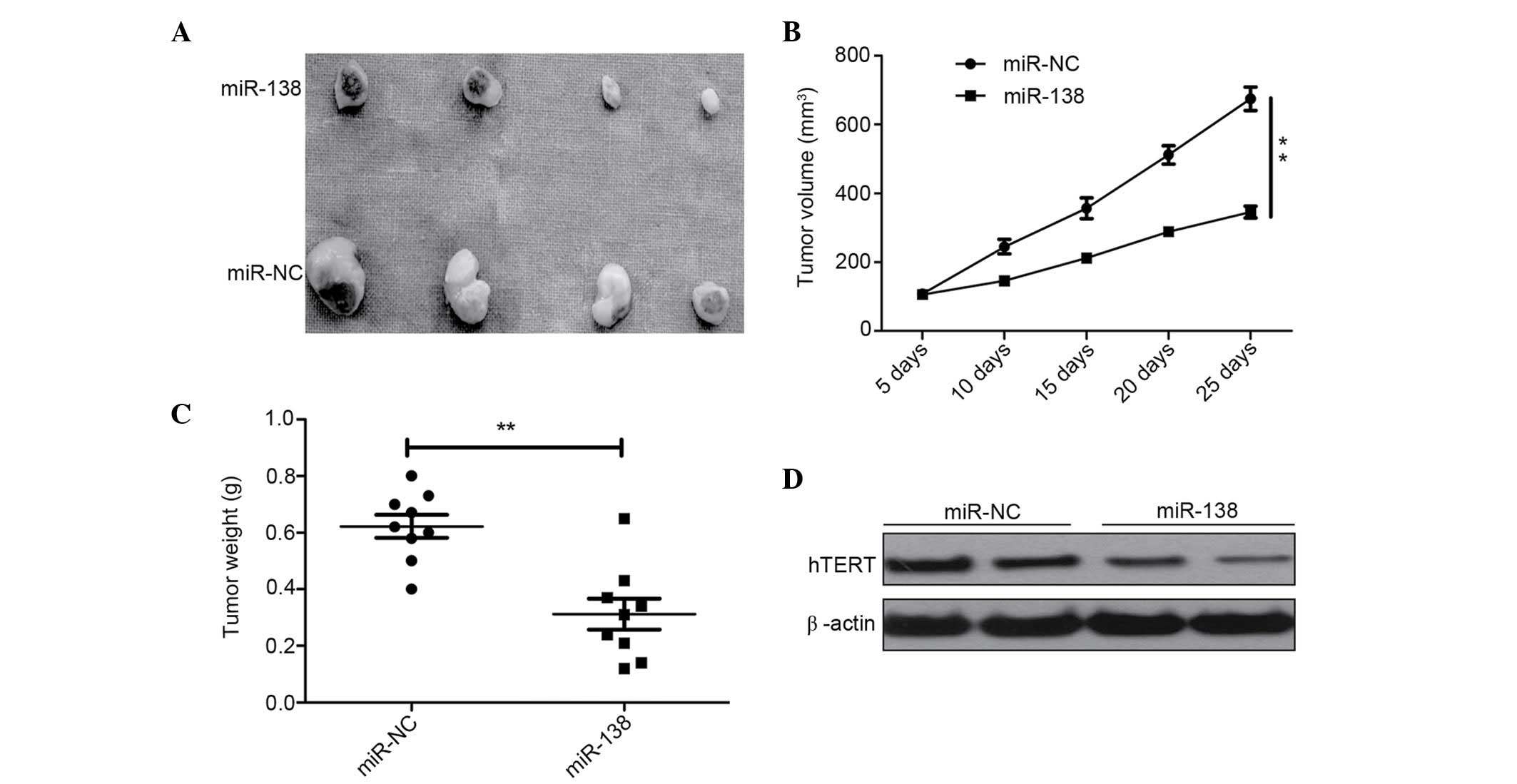
miR-138 suppresses tumor growth in
vivo
To determine whether miR-138 is involved in
tumorigenesis in vivo, the present study examined the
potential activity of miR-138 in tumorigenesis using a HeLa
xenograft model. Tumors grew slower in the HeLa/miR-138 group
compared with the HeLa/miR-NC group (Fig.
5A). At the end of the treatment, a significant decrease in
tumor size (Fig. 5B) and weight
(Fig. 5C) was observed in mice from
the group injected with HeLa/miR-138 compared with the mice
injected with HeLa/miR-NC (Fig. 5B and
C). Furthermore, hTERT expression was also determined in tumor
tissue using western blot analysis. Western blot analysis
demonstrated that the overexpression of miR-138 significantly
decreased hTERT expression. These findings suggest that miR-138
could suppress the growth of cervical cancer tumors in vivo
by targeting hTERT.
Discussion
Discovering the molecules involved in cervical
cancer cell initiation and progression and understanding the
associated mechanisms are critical for developing effective
therapeutic strategies to improve the survival and prognosis of
patients suffering from cervical cancer. Growing evidence has
demonstrated that numbers of miRNAs are crucial for the initiation,
progression and metastasis of cervical cancer by regulating various
processes, including cancer cell proliferation, differentiation,
apoptosis, adhesion, cell cycle arrest, migration and invasion
(11,24,25). For
instance, Zhou et al (26)
demonstrated that that miR-107 directly targeted myeloid cell
leukemia-1 and activated the ATR serine/threonine kinase/checkpoint
kinase 1 pathway to inhibit the proliferation, migration and
invasiveness of cervical cancer cells. Li et al (27) reported that the overexpression of
miR-342-3p inhibits cell proliferation, migration and invasion in
cervical cell lines by targeting mammalian transcription factor
forkhead box M1. Wen et al (28) reported that miR-506 induced cell cycle
arrest at the G1/S transition, enhanced the apoptosis and
chemosensitivity of cervical cancer cells, and inhibited cervical
cancer growth in vitro and in vivo. Data from the
current study provides evidence that miR-138 can inhibit
proliferation, colony formation, migration and invasion, induce
cell apoptosis in cervical cancer cells, and suppress tumor growth
in nude mice models.
miRNAs can act as either tumor suppressors or
promoters and therefore affect tumor development, proliferation,
differentiation, migration and invasion (10). The expression of miR-138 is generally
low in tumors, including in non-small lung cancer, colorectal
cancer, neuroblastoma, esophageal squamous cell carcinoma,
nasopharyngeal carcinoma and hepatocellular carcinoma (14–19).
However, an association between miR-138 expression and cervical
cancer has not been previously reported. To the best of our
knowledge, the present study is the first to demonstrate that
miR-138 expression is significantly downregulated in cervical
cancer tissues and cell lines, and that low miR-138 expression is
negatively associated with advanced FIGO stage and lymph node
metastasis. In addition, miR-138 has been previously reported to
function as a tumor suppressor in numerous malignancies by
targeting various molecules (13–19). For
cervical cancer, a study showed that miR-138 could significantly
inhibit HeLa cell migration by targeting required for meiotic
nuclear division 5 homolog A (29).
However, the effect of miR-138 in cervical development and
progression remains largely unknown. The present study elucidates
the functionality and mechanism of the involvement of miR-138 in
cervical cancer processes, and shows that miR-138 could act as
tumor suppressor in cervical cancer and inhibit cervical tumor
growth in vitro and in vivo by targeting hTERT.
hTERT, a catalytic subunit of telomerase, is a core
component of the telomerase holoenzyme and is involved in
regulating telomerase activity (23,30). hTERT
has been reported to be important for cancer tumorigenesis, growth,
migration and invasion (31,32). Previously, studies showed that hTERT
is a target gene of miR-138 in thyroid carcinoma cell lines
(33) and colorectal cancer (34). Consistent with these results, the
present study identified hTERT as potential target of miR-138 in
cervical cancer cells.
In conclusion, the present study has demonstrated
that miR-138 expression is significantly downregulated in cervical
cancer tissues and cell lines, and that decreased miR-138
expression is negatively associated with advanced FIGO stage and
lymph node metastasis. miR-138 acts as a tumor suppressor in
cervical cancer by suppressing cancer growth, inhibiting cell
migration and invasion, and enhancing apoptosis. Furthermore,
miR-138 was found to exert its function by directly targeting
hTERT. These findings indicate that miR-138 may act as a novel
potential therapeutic agent for the treatment of cervical
cancer.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Duenas-Gonzalez A, Serrano-Olvera A,
Cetina L and Coronel J: New molecular targets against cervical
cancer. Int J Womens Health. 6:1023–1031. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
de Freitas AC, Leitão Mda C Gomes and
Coimbra EC: Prospects of molecularly-targeted therapies for
cervical cancer treatment. Curr Drug Targets. 16:77–91. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Keys HM, Bundy BN, Stehman FB, Muderspach
LI, Chafe WE, Suggs CL III, Walker JL and Gersell D: Cisplatin,
radiation and adjuvant hysterectomy compared with radiation and
adjuvant hysterectomy for bulky stage IB cervical carcinoma. N Engl
J Med. 340:1154–1161. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Smith RA, Brooks D, Cokkinides V, Saslow D
and Brawley OW: Cancer screening in the United States, 2013: A
review of current American Cancer Society guidelines, current
issues in cancer screening, and new guidance on cervical cancer
screening and lung cancer screening. CA Cancer J Clin. 63:88–105.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Waggoner SE: Cervical cancer. Lancet.
361:2217–2225. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cummins JM and Velculescu VE: Implications
of micro-RNA profiling for cancer diagnosis. Oncogene.
25:6220–6227. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Miska EA: How microRNAs control cell
division, differentiation and death. Curr Opin Genet Dev.
15:563–568. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He L, Thomson JM, Hemann MT,
Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe
SW, Hannon GJ and Hammond SM: A microRNA polycistron as a potential
human oncogene. Nature. 435:828–833. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Munker R and Calin GA: MicroRNA profiling
in cancer. Clin Sci (Lond). 121:141–158. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li C, Feng Y, Coukos G and Zhang L:
Therapeutic microRNA strategies in human cancer. AAPS J.
11:747–757. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liang J, Zhang Y, Jiang G, Liu Z, Xiang W,
Chen X, Chen Z and Zhao J: MiR-138 induces renal carcinoma cell
senescence by targeting EZH2 and is downregulated in human clear
cell renal cell carcinoma. Oncol Res. 21:83–91. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ye XW, Yu H, Jin YK, Jing XT, Xu M, Wan ZF
and Zhang XY: miR-138 inhibits proliferation by targeting
3-phosphoinositide-dependent protein kinase-1 in non-small cell
lung cancer cells. Clin Respir J. 9:27–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Long L, Huang G, Zhu H, Guo Y, Liu Y and
Huo J: Down-regulation of miR-138 promotes colorectal cancer
metastasis via directly targeting TWIST2. J Transl Med. 11:2752013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chakrabarti M, Banik NL and Ray SK:
miR-138 overexpression is more powerful than hTERT knockdown to
potentiate apigenin for apoptosis in neuroblastoma in vitro and in
vivo. Exp Cell Res. 319:1575–1585. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gong H, Song L, Lin C, Liu A, Lin X, Wu J,
Li M and Li J: Downregulation of miR-138 sustains NF-κB activation
and promotes lipid raft formation in esophageal squamous cell
carcinoma. Clin Cancer Res. 19:1083–1093. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu X, Lv XB, Wang XP, Sang Y, Xu S, Hu K,
Wu M, Liang Y, Liu P, Tang J, et al: MiR-138 suppressed
nasopharyngeal carcinoma growth and tumorigenesis by targeting the
CCND1 oncogene. Cell Cycle. 11:2495–2506. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang W, Zhao LJ, Tan YX, Ren H and Qi ZT:
MiR-138 induces cell cycle arrest by targeting cyclin D3 in
hepatocellular carcinoma. Carcinogenesis. 33:1113–1120. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Solomon D, Davey D, Kurman R, Moriarty A,
O'Connor D, Prey M, Raab S, Sherman M, Wilbur D, Wright T Jr and
Young N: Forum Group Members; Bethesda 2001 Workshop: The 2001
Bethesda System: Terminology for reporting results of cervical
cytology. JAMA. 287:2114–2119. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang H, Luo J, Liu Z, Zhou R and Luo H:
MicroRNA-138 regulates DNA damage response in small cell lung
cancer cells by directly rargeting H2AX. Cancer Invest. 33:126–136.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi YA, Zhao Q, Zhang LH, Du W, Wang XY,
He X, Wu S and Li YL: Knockdown of hTERT by siRNA inhibits cervical
cancer cell growth in vitro and in vivo. Int J Oncol. 45:1216–1224.
2014.PubMed/NCBI
|
|
24
|
Banno K, Iida M, Yanokura M, Kisu I, Iwata
T, Tominaga E, Tanaka K and Aoki D: MicroRNA in cervical cancer:
OncomiRs and tumor suppressor miRs in diagnosis and treatment.
Scientific World Journal. 2014:1780752014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ribeiro J and Sousa H: MicroRNAs as
biomarkers of cervical cancer development: A literature review on
miR-125b and miR-34a. Mol Biol Rep. 41:1525–1531. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou C, Li G, Zhou J, Han N, Liu Z and Yin
J: miR-107 activates ATR/Chk1 pathway and suppress cervical cancer
invasion by targeting MCL1. PloS One. 9:e1118602014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li XR, Chu HJ, Lv T, Wang L, Kong SF and
Dai SZ: miR-342-3p suppresses proliferation, migration and invasion
by targeting FOXM1 in human cervical cancer. FEBS Lett.
588:3298–3307. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wen SY, Lin Y, Yu YQ, Cao SJ, Zhang R,
Yang XM, Li J, Zhang YL, Wang YH, Ma MZ, et al: miR-506 acts as a
tumor suppressor by directly targeting the hedgehog pathway
transcription factor Gli3 in human cervical cancer. Oncogene.
34:717–725. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li J, Chen Y, Qin X, Wen J, Ding H, Xia W,
Li S, Su X, Wang W, Li H, et al: MiR-138 downregulates miRNA
processing in HeLa cells by targeting RMND5A and decreasing
Exportin-5 stability. Nucleic Acids Res. 42:458–474. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang W and Xing L: RNAi gene therapy of
SiHa cells via targeting human TERT induces growth inhibition and
enhances radiosensitivity. Int J Oncol. 43:1228–1234.
2013.PubMed/NCBI
|
|
31
|
Cifuentes-Rojas C and Shippen DE:
Telomerase regulation. Mutat Res. 730:20–27. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Noël JF and Wellinger RJ: Exposing secrets
of telomere-telomerase encounters. Cell. 150:453–454. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mitomo S, Maesawa C, Ogasawara S, Iwaya T,
Shibazaki M, Yashima-Abo A, Kotani K, Oikawa H, Sakurai E, Izutsu
N, et al: Downregulation of miR-138 is associated with
overexpression of human telomerase reverse transcriptase protein in
human anaplastic thyroid carcinoma cell lines. Cancer Sci.
99:280–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Qin YZ, Xie XC, Liu HZ, Lai H, Qiu H and
Ge LY: Screening and preliminary validation of miRNAs with the
regulation of hTERT in colorectal cancer. Oncol Rep. 33:2728–2736.
2015.PubMed/NCBI
|